





 та дезоксирибонуклеїнові (ДНК) кислоти")



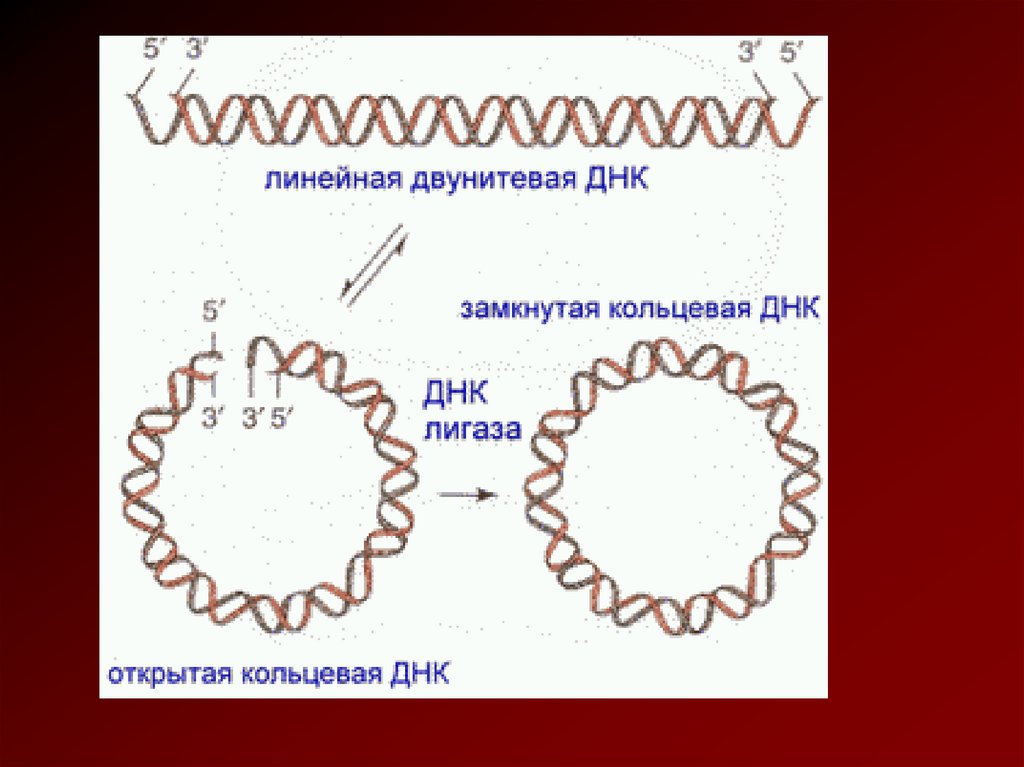
 ядро; 2) цитоплазму; 3) оболонку")
 забезпечує процеси біосинтезу й передачі спадкової інформації та складається із:")





")


")

















 чоловіка")









 biology
biologySimilar presentations:




")
")




Будова ядра
1.
Львівський національний університет імені Івана ФранкаКафедра біофізики та біоінформатики
Будова ядра
Викладач:
доц. Отчич Віра Петрівна
2. План:
1. Будова нуклеїнових кислот.2. Будова клітинного ядра.
3. Будова,
типи
і
властивості
метафазних хромосом.
3. Будова нуклеїнових кислот
У залежності від природи вуглеводу,
що входить до складу нуклеотидів,
нуклеїнові кислоти розподіляють на
два види дезоксирибонуклеїнові
кислоти (ДНК), які містять вуглевод
2-дезокси- D-рибоза
та
2-дезокси-D-рибозу,
рибонуклеїнові кислоти (РНК), які
вміщують вуглевод D-рибозу.
2-дезокси-D-рибоза
та
D-рибоза
знаходяться в нуклеїнових кислотах
у β-фуранозній формі:
D-рибоза
4.
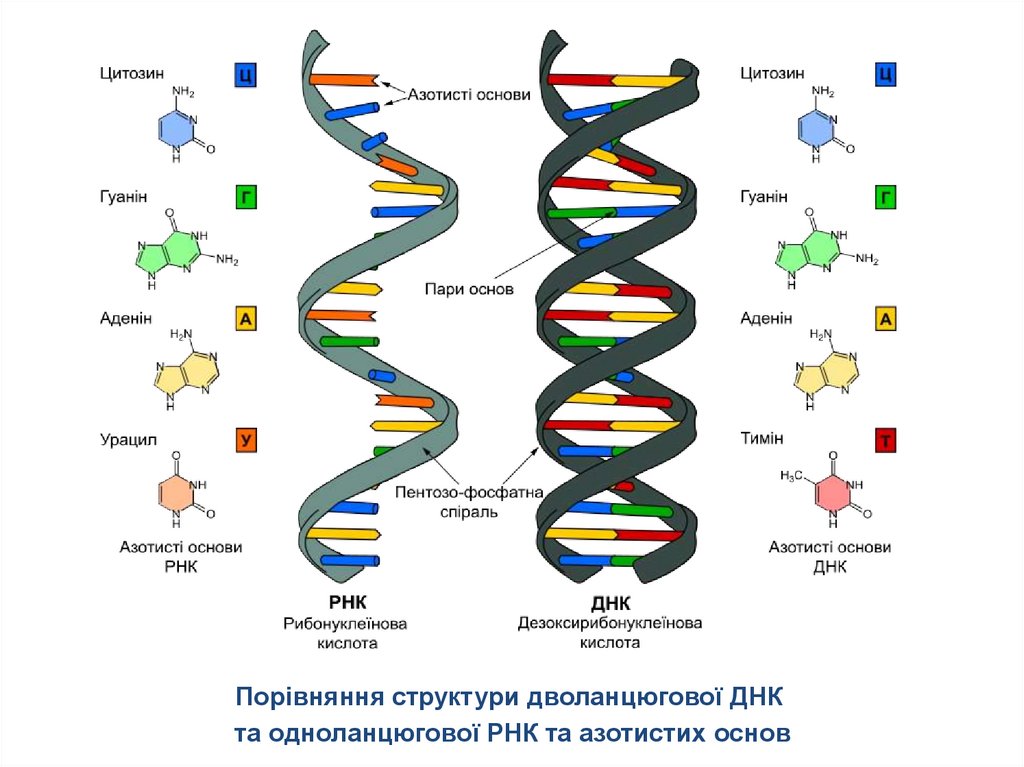
Порівняння структури дволанцюгової ДНКта одноланцюгової РНК та азотистих основ
5. Азотисті основи
цитозинаденін
тимін
урацил
гуанін
6.
NH2N
N
H
N
N
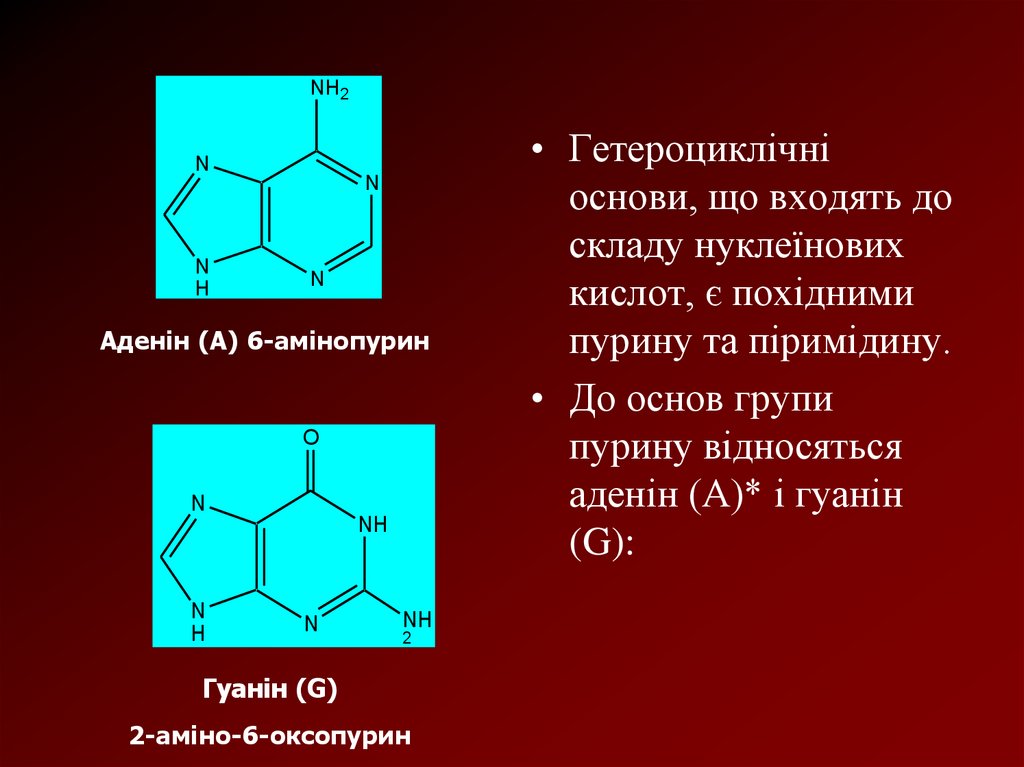
Аденін (А) 6-амінопурин
O
N
NH
N
H
N
NH
2
Гуанін (G)
2-аміно-6-оксопурин
• Гетероциклічні
основи, що входять до
складу нуклеїнових
кислот, є похідними
пурину та піримідину.
• До основ групи
пурину відносяться
аденін (А)* і гуанін
(G):
7.
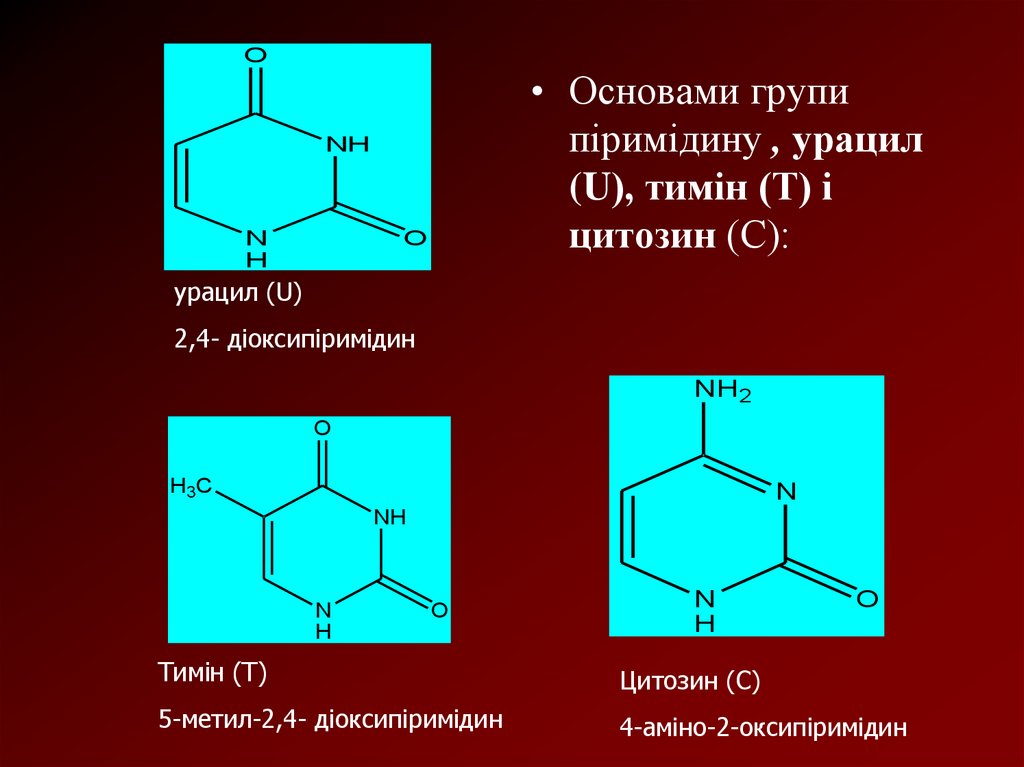
O• Основами групи
піримідину , урацил
(U), тимін (Т) і
цитозин (С):
NH
N
H
O
урацил (U)
2,4- діоксипіримідин
NH2
O
H3 C
N
NH
N
H
O
N
H
O
Тимін (Т)
Цитозин (С)
5-метил-2,4- діоксипіримідин
4-аміно-2-оксипіримідин
8.
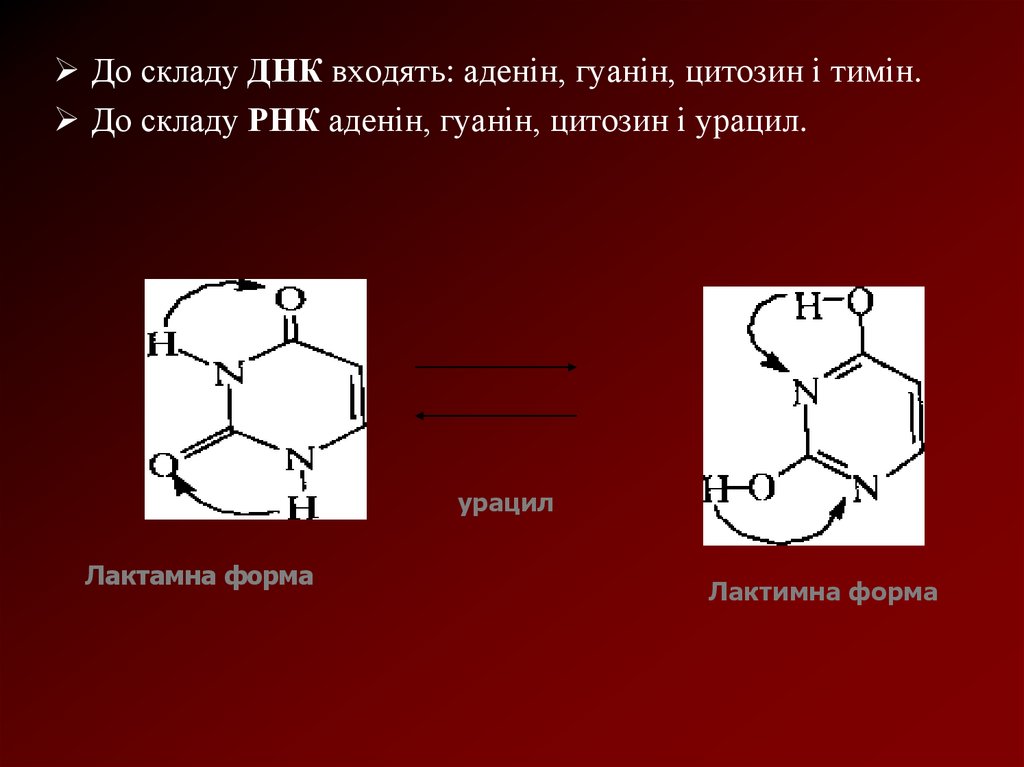
До складу ДНК входять: аденін, гуанін, цитозин і тимін.До складу РНК аденін, гуанін, цитозин і урацил.
урацил
Лактамна форма
Лактимна форма
9.
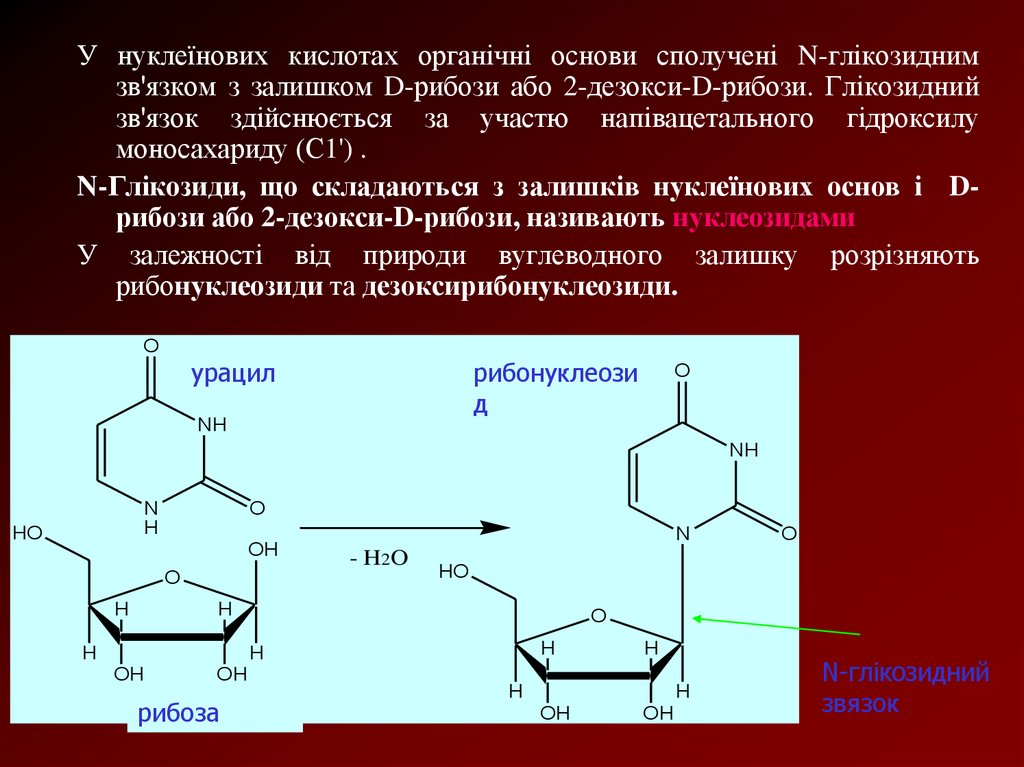
У нуклеїнових кислотах органічні основи сполучені N-глікозиднимзв'язком з залишком D-рибози або 2-дезокси-D-рибози. Глікозидний
зв'язок здійснюється за участю напівацетального гідроксилу
моносахариду (С1') .
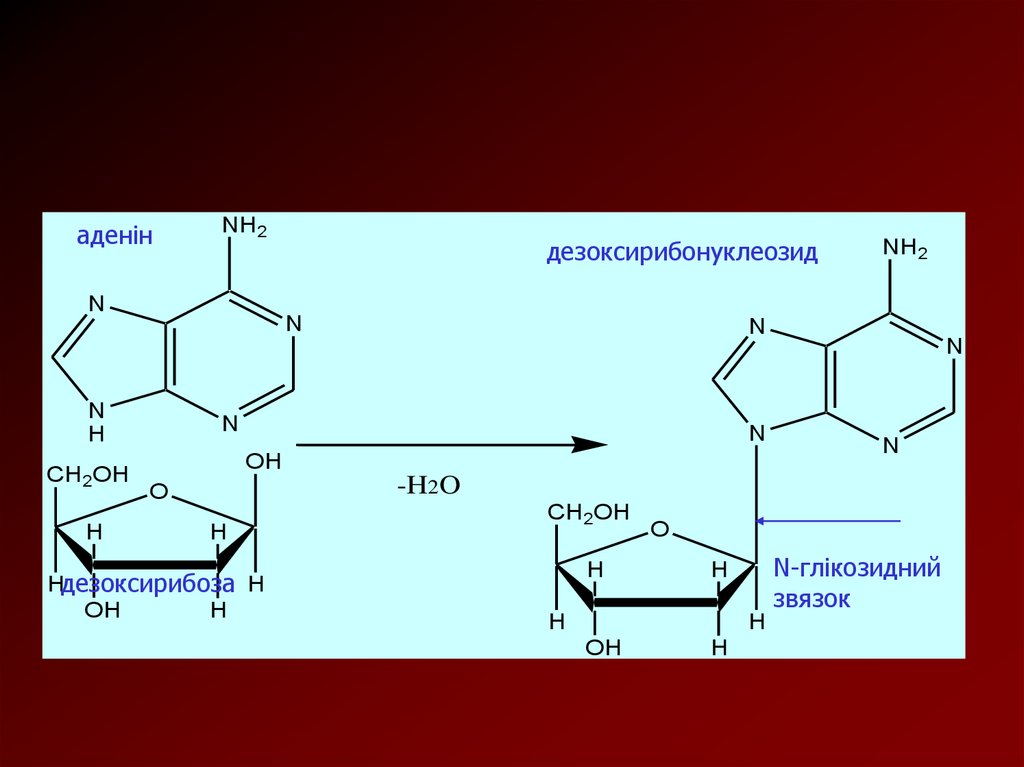
N-Глікозиди, що складаються з залишків нуклеїнових основ і Dрибози або 2-дезокси-D-рибози, називають нуклеозидами
У залежності від природи вуглеводного залишку розрізняють
рибонуклеозиди та дезоксирибонуклеозиди.
O
урацил
рибонуклеози
д
NH
O
NH
N
H
HO
O
OH
O
H
N
- H2O
HO
H
H
O
H
H
OH
H
OH
рибоза
O
H
H
OH
OH
N-глікозидний
звязок
10.
аденінNH2
N
N
N
H
CH2OH
H
дезоксирибонуклеозид
NH2
N
N
N
OH
N
N
-H2O
O
H
Hдезоксирибоза H
OH
H
CH2OH
H
O
N-глікозидний
звязок
H
H
H
OH
H
11.
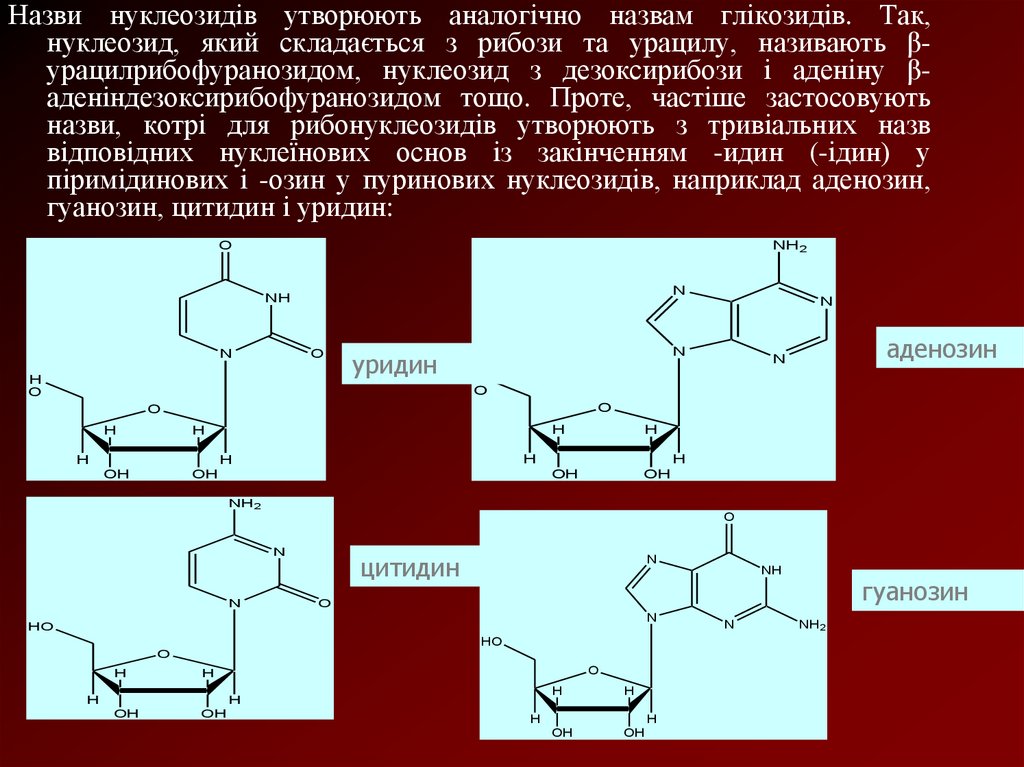
Назви нуклеозидів утворюють аналогічно назвам глікозидів. Так,нуклеозид, який складається з рибози та урацилу, називають βурацилрибофуранозидом, нуклеозид з дезоксирибози і аденіну βаденіндезоксирибофуранозидом тощо. Проте, частіше застосовують
назви, котрі для рибонуклеозидів утворюють з тривіальних назв
відповідних нуклеїнових основ із закінченням -идин (-ідин) у
піримідинових і -озин у пуринових нуклеозидів, наприклад аденозин,
гуанозин, цитидин і уридин:
NH2
O
N
NH
N
O
H
O
уридин
N
N
аденозин
N
H
O
O
O
H
H
OH
OH
H
H
H
OH
OH
H
H
H
NH2
O
N
N
цитидин
N
NH
N
HO
HO
O
H
H
OH
OH
H
гуанозин
O
O
H
H
H
OH
OH
H
H
N
NH2
12.
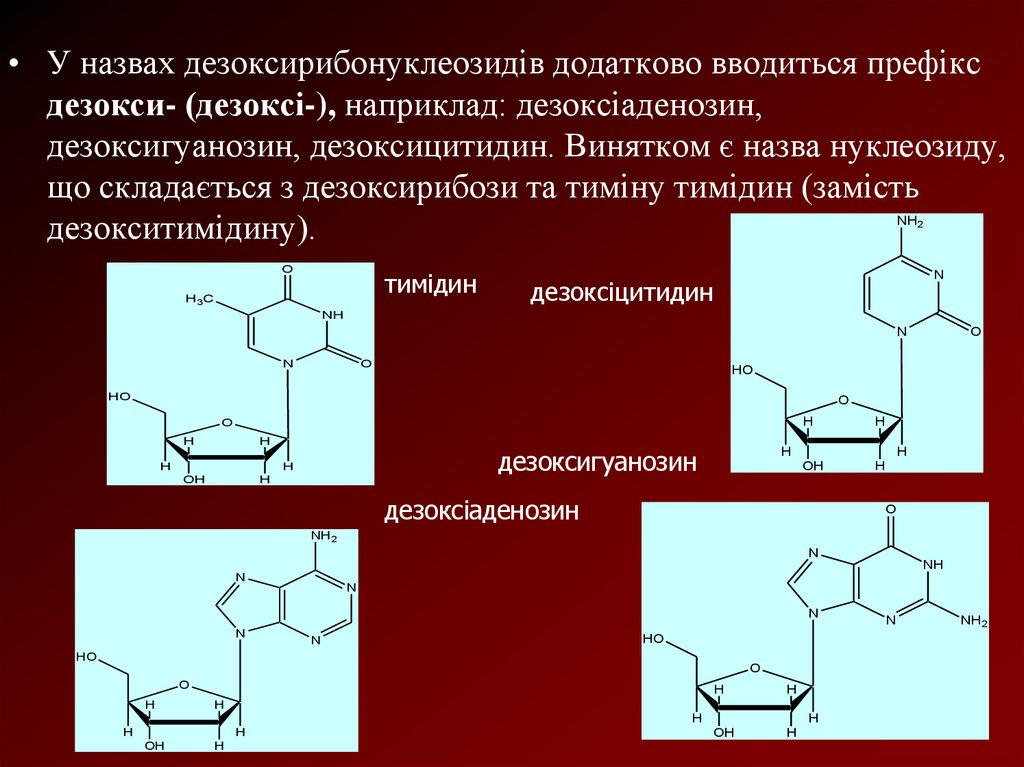
• У назвах дезоксирибонуклеозидів додатково вводиться префіксдезокси- (дезоксі-), наприклад: дезоксіаденозин,
дезоксигуанозин, дезоксицитидин. Винятком є назва нуклеозиду,
що складається з дезоксирибози та тиміну тимідин (замість
дезокситимідину).
NH2
O
тимідин
H3C
N
дезоксіцитидин
NH
N
N
O
O
HO
HO
O
O
H
H
H
дезоксигуанозин
H
OH
H
H
H
OH
H
H
H
дезоксіаденозин
O
NH2
N
NH
N
N
N
N
N
HO
HO
O
O
H
H
H
OH
H
H
H
H
H
OH
H
H
N
NH2
13.
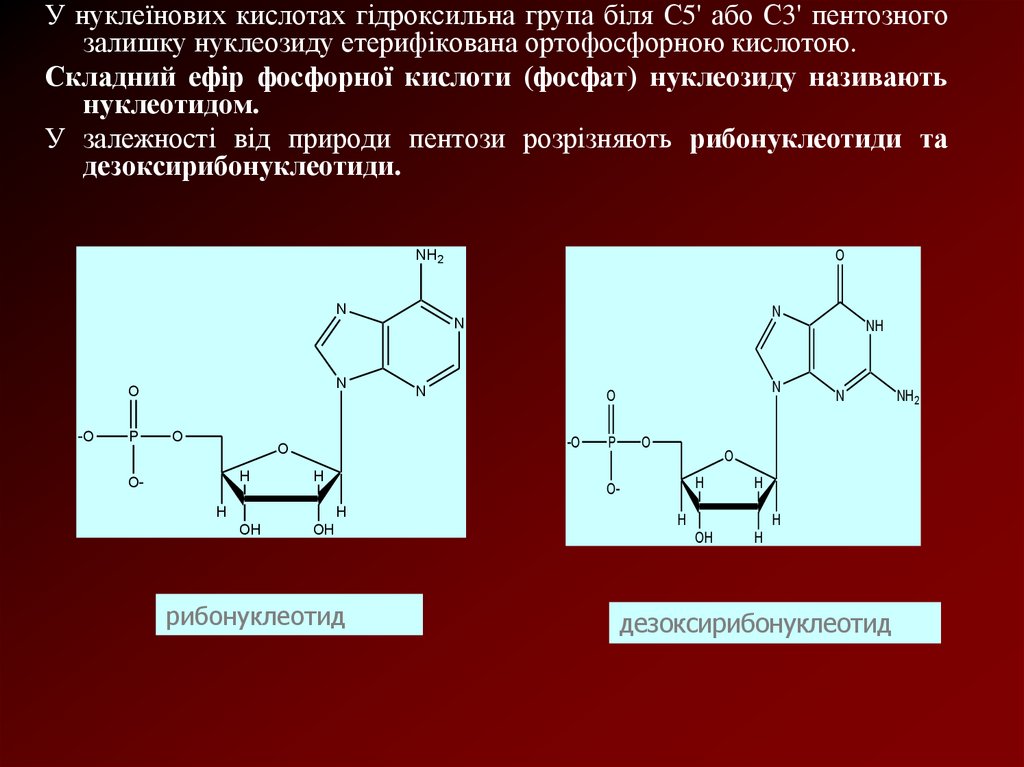
У нуклеїнових кислотах гідроксильна група біля С5' або С3' пентозногозалишку нуклеозиду етерифікована ортофосфорною кислотою.
Складний ефір фосфорної кислоти (фосфат) нуклеозиду називають
нуклеотидом.
У залежності від природи пентози розрізняють рибонуклеотиди та
дезоксирибонуклеотиди.
NH2
N
N
O
-O
P
O
N
N
N
H
H
OH
OH
H
N
O
-O
O
O-
O
P
O
рибонуклеотид
N
O
OH
NH
H
H
OH
H
H
H
дезоксирибонуклеотид
NH2
14. Номенклатура нуклеотидів
Назви нуклеотидів як монофосфатівНазви нуклеотидів як
кислот
Аденозин-5'-монофосфат (АМФ)
5'-аденілова кислота
Гуанозин-5'-монофосфат (ГМФ)
5'-гуанілова кислота
Цитидин-5'-монофосфат (ЦМФ)
5'-цитидилова кислота
Уридин-5'-монофосфат (УМФ)
5'-уридилова кислота
Дезоксіаденозин-5'-монофосфат
(ДАМФ)
дезоксіадепілова кислота
Дезоксигуанозин-5'моІюфосфат(ДГМФ)
дезоксигуанілова кислота
Дезоксицитидип-5'-моІюфосфат
(ДЦМФ)
дезоксицитидилова кислота
Тимідин-5'-монофосфат (ДТМФ)
тимідилова кислота
15.
• Нуклеїнові кислоти являють собою продукти полімеризаціїмоно-нуклеотидів. Нуклеотиди сполучаються в довгі
ланцюги за допомогою фосфодіефірних зв'язків, які
утворюються за участю гідроксилу при С*' попередньої
нуклеотидної ланки та гідроксилу, що належить С ' дальшої
нуклеотидної ланки.
• Мононуклеотиди, їх похідні та динуклеотиди присутні в
клітинах також у вільному вигляді та виконують важливу
роль в обміні речовин. В усіх тканинах організму, поряд з
нуклеозидмонофосфатами, містяться ди- та трифосфати
нуклеозидів.
• Особливо широко відомі аденозин-5’- фосфат (АМФ),
аденозин-5’-дифосфат (АДФ) і аденозин-5'-трифосфат
(АТФ).
• Ці нуклеотиди здатні до взаємоперетворення шляхом
фосфорилювання (приєднання одного або двох залишків
фосфорної кислоти до АМФ) або ж дефосфорилювання
(відщеплення одного або двох залишків фосфорної кислоти
від АТФ). При дефосфорилюванні виділяється значна
кількість енергії, котра використовується в організмі для
проходження тих чи інших біологічних процесів, наприклад,
у біосинтезі білка.
16. Рибонуклеїнові (РНК) та дезоксирибонуклеїнові (ДНК) кислоти
Схема ділянки полінуклеотидного ланцюга молекули нуклеїнової кислоти• Нуклеїнові кислоти являють собою високомолекулярні
гетерополімери, які складаються з залишків ортофосфорної
кислоти та рибози або дезоксирибози, що чергуються, сполучених
з нуклеїновими основами, котрі виступають у полімерному
ланцюзі як «бокові групи»
17.
Певнапослідовність
нуклеотидних
ланок
у
полінуклеотидному ланцюзі називається первинною
структурою нуклеїнових кислот.
(Просторова орієнтація полінуклеотидних ланцюгів у
молекулі
називається
вторинною
структурою
нуклеїнових кислот.
Вперше вторинну структуру ДНК у вигляді моделі з
подвійної спіралі описали американський біохімік Дж.
Уотсон і англійський біохімік Ф. Крик (1953 p.).
Узагальнивши роботи Л. Полінга, А. Тод-да, Е. Чаргаффа,
М. Уілкінса та інших, вони дійшли висновку, що молекула
ДНК являє собою дві паралельні правозакручені спіралі
(подвійна спіраль), фіксовані між собою ван-дерваальсовими силами притягання, що діють вздовж спіралі
між ядрами нуклеїнових основ (міжплощинна вертикальна
взаємодія). Крім того, вторинна структура стабілізується
водневими зв'язками між залишками нуклеїнових основ
двох паралельних спіралей.
18.
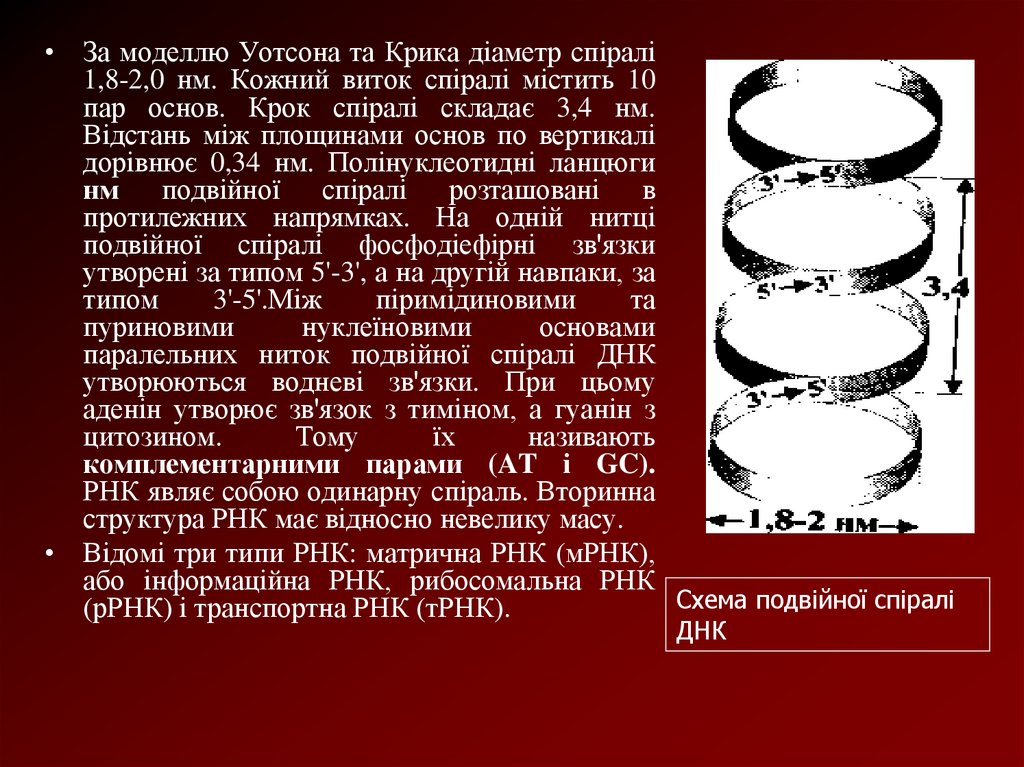
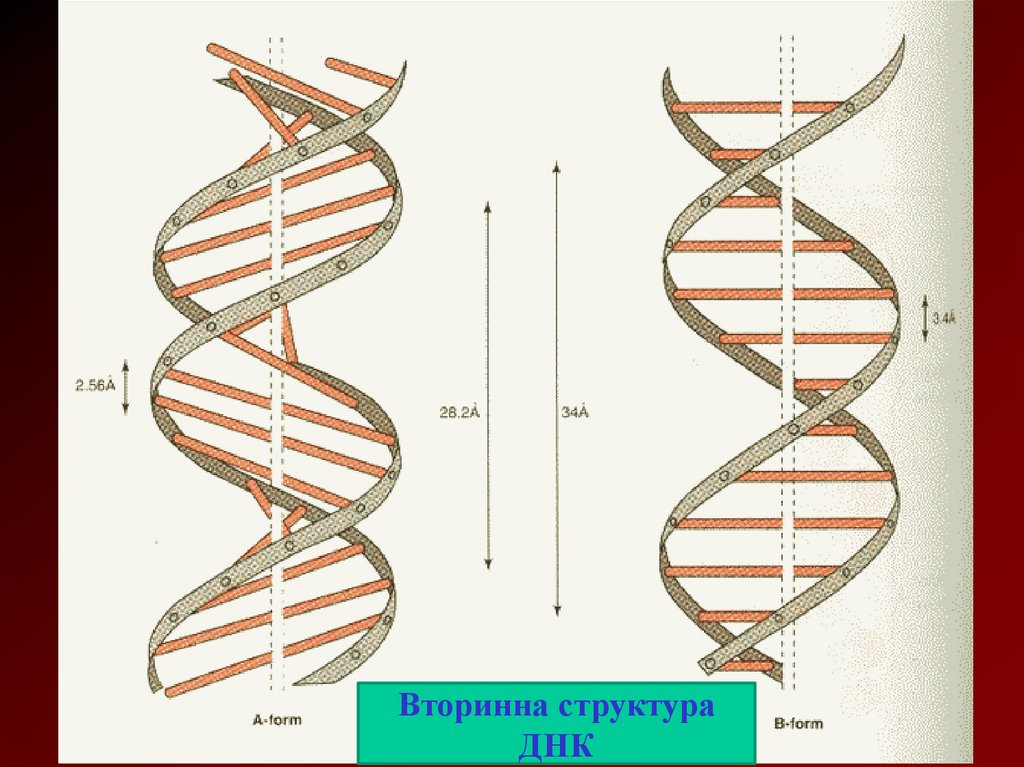
• За моделлю Уотсона та Крика діаметр спіралі1,8-2,0 нм. Кожний виток спіралі містить 10
пар основ. Крок спіралі складає 3,4 нм.
Відстань між площинами основ по вертикалі
дорівнює 0,34 нм. Полінуклеотидні ланцюги
нм подвійної спіралі розташовані в
протилежних напрямках. На одній нитці
подвійної спіралі фосфодіефірні зв'язки
утворені за типом 5'-3', а на другій навпаки, за
типом
3'-5'.Між
піримідиновими
та
пуриновими
нуклеїновими
основами
паралельних ниток подвійної спіралі ДНК
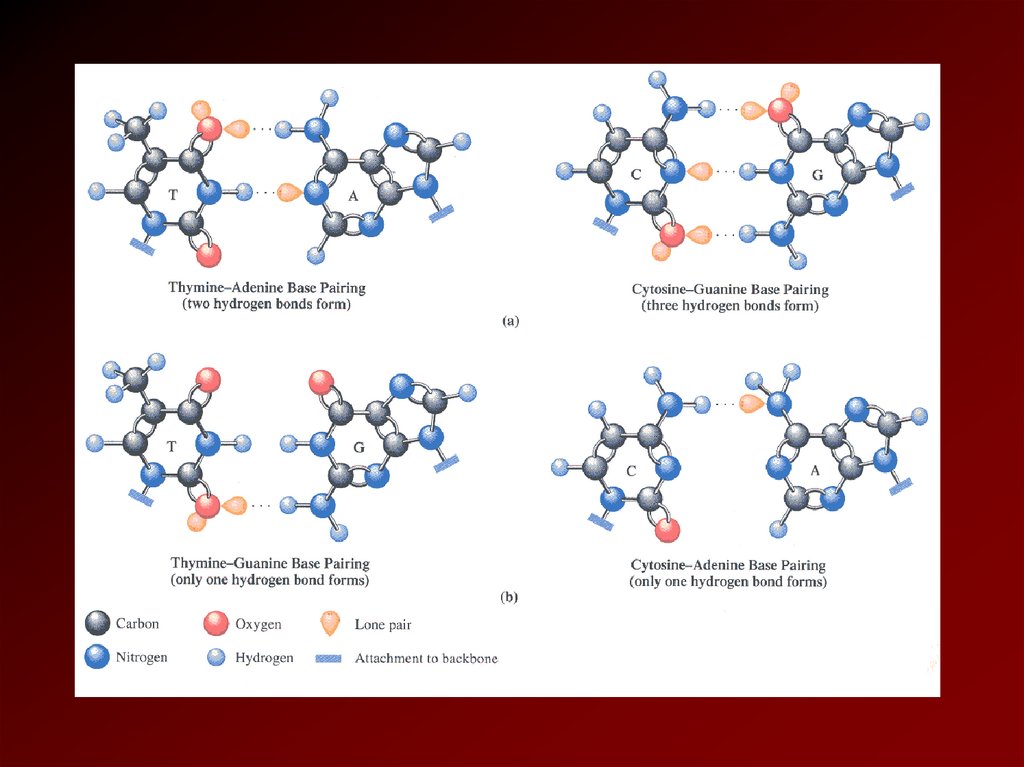
утворюються водневі зв'язки. При цьому
аденін утворює зв'язок з тиміном, а гуанін з
цитозином.
Тому
їх
називають
комплементарними парами (AT і GC).
РНК являє собою одинарну спіраль. Вторинна
структура РНК має відносно невелику масу.
• Відомі три типи РНК: матрична РНК (мРНК),
або інформаційна РНК, рибосомальна РНК
Схема подвійної спіралі
(рРНК) і транспортна РНК (тРНК).
ДНК
19. Утворення нуклеотидів
• Нуклеотиди утворюються при взаємодії пентози з азотистоюосновою (утворюючи N-глікозидний зв'язок при взаємодії з
глікозидним гідроксилом пентози та нітрогеном N-1
піримідинових та N-9 пуринових основ) і залишком
фосфорної кислоти (вз 3 або 5 гідроксилом) :
20. Структура
• ПервинаСтруктура
21.
22.
Вторинна структураДНК
23.
24.
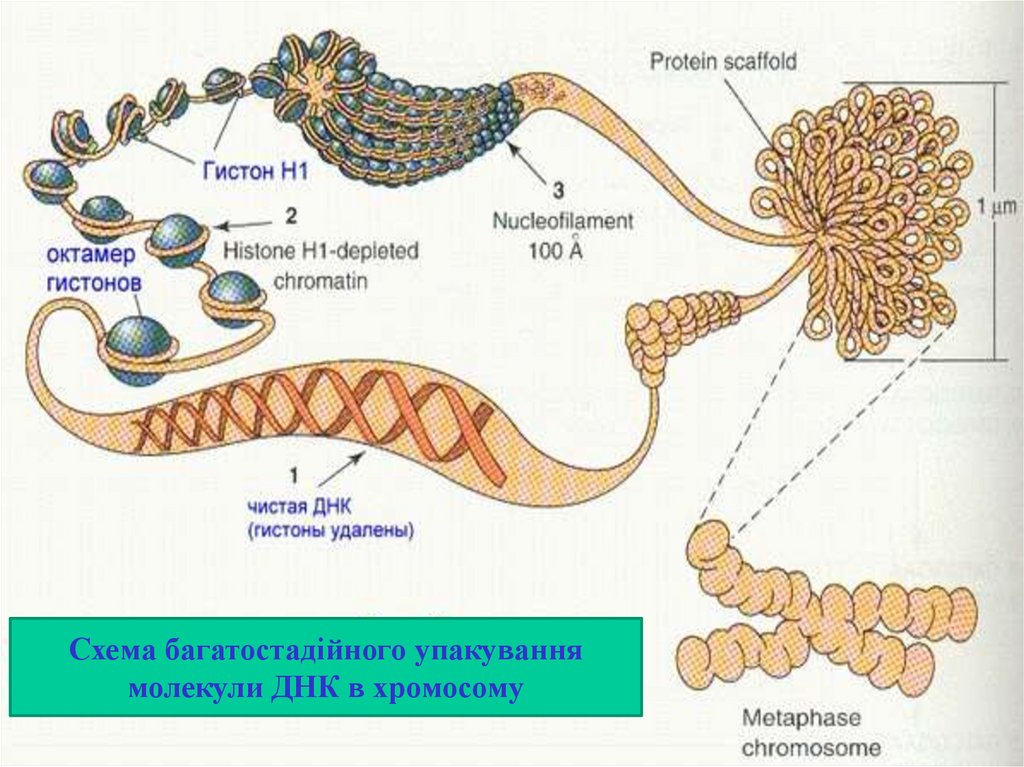
Схема багатостадійного упакуваннямолекули ДНК в хромосому
25.
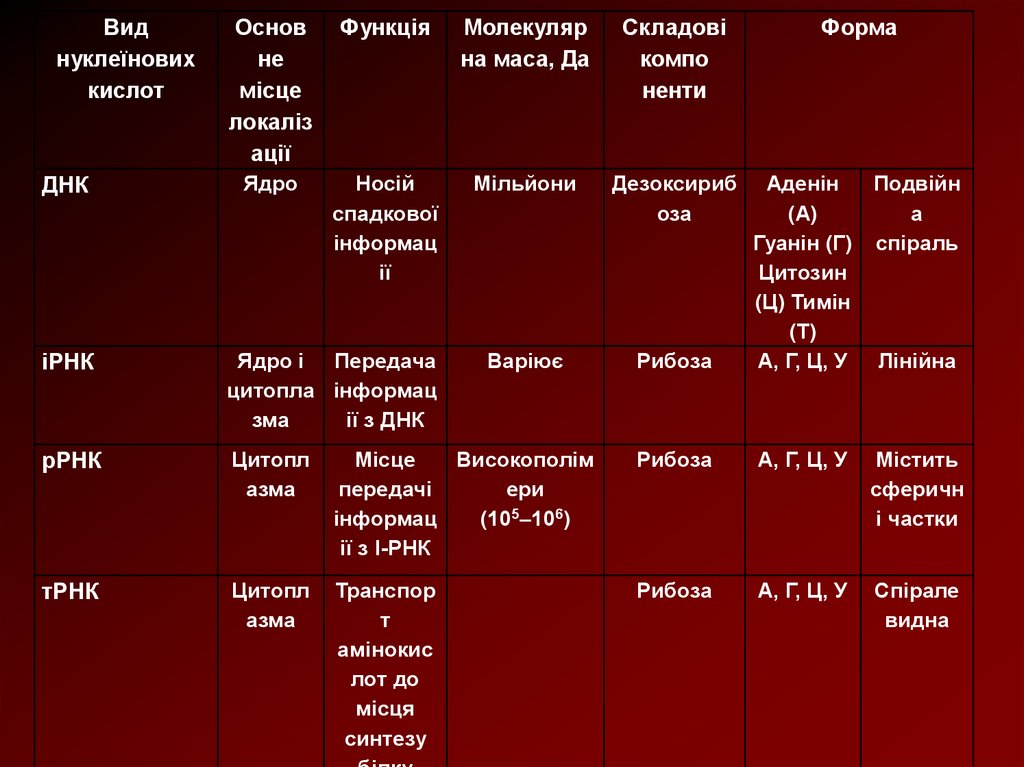
Виднуклеїнових
кислот
ДНК
Основ
не
місце
локаліз
ації
Функція
Молекуляр
на маса, Да
Складові
компо
ненти
Форма
Ядро
Носій
спадкової
інформац
ії
Мільйони
Дезоксириб
оза
Варіює
Рибоза
Аденін
Подвійн
(А)
а
Гуанін (Г) спіраль
Цитозин
(Ц) Тимін
(Т)
А, Г, Ц, У Лінійна
іРНК
Ядро і Передача
цитопла інформац
зма
ії з ДНК
рРНК
Цитопл
азма
Місце
Високополім
передачі
ери
інформац
(105–106)
ії з І-РНК
Рибоза
А, Г, Ц, У
Містить
сферичн
і частки
тРНК
Цитопл
азма
Транспор
т
амінокис
лот до
місця
синтезу
Рибоза
А, Г, Ц, У
Спірале
видна
26. Еукаріотична клітина має три основні частини: 1) ядро; 2) цитоплазму; 3) оболонку
27. Ядро (лат. nucleus, грец. karion) забезпечує процеси біосинтезу й передачі спадкової інформації та складається із:
1) ядерної оболонки;2) ядерця;
3) каріоплазми;
4) хроматину
28.
У 1825 р. Я. Пуркіньє вперше спостерігав клітинні ядрав яйцеклітині курки.
У 1833 р. Р. Броун описав ядро у рослинних клітинах.
У 1839 р. Т. Шванн описав ядро у тваринних клітинах.
Клітини
бувають
двоядерні
(клітини деяких водоростей).
(хрящові),
багатоядерні
29.
Функції ядраЗбереження генетичної (спадкової) інформації серед
клітинних поколінь;
Підтримання постійної структури ДНК за допомогою так
званих репараційних ферментів, які можуть відновити молекулу
ДНК після її ушкоджень (у тому числі, радіаційних);
Редуплікація молекул ДНК (тобто якісне і кількісне
подвоєння генетичного матеріалу);
Розподіл генетичного матеріалу між дочірніми клітинами
під час мітозу; рекомбінація генетичного матеріалу в процесі
мейозу.
Реалізація генетичної інформації, тобто полягає у створенні
апарату білкового синтезу.
Синтез усіх видів РНК (інформаційної, транспортної,
рибосомної), а також побудова рибосом.
30.
Форма ядра:Найчастіше сферична;
зустрічаються
паличкоподібна,
бобовидна,
кільцеподібна,
сегментована.
Форма ядра залежить від:
форми
клітини
(видовжені
клітини
гладких
м'язів
мають
видовжене паличкоподібне ядро);
від кількості включень (ядро жирової клітини набуває сплющеної
форми під впливом великої жирової краплі, що займає майже всю
клітину);
розташування органел (форма ядра моноцита бобовидна завдяки
розташуванню в місці його заглибини центросоми).
31.
В еукаріотичних клітинах основний генетичний матеріалзосереджений в ядрі.
До складу сухої речовини ядра входить 80% білків, 12%
ДНК, 5% РНК, 3% ліпідів і деяка кількість Мангану і
Марганцю.
Усі клітини людського організму містять ядро, за винятком
високоспеціалізованих клітин крові — еритроцитів, що
втрачають ядро у процесі свого розвитку і є без'ядерними.
Переважна більшість клітин містить одне ядро, але бувають
двоядерні клітини (20% клітин печінки є двоядерними), а
також багатоядерні (остеокласти — клітини кісткової
тканини).
32. Остеокласт
Адипоцити33. Клітини крові (різна форма ядер)
ТромбоцитиКісткова тканина
Нейтрофільний
гранулоцит
Еозинофільний
гранулоцит
Еритроцит
Базофільний
гранулоцит
Моноцит
Лімфоцит
Клітини крові (різна форма ядер)
34.
Ядро локалізується завжди у певному місці клітини. Наприклад, уциліндричних клітинах шлунка, кишки воно займай базальне положення.
Розміри ядер під 3-4 до 40 мкм. Кожний тип клітини має своє постійне
співвідношення між об'ємом ядра і цитоплазми. Ця константа носить
назву індексу Гертвіга. Згідно значень цього індексу клітини поділяють
на ядерні (з великим індексом Гертвіга) та цитоплазматичні (з малим
індексом Гертвіга).
Ядро може бути в двох станах — мітотичному (під час поділу)
та інтерфазному (між поділами). Останнє називають також метаболічним
ядром, що підкреслює його функціональний стан.
У живій клітині інтерфазне ядро виглядає оптично пустим, видно
лише ядерце. Структури ядра у вигляді ниточок, зерняток у живій клітині
можна спостерігати лише при дії на неї пошкоджуюючих агентів, коли
клітина переходить у стан так званого паранекрозу (стан на межі життя і
смерті). З цього стану клітина може або повернутися до нормальної
життєдіяльності, або загинути. Морфологічно розрізняють такі зміни ядра
при загибелі клітини: каріопікноз (ущільнення), каріорексис (розлад),
каріолізис (розчинення).
35.
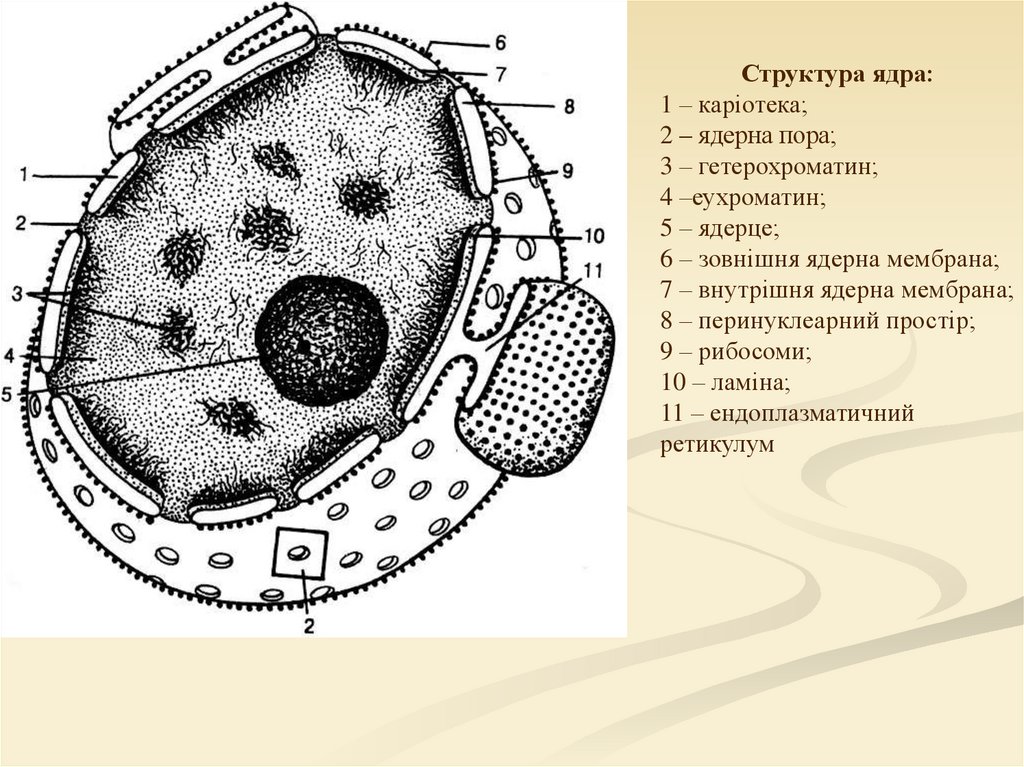
Структура ядра:1 – каріотека;
2 – ядерна пора;
3 – гетерохроматин;
4 –еухроматин;
5 – ядерце;
6 – зовнішня ядерна мембрана;
7 – внутрішня ядерна мембрана;
8 – перинуклеарний простір;
9 – рибосоми;
10 – ламіна;
11 – ендоплазматичний
ретикулум
36. Електронна мікрофотографія інтерфазного ядра
37. Ядерна оболонка (нуклеолема, каріолема, каріотека)
• Структура, що відокремлює ядро відцитоплазми, дає змогу здійснюватися обміну
між ядром і цитоплазмою
Складається із зовнішньої та внутрішньої
мембрани, товщина кожної приблизно 8 нм.
Мембрани
розділені
між
собою
перинуклеарним простором, завширшки 25
нм.
Значення
розділу цих структур дуже
важливе: це забезпечує відособлення
процесів синтезу білка і процесів синтезу
нуклеїнових кислот, що дозволяє, на відміну
від прокаріотів, регулювати генну активність
та реалізовувати її у вигляді синтезу
специфічних білків.
38.
Зовнішня мембрана зовнішньої оболонки частопереходить
у
мембрани
гранулярного
ендоплазматичного ретикулума.
Цитоплазматичний
бік
ядра
часто
вкритий
рибосомами.
Найбільш
характерними
структурами
ядерної
оболонки є ядерні пори, які утворюються внаслідок
злиття зовнішньої та внутрішньої ядерних мембран у
вигляді округлих наскрізних отворів або перфорацій з
діаметром біля 100 нм.
Через пори здійснюється вибірковий транспорт
молекул: з цитоплазми в ядро транспортуються
ферменти для синтезу РНК, гормони, деякі регулятори
синтетичних процесів, а з ядра в цитоплазму – різні
види РНК.
39.
Функції ядерної оболонкибар'єрна функція: ядерна оболонка відокремлює вміст ядра,
його генетичний матеріал від цитоплазми, обмежує вільний доступ
в ядро та вихід із нього різних речовин.
регуляція транспорту макромолекул між ядром і цитоплазмою.
Наприклад, відомо, що гістони та інші негістонові білки після
синтезу в цитоплазмі мігрують у ядро. Відомий також і зворотний
процес транспорту речовин з ядра в цитоплазму. Це, у першу чергу,
стосується транспорту РНК, що синтезуються виключно в ядрі.
Механізм транспорту високомолекулярних сполук, а також рибосом
через ядерну оболонку не зовсім зрозумілий, можливо, він
здійснюється через пори.
участь у створенні внутрішньоядерного порядку шляхом
фіксації хромосомного матеріалу в інтерфазі до внутрішньої
ядерної мембрани.
40.
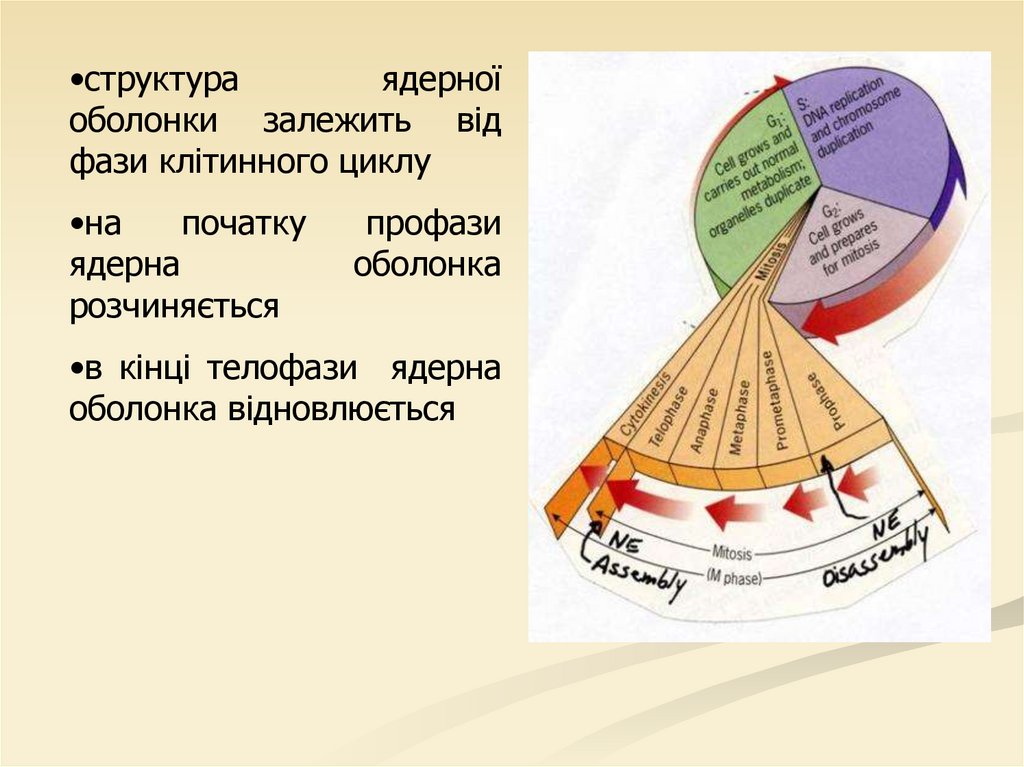
•структураядерної
оболонки залежить від
фази клітинного циклу
•на
початку
ядерна
розчиняється
профази
оболонка
•в кінці телофази ядерна
оболонка відновлюється
41.
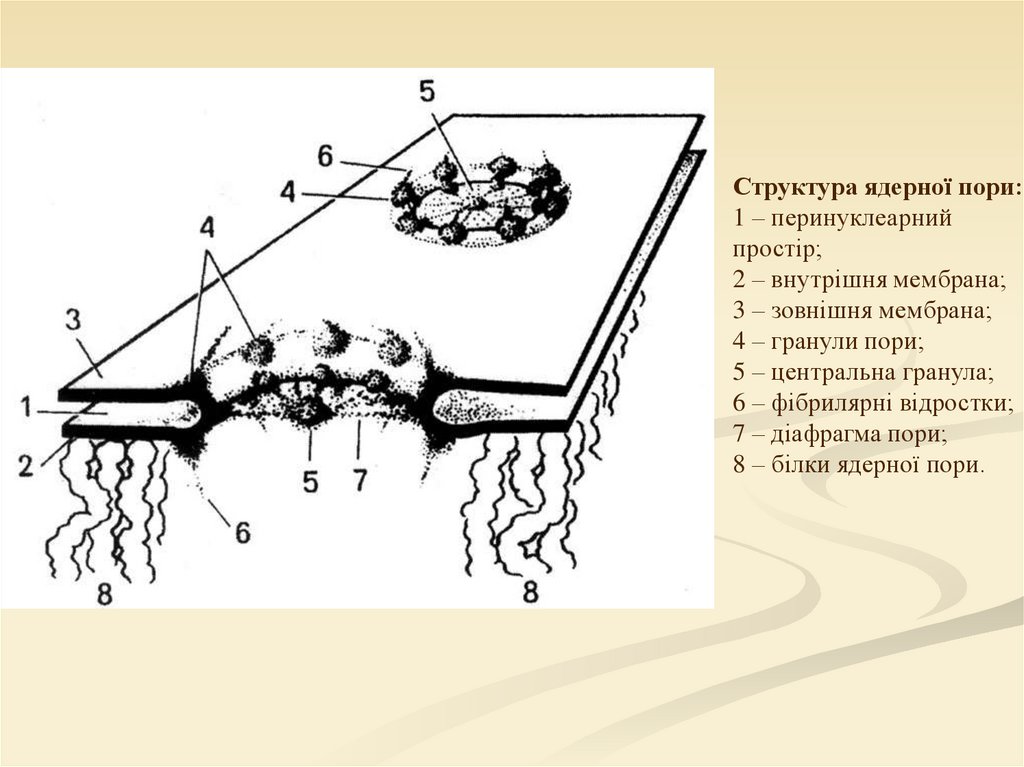
Структура ядерної пори:1 – перинуклеарний
простір;
2 – внутрішня мембрана;
3 – зовнішня мембрана;
4 – гранули пори;
5 – центральна гранула;
6 – фібрилярні відростки;
7 – діафрагма пори;
8 – білки ядерної пори.
42. Каріоплазма, або ядерний сік
• рідка частинаядра,
желеподібний
колоїд
• середовище в
якому
перебувають
ядерця та
хромосоми
43. Хроматин
• інтерфазна форма існування хромосом(структурний аналог хромосом)
• хромосоми видно лише під час поділу
клітини
• це комплекс ДНК і білків – дезоксирибонуклеопротеїн
• складається з хромосомних фібрил
(ниток) товщиною 20-25 нм
• у хроматині виявлено невелику
кількість РНК — продуктів процесу
транскрипції. Співвідношення вказаних
хімічних компонентів у хроматині
ДНК : білок : РНК = 1: 1,3 : 0,2.
• основу структурної організації фібрил
складають нуклеосоми.
• при поділі клітини ДНК спіралізується, і
хроматин перетворюється в хромосоми.
44.
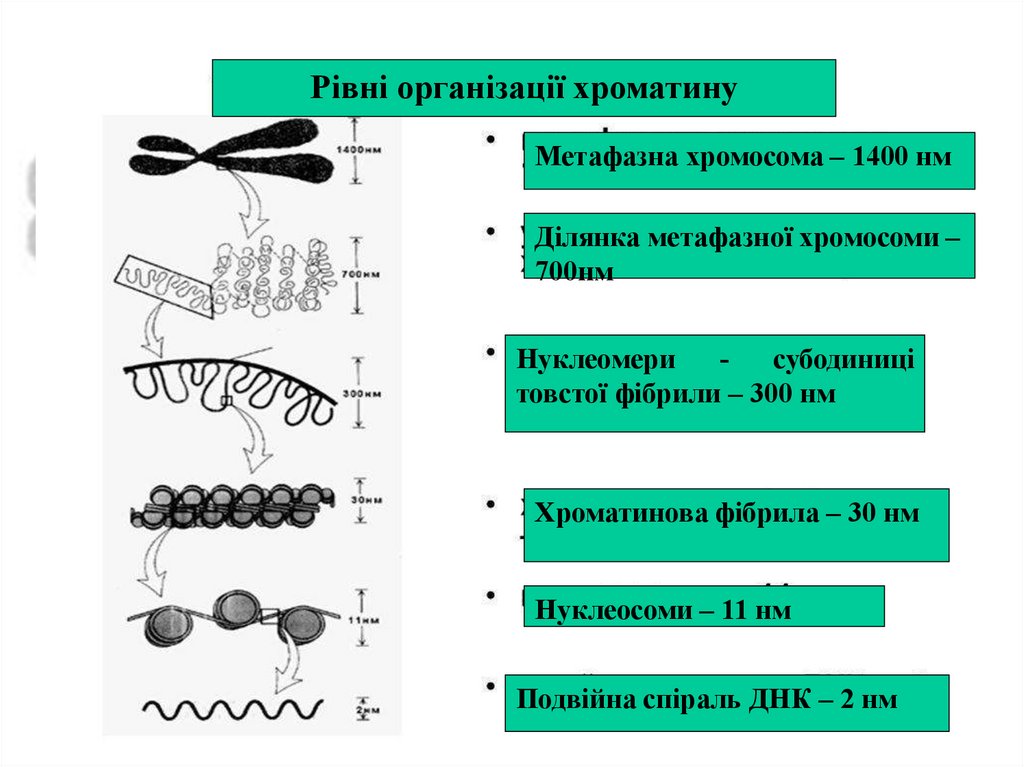
Рівні організації хроматинуМетафазна хромосома – 1400 нм
Ділянка метафазної хромосоми –
700нм
Нуклеомери
субодиниці
товстої фібрили – 300 нм
Хроматинова фібрила – 30 нм
Нуклеосоми – 11 нм
Подвійна спіраль ДНК – 2 нм
45. Будова хроматину
46.
• Нуклеосома – це часточка, що має вигляд нанизанихна нитку намистин.
• Щільно упаковані у вигляді спіралі товщиною 36 нм,
на кожний виток припадає приблизно 6 нуклеосом.
• Нуклеосому можна уявити у вигляді циліндра з 2
нитками ДНК, які закручені зовні навколо нього.
47.
Види хроматину• Гетерохроматин відповідає конденсованим під час
інтерфази ділянкам хромосом; він є функціонально
неактивним. Цей хроматин добре забарвлюється, саме
його
можна
бачити
на
гістологічному
препараті.
Гетерохроматин
поділяється
на структурний (це ділянки хромосом, що постійно
конденсовані)
та
факультативний
(може
деконденсунатись і переходити в еухроматин).
• Еухроматин відповідає деконденсованим в інтерфазі
ділянкам хромосом. Це робочий, функціонально
активний хроматин. Він не забарвлюється, його не
видно на гістологічному препараті. Під час мітозу
весь еухроматин конденсується і входить до складу
хромосом.
48. Рівні спіралізації хромосом
ДНК
Нуклеосома
Хроматинова фібрила
Хроматида
Конденсована хромосома
49. Взаємозв’язок між структурою хроматину та організацією хромосом
50. Хромосоми
Хромосоми — це щільні паличко- або ниткоподібнітільця діаметром 0,2-2 мкм і довжиною у людини від
1,5 до 10 мкм, які добре забарвлюються основними
барвниками і які помітні в ядрі клітини під час
мітотичного поділу. Назву їм дав В. Вальдеєр.
51. Структурна організація метафазних хромосом
• Кожна хромосомаскладається з двох
хроматид –
конденсованих подвійних
ланцюгів ДНК
• Сестринські хроматиди
сполучаються між собою
в ділянці центромери
52. Структурна організація метафазних хромосом
1 - довге плече;2 - коротке плече;
3 - центромера;
4- вторинна перетяжка;
5 - супутник;
6 - хроматиди
53.
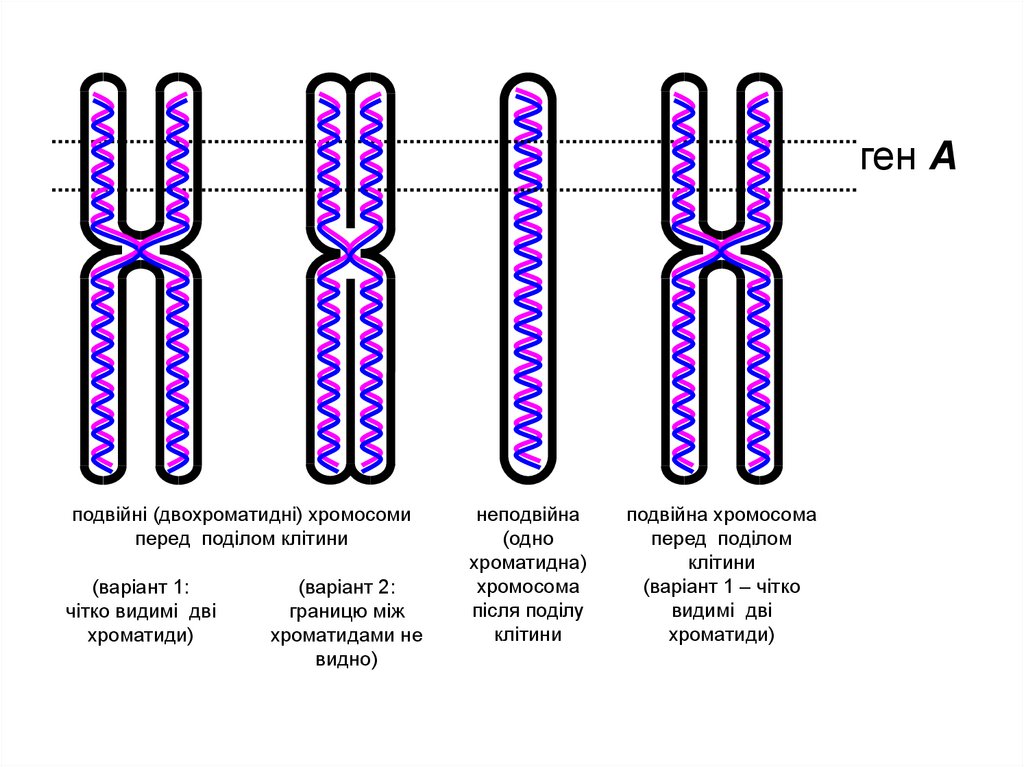
ген Аподвійні (двохроматидні) хромосоми
перед поділом клітини
(варіант 1:
чітко видимі дві
хроматиди)
(варіант 2:
границю між
хроматидами не
видно)
неподвійна
(одно
хроматидна)
хромосома
після поділу
клітини
подвійна хромосома
перед поділом
клітини
(варіант 1 – чітко
видимі дві
хроматиди)
54. Типи метафазних хромосом людини
1 – метацентрична;2 – субметацентрична;
3 – акроцентрична з супутником;
4 – акроцентрична;
а – первинна перетяжка (центромера)
б – вторинна перетяжка
55. Каріотип – диплоїдний набір хромосом клітини, який характеризується: кількістю хромосом, певними розмірами, формою, будовою
Диплоїднийнабір хромосом
(2n) – повний
набір хромосом,
міститься у
соматичних
клітинах
Гаплоїдний набір
хромосом (n) –
половинний набір
хромосом,
міститься у
статевих
клітинах
56. ДНК в хромосомах
• ДНК в складіхромосом
зв’язана з
білками-гістонами
• Один комплекс із
гістонів і ДНК
називається
нуклеосома
• Послідовність
нуклеосом
багатократно
спіралізована.
57. Каріотип людини
58. Ідіограма – розташування пар хромосом у порядку зменшення їх розмірів
• Гомологічні хромосоми –хромосоми однієї пари, які
однакові за розмірами, формою
• Хромосомні набори чоловіка
та жінки відрізняються між
собою за статевими
хромосомами (2)
• Автосоми (44) – хромосоми,
які однакові в обох статей
• 2n=44a+XX (каріотип жінки)
2n=44a+XY (каріотип чоловіка)
59. Хромосомний набір (каріотип) чоловіка
60.
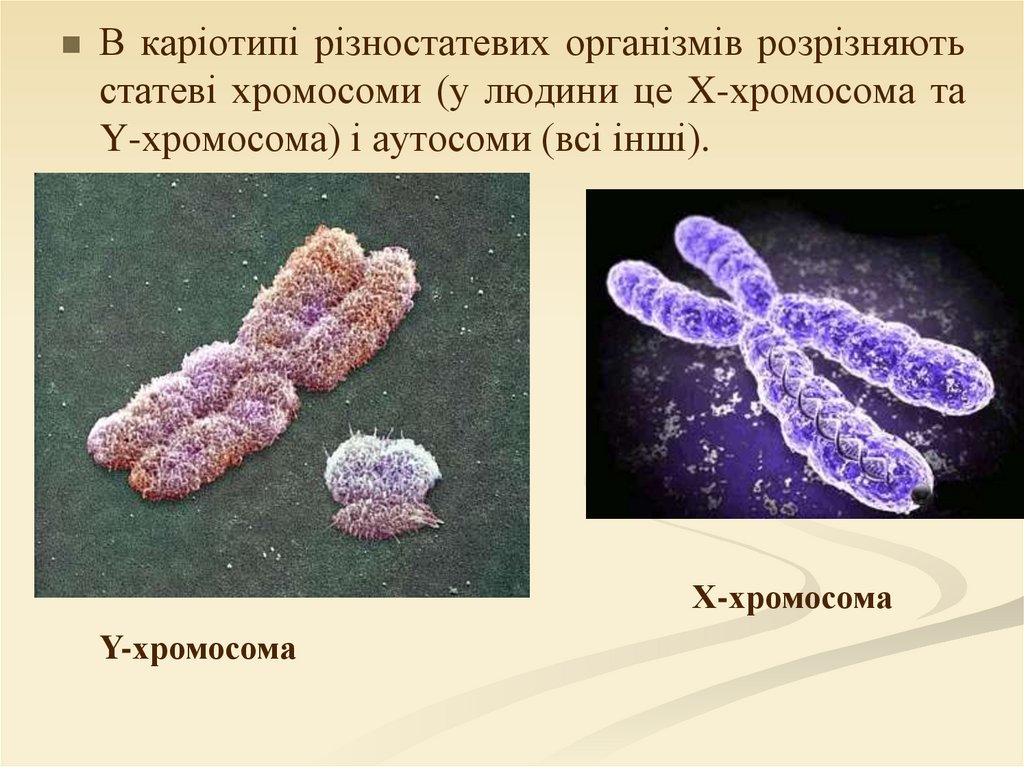
В каріотипі різностатевих організмів розрізняютьстатеві хромосоми (у людини це X-хромосома та
Y-хромосома) і аутосоми (всі інші).
Х-хромосома
Y-хромосома
61. ПОРУШЕННЯ СТРУКТУРИ ХРОМОСОМ
• Порушення структури хромосом відбувається в результатіспонтанних змін:
• Генні мутації (зміни на молекулярному рівні)
• Хромосомні мутації (мікроскопічні зміни):
–
–
–
–
делеції
дуплікації
транслокації
інверсії
62. Хромосомна мутація: ДЕЛЕЦІЯ
-від лат. deletio — знищення —хромосомна аберація (перебудова),
при якій відбувається втрата ділянки
хромосоми.
63. Хромосомна мутація: ДУПЛІКАЦІЯ
від лат. duplicatio — подвоєння —структурна
полягає
в
хромосоми.
хромосомна
мутація,
подвоєнні
ділянки
64. Хромосомна мутація: ТРАНСЛОКАЦІЯ
• Впроцесі
ділянками
транслокації
відбувається
негомологічних
хромосом,
загальне число генів не змінюється.
обмін
але
65. Хромосомна мутація: ІНВЕРСІЯ
• Змінаструктури
хромосоми,
яка
викликана
поворотом на 180° однієї з внутрішніх її ділянок.
66.
ЯДЕРЦЕЯдерце — це найбільш щільна структура ядра (щільність ядерця в 1,5 раза
перевищує щільність ядра), яка добре помітна у живій незабарвленій клітині.
Форма ядерець сферична, розмір 1-5 мкм.
Ядерце добре забарвлюється, особливо основними барвниками. Це пов'язано з
наявністю у ньому великої кількості РНК, концентрація якої тут у 2—8 разів вища, ніж
у ядрі, і в 2-3 рази перевищує концентрацію у цитоплазмі. Кількість ядерець, як
правило, відповідає кількості хромосомних наборів. Тому в диплоїдних клітинах їх
буває два на ядро.
Ядерце — це не самостійна організована структура, а похідне хромосом, які
містять так звані ядерцеві організатори, що здебільшого розташовані у зонах
вторинних перетяжок. Останні являють собою локуси хромосом з найбільш високою
концентрацією і активністю синтезу РНК в інтерфазі.
Ядерце — це місце утворення рибосомних РНК і самих рибосом. ДНК ядерцевого
організатора складається із множинних копій генів рРНК: на кожному з них
синтезується попередник рРНК, який у зоні ядерця одягається білком; потім тут
утворюються субодиниці рибосом.
Субмікроскопічна будова ядерця характеризується наявністю двох основних
структур: гранул діаметром 15-20 нм і фібрил товщиною 6-8 нм.
Гранулярний компонент часто розташовується у вигляді нитки, нуклеолонеми
товщиною 0,2 мкм.
Фібрилярний компонент ядерця — це рибонуклеопротеїнові тяжі, попередники
рибосом, а гранули — субодиниці рибосом, що дозрівають. Навколо ядерця
знаходиться компактна зона навколоядерцевого гетерохроматину. Конденсований
хроматин також розміщений поміж петлями нуклеолонеми.
67. Ядерця
• щільні, сильнозабарвлені
гранулярні
утворення, які не
мають мембрани
• відбувається
синтез
рибосомальної РНК
• утворення
субодиниць рибосом
• синтез ядерних
білків (гістонів)