





















 biology
biologySimilar presentations:
")



")





Метаболизм белков и аминокислот
1. МЕТАБОЛИЗМ БЕЛКОВ И АМИНОКИСЛОТ
2.
Гидролитическое расщепление белков иполипептидов, поступающих вместе с пищей,
происходит под действием протеолитических
специфических и неспецифических ферментов
(протеиназ и пептидаз):
• Эндо- и экзопептидазы;
• Амино- и карбоксипептидазы;
• Сериновые, цистеиновые и др. пептидазы
Образующиеся аминокислоты поступают в кровь и
переносятся к различным органам и тканям.
3.
Внутриклеточный протеолиз осуществляетсяразличными лизосомальными протеазами,
амино- и карбоксипептидазами, дипептидазами.
Большинство внутриклеточных протеолитических
ферментов заключено в протеасомы.
4.
⅔ аминокислот, поступающих в клетки иобразующихся в процессе внутриклеточного
протеолиза, вовлекаются в биосинтез белка.
Остальные подвергаются катаболизму.
Основными катаболическими
превращениями аминокислот являются:
• дезаминирование;
• трансаминирование;
• декарбоксилирование.
5.
Продуктами декарбоксилированияаминокислот являются биогенные амины:
• гистамин (продукт декарбоксилирования
гистидина),
• тирамин (из тирозина),
• кадаверин (из лизина),
• -аминомасляная кислота (из глутамата),
• этаноламин (из серина),
• дофамин (из тирозина),
• серотонин (из окситрипрофана) и др.
6.
Декарбоксилирование аминокислотнеобратимый ферментативный процесс,
катализируемый декарбоксилазами
аминокислот.
Кофактор декарбоксилаз аминокислот –
пиридоксальфосфат.
7.
Дезаминирование аминокислот – отщеплениеα-аминогруппы – может происходить
различными путями:
восстановительное,
гидролитическое,
внутримолекулярное (элиминирующее),
окислительное.
Основным типом является окислительное
дезаминирование.
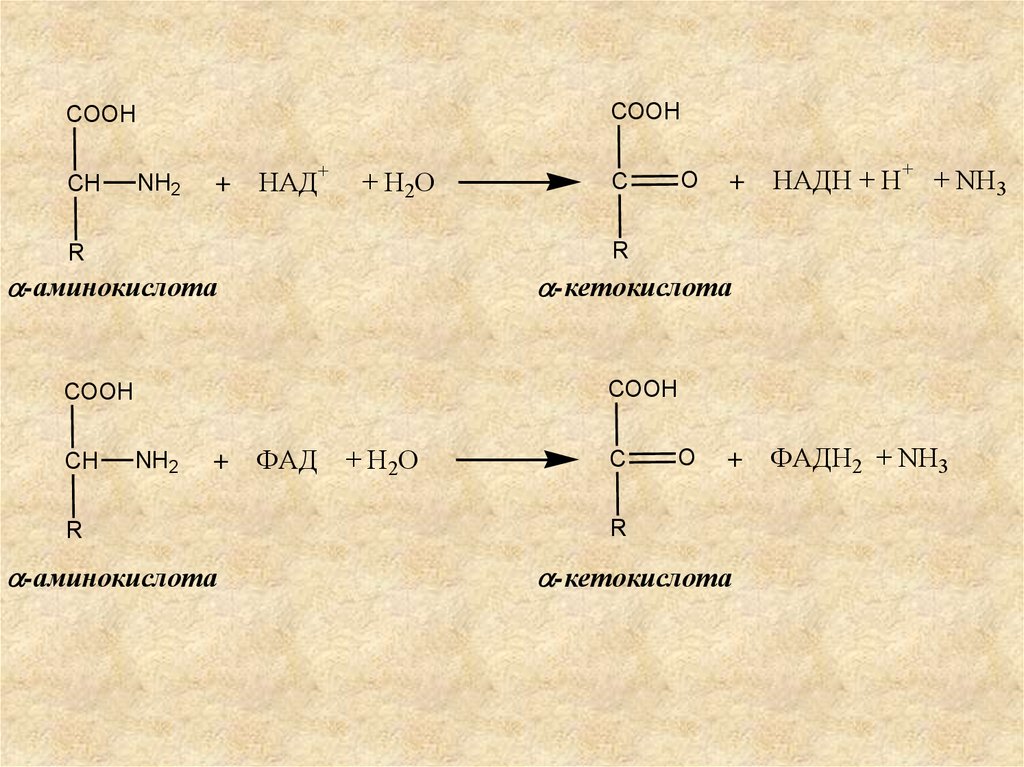
8.
Окислительное дезаминирование катализируется:- НАД-зависимыми дегидрогеназами
аминокислот;
- ФАД (ФМН)-зависимыми оксидазами
аминокислот.
Продукты окислительного дезаминирования –
α-кетокислоты.
9.
COOHCOOH
CH
NH2
+
НАД+
+ Н2О
O
+
НАДН + H+ + NH3
R
R
-аминокислота
-кетокислота
COOH
COOH
CH
C
NH2
+
ФАД
+ Н 2О
C
O
+
ФАДН2 + NH3
R
R
-аминокислота
-кетокислота
ТРАНСАМИНИРОВАНИЕ АМИНОКИСЛОТ
10.
ТРАНСАМИНИРОВАНИЕ АМИНОКИСЛОТТрансаминирование (переаминирование)
аминокислот – реакция межмолекулярного
переноса аминогруппы от -аминокислоты
на -кетокислоту без промежуточного
образования аммиака.
Ферменты: аминотрансферазы
(трансаминазы)
Кофермент: пиридоксальфосфат
11.
COOHCOOH
CH
R1
NH2
-аминокислота
α-аминок-таs
+
C
R2
COOH
O
-кетокислота
α-кеток-таs
C
COOH
O
R1
α-кеток-тар
+
CH
NH2
R2
α-аминок-тар
α-аминокислотаs + пиридоксальфосфат-Е →
→ α-кетокислотаp + пиридоксаминфосфат-Е
α-кетокислотаs + пиридоксаминфосфат-Е →
→ α-аминокислотаp + пиридоксальфосфат-Е
12.
Аммиак, образующийся при дезаминировании,используется:
• для синтеза заменимых аминокислот –
восстановительное аминирование;
• для синтеза азотсодержащих соединений.
Избыточный аммиак – продукт катаболизма –
должен быть инактивирован и выведен
из организма.
13.
ТИПЫ АЗОТИСТОГО ОБМЕНАв зависимости от формы выведения аммиака
Аммониотелический тип у водных
животных. Конечный продукт – аммиак,
выделяющийся непосредственно в воду.
Уреотелический тип у наземных
позвоночных. Конечный продукт – мочевина.
Урикотелический тип у рептилий и
птиц. Конечный продукт – мочевая кислота.
14.
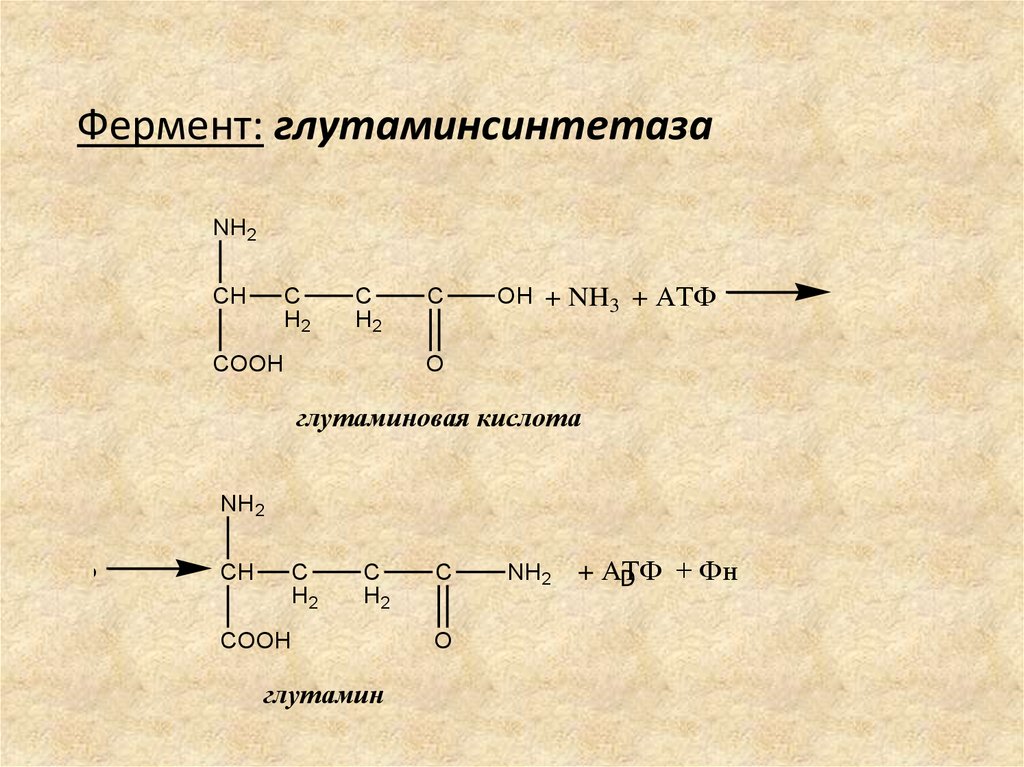
БИОСИНТЕЗ ГЛУТАМИНАОбразование амидов (глутамина и аспарагина) –
процесс первичного связывания аммиака в клетках.
Биосинтез глутамина – наиболее распространенный
путь связывания и обезвреживания аммиака в
организме.
Глутамин – нетоксичная форма транспортировки и
хранения аммиака.
Фермент: глутаминсинтетаза
15.
3Фермент: глутаминсинтетаза
NH2
NH2
CH
C
H2
C
H2
COOH
C
OH + NH3 + АТФ
O
NH2
CH
C
H2
C
H2
COOH
глутамин
C
O
C
H2
C
H2
COOH
глутаминовая кислота
+ АТФ
CH
NH2
+ АТФ
+ Фн
D
глутамин
16.
ОРНИТИНОВЫЙ ЦИКЛ МОЧЕВИНООБРАЗОВНИЯ1. Синтез карбамоилфосфата
Фермент: карбамоилфосфат-синтетаза
O
NH4+ + СО2 + 2АТФ + Н2О
O
Н2 О
H2N
C
H2N
C
O PO3H2
карбамолилфосфат
O PO3H2
+ 2АДФ + Фн
карбамолилфосфат
Синтез карбамоилфосфата происходит в митохондриях
клеток печени. Донор азота только аммиак
(а не амины и другие азотсодержащие соединения).
17.
2. Образование цитруллинаH N
CO
Фермент: орнитинкарбамоилтрансфераза
2
O
H2N
H2N
C
O PO3H2 +
C
H2
C
H2
CH2
CH
HN
C
H2
NH2
COOH
орнитин
орнитин
H2 N
C
H2
нитин
CH2
CH
CO
HN
NH2
COOH
цитр
C
H2
C
H2
CH2
CH
+ Фн
NH2
COOH
цитруллин
цитруллин
18.
3. Взаимодействие цитруллина с аспарагиновой кислотойФермент: аргининосукцинатсинтетаза
H2N
CO
HN
CH2 C
NH2
C C CH2
H2 H2
+
CH NH2
COOH
цитруллин
CH COOH
HN
+ АТФ
HN
H2C COOH
аспартат
H COOH
цитруллин
HN C N
COOH
аспартат
+ АМФ
CH2
+ ФФн
H
+ АТФ
COOH
CH
HN
C
H2
C
H2
CH NH2
COOH
аспартат
C
H2
CH
C
H2
C
C
C
C
CH2 COOH
H2
C N
H
аргининосукцинат
аргининосукцинат
+ АМФ + ФФн
аргининосукцин
19.
4. Образование аргининаФермент: аргининосукцинатлиаза
CH2 COOH
HN
C N
H
HN
C
H2
CH
C
H2
COOH
CH2
HN
C
HN
NH2
C
H2
C
H2
CH NH2
CH NH2
CH
COOH
+
HC
COOH
COOH
COOH
аргининосукцинат
CH2
аргинин
фумарат
20.
5. Гидролиз аргинина с образованием мочевиныФермент: аргиназа
HN
C NH2
H2N
HN
C
H2
C
H2
CH2
+ H2O
C
H2
C
H2
CH2
CH NH2
H2N
C
+
O
CH NH2
COOH
COOH
аргининаргинин
орнитин
орнитин
H2N
мочевина
мочевина
21.
Суммарное уравнение мочевинообразованияСО2 + NH3 + аспартат + 3АТФ + 2Н2О
мочевина + фумарат + 2АДФ +АМФ + 2Фн + ФФн
22. БИОСИНТЕЗ АМИНОКИСЛОТ
Аминокислоты, образующиеся при гидролизебелков:
2/3 расходуются на синтез белка;
1/3 катаболизируются.
Т.е. 1/3 аминокислот должна синтезироваться вновь.
23.
БИОСИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТЗаменимые аминокислоты:
ала, асн, асп, гли, глн, глу, про, сер, тир, цис
Незаменимые аминокислоты:
вал, иле, лей, лиз, мет, тре, три, фен, арг, гис
Углеродный скелет образуется из промежуточных
метаболитов:
гликолиза,
пентозомонофосфатного пути
цикла Кребса.
24.
Пути синтеза:
прямое аминирование α-кетокислот или
ненасыщенных карбоновых кислот;
переаминирование;
взаимопревращение аминокислот.