







 biology
biologySimilar presentations:


")


")




Биофизика клетки. Слияние мембран in vivo
1.
Биофизика клетки:двенадцатая лекция
Рассматриваемые вопросы:
1. Слияние мембран in vivo
2. Модельные системы, описывающие слияние мембран
3. Простейшая теория слияния мембран
4. Энергетическая траектория слияния и интермедиаты:
сталк, диафрагма полуслияния, пора слияния
5. Введение деформации наклона для описания слияния
6. Гидратационное отталкивание
2.
Энергия ХельфрихаB 2
2
B
W C1 C2 C0 C0 dS
2
2
суммарная кривизна
мембраны равна C1+C2
спонтанная кривизна липида:
кривизна мембраны, состоящей
только из липидов данного сорта,
в отсутствие внешних сил
поверхностное натяжение:
энергия единицы поверхности
мембраны без учета кривизны
3.
Примеры слияния мембран in vivo.1. Слияние синаптических везикул
4.
Слияние мембран происходит быстро подвоздействием белков
Palfreyman and Jorgensen, Contemporary Neuroscience, 2008
Jurado, Front. Cell. Neurosci., 2014
https://www.hhmi.org/research/speed-synapse
5.
Работа комплекса SNAREБелковый комплекс SNARE
изменяет свою конформацию и
«застегивается» подобно
застежке-молнии.
Работа одного комплекса SNARE
в процессе его конформационной
перестройки оценивается в 35 kT
Risselada, H. J., &
Grubmüller, H. (2021).
How proteins open fusion
pores: insights from
molecular
simulations. European
Biophysics Journal, 50,
279-293.
6.
Примеры слияния мембран in vivo.2. Слияние мембран в жизненном цикле
вируса гриппа
https://micro.magnet.fsu.edu/cells/viruses/influenzavirus.html
https://en.wikipedia.org/wiki/Influenza
7.
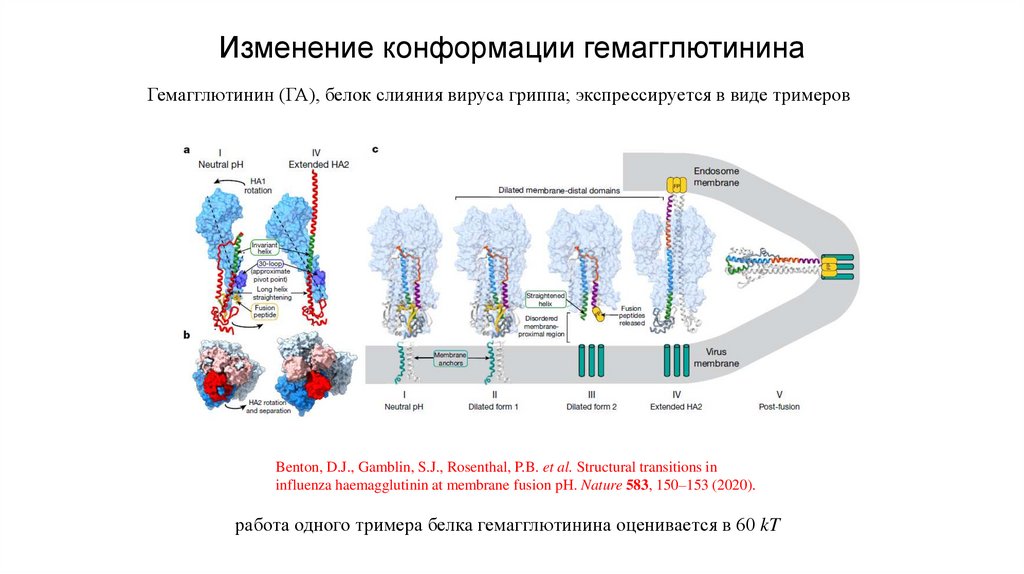
Изменение конформации гемагглютининаГемагглютинин (ГА), белок слияния вируса гриппа; экспрессируется в виде тримеров
Benton, D.J., Gamblin, S.J., Rosenthal, P.B. et al. Structural transitions in
influenza haemagglutinin at membrane fusion pH. Nature 583, 150–153 (2020).
работа одного тримера белка гемагглютинина оценивается в 60 kT
8.
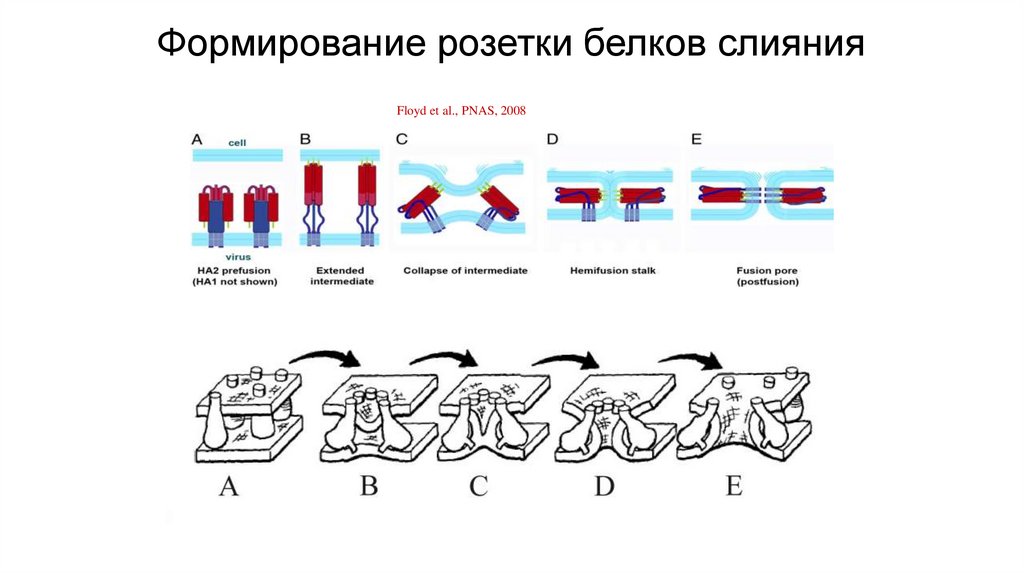
Формирование розетки белков слиянияFloyd et al., PNAS, 2008
9.
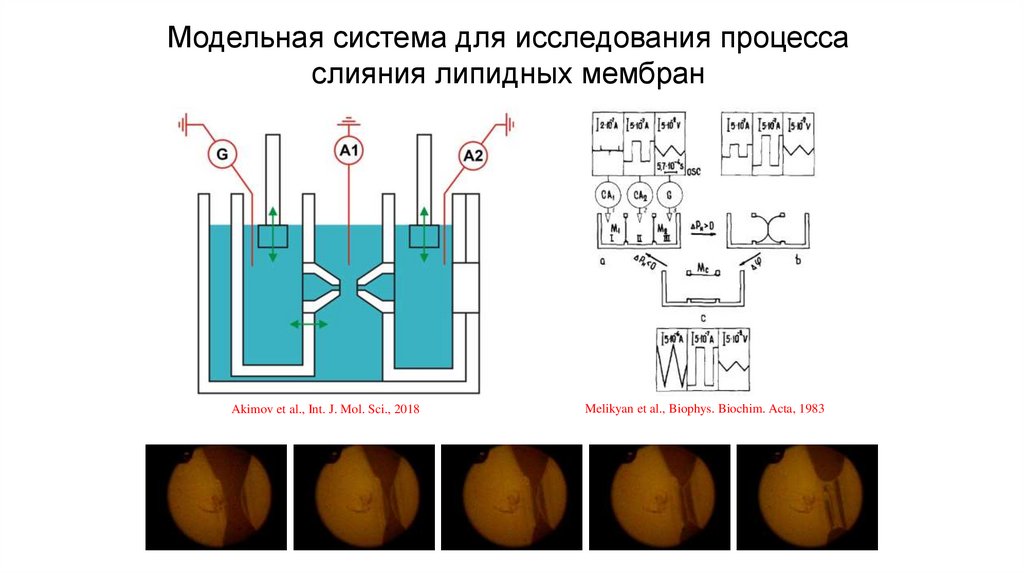
Модельная система для исследования процессаслияния липидных мембран
Akimov et al., Int. J. Mol. Sci., 2018
Melikyan et al., Biophys. Biochim. Acta, 1983
10.
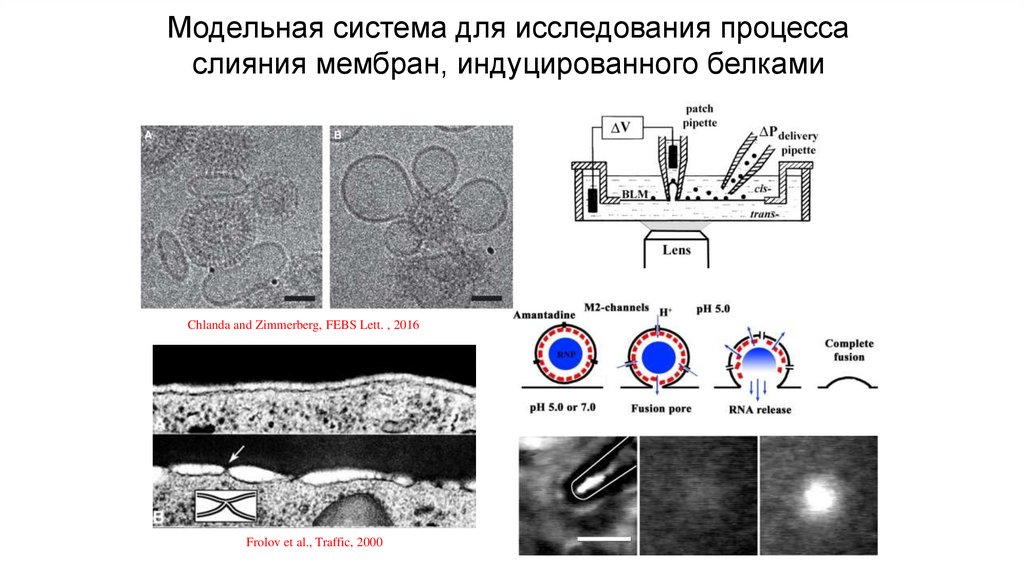
Модельная система для исследования процессаслияния мембран, индуцированного белками
Chlanda and Zimmerberg, FEBS Lett. , 2016
Frolov et al., Traffic, 2000
11.
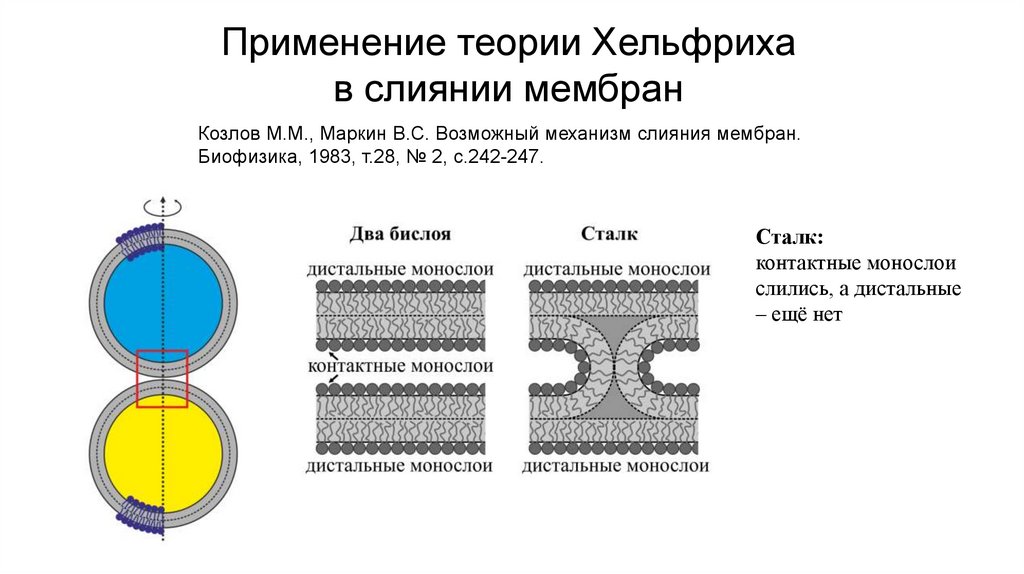
Применение теории Хельфрихав слиянии мембран
Козлов M.M., Маркин B.C. Возможный механизм слияния мембран.
Биофизика, 1983, т.28, № 2, с.242-247.
Сталк:
контактные монослои
слились, а дистальные
– ещё нет
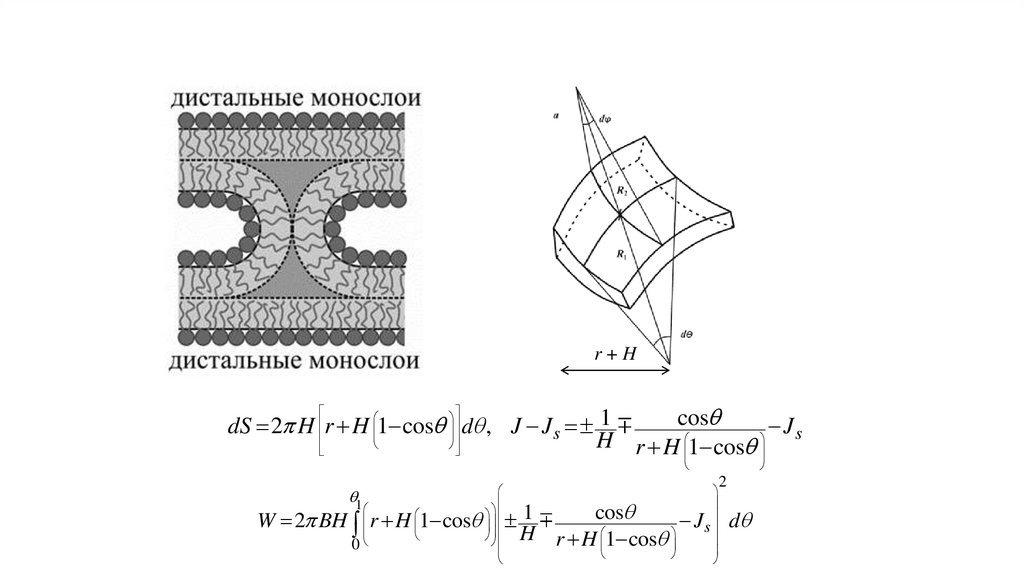
12.
r+Hcos
dS 2 H r H 1 cos d , J J s 1
J
H r H 1 cos s
1
W 2 BH r H 1 cos
0
cos
1
J
H r H 1 cos
s
2
d
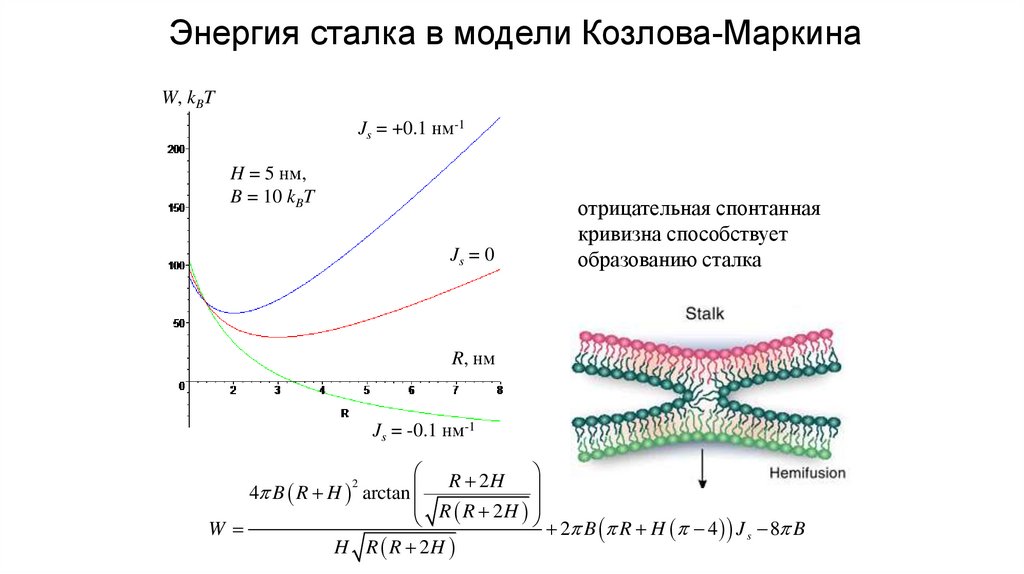
13.
Энергия сталка в модели Козлова-МаркинаW, kBT
Js = +0.1 нм-1
H = 5 нм,
B = 10 kBT
Js = 0
отрицательная спонтанная
кривизна способствует
образованию сталка
R, нм
Js = -0.1 нм-1
R 2H
4 B R H arctan
R R 2H
2 B R H 4 J 8 B
W
s
H R R 2H
2
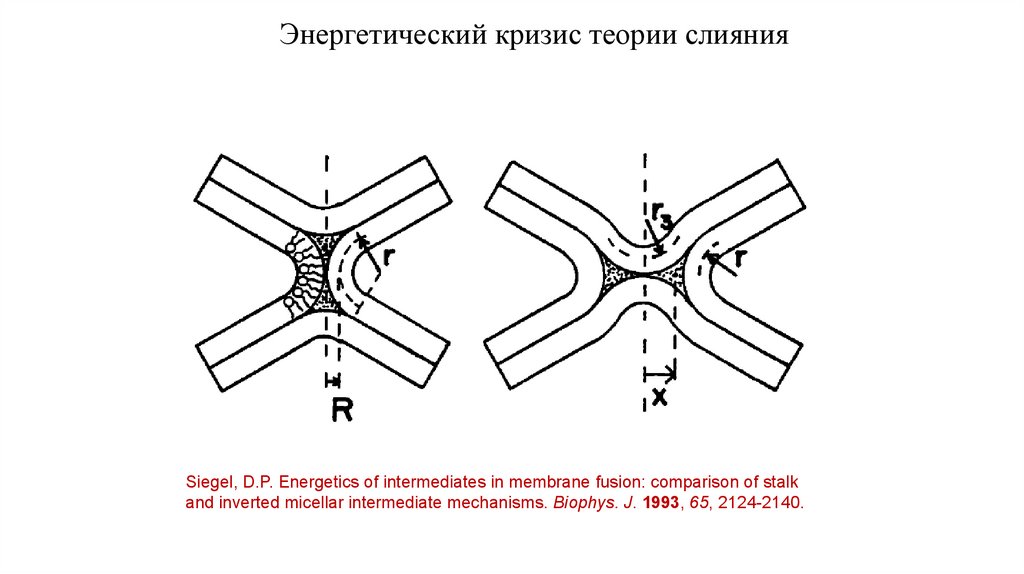
14.
Энергетический кризис теории слиянияSiegel, D.P. Energetics of intermediates in membrane fusion: comparison of stalk
and inverted micellar intermediate mechanisms. Biophys. J. 1993, 65, 2124-2140.
15.
Разрешение энергетического кризиса:введение деформации наклона
K
B
2
B
W divn C0 C02 t t 2 dS
2
2
2
наклон
Helfrich, W. Elastic properties of lipid
bilayers: theory and possible experiments.
Z. Naturforsch. C 1973, 28, 693-703.
Kuzmin et al., PNAS, 2001
Kuzmin, P.I.; Zimmerberg, J.; Chizmadzhev,
Y.A.; Cohen, F.S. A quantitative model for
membrane fusion based on low-energy
intermediates. Proc. Natl. Acad. Sci. USA 2001,
98, 7235-7240.
Markin, V.S.; Albanesi, J.P. Membrane
fusion: stalk model revisited. Biophys. J. 2002, 82,
693-712.
Kozlovsky, Y.; Kozlov, M.M. Stalk model of
membrane fusion: solution of energy crisis.
Biophys. J. 2002, 82, 882-895.
16.
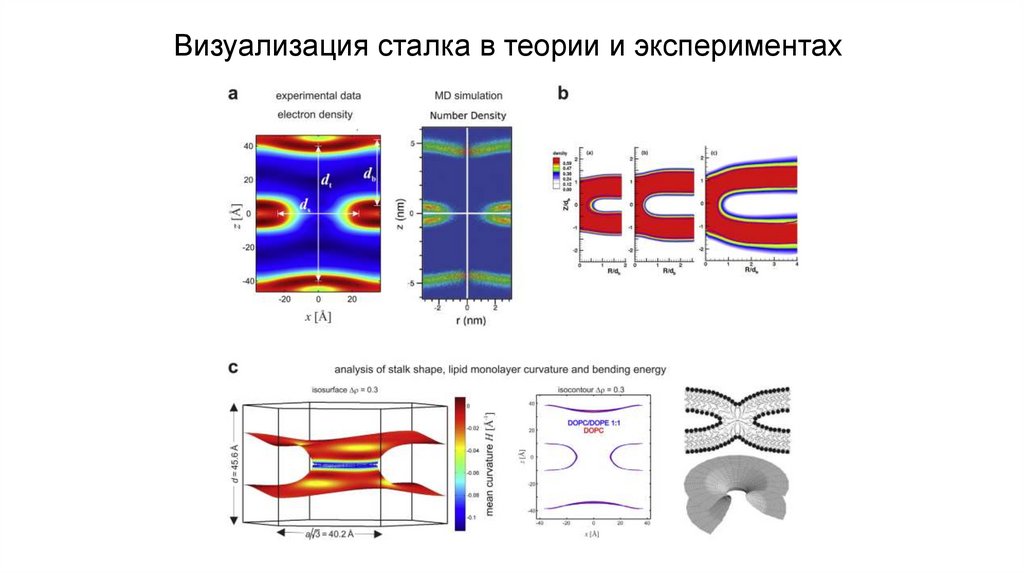
Визуализация сталка в теории и экспериментах17.
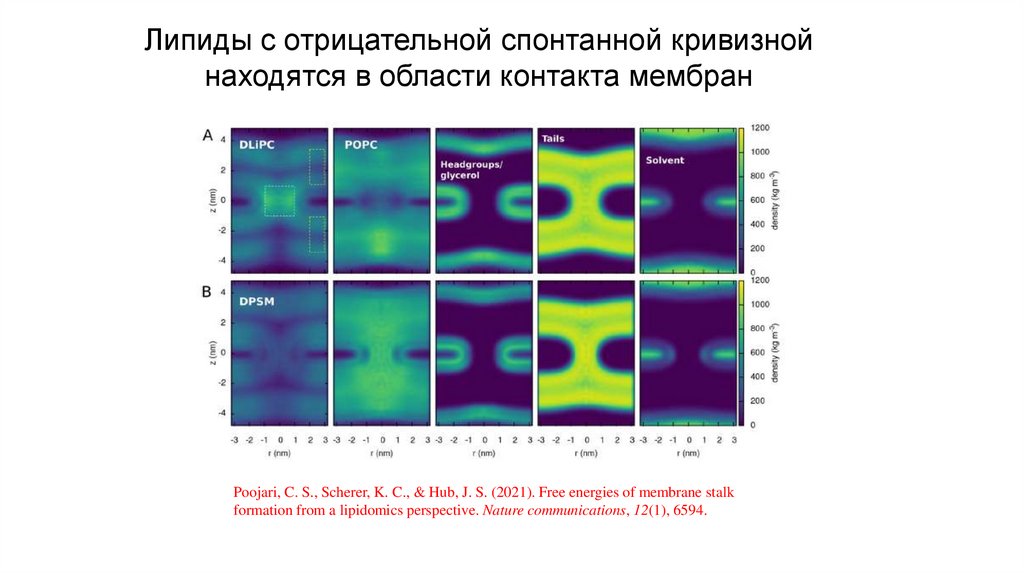
Липиды с отрицательной спонтанной кривизнойнаходятся в области контакта мембран
Poojari, C. S., Scherer, K. C., & Hub, J. S. (2021). Free energies of membrane stalk
formation from a lipidomics perspective. Nature communications, 12(1), 6594.
18.
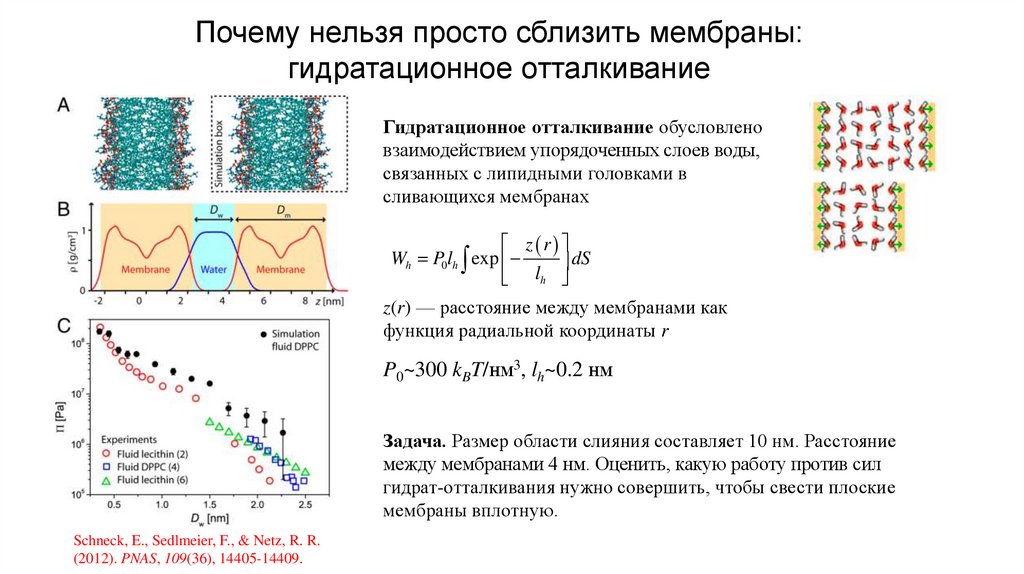
Почему нельзя просто сблизить мембраны:гидратационное отталкивание
Гидратационное отталкивание обусловлено
взаимодействием упорядоченных слоев воды,
связанных с липидными головками в
сливающихся мембранах
z r
Wh P0lh exp
dS
l
h
z(r) — расстояние между мембранами как
функция радиальной координаты r
P0~300 kBT/нм3, lh~0.2 нм
Задача. Размер области слияния составляет 10 нм. Расстояние
между мембранами 4 нм. Оценить, какую работу против сил
гидрат-отталкивания нужно совершить, чтобы свести плоские
мембраны вплотную.
Schneck, E., Sedlmeier, F., & Netz, R. R.
(2012). PNAS, 109(36), 14405-14409.
19.
Плотный контакт мембран.Образование гидрофобных дефектов
Мембранный процесс может проходить
самопроизвольно за время ~10 мин, если его
энергетической барьер не превышает ~40 kBT
Kuzmin et al., PNAS, 2001
Гидрофобные дефекты представляют собой зародыши будущего
контакта цис-монослоев мембран, поскольку они локально
разупорядочивают гидратационные слои, что приводит к притяжению
между мембранами. Притяжение увеличивается с уменьшением
расстояния между дефектами, что в конце концов приводит к их слиянию.
Поверхностная плотность энергии
гидрофобного притяжения
Akimov, S.A.; Polynkin, M.A.; Jiménez-Munguía, I.; Pavlov, K.V.;
Batishchev, O.V. Phosphatidylcholine membrane fusion is pHdependent. Int. J. Mol. Sci. 2018, 19, 1358.
d
lf
w f W 1 e ,
W ~ 12 kBT/нм2, lf ~ 1 нм
20.
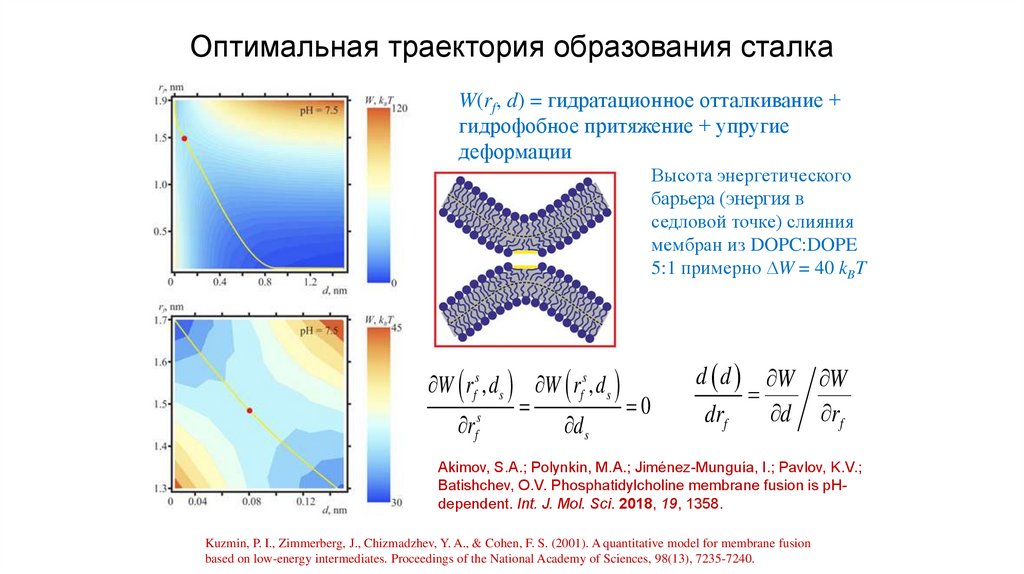
Оптимальная траектория образования сталкаW(rf, d) = гидратационное отталкивание +
гидрофобное притяжение + упругие
деформации
Высота энергетического
барьера (энергия в
седловой точке) слияния
мембран из DOPC:DOPE
5:1 примерно W = 40 kBT
W rfs , d s W rfs , d s
0
s
rf
d s
d d W W
d rf
drf
Akimov, S.A.; Polynkin, M.A.; Jiménez-Munguía, I.; Pavlov, K.V.;
Batishchev, O.V. Phosphatidylcholine membrane fusion is pHdependent. Int. J. Mol. Sci. 2018, 19, 1358.
Kuzmin, P. I., Zimmerberg, J., Chizmadzhev, Y. A., & Cohen, F. S. (2001). A quantitative model for membrane fusion
based on low-energy intermediates. Proceedings of the National Academy of Sciences, 98(13), 7235-7240.
21.
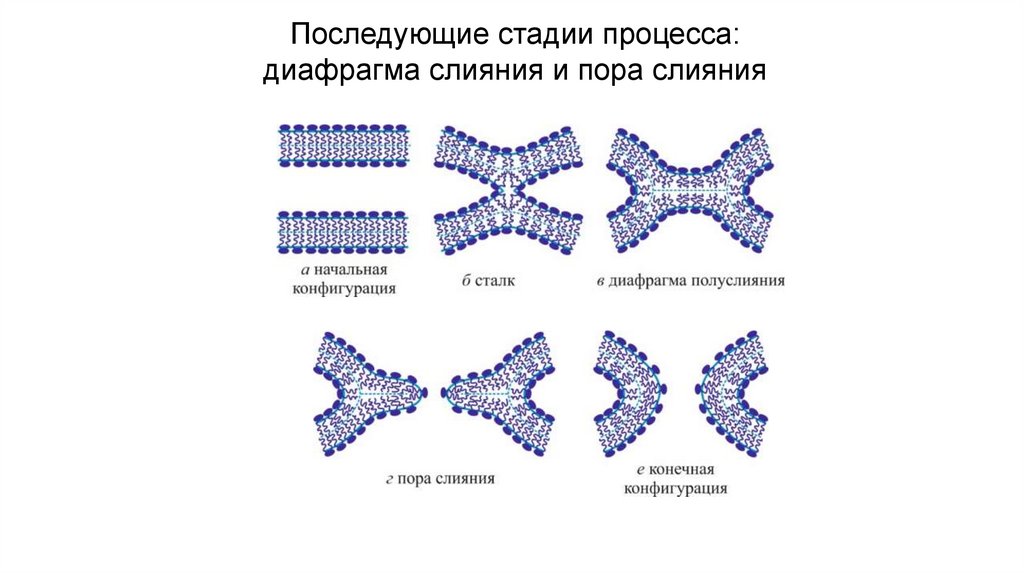
Последующие стадии процесса:диафрагма слияния и пора слияния
22.
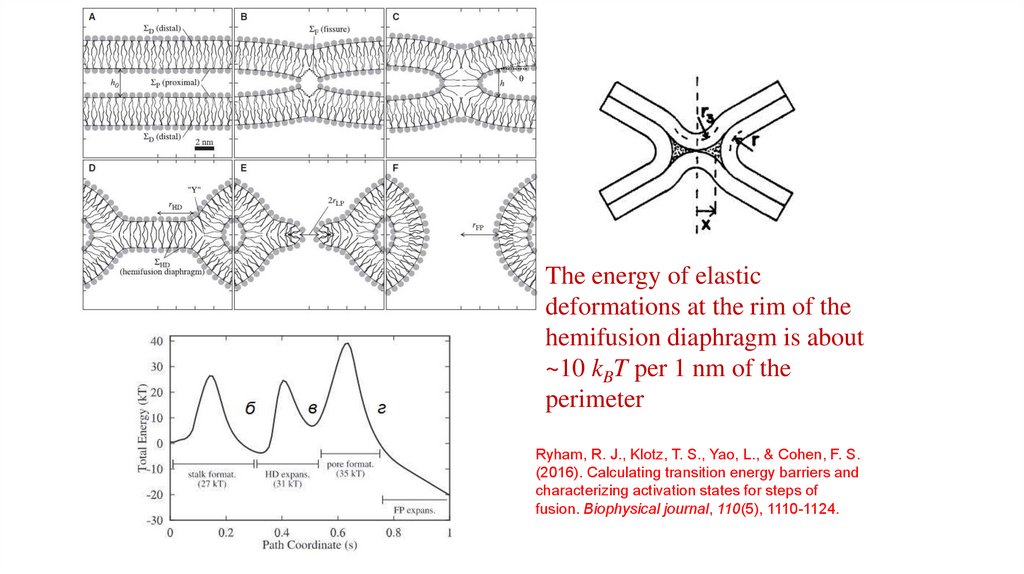
The energy of elasticdeformations at the rim of the
hemifusion diaphragm is about
~10 kBT per 1 nm of the
perimeter
Ryham, R. J., Klotz, T. S., Yao, L., & Cohen, F. S.
(2016). Calculating transition energy barriers and
characterizing activation states for steps of
fusion. Biophysical journal, 110(5), 1110-1124.