









































































































 biology
biologySimilar presentations:



")
")





Будова ядра. Генетичний апарат. Нуклеїнові кислоти
1. Будова ядра. Генетичний апарат. Нуклеїнові кислоти
К.б.н. Полковенко О.В.2.
Включення 3Н-тімідіну в ядра остеокластів через 72 год. після введеннярадіонукліда. Гіпокінезія. Гісторадіоавтограф, гематоксилін-тіонін-еозін.
Об.Х100, ок. 12,5.
3.
Structure of the nucleus:1 — internal membrane;
2 — external membrane;
3 — pore;
4 — nucleoli;
5 — chromatin;
6 — nuclear sap
4.
5.
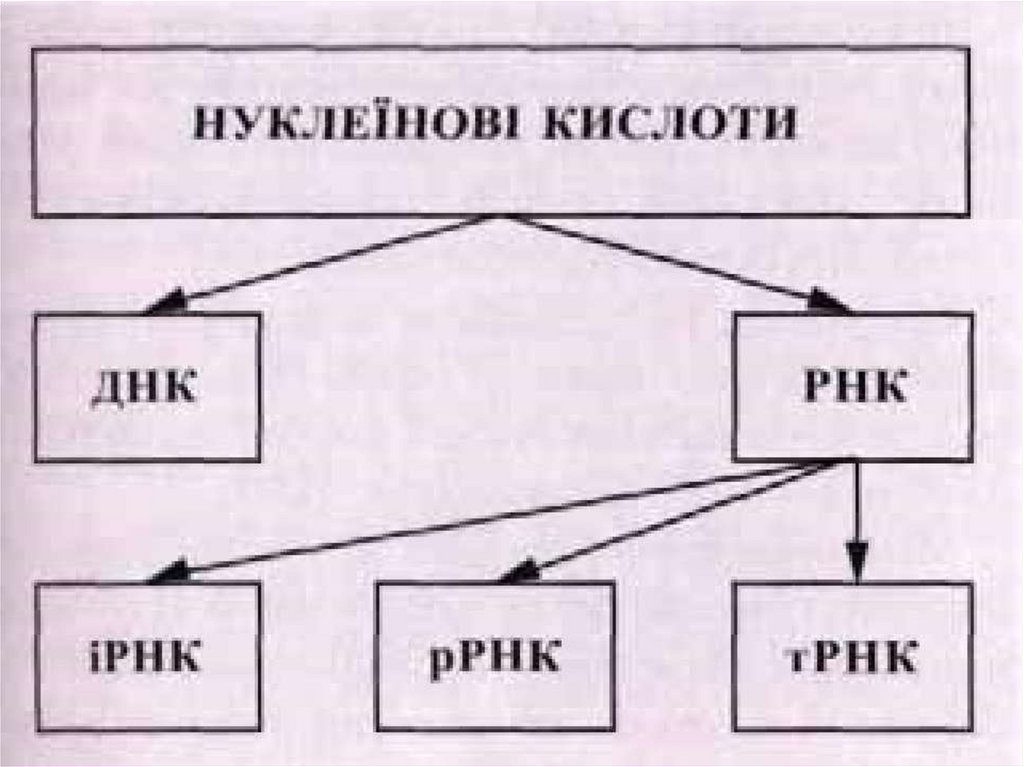
Нуклеїнові кислоти (лат. nucleus —ядро). Ці речовини вперше було
виявлено і виділено з ядер клітин. Є два
види нуклеїнових кислот:
дезоксирибонуклеїнова (ДНК) і
рибонуклеїнова (РНК). Основна кількість
ДНК зосереджена в хромосомах клітини
і лише невелика її кількість міститься в
мітохондріях і пластидах. РНК міститься
в ядерцях, а також у цитоплазмі.
Молекула ДНК — це дуже довгий
полінуклеотидний ланцюг, довжина
його може досягати десяти міліметрів.
6.
7.
8.
Так, вважають, що сумарна довжинамолекул ДНК 46 хромосом однієї клітини
людини становить 170—180 см.
Відповідно дуже велика і молекулярна
маса ДНК (сотні мільйонів умовних
одиниць). Кожна молекула ДНК
складається з двох сполучених між
собою ланцюгів нуклеотидів. До складу
кожного нуклеотиду входять азотиста
основа, дезоксирибоза і фосфорна
кислота. Всього в ДНК є чотири види
азотистих основ: аденін (А), гуанін (Г),
тимін (Т) і цитозин (Ц)
9.
Нуклеотиди різняться лише азотистимиосновами. Назва нуклеотидів також
пов'язана з назвою нуклеозидів (сполук
азотистих основ з пентозою) цих основ.
Наприклад, нуклеотид, який містить аденін,
утворюється приєднанням залишку
фосфорної кислоти до аденозинового
нуклеозиду (аденін + рибоза або
дезоксирибоза), називається аденіловим.
Відповідно утворюються нуклеотиди
гуанозинового, уридинового, цитидинового і
тимідинового нуклеозидів.
10.
Два ланцюги, які складаються з десятківі сотень нуклеотидів, скручуються між
собою й утворюють подвійну спіраль.
Діаметр цієї спіралі становить 2 • 10~9
м (2 нм), а відстань між сусідніми
нуклеотидами — 3,4 • 10~10 м (0,34 нм;
1 нм = 10~9 м). Розмір витка спіралі
(крок спіралі) становить 3,4 • 10~9 м
(3,4 нм), у ньому розміщується 10 пар
нуклеотидів.
11.
Азотисті основи одного ланцюгасполучені з основами другого ланцюга
за допомогою водневих зв'язків у
такому порядку: аденін одного ланцюга
сполучений з тиміном другого ланцюга;
гуанін сполучений так само з
цитозином. Сполучення інших типів у
нормі ніколи не виникають. Це
пояснюється тим, що між такими парами
нуклеотидів, як аденів С — тимін і
гуанін — цитозин, існує особливий тип
зв'язку, який дістав назву
комплементарного (принцип
комплементарності; лат. complementum
— доповнення).
12.
Подвійна спіраль молекули ДНК здатна розкручуватися,при цьому водневі зв'язки розриваються й окремі
ланцюги ДНК відходять один від одного. Іноді таке
роз'єднання ланцюгів відбувається не по всій довжині
молекули, а лише на певній ділянці. Ланцюги ДНК
можуть знову спіралізуватися, відновлюючи свою
попередню структуру.
Редуплікація (лат. duplicatio — подвоєння), реплікація
(англ. replica — відбиток) ДНК — процес
самовідтворення макромолекул нуклеїнових кислот,
який забезпечує точне копіювання генетичної
інформації і передавання її з покоління в покоління.
Самоподвоєння молекули ДНК відбувається в період
інтерфази перед поділом клітин. При цьому молекула
ДНК розкручується і з одного кінця спіраль
розділяється на окремі ланцюги.
13.
14.
Біля кожного з них із вільних нуклеотидів, якіє в ядрі клітини, розпочинається синтез
другого ланцюга ДНК. Цей синтез
відбувається за принципом
комплементарності. В результаті замість однієї
молекули ДНК утворюються дві молекули
такого самого нуклеотидного складу, як і
початкова. Один ланцюг у кожній
новоутвореній молекулі ДНК походить від
початкової молекули (материнський ланцюг),
а другий синтезується заново (дочірній). Як
процес розділення молекули ДНК на два
ланцюги, так і процес синтезу нових ланцюгів
здійснюється за рахунок дії низки ферментів
(ДНК-полімерази, ДНК-лігази).
15.
Генетична інформація геному складається з генів. Ген єелементарною структурно-функціональною одиницею
спадковості, що визначає розвиток певної ознаки клітини
або організму. Внаслідок передачі генів у ряді поколінь
забезпечується успадкування ознак батьків.
Г. Мендель був першим, хто в 1865 р. стверджував про одиницю
спадковості, він назвав її "спадковим фактором". Слово "ген" було
введено В. Йогансеном у 1909 р. для позначення одиниці
спадковості, що займає особливе місце (локус) у хромосомі. У
1948 р. Дж. Бідл і Е. Тейтем запропонували гіпотезу "один ген один поліпептид" і розглядали ГЕН ЯК ОДИНИЦЮ
СПАДКОВОГО МАТЕРІАЛУ, ЩО МІСТИТЬ ІНФОРМАЦІЮ
ДЛЯ УТВОРЕННЯ ОДНОГО БІЛКА.
16.
Відповідно до сучасної концепції, ген — одиницяпередачі спадкової інформації у вигляді
безперервної ділянки ДНК, яка обумовлює
певну функцію в організмі або забезпечує
транскрипцію іншого гена. Отже, одна і та ж
ДНК-послідовність може кодувати не один, а
декілька різних поліпептидів.
Гени в ДНК розташовані у лінійному порядку.
Кожен ген має своє місце розташування (локус).
Теломерні та центромерні ділянки хромосом не
містять генів.
Сукупність всіх одиниць інформації (генів), які
містяться в клітині називають геномом, а сукупність
генів даної клітини або організму складає його
генотип.
17.
Ген як одиниця генетичної інформаціїзабезпечує такі функції:
• зберігання спадкової інформації;
• керування біосинтезом білків та інших сполук у
клітині;
• редуплікації ДНК і РНК (подвоєння генів під час
поділу);
• репарації (відновлення) пошкоджених ДНК і РНК;
• забезпечення спадкової мінливості клітин і
організмів;
• контроль за індивідуальним розвитком клітин і
організмів;
• явище рекомбінації.
18.
У геномах еукаріотів спостерігається надлишковість ДНК, тобтонезначна частина загальної послідовності геному (наприклад, 1,5 %
геному людини) кодує білки – тобто містить кодуючі гени (екзони),
які чергуються з послідовностями, які не транслюються в
амінокислотні послідовності (інтрони). Крім того, значна частина
геному використовується для здійснення процесів ембріонального
розвитку, диференціювання, росту і надалі не експресується.
Екзон — ділянка гена (ДНК) еукаріот, яка несе генетичну
інформацію, що кодує синтез білка. Ділянки ДНК, які відповідають
екзонам, на відміну від інтронів, повністю представлені в молекулі
інформаційної РНК, що кодує первинну структуру білка.
Інтрон— ділянка гена (ДНК) еукаріот, яка, як правило, не несе
генетичної інформації, що відповідає за синтез білка, кодованого
даним геном. Інтрони розміщені між екзонами і представлені
лише в первинному транскрипті — попередникові іРНК (про-іРНК),
при дозріванні іРНК вони видаляються (а екзони залишаються).
19.
Крім того, до некодуючих послідовностейДНК також відносяться:
промотори – гени, що безпосередньо
передують відкритим рамкам зчитування;
псевдогени — копії генів, інактивовані в
результаті мутацій, які іноді можуть
служити початковим матеріалом для
дуплікації і подальшої дивергенції генів;
20.
Залежно від організації нуклеотидів у ДНК, структури тафункцій розрізняють такі типи нуклеотидних
послідовностей:
1) структурні гени – несуть інформацію про структуру певних
поліпептидів, із цих ділянок ДНК транскрибується іРНК, яка спрямовує
синтез ферментів і структурних білків;
2) регуляторні гени (промотори, оператори) – контролюють і
регулюють активність інших генів та процес біосинтезу білка;
3) сателітна ДНК – містить велику кількість повторюваних груп
нуклеотидів, що не мають змісту і не транскрибуються, проте поодинокі
гени серед сателітної ДНК, звичайно, мають регуляторну або
посилювальну дію на структурні гени;
4) кластери генів – групи різних структурних генів у певній ділянці
хромосоми, об'єднані загальними функціями;
5) спейсери – великі неінформативні ділянки ДНК, що знаходяться між
кластерами, відокремлюють гени один від одного і не транскрибуються;
6) повторювані гени – один і той самий ген багаторазово
повторюється (декілька сотень раз); не відокремлюючись один від
одного, вони створюють тандеми.
21.
У клітині виявлено білок-репресор. Якийген кодує амінокислотну послідовність
цього білка ?
– Ген-промотор
– Ген-термінатор
+ Ген-регулятор
– Ген-модифікатор
– Ген-оператор
22.
A repressor protein has been found inthe cell. What gene encodes the amino
acid sequence of this protein?
- The promoter-general
- The gene terminator
+ Gene regulator
- Gen-modifier
- The general operator
23.
Таким чином, гени – це ділянки кодуючоїпослідовності ДНК клітин разом із регуляторними
ділянками.
З’ясовано, що гени еукаріотів є переривчастими,
мозаїчними, вони складаються з кодуючих ділянок —
екзонів, розділених некодуючими — інтронами.
Структурні гени — це кістяк геному, оскільки вони
кодують структуру білків, визначають чергування
амінокислотних залишків у поліпептидному ланцюгу.
Регуляторні гени виконують регуляторні функції і
контролюють експресію (прояв) структурних генів.
Виявлені і так звані псевдогени, які не функціонують.
24.
Поряд із стаціонарними генами, що локалізовані у певних ділянкаххромосом, у геномах еукаріотів також містяться мобільні (нестабільні)
генетичні елементи — послідовності ДНК, здатні переміщатися усередині
геному живих організмів. Існує декілька класів мобільних елементів геному,
що відрізняються за будовою і способом переміщення:
транспозони – генетичні паразити і віруси або схожі на них
послідовності. Можуть переміщатися в межах геному однієї клітини за
допомогою процесу транспозиції забезпечують переміщення невеликих
ділянок генетичного матеріалу (транспозонів) у межах однієї хромосоми чи
між різними хромосомами. в результаті чого можуть викликати мутації і
змінювати кількість ДНК в геномі. Транспозони також відомі під назвою
«стрибаючі гени» й іноді виступають в якості гомологічних ділянок при
гомологічній рекомбінації;
інсерції – короткі ділянки ДНК, що кодують лише білки, залучені в процес
транспозиції (на відміну від транспозонів, що зазвичай несуть допоміжні
гени, наприклад гени резистентності);
профаги — латентна форма помірних бактеріофагів;
плазміди.
25.
Реалізація генетичної інформації.У генах закодована інформація, яка необхідна для синтезу амінокислотної
послідовності білків. Фраза «закодувати» досить часто уживається, щоб
позначити інформацію, яка міститься у генах, і необхідна для певної
структури білка: «гени кодують білки». Гени також кодують нуклеотидну
послідовність різних типів РНК, найважливішими з яких є інформаційні, або
матричні (іРНК, або мРНК), рибосомальні (рРНК) і транспортні (тРНК). За
допомогою цих трьох різновидів РНК відбувається синтез білків, які
здійснюють метаболізм і зумовлюють розвиток ознак.
ДНК є носієм генетичної інформації, записаної у вигляді нуклеотидної
послідовності за допомогою генетичного коду, який складається із
тринуклеотидних послідовностей (триплетів нуклеотидів або кодонів).
М.Гамов ще в 1954 р. припустив, що кодування інформації в ДНК може
здійснюватися сполученням декількох нуклеотидів. Одне з визначних
досягнень біології XX століття - розшифрування триплетного генетичного
коду. Антикодон складається з п'яти нуклеотидів, у центрі - три
специфічних рибонуклеотиди (триплет). Азотисті основи антикодона мають
комплементарний триплет (кодон) на ланцюгу іРНК. У період синтезу білка
антикодон знаходить відповідний йому кодон на іРНК і тимчасово
приєднується до нього водневими зв'язками.
26.
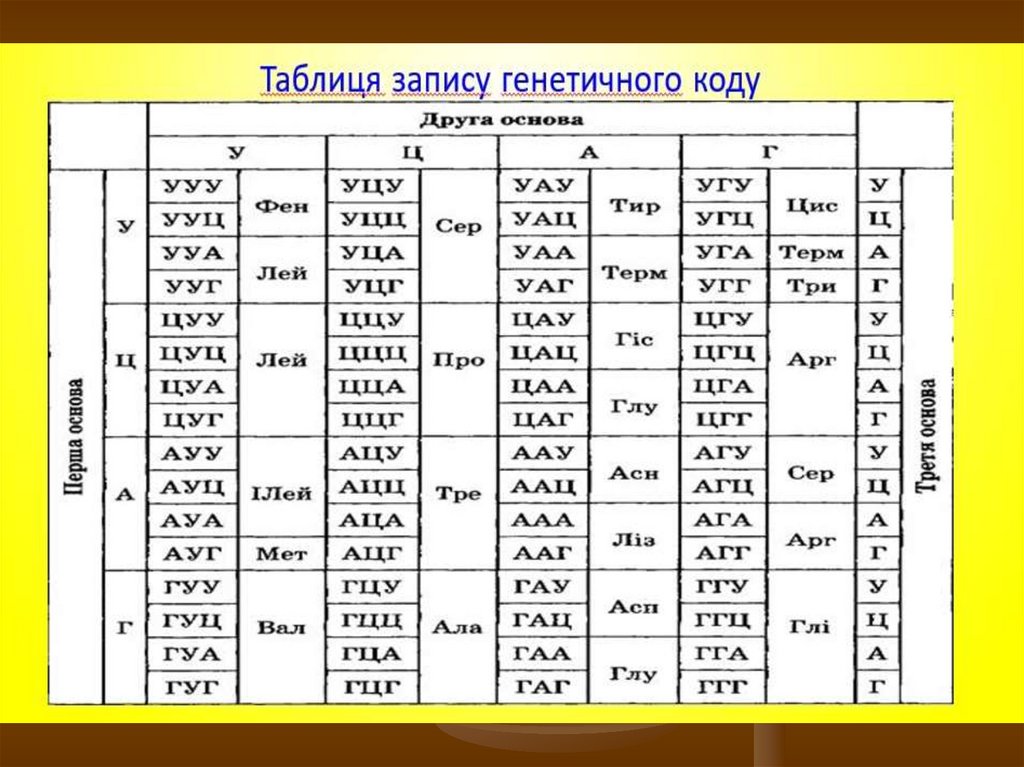
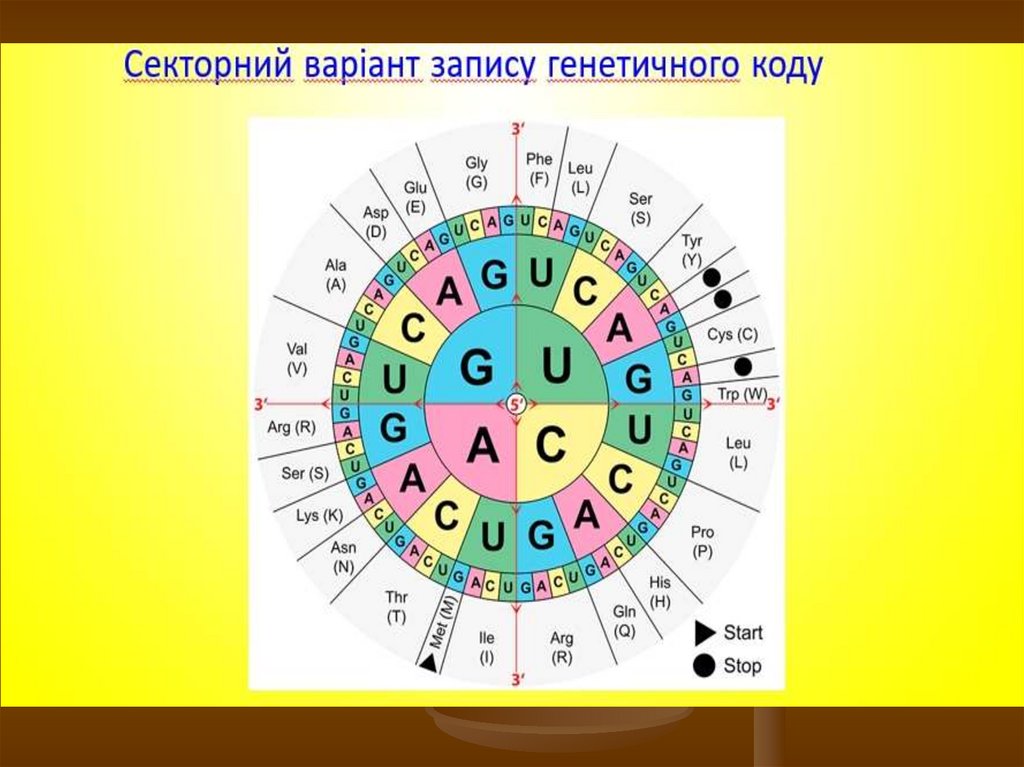
Таким чином, генетичний код - є системою кодуванняпослідовності амінокислот у молекулі білка у вигляді певної
послідовності триплетів нуклеотидів у молекулі ДНК і РНК.
Чотири азотистих основи в комбінаціях по 3, тобто 43,
можуть утворити 64 різних кодони. Серед 64 кодонів три є
сигналами зупинки синтезу білка (стоп-кодони, або
нонсенс-кодони), решта (61 змістовний кодон) відповідають
20 амінокислотам.
Оскільки можливих варіантів кодонів 64, амінокислот - 20,
із яких 18 амінокислот кодуються кількома синонімічними
триплетами (кодонами-синонімами) і тільки дві
амінокислоти – триптофан і метіонін – кодуються лише
одним кодоном кожна.
27.
У багатьох випадках та сама амінокислотакодується не одним триплетом, а кількома –
двома, чотирма, шістьма. У цих випадках
перші два нуклеотиди таких триплетів
однакові, а третій змінюється. Із 64
триплетів 61 є значущим, тобто кодуючим
амінокислоти. Два значущих кодони у складі
інформаційної РНК – AУГ і ГУГ (лише у певних
організмів планети) – називають ініціюючими
чи кодонамиініціаторами, бо саме з них
рибосома розпочинає синтез генного
продукту, тобто поліпептиду.
28.
Три триплети у складі інформаційної РНК– УАА, УАГ і УГА, – називають нонсенскодонами або беззмістовними чи
стоп-кодонами, бо вони не кодують
амінокислот. Ці кодони визначають
закінчення синтезу поліпептидного
ланцюга, отже несуть дуже важливе
смислове навантаження. Тому більш
коректно їх називати термінуючими
триплетами.
29.
Генетичний код ДНК має такі фундаментальні характеристики:1) триплетність (триплет нуклеотидів – це три сусідні азотисті основи,які
кодують одну амінокислоту);
2) специфічність (кожен окремий триплет кодує тільки одну певну
амінокислоту);
3) неперекривність (жодна азотиста основа одного кодону ніколи не
входить до складу іншого кодону);
4) відсутність розділових знаків (генетичний код не має "знаків
пунктуації" між кодуючими триплетами у структурних генах);
5) універсальність (генетичний код однаковий для всіх живих організмів,
від бактерій до ссавців, тобто у всіх живих організмів той самий триплет
кодує ту ж амінокислоту);
6) надмірність або виродженість (деякі амінокислоти часто кодуються
більш ніж одним кодовим триплетом);
7) колінеарність (ДНК є лінійним полінуклеотидом, а білок – лінійним
поліпептидом. Послідовність амінокислот у білку відповідає послідовності
триплетів у його гені. Тому ген і поліпептид, який він кодує, називають
колінеарними);
8) відповідність (гени – поліпептиди).
30.
31.
Під дією мутагену в гені змінився склад кількохтриплетів, але клітина продовжує синтезувати
той самий білок. З якою властивістю
генетичного коду це може бути пов'язано?
Специфічністю
Універсальністю
Триплетністю
+ Виродженістю
– Колінеарністю
32.
Молекула РНК має простішу будову. Вона складається зодного полінуклеотидного ланцюга, який теж містить багато
нуклеотидів чотирьох видів — гуаніловий, цитидиловий,
аденіловий та уридиловий (у молекулі РНК замість основи
тиміну міститься азотиста основа урацил, яка за структурою
близька до тиміну). До нуклеотидів РНК входить не
дезоксирибоза, а рибоза. Молекули РНК значно коротші, ніж
молекули ДНК, і мають набагато меншу молекулярну масу,
яка рідко перевищує 100 тис. у. о. Існує кілька видів РНК, що
різняться за структурою і функціями. Так, рибосомальні РНК
(р-РНК) містяться в рибосомах, транспортні РНК (тРНК),
найбільші за розміром, беруть участь у транспортуванні
амінокислот до місця синтезу білків, інформаційні, або
матричні РНК (і-РНК, або м-РНК), синтезуються на ділянці
одного з ланцюгів ДНК і передають інформацію про структуру
білка з ядра клітини до рибосом.
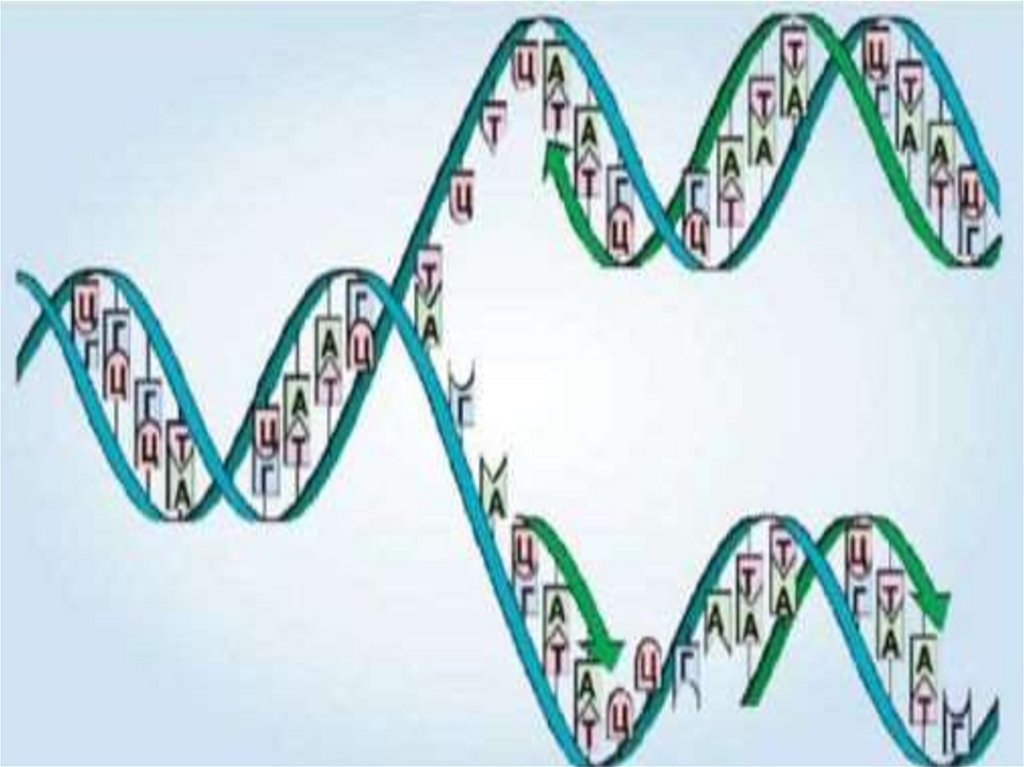
33.
Всі ці типи РНК синтезуються на матриці ДНК(тобто за рахунок копіювання послідовності
ДНК у послідовність макромолекули, що
синтезується) у процесі транскрипції і беруть
участь у біосинтезі білків (процесах сплайсингу
і трансляції).
Генетична інформація, закодована в ДНК,
зчитується при експресії генів. У більшості
випадків вона використовується для
біосинтезу білків у процесах транскрипції
(синтезу молекул РНК на матриці ДНК) і
трансляції (синтезу білків на матриці РНК).
34.
35. ОСНОВНІ ПРАВИЛА, ЩО ДОПОМАГАЮТЬ РОЗВ’ЯЗУВАТИ ЗАДАЧІ
молекула білка у середньому складається з 200 амінокислот.
кожну амінокислоту кодує триплет нуклеотидів і-рнк (під час
трансляції)
трансляція здійснюється згідно з генетичним кодом
для всіх ДНК дійсним є правило Чаргаффа: А=Т
ГΞЦ
вміст пуринових азотистих основ А і Г = вмісту піримідинових
азотистих основ – Т і Ц
сума всіх нуклеотидів в молекулі ДНК або рнк становить 100 %:
для днк = А+Т+Г+Ц
для рнк = А+У+Г+Ц
довжина одного нуклеотида, тобто відстань між двома сусідніми
вздовж осі днк, становить 0,34 нм
один виток спіралі включає 10 нуклеотидів і має довжину 3,4 нм
36. ОСНОВНІ ПРАВИЛА, ЩО ДОПОМАГАЮТЬ РОЗВ’ЯЗУВАТИ ЗАДАЧІ
середня молекулярна маса одного нуклеотида 345 умовниходиниць
середня молекулярна маса однієї амінокислоти дорівнює
100 умовних одиниць
для визначення довжини гена вираховують кількість
нуклеотидів, що знаходяться в одному ланцюзі Днк
для визначення молекулярної маси гена знаходять кількість
нуклеотидів у двох ланцюгах ДНК
триплети УГА, УАА, УАГ – є кодонами термінаторами, не
несуть спадкової інформації, вони сигналізують про
закінчення синтезу.
кодон АУГ – є стартовим, з нього починається синтез.
37.
Життєвий цикл клітини включає процесавторепликації ДНК. Внаслідок цього
монохроматидні хромосоми
перетворюються на біхроматидні. У який
період клітинного циклу потрапляє це
явище?
A. G0
B. G1
C. * S
D. G2
E. -
38.
Функції нуклеїнових кислот — збереження,відтворення в точних копіях і передача
генетичної інформації в ряді клітинних поколінь.
ДНК розташована переважно в ядрі і містить код
(інформацію) на синтез білка — порядок
розміщення амінокислот у поліпептидному
ланцюгу, а також регулює процеси транскрипції
— “переписування” інформації з ДНК на
інформаційну РНК. Розрізняють три види РНК:
інформаційну (іРНК), або матричну (мРНК), яка
служить матрицею для синтезу білка,
транспортну (тРНК), що використовується для
перенесення амінокислот до рибосом і
рибосомальну (рРНК). Остання належить до
складу рибосом, на яких безпосередньо
відбувається збирання поліпептидного ланцюга
39.
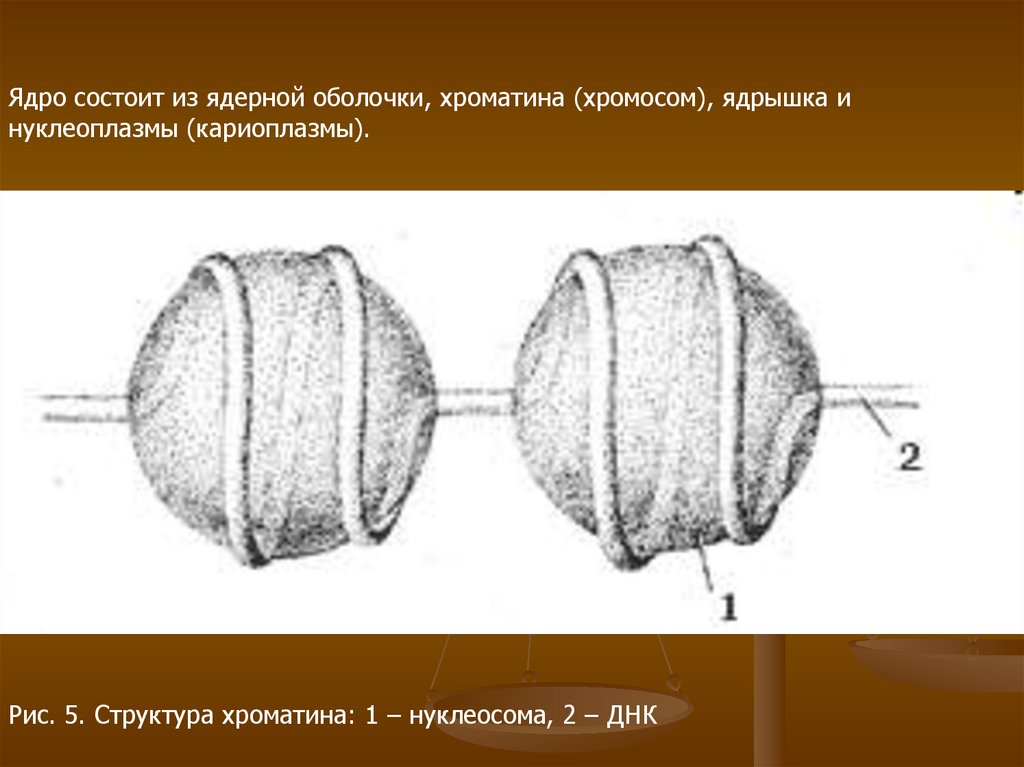
Ядро состоит из ядерной оболочки, хроматина (хромосом), ядрышка инуклеоплазмы (кариоплазмы).
Рис. 5. Структура хроматина: 1 – нуклеосома, 2 – ДНК
40.
Количество молекул ДНК в ядре равно числухромосом. Количество и форма хромосом являются
уникальной характеристикой вида. В состав каждой
из хромосом входит одна молекула ДНК, состоящая
из двух связанных между собой нитей и имеющая
вид двойной спирали толщиной 2 нм. Длина ее
значительно превышает диаметр клетки: она
может достигать нескольких сантиметров.
Молекула ДНК заряжена отрицательно, поэтому
сворачиваться (конденсироваться) она может
только после связывания с положительно
заряженными белками-гистонами
41.
Сначала двойная нить ДНК закручивается вокруг отдельныхблоков гистонов, в каждый из которых входит 8 молекул
белка, образуя структуру в виде «бусин на нитке» толщиной
около 10 нм. Бусины называются нуклеосомами. В результате
формирования нуклеосом длина молекулы ДНК уменьшается
примерно в 7 раз. Далее нить с нуклеосомами сворачивается,
формируя структуру в виде каната толщиной около 30 нм.
Затем такой канат, изогнутый в виде петель, прикрепляется к
белкам, образующим основу хромосомы. В результате
образуется структура с толщиной около 300 нм. Дальнейшая
конденсация этой структуры приводит к образованию
хромосомы.
В период между делениями хромосома частично
разворачивается. В результате этого отдельные участки
молекулы ДНК, которые должны экспрессироваться в данной
клетке, освобождаются от белков и вытягиваются, что делает
возможным считывание с них информации путем синтеза
42.
Ядерная оболочка состоит из двух мембран: внешней и внутренней, разделенныхмежмембранным пространством. Внешняя мембрана соприкасается с цитоплазмой,
на ней могут находиться полирибосомы, а сама она может переходить в мембраны
эндоплазматического ретикулума. Внутренняя мембрана связана с хроматином.
Таким образом, ядерная оболочка обеспечивает фиксацию хромосомного
материала в трехмерном пространстве ядра.
Оболочка ядра имеет круглые отверстия – ядерные поры (рис. 7). В области поры
внешняя и внутренняя мембраны смыкаются и образуют отверстия, заполненные
фибриллами и гранулами. Внутри поры располагается сложная система из белков,
обеспечивающих избирательное связывание и перенос макромолекул. Количество
ядерных пор зависит от интенсивности метаболизма клетки.
Поры в ядерной мембране
43.
Хромосома — це велика молекулярна структура, де міститься близько 90% ДНКклітини. Всі хромосоми містять дуже довгий безперервний полімеризований ланцюг
ДНК (єдину ДНК-молекулу), що містить гени, регуляторні елементи та проміжні
нуклеотидні послідовності.
Слово «хромосома» походить від грецьких слів «хрома» — колір та «сома» — тіло.
Хромосоми еукаріот складаються з лінійної макромолекули ДНК, що намотана на
специфічні білки-гістони, формуючи матеріал під назвою «хроматин».
В клітинах прокаріот звичайно міститься єдина хромосома, яка, на відміну від
еукаріот, є кільцевою та позбавленою гістонів. Втім, це правило не є абсолютним:
існують бактерії з більше, ніж одною хромосомою; у деяких бактерій хромосоми є
лінійними; у кількох видів архей виявлені специфічні гістони.
Хромосоми можуть перебувати в двох структурно-функціональних станах: в
конденсованому (спіралізованому) та деконденсованому (деспіралезованому). В
інтерфазі хромосоми живої клітини невидимі, спостерігати можна лише гранули
хроматину, бо в цей період хромосоми частково або повністю деконденсовані. Це є
їхнім робочим станом, бо в більш дифузному хроматині активніші процеси синтезу.
Під час мітотичного поділу клітини, коли відбувається конденсація
хроматину,хромосоми добре помітні.
44.
45.
ХроматинХроматином називають комплекс ДНК та білків. До складу хроматину входять два
типи білків - гістонові та негістонові.
ДНП
Найменшими структурними компонентами хромосом є нуклеопотеідні фібрили, їх
видно винятково в електронний мікроскоп. Хромосомні нуклеопротеіди — ДНП
(дезоксірібонуклеопротеїди) - складаються з ДНК і білків (переважно гістонів).
Нуклеосоми, хромонеми, хроматиди
Молекули гістонів утворюють групи - нуклеосоми. Кожна нуклеосома містить в собі 8
білкових молекул. Розмір нуклеосоми приблизно 8 нм. З кожною нуклеосомою
пов"язана ділянка ДНК, що спірально оплетає нуклеосому ззовні. В такій ділянці ДНК
знаходиться 140 нуклеотидів загальною довжиною 50 нм, але завдяки спіралізаціі
довжина скорочується в 5 раз.
В хроматині близько 87 - 90% довжини ДНК зв"язано з нуклеосомами.
Фібрили ДНП попарно закручуються, утворюючи хромонеми (від гр. chroma - колір,
nema - струна), які входять до комплексів вищого порядку - також спірально
закручених полухроматид. Пара полухроматид утворює хроматиду, а пара хроматид
-хромосому.
На різних ділянках однієї хромосоми спіралізація, компактність її основних елементів
неоднакова; із цим пов"язана різна інтенсивність окращування окремих ділянок.
46.
ЦентромераПервинна перетяжка, або центромера, - найменш спіралізована частина
хромосоми.
На ній розміщений кінетохор (гр. kinesis - рух, phoros - той, що несе), до
якогго під час поділу клітини кріпляться ниті веретена поділу.
Місце розташування центромери в кожної пари хромосом постійне, воно
обумовлює їхню форму.
В залежності від розташування центромери виділяють три типи
Метацентричні,
субметацентричні і
акроцентричні.Метацентричні хромосоми мають плечі
хромосом:
майже однакової довжини; в субметацентричних плечі нерівні;
акроцентричні хромосоми мають палочковидну форму з дуже коротким,
майже непомітним другим плечем.
Можуть виникати і телоцентричні хромосоми - як результат відриву
одного плеча, коли центромера розташована на кінці хромосоми. В
нормальному каріотипі такі хромосоми не зустрічаються.
47. Tипи хромосом в залежності від місцерозташування центромери
48.
Хромосомний фрагмент жінки міститьхромосому з плечами р і q однакової
довжини. До якого морфологічного
типу належить ця хромосома?
* Метацентричний
Телоцентричний
Акроцентричний
Субметацентричний
Субакроцентричний
49.
Chromosomal complement of a womancontains a chromosome with arms p and q
of equal length. What morphological type
does this chromosome belong to?
*Metacentric
Telocentric
Acrocentric
Submetacentric
Subacrocentric
50.
ТеломериКінці плечей хромосоми називаюиь теломерами, це
спеціалізовані ділянки, які перешкоджають з"єднанню
хромосом між собою або з їхніми фрагментами. Кінець
хромосоми, який не має теломери, стає "ненасиченим",
"липким", і легко приєднує фрагменти хромосом або
з"єднується з подібними ділянками. У нормі ж теломери
зберігають хромосому як дискретну індивідуальну одиницю.
Супутники
Деякі хромосоми мають глибокі вторинні перетяжки, що
відділяють окремі ділянки хромосоми - супутники. Такі
хромосоми можуть зближуваьись і утворювати асоціації, а
тонкі ниті, які з"єднують супутники з плечами хромосом,
при цьому беруть участь в утворенні ядерець. Саме ці
ділянки в хромосомах людини є організаторами ядерець. У
людини вторинні перетяжки є на довгому плечі 1, 9 та 16
хромосом та на кінцевих ділянках коротких плечей 13, 14,
15, 21, 22 хромосом.
51.
ПРАВИЛО ПОСТІЙНОСТІ ЧИСЛА ХРОМОСОМКількість хромосом та характерні особливості їхньої будови видова ознака. Це є правилом постійності числа хромосом. Це
число не залежить від висоти організації і не завжди вказує на
філогенетичну спорідненість. Наприклад, в ядрах усіх клітин
конячої аскариди Paraascaris megalocephala univalenus
знаходиться по 2 хромосоми, в мухи-дрозофіли Drosophila
melanogaster -по 8, в людини - по 46, а в річкового рака Astacus
fluviatalis - по 116.
Кількість хромосом не залежить від висоти організації, а також
не завжди вказує на філогенетичне рідство: одне й те саме
число може траплятись у дуже далеких форм, а в близьких
видів - дуже відрізнятися. Однак, дуже важливо, що в
представників одного виду число хромосом в ядрах всіх клітин
постійне.
ПРАВИЛО ПАРНОСТІ ХРОМОСОМ
Існує також правило парності хромосом, за яким число
хромосом завжди є парним, бо в каріотипі хромосоми
об'єднуються в гомологічні пари.
52.
ПРАВИЛО ІНДИВІДУАЛЬНОСТІ ХРОМОСОМГомологічні хромосоми однакові за формою та
будовою, розташуванням центромер, хромомер,
інших деталей будови. Негомологічні хромосоми
завжди мають відмінності. Тому маємо правило
індивідуальності хромосом: кожна пара гомологічних
хромосом характеризується своїми особливостями.
ПРАВИЛО НЕПРЕРИВНОСТІ ХРОМОСОМ
Правило непреривності хромосом: в послідовних
генераціях число та індивідуальність хромосом
зберігається завдяки здатності хромосом до
авторепродукції під час поділу клітини.
53.
Набори хромосомДиплоїдний
В ядрах соматичних клітин (тобто клітин тіла) міститься повний подвійний набір хромосом.
В ньому кожна хросома має свого гомологічного "партнера". Такий набір хромосом
називають диплоїдним і позначають "2n"
Гаплоїдний
В ядрах гамет (статевих клітин) на відміну від соматичних, є лише по одній хромосомі з
кожної гомологічної пари.Всі вони різні. негомологічні. Такий одинарний набор хромосом
називають гаплоїдним і позначають як "n".
При заплідненні відбувається злиття гамет , кожна з яких вносить в зиготу гаплоїдний
набір хромосом, і відновлюється диплоїдний набір: n + n = 2n.
В деяких організмів може внаслідок мутацій змінюватись диплоїдний набір хромосом поліплоїдія або анеуплоїдія.
Гетерохромосоми й аутосоми
При порівнянні хромосомних наборів чоловічих та жіночих осіб одного виду
спостерігається відмінність в одній парі хромосом. Ця пара отримала назву статевих
хромосом, або гетерохромосом.
Решта пар гомологічних пар хромосом, однакових в обох статей, мають загальну назву
аутосоми.
54.
Хромосомні аберації ( від лат. aberratіo — ухилення), абоперебудови — зміни структури хромосом, викликані дією на
клітини мутагенних факторів (іонізуючого випромінювання,
канцерогенів тощо). Розрізняють декілька різновидів
хромосомних аберацій.
Делеція (від лат. deletio — нестача) — випадання частини
хромосоми при її розривах з втратою відірваного сегмента.
Розрізняють кінцеву делецію — відрив кінцевої ділянки з
вкороченням хромосоми та інтерстиціальну делецію— втрату
внутрішнього фрагмента хромосоми.
Дуплікація (від лат. duplex — подвійний) — подвоєння
певного сегмента в хромосомі. Якщо подвоєння ділянки в
хромосомі відбулося в результаті переміщення його з іншої
хромосоми (частіше гомологічної), то таку дуплікацію
називають переміщеною. Коли дуплікований сегмент
знаходиться в хромосомі поряд з вихідним, то це дуплікація
повторень.
55.
Транслокації (від лат. trans — через і locus — місце) —взаємообмін відділеними уламками (фрагментами) між
гомологічними і негомологічними хромосомами в процесі
кросинговеру, як статевого (при мейозі), так і
соматичного. Такий взаємообмін можливий при двох
одночасних розривах в різних хромосомах. Якщо при
транслокації зіллються два сегменти, які містять
хромомери, то виникне дицентрична хромосома, а
з’єднання двох сегментів без центромер дає ацентричну
хромосому.
Інверсія (від лат. inversio — перевертання,
переставлення) відбувається внаслідок перевертання
уламка хромосоми на 180 градусів після її розриву в двох
місцях з наступним з’єднанням двох кінцевих і
переверненого серединного сегментів. В інвертованому
сегменті порядок розміщення генів міняється на
обернений.
56.
Унаслідок дії випромінювання напослідовність нуклеотидів ДНК загублені
2 нуклеотиди. Яка з перелічених видів
мутацій відбулася в ланцюзі ДНК?
– Інверсія
+ Делеція
– Дуплікація
– Реплікація
57. Біосинтез білка
І етап. Транскрипція —передача інформації про
структуру білка з молекули ДНК на ІРНК. Особливий
фермент РНК-полімераза, просуваючись по молекулі ДНК, за
принципом комплементарності підбирає нуклеотиди і з’єднує
їх в один ланцюг. Ділянка ДНК (ген або група генів) є
матрицею для відповідної іРНК. На початку кожної групи
генів є своєрідний посадочний майданчик для ферменту
РНК-полімерази — промотор. Тільки приєднавшись до неї,
РНК-полімераза здатна почати синтез іРНК. У кінці групи
генів РНК-полімераза зустрічає стоп-сигнал — термінатор (у
вигляді певної послідовності нуклеотидів), який сигналізує
про припинення процесу транскрипції. Синтезовані молекули
іРНК переходять із ядра в цитоплазму, а ДНК відновлює
свою структуру.
58. ТРАНСКРИПЦІЯ
Транскрипція - це синтез іРНК. Уеукаріот транскрипція має свої
особливості. Ген еукаріот
складається з екзонів та інтронів.
Інтрони не кодують білок. Вони
вирізаються з іРНК. Таким чином,
транскрипція у еукаріот включає два
етапи: 1. Синтез про-іРНК (незрілої
іРНК), яка повністю комплементарна
гену.
59.
2. Процесинг-дозрівання іРНК.Процесинг включає: * сплайсинг
(вирізання інтронів і зшивання
екзонів), *утворення кепу та полі-Ахвоста. Кеп (метильований гуанін)
прикріплюється до початкового кінця
іРНК, полі-А-хвіст(велика кількість Ануклеотидів) прикріплюється до кінця
іРНК. Кеп і хвіст забезпечують
стабільність іРНК в цитоплазмі.
60.
II етап. Активація амінокислот.Цей процес відбувається в цитоплазмі.
Активовані молекули амінокислот з’єднуються
з відповідними молекулами транспортних РНК.
У молекулі тРНК є дві важливі ділянки:
акцепторна ділянка, до якої прикріплюється
відповідна амінокислота, антикодон — триплет
нуклеотидів, який комплементарний кодону
ІРНК даної амінокислоти. Активовані
амінокислоти, сполучені з тРНК, надходять до
рибосом.
61.
Активація амінокислот і з'єднання з тРНК.У клітинах еукаріот близько 50 видів тРНК (у
зв'язку з надмірністю генетичного коду). Кожна
тРНК має антикодон (для взаємодії з кодоном
іРНК) і акцепторну ділянку (куди приєднується
амінокислота). З'єднання тРНК з амінокислотою
каталізує фермент аміноацил-тРНК-синтетаза.
Процесу передує активація амінокислот
(з'єднання із залишком АТФАМФ). Амінокислота
+ АТФ = Амінокислота + АМФ (АК + АМФ). АК
+ АМФ + тРНК = АК + тРНК + АМФ.
62.
III етап. Трансляція — синтез поліпептиднихланцюгів. Відбувається так: молекула іРНК рухається
між двома субодиницями рибосом, і до неї послідовно
приєднуються молекули тРНК з амінокислотами. При
цьому за принципом комплементарності кодони ІРНК
вступають у зв’язок з антикодонами тРНК.
Послідовність розташування амінокислот при цьому
визначається порядком чергування триплетів у
молекулі ІРНК. Про завершення синтезу
поліпептидного ланцюга сигналізує термінуючий кодон
іРНК (УАА, УАГ, УГА). Процес синтезу молекули білка
потребує великих витрат енергії. На сполучення кожної
амінокислоти з тРНК витрачається енергія двох
молекул АТФ. Крім того, енергія ще двох молекул АТФ
потрібна для пересування рибосоми по ІРНК.
63. ТРАНСЛЯЦІЯ
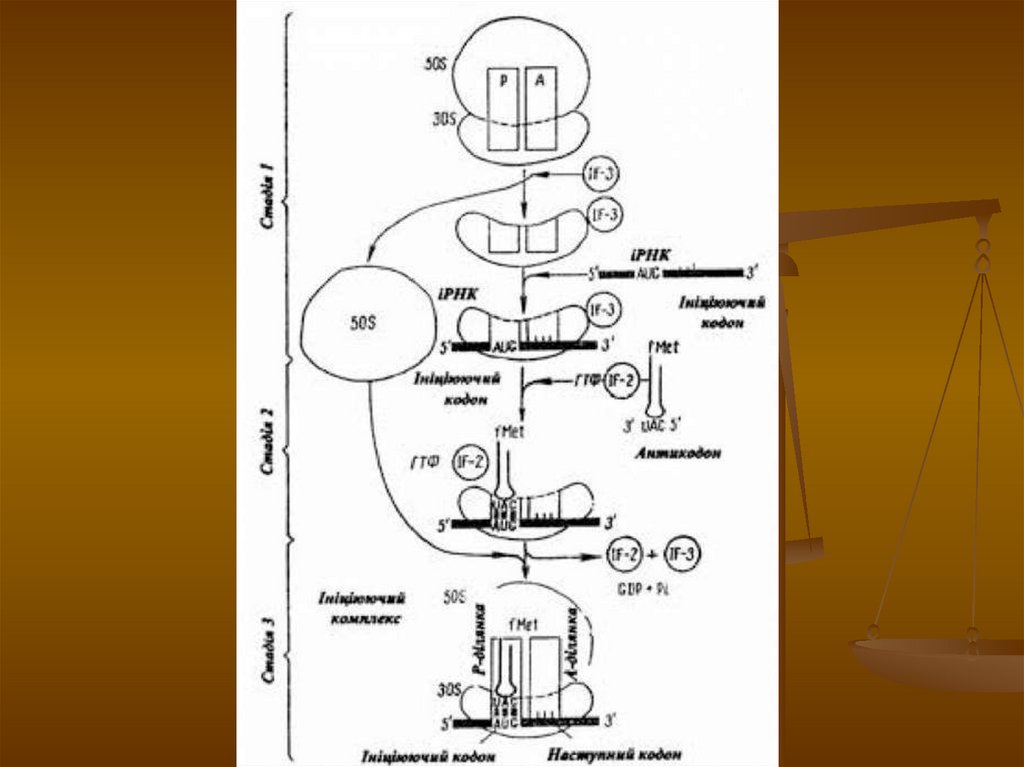
Трансляція - синтез первинної структурибілка в рибосомі. Етапи трансляції:
1) Ініціація - початок трансляції. Рибосома
з'єднується з іРНК і захоплює два кодони
(перший - ініціальний-опиняється в
пептидильному центрі). До ініціального
триплету підходить тРНК з ініціальний
метіоніном. Утворюється ініціальний комплексрибосоми, ініціальний триплет, тРНК.
2) Елонгація - синтез поліпептиду. До
другого кодону іРНК підходить друга тРНК з
амінокислотою.
64.
Якщо антикодон тРНК комплементарнийкодону іРНК, дві амінокислоти з'єднуються
пептидним зв'язком. Потім перша тРНК
виходить з рибосоми, рибосома переміщується
на один триплет уперед. До цього триплета
підходить нова тРНК з амінокислотою. Якщо
антикодон тРНК комплементарний кодону
іРНК, то між двома останніми амінокислотами
знову утворюється пептидний зв'язок і процес
повторюється. Процес продовжується до тих
пір, поки рибосома не дійде до стоп-кодону.
65.
3) Термінація транскрипції - закінчення.Рибосома доходить до стоп-кодону. Синтез
поліпептиду зупиняється.
4) Посттрансляційні процеси - утворення
вторинної, третинної та четвертинної
структури білка, модифікація амінокислот.
Процес може відбуватися в цитоплазмі,
гранулярній ЕПС, комплексі Гольджі. Після
того, як білок утворив третинну або
четвертинну структуру, він може виконувати
свої функції.
66.
Під час трансляції до кожної іРНКприєднується одночасно кілька рибосом, які
розміщені вздовж її молекули на певній
відстані одна від одної. Як називається
трансляційний комплекс, що складається з
однієї іРНК та розміщених на ній кількох
рибосом?
Центросома
Лізосома
Фагосома
Нуклеосома
+ Полісома
67.
Етапи трансляціїАктивація амінокислот
Ініціація пептидного
ланцюга
Елонгація
Термінація
Необхідні компоненти
20 амінокислот, 20 амінозцил-тРНК-синтетаз. 20
або більше тРНК, АТФ. Mg2+
іРНК, N-формідметіоніп тРНК, що ініціює кодон
іРИК (AUG). ЗОS-рибосомна субчастина. 50S
рибосомна субчастина, ГТФ. Мg2+фактори
ініціації (іF=1, iF=2, iF=3)
70S· рибосома (ініціюючий комплекс), набір
аміноацил-тРНК, відповідних кадонам іРНК,
Мg2+, фактори елонгації (Тu, Тв, і G). ГТФ,
пептидилтрамсфераза
АТФ, термінуючий кодом іРНК. фактори
звільненя поліпептида (R1, R2, S)
Згортання і процесинг
Специфічні ферменти і кофактори
68.
Ініціація поліпептидного ланцюга. Було встановлено, що вЕ.соlі й інших прокаріот N-кінцевою амінокислотою при
збірці поліпептидного ланцюга завжди є залишок Nформілметіоніну. Цей та інший факти стали підставою для
припущення про значення формільованого метіоніну як
ініціатора в процесі синтезу поліпептидного ланцюга.
Формільований метіонін одержується в результаті двох
послідовних реакцій. При цьому варто вказати на існування
двох тРНК: тРНКМеt і тРНКfМеt, що здійснюють акцепцію
метіоніну, а також звернути увагу на те, що фермент
формілтрансфераза (трансформілаза) не спроможний
формілювати метіонін, що знаходиться у вільному стані.
Однак і в комплексі з тРНК не завжди можливе
формілювання залишку метіоніну. Ця реакція можлива
тільки в тому випадку, коли тРНК виявляється специфічною
тРНКfМеt
69.
70.
Упаковка і процесинг поліпептидноголанцюга.
Первинна структура (послідовність амінокислот) є
визначальним моментом у формуванні тривимірної
структури, завдяки якій білок стає функціонально активним.
Однак нерідко білкова молекула набуває біологічно
активної конформації тільки в результаті процесингу чи
ковалентної (посттрансляційної) модифікації, що проходить
у різних білків по-різному і включає реакції видалення чи
приєднання атомних груп. Так, формільна група, що
входить до складу N-кінцевого N-формілметіоніну, у білків
бактеріального походження в ході процесингу
деформілюється. Під дією особливих амінопептидаз
відбувається гідролітичне відщеплення одного чи декількох
N-кінцевих амінокислотних залишків, у зв’язку з чим у
багатьох остаточно сформованих білках їх не вдається
знайти.
71.
Синтез одного білка триває від 20 до 560 секунд. Алеця швидкість буде підвищена, якщо синтез
поліпептидного ланцюга відбудеться на
полірибосомальному комплексі (полісомі) — скупченні
рибосом (до 80 й більше), коли вони об’єднані однією
ІРНК в групу.
IV етап. Утворення вторинної і третинної
структур білкової молекули. Цей етап здійснюється
в цитоплазмі шляхом скручування, згортання
поліпептидного ланцюга. Потім до нього приєднуються
різні органічні молекули — вуглеводи, жирні кислоти
тощо.
72.
КЛІТИННИЙ ЦИКЛ (син.: життєвий цикл) існування клітини від поділу до поділу абосмерті. У клітинах, які здатні до розмноження,
клітинний цикл складається з чотирьох
періодів (три перших включають інтерфазу),
які послідовно змінюють один одного.
Перший період інтерфази - пресинтетичний,
або постмітотичний (G1, від англ. grow рости, збільшуватись), другий - синтетичний
(S, від англ. synthesis - синтез), третій постсинтетичний, або премітотичний (G2).
За інтерфазою наступає четвертий період
циклу - мітоз (M).
73.
У клітину введено хімічну речовину, що блокуєроботу ферментів, які беруть участь у
деспіралізації ДНК. Які процеси й у який
період мітотичного циклу клітини
порушуються?
Реплікація ДНК у метафазі
Порушується деспіралізація хромосом і
формування ядерної оболонки в телофазі
Поділ ділянки центромери на окремі
хроматиди в анафазі
+ Реплікація ДНК у синтетичному
періоді
Дочірні хромосоми не досягають полюсів
клітини в анафазі
74.
A chemical is introduced into the cell that blocksthe work of enzymes involved in DNA desirability.
What processes and during which period of the
mitotic cell cycle are disrupted?
DNA replication in metaphase
Chromosome despiralization and nuclear
membrane formation in telophase are impaired
Separation of the centromere site into
individual chromatids in anaphase
+ DNA replication in the synthetic period
The daughter chromosomes do not reach the
cell poles in anaphase
75. РЕГУЛЯЦІЯ ЕКСПРЕСІЇ ГЕНІВ У ПРОКАРІОТІВ. ОПЕРОН.
У прокаріот кільцевидна ДНК, що кодує невеликукількість білків (у кишкової палички- біля 4000). Для
багатьох генів характерна оперонна регуляція
активності. Оперон - це група структурних генів, які
кодують білки-ферменти одного метаболічного циклу
та робота яких знаходиться під контролем загальних
регуляторних генів. Оперони дозволяють ДНК кодувати
багато білків. Оперон був відкритий в 1961 р.
французькими вченими Жакобом і Моно. Вони відкрили
лактозний оперон у кишкової палички. Якщо кишкову
паличку помістити в середовище, що містить лактозу,
то вона починає виробляти три ферменти, що беруть
участь у метаболізмі лактози.
76.
Ферменти кодують три структурні гена:lacZ - галактозидаза - розщеплює лактозу на
глюкозу та галактозу.
Lac Y-фермент пермеаза (забезпечує
надходження лактози в клітину).
lacA - трансацетілаза, бере участь у видаленні
з клітини токсичних продуктів розщеплення
лактози.
77.
Структурні гени знаходяться в оточеннірегуляторних генів:
Ген-регулятор - кодує білок-репрессор.
Ген-промотор - місце приєднання РНК-полімерази
для початку транскрипції.
Ген-оператор. Якщо до нього приєднаний білокрепрессор,
то
він
блокує
транскрипцію.
Термінатор - на ньому закінчується транскрипція.
Оперон інактивований, якщо білок-репрессор
з'єднаний
з
геном-оператором.
Оперон
в
активному стані, якщо в клітку потрапляє лактоза.
Вона з'єднується з білком-репрессором та
інактивує його. Починається синтез трьох
ферментів.
78. РЕПАРАЦІЯ ДНК.
Репарація ДНК – виправлення помилок ДНК.Якщо помилки залишаються, то вони можуть
призвести до генних мутацій і генних хвороб.
Репарація підтримує генетичну цілісність
організму та його виживання.
1) Фоторепарація у прокаріот. Опромінення
клітини ультрафіолетовими променями
викликає утворення в ДНК тимінових димерів.
УФ- промені активують фермент
фотореактивації, який зв'язується з тимінових
димерами і розриває їх.
79.
2) Ексцизійна репарація в прокаріот і еукаріот- ферменти-нуклеази вирізають помилкову
основу або ділянку пошкодженого ланцюга
ДНК, фермент ДНК-полімераза 1 типу
вбудовує нормальні нуклеотиди, ферменти
лігази зшивають фрагменти.
3) Репарація під час реплікації - самокорекція
ДНК .
80.
4) Постреплікативна репарація - якщо невидалені помилкові нуклеотиди під час
реплікації, то відбувається рекомбінація
пошкодженого ланцюга з ланцюгом ДНК у
другій дочірній молекулі та помилка
усувається.
5) SOS-репарація - при реплікації ДНКполімераза перескакує місце пошкодження та
продовжує реплікацію без розривів, але
послідовність нуклеотидів змінюється.
81.
ХВОРОБИ РЕПАРАЦІЇ ДНК.При порушенні репарації ДНК у клітинах
накопичуються мутації, що з часом
призводить до:
1) розвитку пухлин,
2) передчасного старіння,
3) спадкових захворювань – хвороб
репарації.
82.
Спадкові хвороби, які обумовлені мутацієюгенів репарації ДНК, називаються хворобами
репарації ДНК. Приклад - пігментна
ксеродерма - генна хвороба з аутосомнорецесивним типом успадкування. У хворих
порушена ексцизійна репарація ДНК, які
пошкоджені УФ променями та ін. мутагенами.
Під дією сонячного світла на шкірі
з'являються ластовиння, пігментні плями, з
часом у всіх хворих розвивається рак шкіри.
83.
МІТОЗЗдатність до поділу - найважливіша властивість
кліток. Без поділу неможливо уявити собі
збільшення числа одноклітинних істот, розвиток
складного багатоклітинного організму з однієї
заплідненої яйцеклітини, поновлення кліток,
тканин і навіть органів, втрачених у процесі
життєдіяльності організму.
Поділ кліток здійснюється поетапно. На кожнім
етапі поділу відбуваються визначені процеси. Вони
приводять до подвоєння генетичного матеріалу
(синтезу ДНК) і його розподілу між дочірніми
клітками. Період життя клітки від одного поділу до
наступного називається клітинним циклом.
84.
Підготовка до поділуЕукаріотичні організми, що складаються з кліток, що мають ядра, починають
підготовку до розподілу на визначеному етапі клітинного циклу, в інтерфазі.
Саме в період інтерфази в клітці відбувається процес біосинтезу білка,
подвоюються всі найважливіші структури клітки. Уздовж вихідної хромосоми з
наявних у клітці хімічних сполук синтезується її точна копія, подвоюється
молекула ДНК. Подвоєна хромосома складається з двох половинок - хроматид.
Кожна з хроматид містить одну молекулу ДНК.
Інтерфаза в клітках рослин і тварин у середньому продовжується 10 - 20 ч.
Потім настає процес розподілу клітки - мітоз.
Під час мітозу клітка проходить ряд послідовних фаз, у результаті яких кожна
дочірня клітка одержує такий же набір хромосом, який був у материнській
льотці.
Мітоз (від греч. mitos- нитка), непрямий розподіл, основний спосіб розподілу
еукаріотних кліток. Біологічне значення мітозу складається в строго однаковому
розподілі редуплікованих хромосом між дочірніми клітками, що забезпечує
утворення генетично рівноцінних кліток і зберігає наступність у ряді клітинних
поколінь. У 1874 И. Д. Чистяков описав ряд стадій (фаз) М. у спорах плаунів, ще
не ясно представляючи собі їхню послідовність. Детальні дослідження з
морфології М. уперше були виконані Е. Страсбургером на рослинах (1876-79) і
В. Флеммінгом на тварин (1882). Тривалість мітозу в середньому 1-2 ч., різна
для різних видів кліток. Процес залежить також і від умов зовнішнього
середовища (температури, світлового режиму й інших показників).
85.
Фази мітозуУ процесі мітозу умовно виділяють декілька стадій, поступово і
безупинно перехідний друг у друга: профазу, прометафазу,
метафазу, анафазу і телофазу. Тривалість стадій мітозу різна і
залежить від типу тканини, фізіологічного стану організму,
зовнішні факторів; найбільш тривалі перша й остання.
У профазі добре видні центріолі - утворення, що знаходяться в
клітинному центрі і грають ролі в розподілі дочірніх хромосом
тварин. (Нагадаємо, що у вищих рослин немає центріолей у
клітинному центрі, що організує розподіл хромосом.) Ми ж
розглянемо мітоз на прикладі тваринної клітки, оскільки
присутність центріолей робить процес розподілу клітки більш
наочним. Центріолі поділяються і розходяться до різних полюсів
клітки. Від центріолів протягаються мікротрубочки, що утворять
нитки веретена розподілу, що регулює розбіжність хромосом до
полюсів клітки, що ділиться
86.
Найважливіші ознаки профази - конденсаціяхромосом, розпад ядерець і початок формування
веретена розподілу, зниження активності
транскрипції (до кінця профази синтез РНК
припиняється). Веретено розподілу утвориться або за
участю центріолів, утворити мітотичний апарат (у
клітках тварин і деяких нижчих рослин), або без них
(у клітках вищих рослин і деяких найпростіших). У
водоростей, нижчих грибів і ряду найпростіших
веретено може формуватися усередині ядра (т.зв.
закритий М.). Прометафаза починається розпадом
ядерної оболонки на фрагменти і безладні рухи
хромосом у центр. частини клітки, що відповідає зоні
колишнього ядра. При "закритому М." оболонка ядра
зберігається протягом усього М. Хромосоми
спірализуються й у результаті цього коротшають і
товщають, і їх уже можна спостерігати у світловий
мікроскоп. Ще краще вони видні на наступній стадії
мітозу - метафазі.
87.
У Метафазі завершується формування веретенарозподілу. Хромосоми перестають рухатися і
вибудовуються по екваторі веретена, утворити
екваторіальну пластинку. При цьому добре видно, що
кожна хромосома, що складається з двох хроматид,
має перетяжку - центроміру (рис 2). Хромосоми
своїми центромірами прикріплюються до ниток
веретена розподілу. Після розподілу центроміри
кожна хроматида стає самостійною дочірньою
хромосомою. Синтез білка знижений на 20-30% у
порівнянні з інтерфазою. На цій стадії М. клітки наиб.
чуттєві до холоду, колхіцину, його похідним і ін.
агентам, вплив яких руйнує веретено поділу і
приводить до зупинення поділу кліток (До-мітоз). При
низьких дозах агентів, що ушкоджують, нормальний
плин М. відновлюється через кілька годин після їх
впливу; більш високі дози приводять або до загибелі
клітки, або до її поліплоїдизації.
88.
Анафаза - сама коротка стадія М. Характеризується поділом сестриниххроматид і розбіжністю хромосом до протилежних полюсівклітки.
Швидкість їхнього руху в середньому 0,2-5 мкм/хв. У ряді випадків рух
хромосом до полюсів клітки супроводжується додатковою розбіжністю
полюсів друг від друга.
Тілофаза триває з моменту припинення руху хромосом до закінчення
процесів, зв'язаних з реконструкцією дочірніх ядер (деспіралізація й
активізація хромосом, утворення ядерної оболонки, формування
ядерець), з руйнуванням веретена розподілу, поділом тіла
материнської клітки на 2 дочірні й утворенням (у клітках тварин)
залишкового тельця Флеммінга. Вона починається після того, як
дочірні хромосоми, що складаються з однієї хроматиди, досягли
полюсів клітки. На цій стадії хромосоми знову деспіралізуются і
здобувають такий же вид, який вони мали до початку розподілу клітки
в інтерфазі (довгі тонкі нитки). Навколо них виникає ядерна оболонка,
а в ядрі формується ядерце, у якому синтезуються рибосоми. У процесі
розподілу цитоплазми всі органоїди (мітохондрії, комплекс Гольджи,
рибосоми й ін.) розподіляються між дочірніми клітками більш-менш
рівномірно.
По завершенні цитотомії клітки вступають в інтерфазу, до-раю
починається G1- періодом наступного клітинного циклу.
89.
90.
У клітину введено хімічну речовину, що блокуєроботу ферментів, які беруть участь у
деспіралізації ДНК. Які процеси й у який період
мітотичного циклу клітини порушуються?
Реплікація ДНК у метафазі
Порушується деспіралізація хромосом і
формування ядерної оболонки в телофазі
Поділ ділянки центромери на окремі
хроматиди в анафазі
+ Реплікація ДНК у синтетичному
періоді
Дочірні хромосоми не досягають полюсів
91.
92.
93. Мейоз, його етапи й фази. Біологічне значення мейозу
І етап – редукційний поділ, або Мейоз І:Профаза І – фаза спіралізації (конденсації) двохроматидних
хромосом. Вона є найтривалішою за часом у мейозі, під час неї
відбувається ряд процесів.
• Спіралізація двохроматидних хромосом. Хромосоми вкорочуються й
ущільнюються та набувають вигляду паличкоподібних структур. Після
цього гомологічні хромосоми зближуються і кон'югують (тісно
прилягають одна до одної по всій довжині, обвиваються,
перехрещуються).
Так утворюються комплекси з 4 хроматид, сполучених між собою в
певних місцях, так звані тетради, або біваленти.
94.
Кон'югація(зближення
і
злиття
ділянок
гомологічних
хромосом)
і кросинговер (обмін певними ділянками між гомологічними хромосомами). У
результаті кросинговеру утворюються нові комбінації спадкового матеріалу. Таким
чином, кросинговер є одним із джерел спадкової мінливості. Через певний час
гомологічні хромосоми починають відходити одна від одної. При цьому стає
помітним, що кожна з них складається з двох хроматид.
• Розходження центріолей до полюсів.
• Зникнення ядерець.
• Розпад ядерної оболонки на фрагменти.
• Формування веретена поділу.
Метафаза І – фаза розташування тетрад на екваторі:
• короткі нитки прикріплюються до центромер лише з одного боку і хромосоми
розташовуються двома лініями;
• на екваторі клітини розташовуються тетради.
Анафаза І – фаза розходження двохроматидних гомологічних хромосом.
• кожна тетрада розділюється на двохроматидні хромосоми;
• нитки веретена поділу скорочуються і розтягують двохроматидні хромосоми до
полюсів. Наприкінці анафази біля кожного з полюсів клітини опиняється гаплоїдний
(половинний) набір хромосом. Розходження хромосом кожної пари є подією
випадковою, що є ще одним джерелом спадкової мінливості.
95.
Телофаза І – фаза деспіралізації двохроматидних хромосом:• утворення двох клітин з гаплоїдним набором двохроматидних хромосом;
• у клітинах тварин та деяких рослин хромосоми деспіралізуються і поділяється
цитоплазма материнської клітини, але в клітинах більшості видів рослин
цитоплазма не ділиться.
Результатом Мейозу І є утворення з однієї материнської клітини двох дочірніх
клітин з гаплоїдним набором двохроматидних хромосом.
Інтерфаза між поділами мейозу коротка або відсутня, оскільки синтез ДНК не
відбувається.
II етап – мітотичний, або Мейоз II
Профаза II – фаза спіралізації двохроматидних хромосом.
Метафаза II – фаза розташування двохроматидних хромосом на екваторі.
■ короткі нитки прикріплюються до центромер;
■ на екваторі клітини в один ряд розташовуються двохроматидні хромосоми.
Анафаза II – фаза розходження однохроматидних хромосом до полюсів клітин:
■ кожна хромосома розділюється на хроматиди;
■ нитки веретена поділу скорочуються і розтягують хроматиди до полюсів.
Телофаза II – фаза деспіралізації однохроматидних хромосом:
■ утворення двох клітин з гаплоїдним набором однохроматидних хромосом.
Отже, загальним результатом мейозу є утворення з однієї материнської клітини 4
дочірніх клітин з гаплоїдним набором однохроматидних хромосом.
Біологічне значення мейозу: 1) забезпечує видозміну спадкового матеріалу; 2)
підтримує сталість каріотипу при статевому розмноженні; 3) лежить в основі
статевого розмноження.
96.
97.
ПОРІВНЯЛЬНА ХАРАКТЕРИСТИКА МІТОЗУ І МЕЙОЗУОзнаки
Мітоз
Мейоз
Кількість поділів
Один
Два
Кількість утворених клітин 3 однієї
Дві
Чотири
Набір хромосом перед поділом у
клітинах
Диплоїдний
Диплоїдний
Набір хромосом у дочірніх клітинах Диплоїдний (2п1с)
Гаплоїдний (1п1с)
Стан спадкової інформації в
клітинах
Незмінений
Видозмінений
Відмінності процесів
у профазімітозу і профазі 1
мейозу
Відсутність кон'югації і
кросинговеру
Наявність кон'югації і кросинговеру
Відмінності процесів
у метафазімітозу і метафазі 1
мейозу
На екваторі хромосоми
розташовуються в один ряд
На екваторі хромосоми
розташовуються в два ряди у
вигляді тетрад
Відмінності процесів у анафазі
мітозу і анафазі 1 мейозу
Розходяться однохроматидні
хромосоми
Розходяться двохроматидні
хромосоми
Відмінності процесів у телофазі
мітозу і телофазі 1 мейозу
Утворюються дві диплоїдні клітини Утворюються дві гаплоїдні клітини
з однохроматидними хромосомами з двохроматидними хромосомами
98.
99.
Лептотена (стадія тонких ниток) — початок конденсації хромосом, вцілому нагадує ранню профазу мітозу, відрізняючись більш тонкими
хромосомами і великими ядрами.
Зиготена (стадія ниток, що зливаються) — зближення та початок
кон’югації гомологічних хромосом; наприкінці її всі гомологи поєднуються в
біваленти.
У пахітені (стадії товстих ниток) відбувається кросинговер — обмін
ідентичними ділянками між гомологічними хромосомами.
Диплотена (стадія подвійних ниток) починається взаємним
відштовхуванням гомологів та появою хіазм; у переважної більшості
організмів у диплотені відбувається подальша спіралізація хромосом та
редукція кількості ядерець; лише в ооцитах тварин, які накопичують багато
жовтка (деякі риби, земноводні, птахи, ссавці), а також у сперматоцитах
деяких комах хромосоми, навпаки, деконденсуються і набувають вигляду
«лампових щіток»; розпушення хромосом супроводжується активацією
процесів синтезу РНК та білка. Це найтриваліший період профази І. У
людини хромосоми типу «лампових щіток» можуть існувати протягом 12–50
років.
Для діакінезу (стадії відокремлення подвійних ниток) характерні
зменшення кількості хіазм та значна компактність бівалентів.
100.
101.
Живі організми існують тому, що постійно:а) в них надходять поживні речовини із
навколишнього середовища;
б) ці речовини перетворюються в організмі;
в) виводяться з організму продукти
життєдіяльності.
Сукупність всіх цих процесів називається
обмін речовин (метаболізм).
102. ЕНЕРГЕТИЧНИЙ ОБМІН.
1. Процеси поглинання із довкілля, засвоєнняі накопичення хімічних речовин, які необхідні
для утворення сполук, необхідних організму,
називаються асиміляцією (біосинтезом).
У кожній живій клітині здійснюється
величезна кількість хімічних реакцій. Всі вони
відбуваються організовано і упорядковано.
Кожна реакція відбувається у конкретно
визначеному місці і за участю ферментів –
каталізаторів, які розміщені на мембранах
мітохондрій та ЕПС.
103.
2. Розрізняють 2 типи реакцій у клітині:І-ший тип – реакції синтезу білків, жирів, вуглеводів,
нуклеїнових кислот, тобто асиміляція.
ІІ-ий тип: - реакція розщеплення складних органічних
речовин до менш складних сполук (СО2 і Н2О), які
супроводжуються виділенням енергії – дисиміляція.
3. Сукупність реакцій біосинтезу називають пластичним
обміном.
4. Сукупність реакцій розщеплення, що забезпечують
клітину енергією, називають енергетичним обміном.
5. енергетичний та пластичний обміни тісно пов’язані між
собою та зовнішнім середовищем і в єдності становлять
обмін речовин і енергії в кожній клітині і в організмі в
цілому.
Процеси асиміляції не завжди врівноважені з процесами
дисиміляції. Так, в організмах, що розвиваються,
переважає асиміляція (накопичуються речовини і росте
організм).
При інтенсивній фізичній роботі, нестачі поживних
речовин та старінні переважають процеси дисиміляції.
Для живих організмів Землі основним джерелом енергії є
сонячне світло.
104.
6. Організми, здатні утворювати органічні сполуки знеорганічних називаються автотрофами.
7. Організми, що використовують для утворення органічних
сполук з неорганічних енергію світла називають
фототрофами (зелені рослини, ціанобактерії).
8. Організми, що використовують для утворення органічних
речовин із неорганічних енергію хімічних реакцій називають
хемотрофами. (сіркобактерії, залізобактерії).
9. Організми, що використовують для утворення своїх
органічних речовин органічні речовини, утворені іншими
організмами (живі організми, їх рештки, продукти
життєдіяльності), які вони одержують з їжею, називають
гетеротрофами.
10. Енергетичний обмін речовин організмів здійснюється у
три послідовних етапи:
а) підготовчий.
б) безкисневий (анаеробне дихання)
в) кисневий (аеробне дихання).
105.
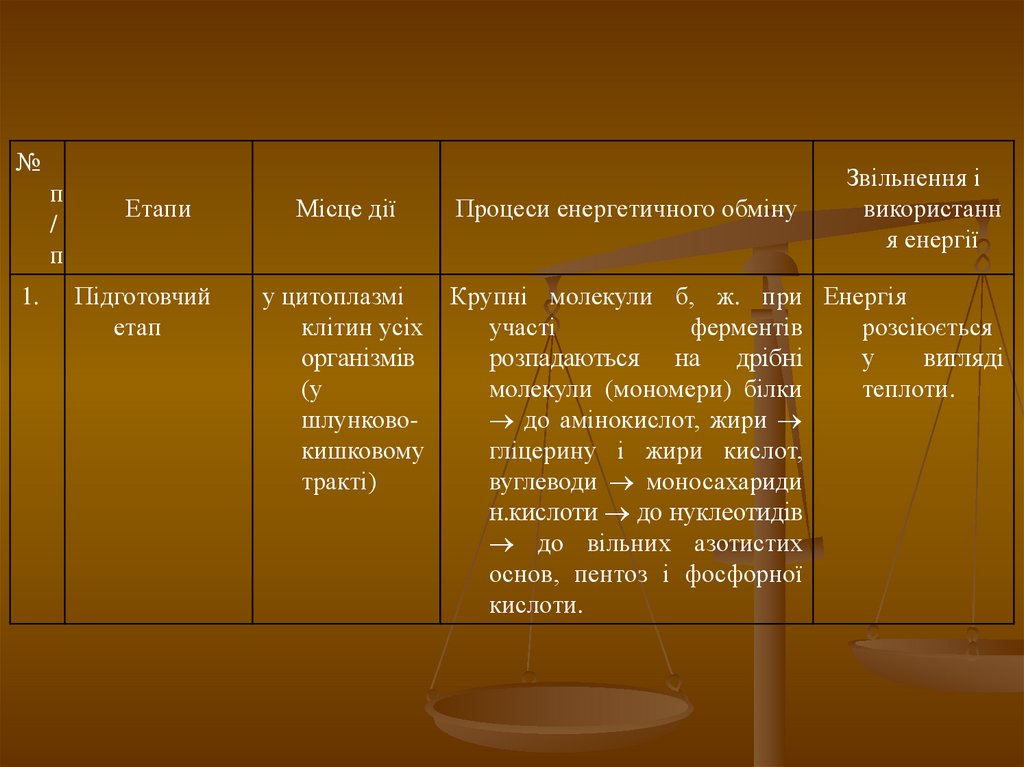
№п
/
п
1.
Етапи
Підготовчий
етап
Місце дії
Процеси енергетичного обміну
Звільнення і
використанн
я енергії
у цитоплазмі
Крупні молекули б, ж. при Енергія
клітин усіх
участі
ферментів
розсіюється
організмів
розпадаються на дрібні
у
вигляді
(у
молекули (мономери) білки
теплоти.
шлунково до амінокислот, жири
кишковому
гліцерину і жири кислот,
тракті)
вуглеводи моносахариди
н.кислоти до нуклеотидів
до вільних азотистих
основ, пентоз і фосфорної
кислоти.
106.
2.Безкисневий
в клітинах
(анаеробни
й) гліколіз
етап
неповне
розщепленн
я
3.
Кисневий
(аеробний)
етап
Амінокислоти, глюкоза та інші Розпад однієї
речовини, що утворюються
молекули
на
підготовчому
етапі,
глюкози дає
розщеплюються далі.
енергію, що
забезпечує
синтез 2х
молекул
АТФ
(виділяється
200 КДж
енергії).
мітохондріамні Дві молекули молочної кислоти Енергія від
мембрани.
розщеплюються за участю
розпаду 2х
АДФ і фосфорної кислоти.
молекул
молочної
кислоти
використову
ється для
синтезу 36
молекул
АТФ
107.
Найважливішим на безкисневому етапіенергетичного обміну є розщеплення в
клітинах молекул глюкози шляхом гліколізу
на дві молекули піровиноградної (С3Н4О3)
або молочної кислоти (С3Н6О3) у м’язових
клітинах:
С6Н12О6 + 2АДФ + 2Н3РО4 2С3Н6О3
+ 2АТФ + 2Н2О
У процесі розпаду глюкози беруть участь 13
різних ферментів, фосфорна кислота і АДФ.
Під час гліколізу виділяється 200 кДж
енергії . 84 кДж використовується на синтез
2х молекул АТФ, а решта (116 кДж)
використовується у вигляді теплоти.
108.
Значення гліколізу: організм дістаєенергію в умовах дефіциту кисню.
Спиртове бродіння – це один тип
перетворення глюкози, коли вона
розпадається на 2 молекули етилового
спирту
(С2Н5ОН) та 2 молекули вуглекислого
газу (СО2)
С6Н12О6 + 2Н3РО4 + 2АДФ = 2СО2
+ 2С2Н5ОН + 2АТФ + 2Н2О
Молочнокисле (молочне) бродіння – вид
безкисневого бродіння.
109.
12. Після завершення гліколізу настає друга стадія– кисневе розщеплення.
13. Процес кисневого розщеплення описується
рівнянням:
2С3Н6О3 + 6О2 + 36АДФ + 36Н3РО4
36АТФ + 6СО2 + 42Н2О
Це дихання
При цьому виділяється енергія (2600 кДж) частина
якої розсіюється у вигляді тепла (45%), 55%
перетворюється в енергію хімічних зв’язків АТФ
(1440 кДж).
14. Сумарне рівняння повного розщеплення
глюкози записується так:
С6Н12О6 + 6О2 + 38АДФ + 38Н3РО4
38АТФ + 6СО2 + 44Н2О
Це процес дихання.
110. ФОТОСИНТЕЗ.
Фототрофи використовують для синтезуорганічних сполук енергію світла. Процес
утворення органічних сполук із неорганічних
завдяки перетворенню світлової енергії в
енергію хімічних зв'язків називають
фотосинтезом. До фототрофних організмів
належать зелені рослини (вищі рослини,
водорості), деякі тварини (рослинні
джгутикові), а також деякі прокаріоти —
ціанобактерії, пурпурові та зелені
сіркобактерії.
111.
Нагадаймо, що коли певна сполука віддає електрон— вона окиснюється, а коли приєднує —
відновлюється. Відновлений НАДФ (НАДФ • Н2)
згодом постачає водень, потрібний для відновлення
атмосферного СО2 до глюкози (тобто сполуки, в якій
запасається енергія).
Подібні процеси відбуваються й у фотосистемі II.
Збуджені електрони, повертаючись на свій
енергетичний рівень, можуть передаватись
фотосистемі І і таким чином п відновлювати.
Фотосистема II відновлюється за рахунок електронів,
які постачають молекули води. Під дією світла за
участю ферментів молекули води розщеплюються
(фотоліз води) на протони водню та молекулярний
кисень, який виділяється в атмосферу, а електрони
використовуються на відновлення фотосистеми II:
112.
У процесі фотосинтезу у зелених рослин і ціанобактерій берутьучасть дві фотосистеми — перша (І) та друга (II), які мають різні
реакційні центри та пов'язані між собою через систему
перенесення електронів.
Процес фотосинтезу відбувається в дві фази — світлову та
темнову. У світлову фазу, реакції якої перебігають у мембранах
особливих структур хлоропластів - тилакоїдів за наявності світла
фотосинтезуючі пігменти вловлюють кванти світла (фотони).
Поглинання фотонів приводить до «збудження» одного з
електронів молекули хлорофілу, який за допомогою молекул —
переносників електронів переміщується на зовнішню поверхню
мембрани тилакоїдів, набуваючи певної потенційної енергії.
У фотосистемі І цей електрон може повертатись на свій
енергетичний рівень і відновлювати її, а може передаватись
такій сполуці, як НАДФ (нікотинамідаденіндинуклеотидфосфат).
Електрони, взаємодіючи з іонами водню, які є в навколишньому
середовищі, відновлюють цю сполуку:
113.
Енергія, вивільнена при поверненні електронів іззовнішньої поверхні мембрани тилакоїдів на попередній
енергетичний рівень, запасається у вигляді хімічних
зв'язків молекул АТФ, які синтезуються під час реакцій в
обох фотосистемах. Деяка її частина витрачається на
випаровування води. Таким чином, під час світлової фази
фотосинтезу утворюються багаті на енергію (яка
запасається у вигляді хімічних зв'язків) сполуки:
синтезується АТФ і відновлюється НАДФ. Як продукт
фотолізу води в атмосферу виділяється молекулярний
кисень.
Реакції темнової фази фотосинтезу перебігають у
внутрішньому середовищі (матриксі) хлоропластів як на
світлі, так і за його відсутності. Як згадувалося раніше, в
ході реакцій темпової фази С02 відновлюється до глюкози
завдяки енергії, що вивільнюється при розщепленні АТФ,
та за рахунок відновленого НАДФ.
Підсумкове рівняння фотосинтезу: