

















 biology
biologySimilar presentations:




")





Биофизика клетки. Четвертая лекция
1.
Биофизика клетки:четвертая лекция
Рассматриваемые вопросы:
1. Электрические потенциалы на мембране
2. Двойной электрический слой и его структура
3. Дзета-потенциал и уравнение Смолуховского
4. Граничный потенциал. Метод компенсации внутримембранного поля
5. Монослойные пленки Ленгмюра-Блоджетт
6. Дипольный момент и уравнение Гельмгольца
2.
Разбор задачЗадача 1
Вариант 2. Что такое электрохимический потенциал? Какие величины в него входят и что они выражают?
Электрохимический потенциал заряженной частицы:
Энергия, приходящаяся на одну частицу
RT ln c zF
0
Стандартный
хим.потенциал
Осмотический вклад
в потенциал
Электрический
вклад в
потенциал
Равенство нулю разности потенциалов —критерий
термодинамического равновесия системы
3.
Разбор задачЗадача 1
Вариант 1. Что такое потенциал Нернста? Чему он равен и в каких предположениях получен?
Потенциал Нернста – разность потенциалов на бесконечно тонкой мембране, проницаемой для ионов калия.
Получается путем приравнивания электрохим. потенциалов для калия с двух сторон от мембраны
A+B-
A+BA+
A (1) A (2)
2
C
2
1
RT ln 1 zF 0
0
0
A (1) A (2)
C
A+
внутри
клетки (1)
здесь калия
больше
снаружи
клетки (2)
здесь калия
меньше
1
kT
C
ln 2 88мВ
e C
4.
Потенциал Нернста: выводA (1) A0 (1) kT ln c A (1) e (1)
A (2) A0 (2) kT ln c A (2) e (2)
kT ln c1 e 1 kT ln c2 e 2
e 2 e 1 kT ln c1 kT ln c2
kT c1
2 1
ln
e c2
A (1) A (2)
A0 (1) A0 (2)
f2
f1
K+
K+
внутри клетки
C1 = 150 мМ
снаружи клетки
С2 = 5 мМ
5.
Задача 2Вариант 1. Концентрация белка в клетке составляет 1 мМ, заряд одной молекулы белка равен 2. Концентрация
электролитов равна 150 мМ. Чему равен потенциал Доннана?
Вариант 2. Концентрация белка в клетке составляет 100 мМ, заряд одной молекулы белка равен 10.
Концентрация электролитов равна 150 мМ. Чему равен потенциал Доннана?
Q=сQzQ Случай Q<<c (1 вариант):
kT Q
1мМ 2
1
25 мВ
25 мВ
0.17 мВ
e 2c
2 150 мМ
150
Случай Q>>c (2 вариант):
kT Q
100 мМ 10
1000
ln 25 мВ ln
25 мВ ln
25 мВ 1.9 47 мВ
e c
150 мМ
150
6.
Задача 3Вариант 1. Концентрация ионов хлора внутри клетки составляет 9 мМ, снаружи клетки 125 мМ. Разность
потенциалов на мембране равна fin ‒ fout = ‒ 0.088 В. Посчитать энергию, требуемую для переноса ионов хлора
изнутри наружу. Ответ выразить в кДж/моль.
Вариант 2. Концентрация ионов кальция внутри клетки составляет 0.01 мМ, снаружи клетки 2.5 мМ. Разность
потенциалов на мембране равна fin ‒ fout = ‒ 0.088 В. Посчитать энергию, требуемую для переноса ионов
кальция изнутри наружу. Ответ выразить в кДж/моль.
out – конечное состояние; in – начальное состояние:
out
C
out in RT ln in zF out in
C
Дж
Кл
Дж
Дж
кДж
125 мМ
300 К ln
1
96500
0.088
В
6559
8492
1.9
моль К
моль
моль
моль
моль
9 мМ
1 вариант: z = ‒1
8.31
2 вариант: z = 2
8.31
Дж
Кл
Дж
Дж
кДж
2.5 мМ
300 К ln
2
96500
0.088
В
13765
16984
30.7
моль К
моль
моль
моль
моль
0.01мМ
7.
Задача 4Вариант 1. Коэффициент распределения g иона на границе вода/мембрана равен g 10–20. Найти изменение
энергии при переходе иона из воды в мембрану. Ответ выразить в единицах kBT.
Вариант 2. Коэффициент распределения g иона на границе вода/мембрана равен g 10–30. Найти изменение
энергии при переходе иона из воды в мембрану. Ответ выразить в единицах kBT.
g e
0
kT
ln g k BT
0
g 10 20 0 ln 10 20 k BT 43.7 k BT 46k BT
g 10 30 0 ln 10 30 k BT 66.8 k BT 69k BT
8.
Краткое содержание предыдущей лекции1. Дебаевская длина экранирования – характеризует
масштаб расстояний, на которых меняется потенциал
вблизи мембраны
1
0 kT
2c0 z 2 e 2
Для расчетов можно пользоваться приближенной формулой:
0.3 / z 2 c0 нм, где с0 измеряется в моль/л
2. Внутри мембраны потенциал меняется линейно:
d
const E x
dx
9.
Сведения из электростатикиДипольный момент р:
10.
Оценка напряженности электрического поляв незаряженной мембране
2 1
d
const E x
dx
L
L
1
2
0.05В; L 5нм
x
В
E 10
м
7
11.
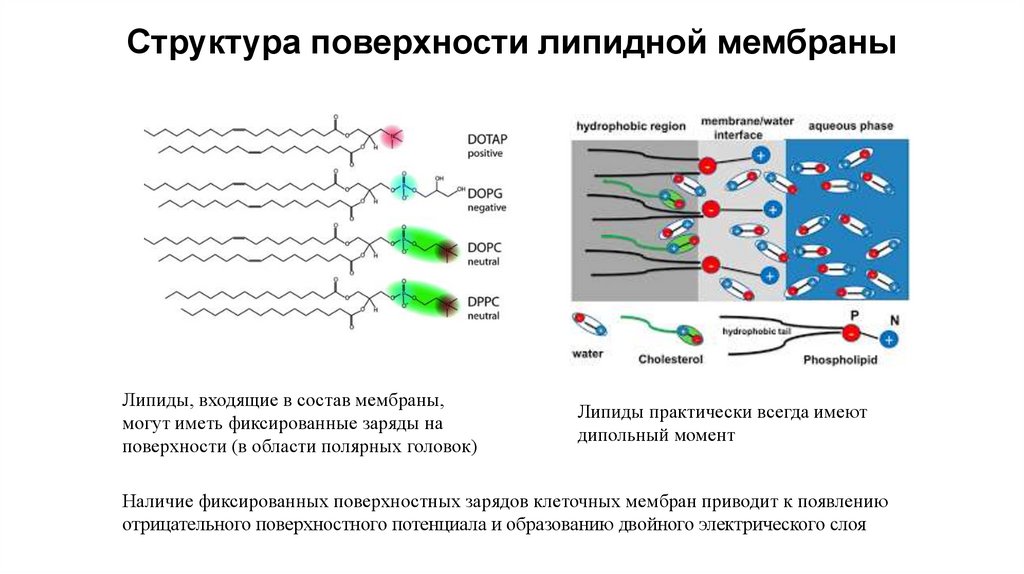
Структура поверхности липидной мембраныЛипиды, входящие в состав мембраны,
могут иметь фиксированные заряды на
поверхности (в области полярных головок)
Липиды практически всегда имеют
дипольный момент
Наличие фиксированных поверхностных зарядов клеточных мембран приводит к появлению
отрицательного поверхностного потенциала и образованию двойного электрического слоя
12.
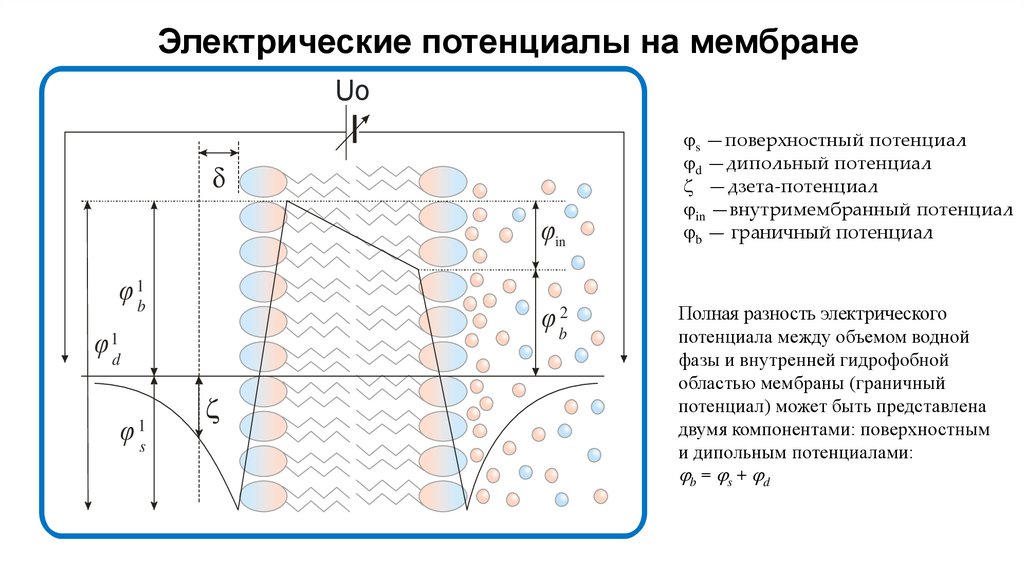
Электрические потенциалы на мембранеUo
d
φin
φ 1b
φ b2
φ 1d
φ
1
s
z
s —поверхностный потенциал
d —дипольный потенциал
z —дзета-потенциал
in —внутримембранный потенциал
b — граничный потенциал
Полная разность электрического
потенциала между объемом водной
фазы и внутренней гидрофобной
областью мембраны (граничный
потенциал) может быть представлена
двумя компонентами: поверхностным
и дипольным потенциалами:
b = s + d
13.
Двойной электрический слойРаспределение ионов около заряженной поверхности определяется двумя факторами: электростатическим
притяжением и тепловым движением (теория Гуи-Чепмена). В устанавливающемся равновесии плотность
распределения ионов убывает с увеличением расстояния от поверхности. Слой раствора с измененными
концентрациями вблизи поверхности называется диффузным, или двойным электрическим слоем.
Связь между плотностью заряда на поверхности и поверхностным потенциалом:
e s
e s
exp
2kT
2kT
2 0 kTc0 exp
14.
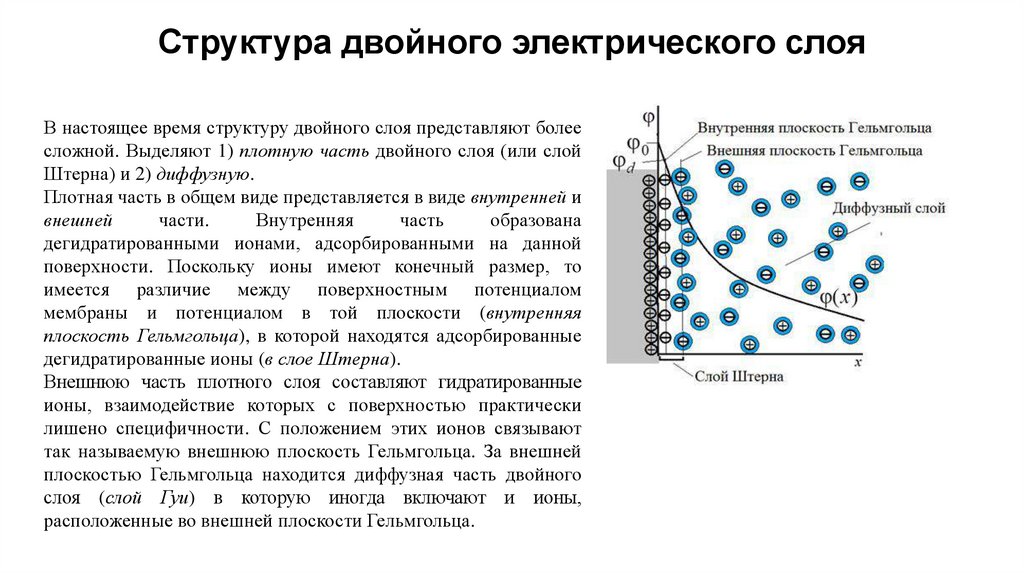
Структура двойного электрического слояВ настоящее время структуру двойного слоя представляют более
сложной. Выделяют 1) плотную часть двойного слоя (или слой
Штерна) и 2) диффузную.
Плотная часть в общем виде представляется в виде внутренней и
внешней
части.
Внутренняя
часть
образована
дегидратированными ионами, адсорбированными на данной
поверхности. Поскольку ионы имеют конечный размер, то
имеется различие между поверхностным потенциалом
мембраны и потенциалом в той плоскости (внутренняя
плоскость Гельмгольца), в которой находятся адсорбированные
дегидратированные ионы (в слое Штерна).
Внешнюю часть плотного слоя составляют гидратированные
ионы, взаимодействие которых с поверхностью практически
лишено специфичности. С положением этих ионов связывают
так называемую внешнюю плоскость Гельмгольца. За внешней
плоскостью Гельмгольца находится диффузная часть двойного
слоя (слой Гуи) в которую иногда включают и ионы,
расположенные во внешней плоскости Гельмгольца.
15.
Дзета-потенциалПри движении частицы двойной электрический слой разрывается. Место разрыва при перемещении
твердой и жидкой фаз друг относительно друга называется плоскостью скольжения. Плоскость
скольжения лежит на границе между диффузными и адсорбционными слоями, либо в диффузном слое
вблизи этой границы. Потенциал на плоскости скольжения называют электрокинетическим или дзетапотенциалом (ζ-потенциал).
Приложенное электрическое поле заставляет перемещаться
подвижные ионы двойного электрического слоя, ионы
увлекают за собой жидкость за счет сил вязкого трения.
16.
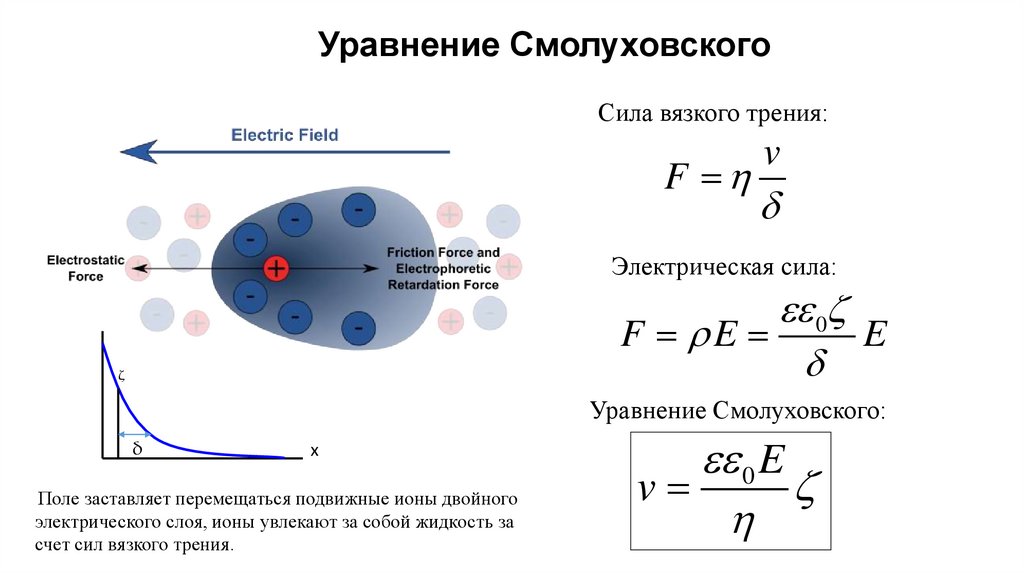
Уравнение СмолуховскогоСила вязкого трения:
F
v
d
Электрическая сила:
0z
F E
E
d
ζ
Уравнение Смолуховского:
δ
x
Поле заставляет перемещаться подвижные ионы двойного
электрического слоя, ионы увлекают за собой жидкость за
счет сил вязкого трения.
0 E
v
z
17.
Использование дзета-потенциалаВажность дзета-потенциала состоит в том, что его значение может быть связано с устойчивостью
коллоидных дисперсий. Дзета-потенциал определяет степень и характер взаимодействия между частицами
дисперсной системы.
Для молекул и частиц, которые достаточно малы, высокий дзета-потенциал будет означать стабильность, т.е.
раствор или дисперсия будет устойчивы по отношению к агрегации. Когда дзета-потенциал низкий,
притяжение превышает отталкивание, и устойчивость дисперсии будет нарушаться. Так, коллоиды с
высоким дзета-потенциалом являются электрически стабилизированными, в то время, как коллоиды с
низким дзета-потенциалом склонны коагулировать или флокулировать.
Значение дзета-потенциала равное 30 мВ (положительное или отрицательное) можно рассматривать как
характерное значение, для условного разделения низко-заряженных поверхностей и высоко-заряженных
поверхностей. Чем больше электрокинетический потенциал, тем устойчивее коллоид.
18.
Граничный потенциалГраничный потенциал складывается из двух компонент — поверхностного и дипольного потенциалов:
b s d
Поверхностный потенциал определяет концентрацию ионов
на границе мембраны:
ze s
cM g c exp
kT
Проводимость мембраны определяется произведением сМ на константу скорости перемещения ионов через мембрану:
ze s
ze d
ze b
G g c exp
k
exp
G
exp
0
0
kT
kT
kT
19.
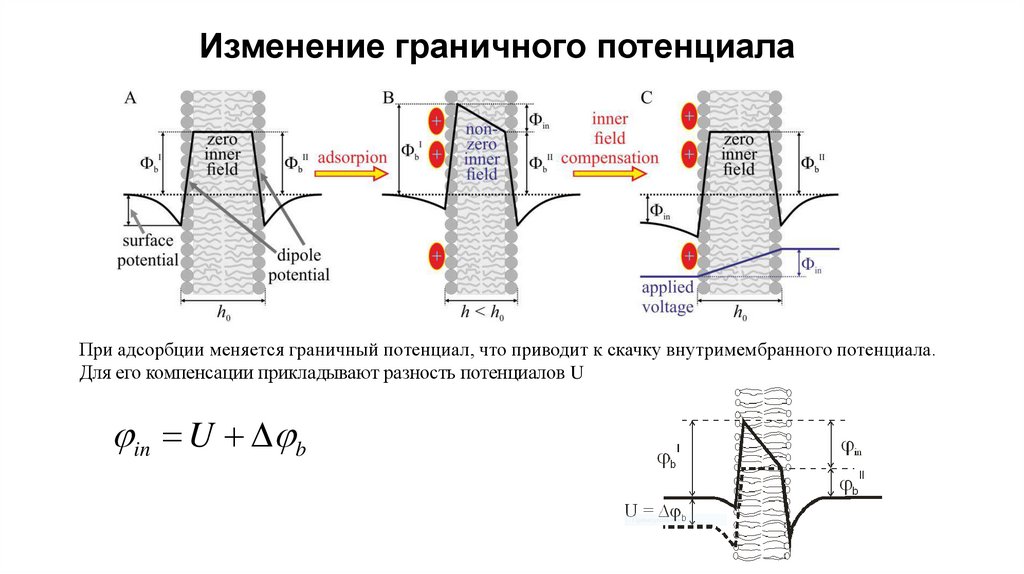
Изменение граничного потенциалаПри адсорбции меняется граничный потенциал, что приводит к скачку внутримембранного потенциала.
Для его компенсации прикладывают разность потенциалов U
in U b
20.
Метод компенсации внутримембранного поляC
U
Зависимость ёмкости от потенциала U
имеет минимум, когда in = 0:
in U b
Количественно выражается закономерностью:
Мембрана как конденсатор с упругостью: его пластины
сближаются и увеличивают емкость настолько, насколько
позволяют упругие свойства
C
S
; f 0 h ; C
h
C C0 1 (U b ) 2
21.
Модуляция второй гармоникиВ качестве U берут переменную величину; методика предложена Соколовым В.С., Кузьминым П.И.
C C0 1 (U b ) 2
I
U U0 Vcos( t)
dQ
dU
dC
C
U
dt
dt
dt
3
2
2
I1 V C0 1 3 U 0 V sin( t )
4
2
I 2 3 C0 V U 0 fb sin(2 t )
Индикатором минимума емкости может служить равенство нулю второй гармоники тока
22.
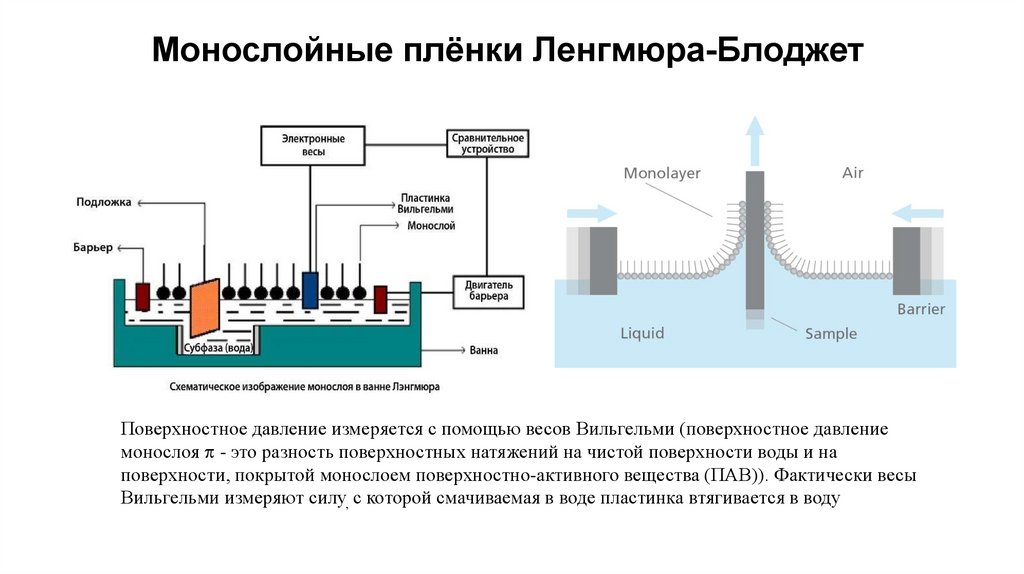
Монослойные плёнки Ленгмюра-БлоджетПоверхностное давление измеряется с помощью весов Вильгельми (поверхностное давление
монослоя - это разность поверхностных натяжений на чистой поверхности воды и на
поверхности, покрытой монослоем поверхностно-активного вещества (ПАВ)). Фактически весы
Вильгельми измеряют силу, с которой смачиваемая в воде пластинка втягивается в воду
23.
Ленгмюровские монослоиComprehensive Study of the Liquid Expanded–Liquid Condensed
Phase Transition in 1,2-Dimyristoyl-sn-glycero-3-phospho-L-serine
Monolayers: Surface Pressure, Volta Potential, X-ray Reflectivity, and
Molecular Dynamics Modeling
Yu. A. Ermakov, V. E. Asadchikov, B. S. Roschin, Yu. O. Volkov, D. A.
Khomich, A. M. Nesterenko, and A. M. Tikhonov
Langmuir 2019 35 (38), 12326-12338
24.
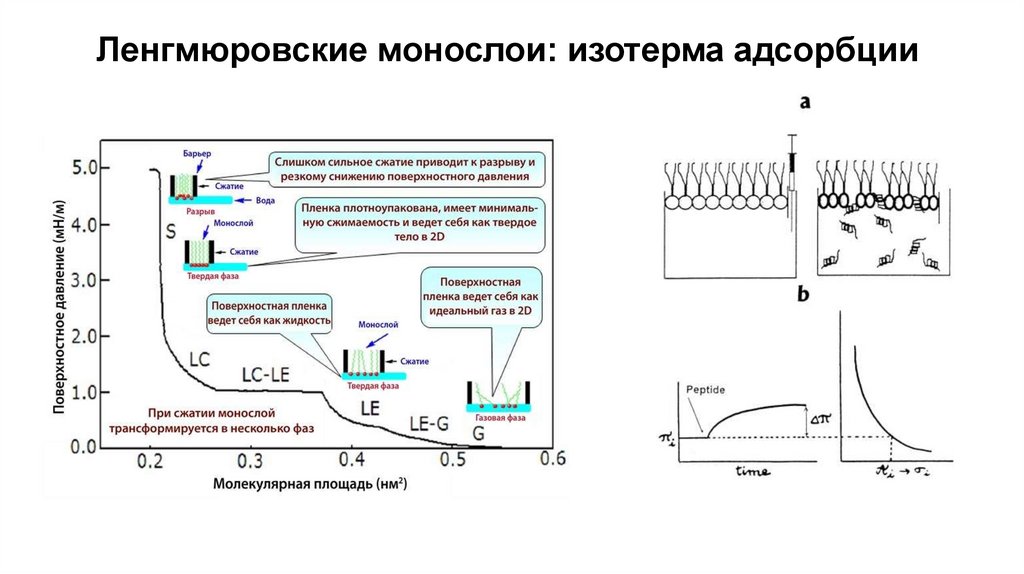
Ленгмюровские монослои: изотерма адсорбции25.
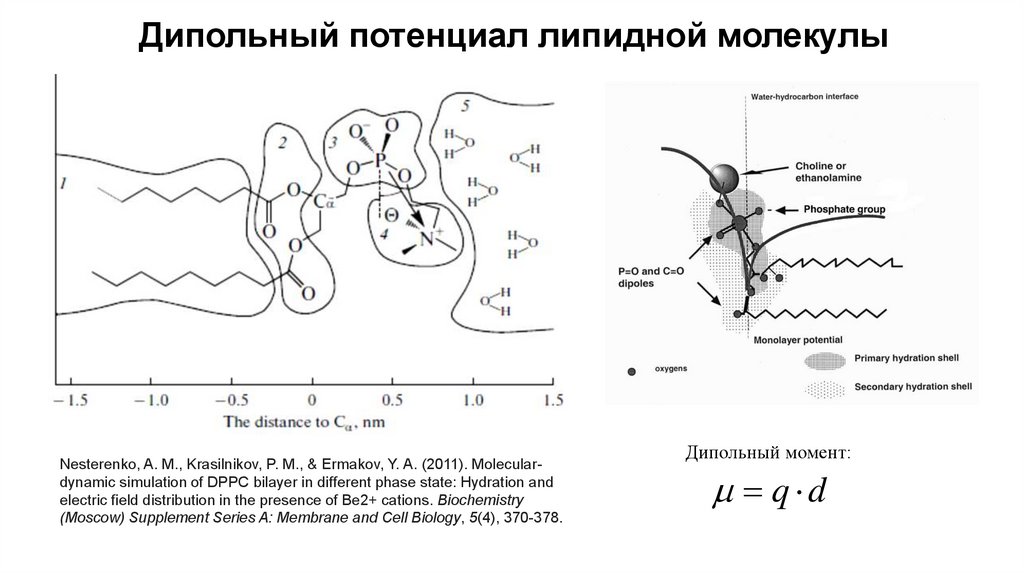
Дипольный потенциал липидной молекулыNesterenko, A. M., Krasilnikov, P. M., & Ermakov, Y. A. (2011). Moleculardynamic simulation of DPPC bilayer in different phase state: Hydration and
electric field distribution in the presence of Be2+ cations. Biochemistry
(Moscow) Supplement Series A: Membrane and Cell Biology, 5(4), 370-378.
Дипольный момент:
q d
26.
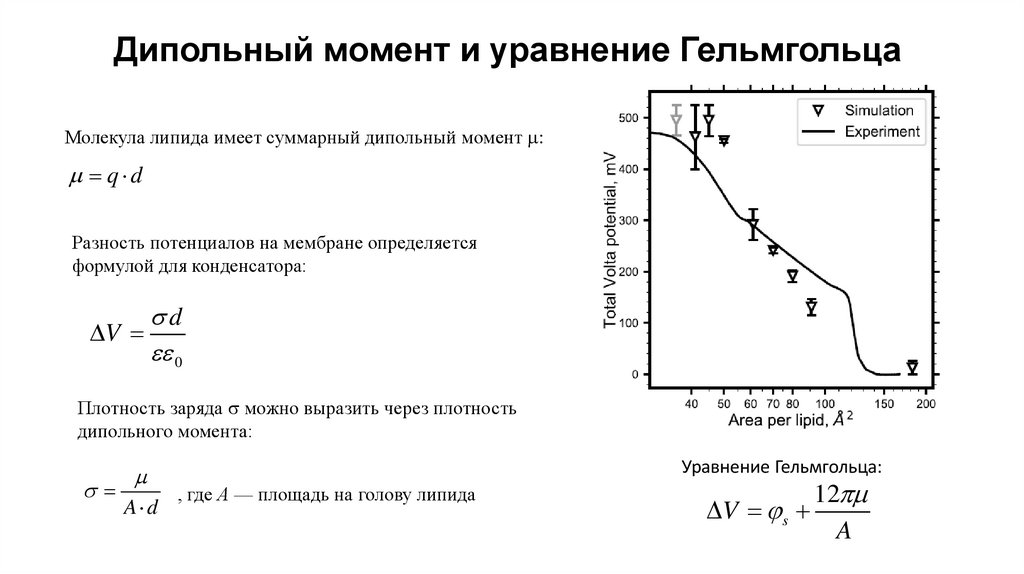
Дипольный момент и уравнение ГельмгольцаМолекула липида имеет суммарный дипольный момент :
q d
Разность потенциалов на мембране определяется
формулой для конденсатора:
d
V
0
Плотность заряда можно выразить через плотность
дипольного момента:
A d
Уравнение Гельмгольца:
, где А — площадь на голову липида
12
V s
A
27.
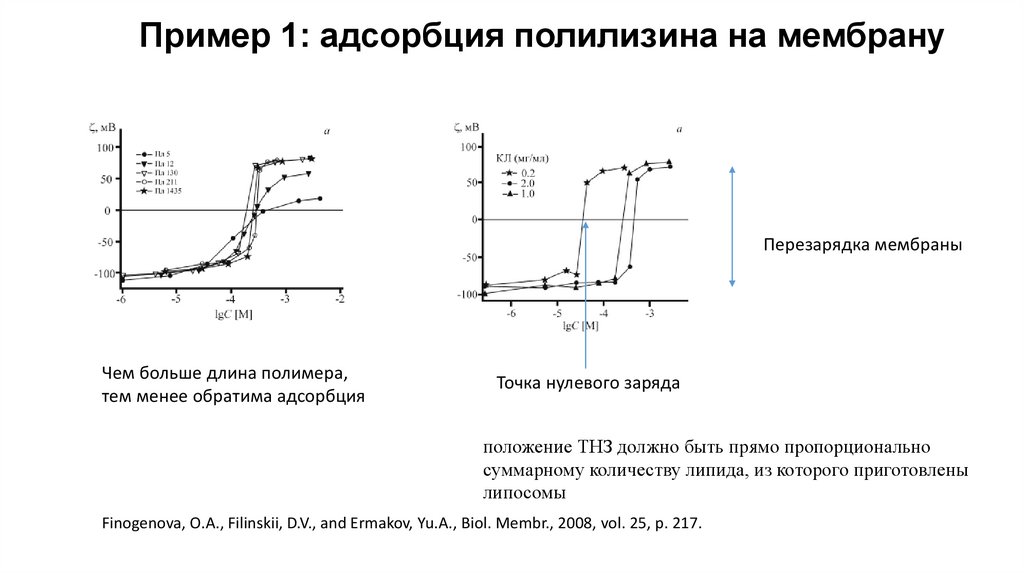
Пример 1: адсорбция полилизина на мембрануПерезарядка мембраны
Чем больше длина полимера,
тем менее обратима адсорбция
Точка нулевого заряда
положение ТНЗ должно быть прямо пропорционально
суммарному количеству липида, из которого приготовлены
липосомы
Finogenova, O.A., Filinskii, D.V., and Ermakov, Yu.A., Biol. Membr., 2008, vol. 25, p. 217.
28.
Конформация полилизина на поверхностиКонформация полимера зависит от
соотношения полимер/липид
Полимер занимает не всю доступную поверхность
мембраны
Poly 1 Lipid
Molotkovsky, R. J., Galimzyanov, T. R., Khomich, D. A., Nesterenko, A. M., & Ermakov, Y. A. (2021). Inhomogeneity of polylysine adsorption layers on lipid
membranes revealed by theoretical analysis of electrokinetic data and molecular dynamics simulations. Bioelectrochemistry, 141, 107828.
29.
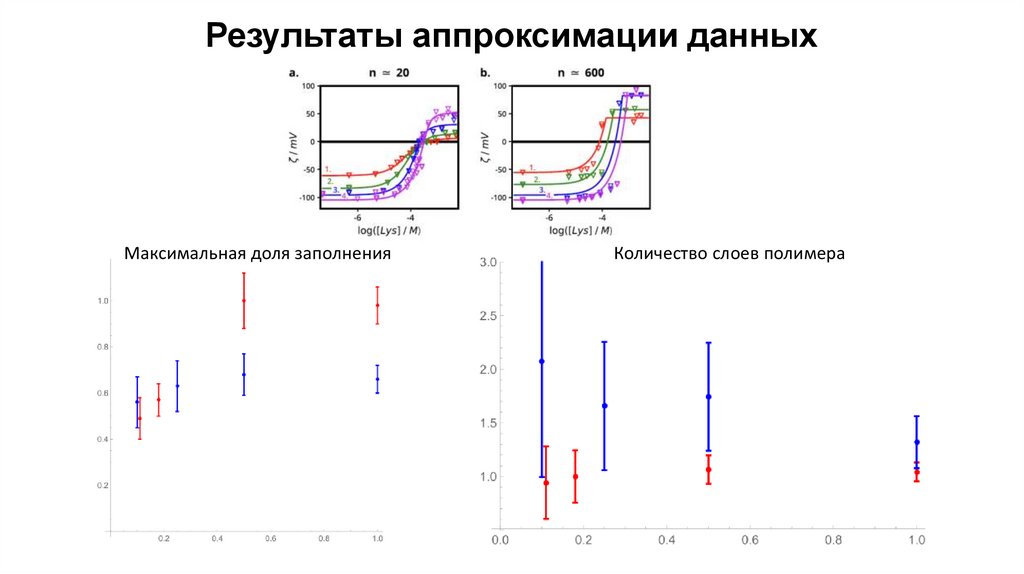
Результаты аппроксимации данныхМаксимальная доля заполнения
Количество слоев полимера
30.
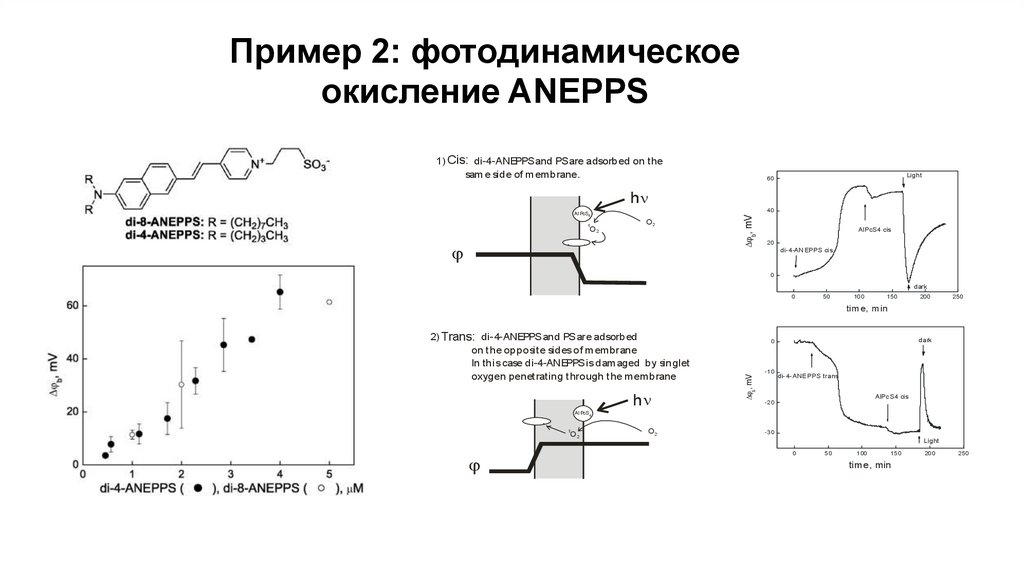
Пример 2: фотодинамическоеокисление ANEPPS
1) Cis: di-4-ANEPPS and PS are adsorbed on the
sam e side of m embrane.
Light
60
hn
1
O2
-
O2
b, mV
40
Al PcS4
AlPcS4 cis
20
+
di-4-ANEPPS cis
0
dark
0
50
10 0
1 50
200
25 0
tim e, m in
hn
dark
0
b, mV
2) Trans: di-4-ANEPPS and PS are adsorbed
on t he opposit e sides of membrane
In this case di-4-ANEPPS is damaged by singlet
oxygen penet rat ing t hrough t he membrane
-1 0
di-4-ANE PPS trans
AlPc S4 cis
-2 0
Al PcS4
+
-
1
O2
O2
-3 0
Light
0
50
1 00
15 0
time, min
20 0
2 50
31.
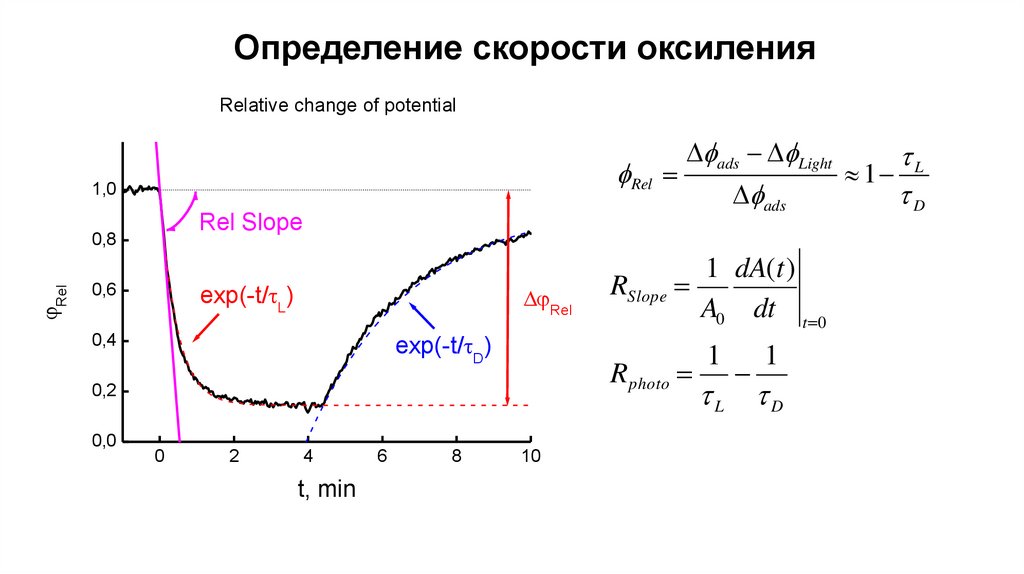
Определение скорости оксиленияRelative change of potential
fRel
1,0
Rel
fads
Rel Slope
0,8
exp(-t/ L)
0,6
Rel
exp(-t/ D)
0,4
0
2
4
t, min
6
8
1 dA(t )
RSlope
A0 dt t 0
R photo
0,2
0,0
fads fLight
10
1
L
1
D
L
1
D
32.
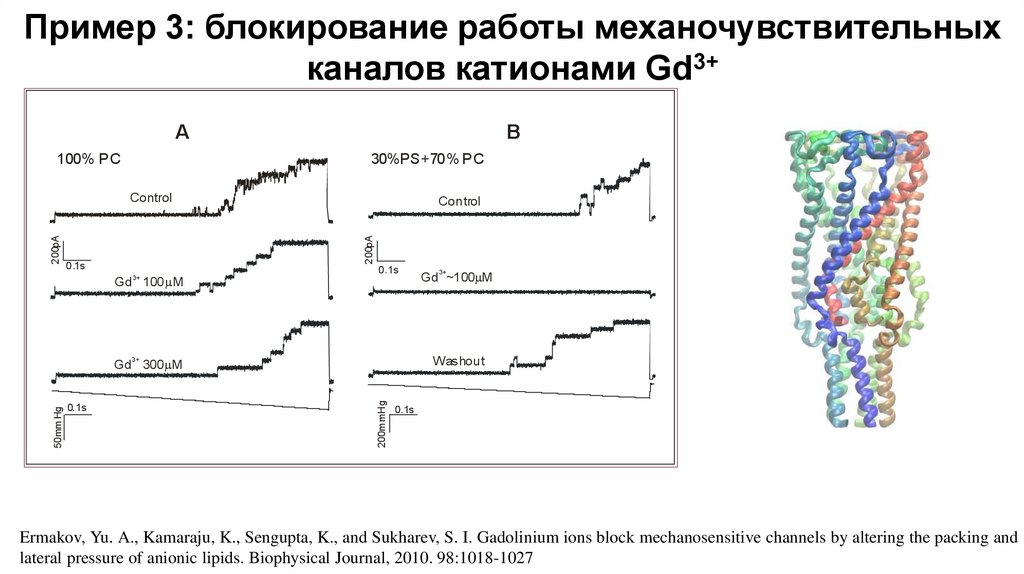
Пример 3: блокирование работы механочувствительныхканалов катионами Gd3+
A
100% PC
B
30%PS+70% PC
Control
200pA
200pA
Control
0.1s
Gd3+ 100 M
0.1s
Gd 300 M
200mmHg
50mm Hg
3+
Washout
3+
0.1s
Gd ~100 M
0.1s
Ermakov, Yu. A., Kamaraju, K., Sengupta, K., and Sukharev, S. I. Gadolinium ions block mechanosensitive channels by altering the packing and
lateral pressure of anionic lipids. Biophysical Journal, 2010. 98:1018-1027
33.
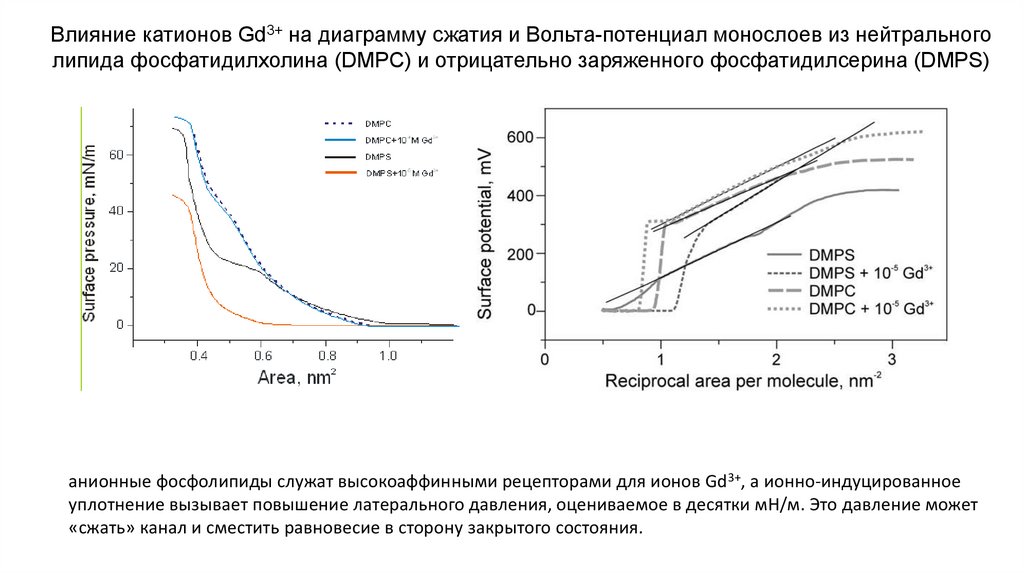
Влияние катионов Gd3+ на диаграмму сжатия и Вольта-потенциал монослоев из нейтральноголипида фосфатидилхолина (DMPC) и отрицательно заряженного фосфатидилсерина (DMPS)
анионные фосфолипиды служат высокоаффинными рецепторами для ионов Gd3+, а ионно-индуцированное
уплотнение вызывает повышение латерального давления, оцениваемое в десятки мН/м. Это давление может
«сжать» канал и сместить равновесие в сторону закрытого состояния.
34.
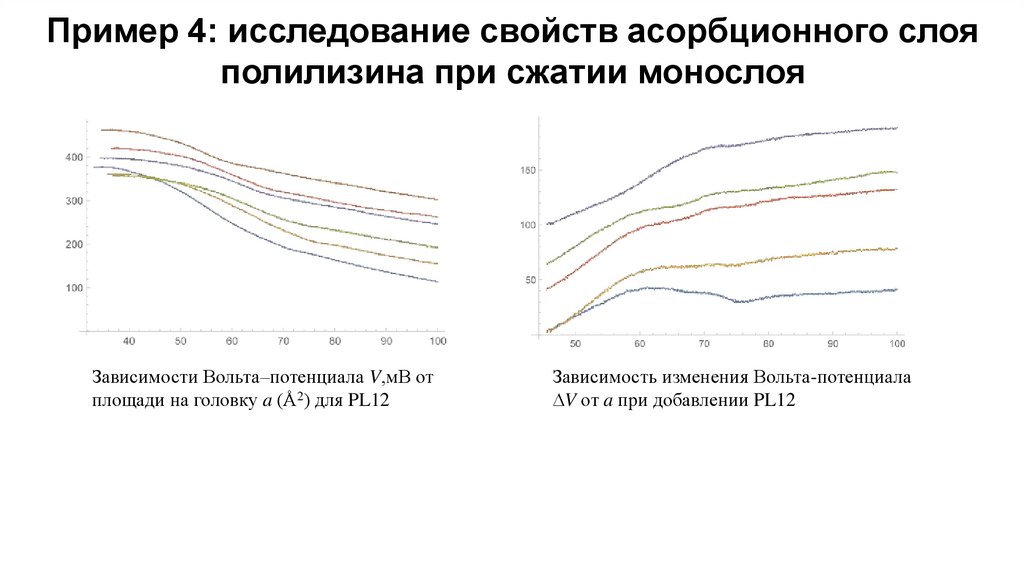
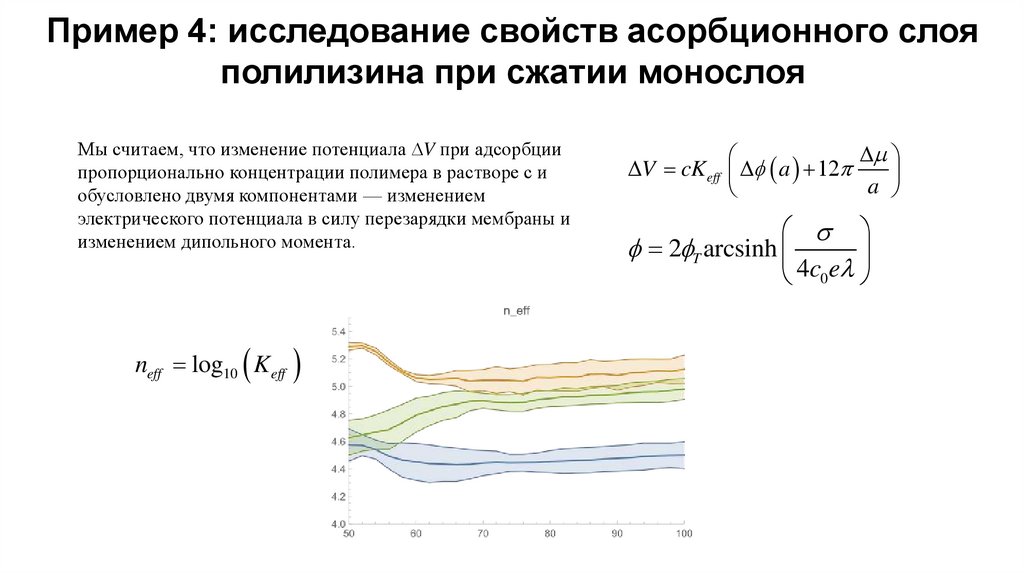
Пример 4: исследование свойств асорбционного слояполилизина при сжатии монослоя
Зависимости Вольта–потенциала V,мВ от
площади на головку a (Å2) для PL12
Зависимость изменения Вольта-потенциала
V от a при добавлении PL12
35.
Пример 4: исследование свойств асорбционного слояполилизина при сжатии монослоя
Мы считаем, что изменение потенциала V при адсорбции
пропорционально концентрации полимера в растворе с и
обусловлено двумя компонентами — изменением
электрического потенциала в силу перезарядки мембраны и
изменением дипольного момента.
neff log10 K eff
V cK eff f a 12
a
f 2fT arcsinh
4c0 e