
























 biology
biologySimilar presentations:




")




")
Механизмы регуляции ферментативной активности
1.
СЗГМУ им. И. И. МЕЧНИКОВАКАФЕДРА БИОЛОГИЧЕСКОЙ И ОБЩЕЙ ХИМИИ
БИОЛОГИЧЕСКАЯ ХИМИЯ-БИОХИМИЯ ПОЛОСТИ РТА
Раздел II
ЛЕКЦИЯ
МЕХАНИЗМЫ РЕГУЛЯЦИИ ФЕРМЕНТАТИВНОЙ
АКТИВНОСТИ.
Доцент Антонова Ж.В.
2022/2023
2.
План лекции1. Способы изменения ферментативной активности. Ингибирование активности ферментов: обратимое и необратимое.
2. Обратимое ингибирование активности ферментов. 2.1. Обратимое конкурентное ингибирование.
2.2. Обратимое неконкурентное ингибирование. 2.3.Смешанное и бесконкурентное ингибирование.
3. Действие конкурентных обратимых ингибиторов на примере фермента СДГ.
4. Антиметаболиты, как лекарственные препараты.
5. Действия конкурентных обратимых ингибиторов на примере ферментов холинэстераз. 5.1.Локализация, биологическая
роль холинэстераз. 5.2. Конкурентные обратимые ингибиторы АХЭ. 5.3.Лекарственные препараты, как конкурентные
обратимые ингибиторы АХЭ.
6. Необратимое ингибирование активности ферментов.
6.1 Необратимое специфическое ингибирование на примере АХЭ. Действие ФОС: инсектицидов, БОВ НПД,
лекарственных препаратов. 6.1.1. Антидоты/реактиваторы АХЭ. Механизм действия оксимов (пралидоксима,
дипироксима, аллоксима).
6.2. Необратимое неспецифическое ингибирование на примере тиоловых ферментов дегидрогеназ.. Механизм действия
«тиоловых ядов». 6.2.1.Антидоты/реактиваторы/протекторы для тиоловых ферментов, механизм действия.
7. Активация ферментов. 7.1. Активация ферментов путем частичного (ограниченного) протеолиза. 7.2. Действие
протекторов. 7.3. Активация ионами металлов.
8. Регуляция активности ферментов. 8.1. Медленная регуляция.
8.2. Быстрая регуляция активности ферментов. 8.2.1. Аллостерическая регуляция вторичными посредниками и
конечными метаболитами по принципу отрицательной обратной связи.
8.2.2. Регуляция путем ковалентной модификации. 8.2.3. Регуляция путем белок-белкового взаимодействия.
3.
Условные обозначенияАХЭ- ацетилхолинэстераза
ХЭ - холинэстераза
АЦ – активный центр
БАЛ- британский антилюизит, антидот
БОВ – боевые отравляющие вещества
КНД – кожно-нарывное действие
НПД – нервно-паралитическое действие
ДФФ – диизопропилфторфосфат
СДГ – сукцинатдегидрогеназа
цАМФ- циклический аденозинмонофосфат
КоЕ - кофермент
4.
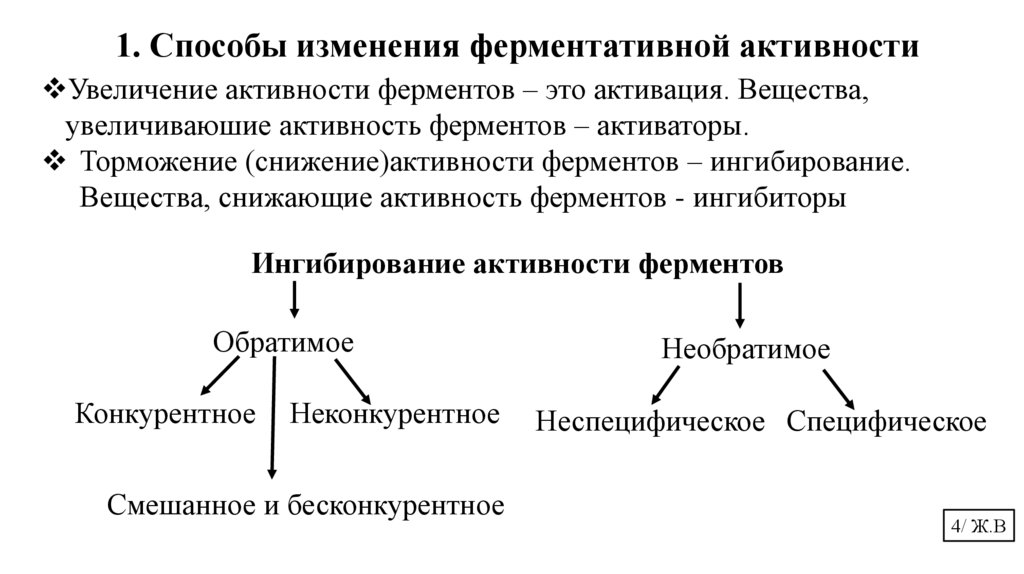
1. Способы изменения ферментативной активностиУвеличение активности ферментов – это активация. Вещества,
увеличиваюшие активность ферментов – активаторы.
Торможение (снижение)активности ферментов – ингибирование.
Вещества, снижающие активность ферментов - ингибиторы
Ингибирование активности ферментов
Обратимое
Конкурентное
Неконкурентное
Смешанное и бесконкурентное
Необратимое
Неспецифическое Специфическое
4/ Ж.В
5.
2. Обратимое ингибирование активности ферментовИнгибитор (I) связывается с ферментом (Е) нековалентными связями и
может легко отделяться от Е при определенных условиях.
2.1. Обратимое конкурентное ингибирование
• I и S – являются структурными аналогами (похожи)
• I и S конкурируют за связывание в АЦ фермента (возможны ЕS и ЕI)
• I связывается с АЦ фермента за счет сил слабого взаимодействия
• I и Е формируют ЕI – комплекс
• ЕI продукт не образует
• Выраженность ингибирования определяется соотношением [I] / [S]
• I может быть вытеснен из АЦ фермента большим количеством S (противоядие)
• Влияние на кинетические параметры: Km , Vmax = const
• Для конкурентного обратимого ингибирования справедливы два уравнения:
Е+S
ЕS
Е+Р
Е+I
ЕI
• После удаления I (диализ, др. виды очистки) активность Е восстанавливается.
5/ Ж.В
6.
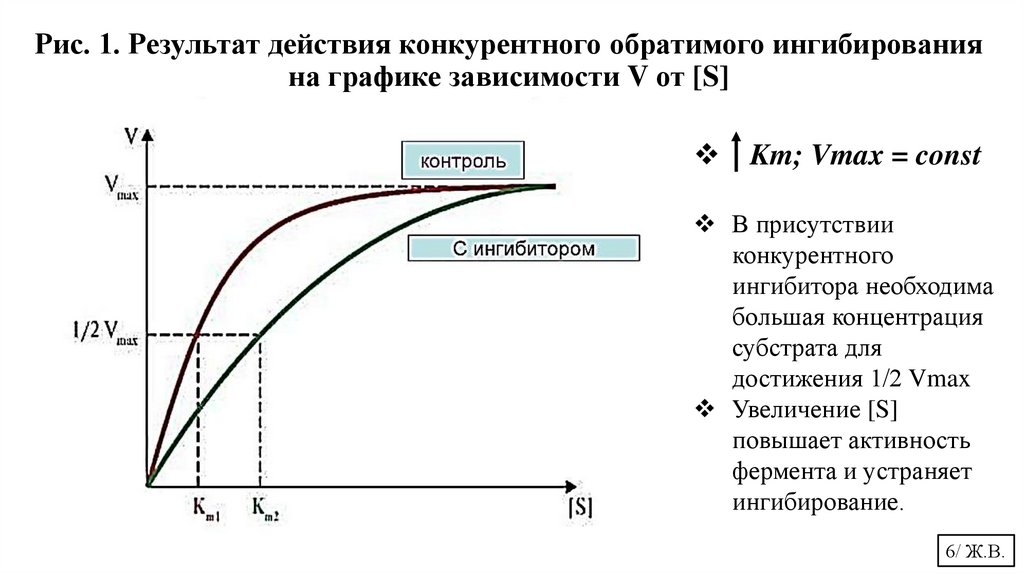
Рис. 1. Результат действия конкурентного обратимого ингибированияна графике зависимости V от [S]
Km; Vmax = const
В присутствии
конкурентного
ингибитора необходима
большая концентрация
субстрата для
достижения 1/2 Vmax
Увеличение [S]
повышает активность
фермента и устраняет
ингибирование.
6/ Ж.В.
7.
2.2.Обратимое неконкурентное ингибирование активности ферментов• I и S – не обладают структурным сходством
• I и S не конкурируют за связывание в АЦ фермента (возможны ЕS, ЕI, ЕSI)
• I связывается с ферментом вне АЦ за счет сил слабого взаимодействия
• I может связаться как с Е (ЕI – комплекс), так и с ES (ЕSI- комплекс)
• ЕSI и ЕI продукты не образуют
• Ингибирование нельзя уменьшить путем простого увеличения концентрации S
• Влияние на кинетические параметры: Km = const, Vmax
• Для неконкурентного обратимого ингибирования справедливы уравнения:
Е+S
ЕS
Е+Р
Е+I
ЕI
ЕS + I
SЕI
• После удаления I (диализ, др. виды очистки) активность Е восстанавливается.
7/Ж.В.
8.
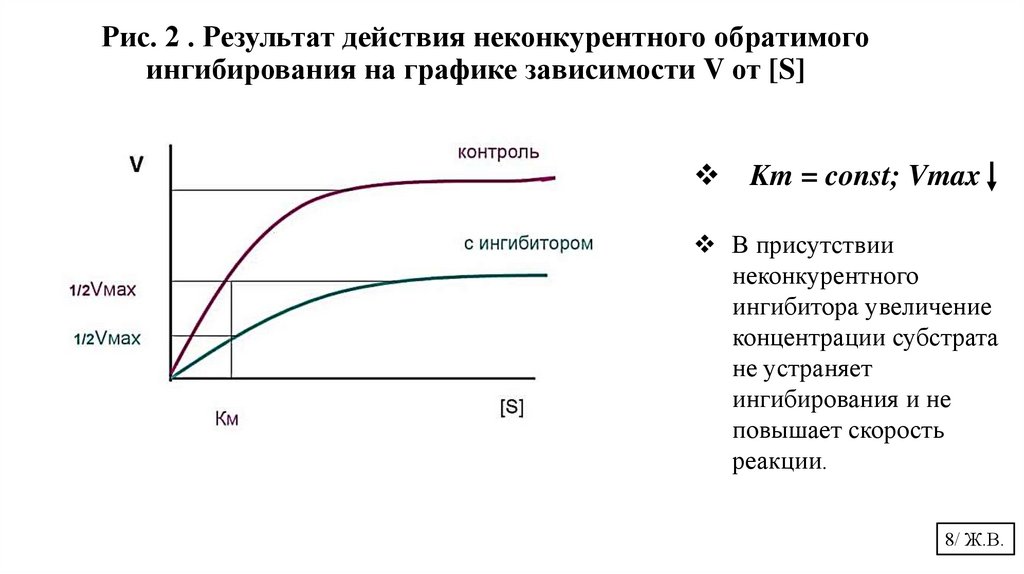
Рис. 2 . Результат действия неконкурентного обратимогоингибирования на графике зависимости V от [S]
Km = const; Vmax
В присутствии
неконкурентного
ингибитора увеличение
концентрации субстрата
не устраняет
ингибирования и не
повышает скорость
реакции.
8/ Ж.В.
9.
2.3. Обратимое смешанное и бесконкурентное ингибирование активности Е.Неконкурентное ингибирование в чистом виде встречается редко. Чаще
наблюдается смешанное ингибирование (частично конкурентное), когда под
действием ингибитора снижается Vmax, а Km – возрастает. Ингибитор может
связаться как с АЦ, так и вне его.
Бесконкурентное ингибирование. Ингибитор связывается вне АЦ только с ЕS
комплексом (ЕSI), изменяя его конформацию. ЕSI – не образует Р
Е+S
ЕS
Е+Р
ЕS + I
ЕSI
• При повышении концентрации субстрата действие бесконкурентного ингибитора
не снижается, так же, как и в случае неконкурентного ингибирования.
9/ Ж.В
10.
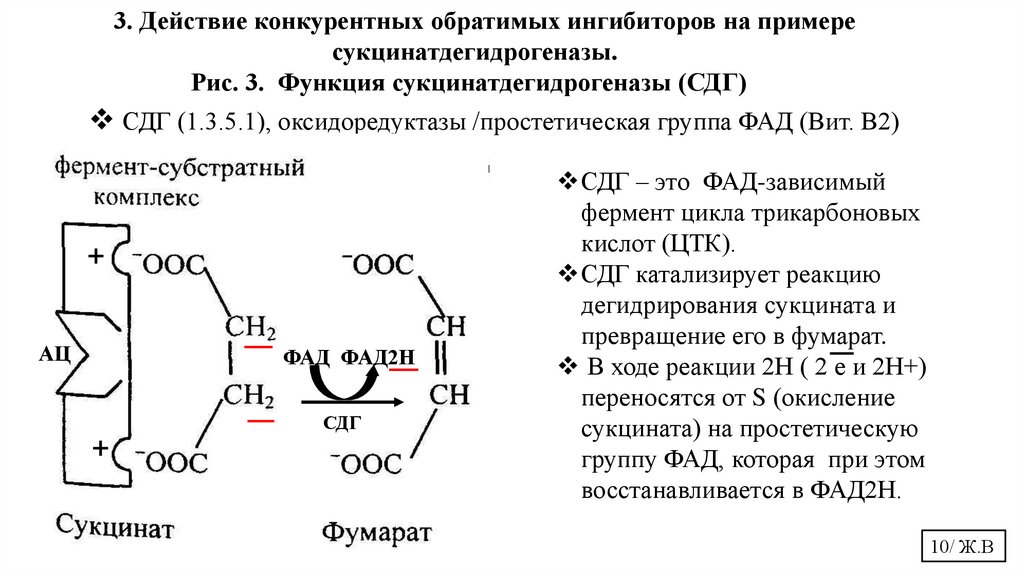
3. Действие конкурентных обратимых ингибиторов на примересукцинатдегидрогеназы.
Рис. 3. Функция сукцинатдегидрогеназы (СДГ)
СДГ (1.3.5.1), оксидоредуктазы /простетическая группа ФАД (Вит. В2)
+
АЦ
ФАД ФАД2Н
СДГ
+
СДГ – это ФАД-зависимый
фермент цикла трикарбоновых
кислот (ЦТК).
СДГ катализирует реакцию
дегидрирования сукцината и
превращение его в фумарат.
В ходе реакции 2Н ( 2 е и 2Н+)
переносятся от S (окисление
сукцината) на простетическую
группу ФАД, которая при этом
восстанавливается в ФАД2Н.
10/ Ж.В
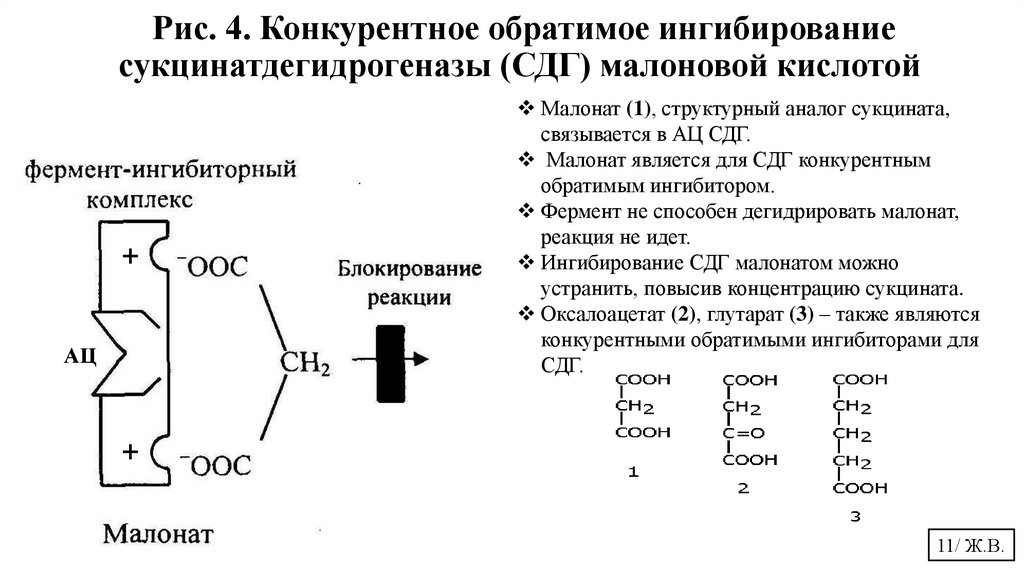
11.
Рис. 4. Конкурентное обратимое ингибированиесукцинатдегидрогеназы (СДГ) малоновой кислотой
+
АЦ
Малонат (1), структурный аналог сукцината,
связывается в АЦ СДГ.
Малонат является для СДГ конкурентным
обратимым ингибитором.
Фермент не способен дегидрировать малонат,
реакция не идет.
Ингибирование СДГ малонатом можно
устранить, повысив концентрацию сукцината.
Оксалоацетат (2), глутарат (3) – также являются
конкурентными обратимыми ингибиторами для
СДГ.
+
11/ Ж.В.
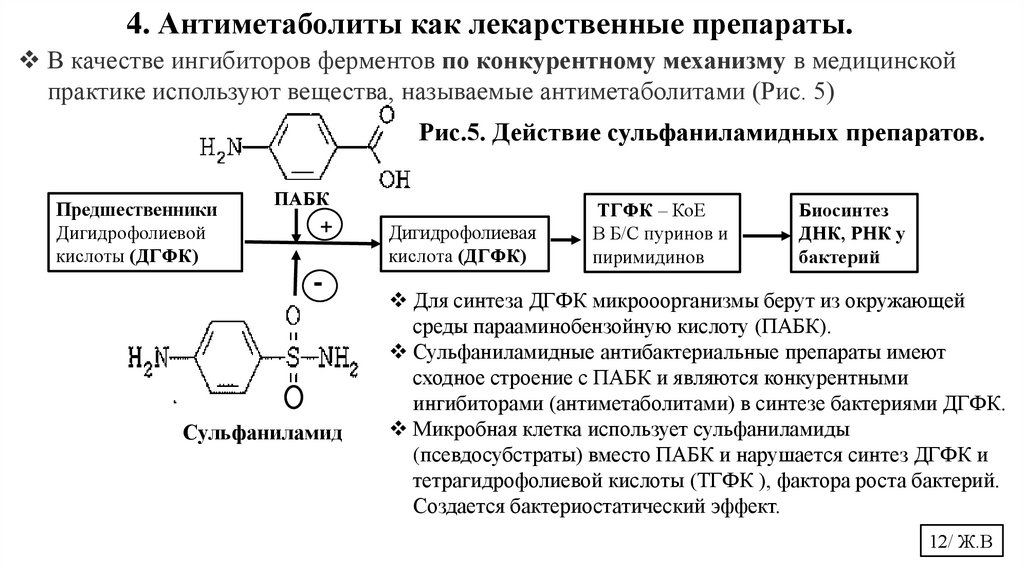
12.
4. Антиметаболиты как лекарственные препараты.В качестве ингибиторов ферментов по конкурентному механизму в медицинской
практике используют вещества, называемые антиметаболитами (Рис. 5)
Рис.5. Действие сульфаниламидных препаратов.
Предшественники
Дигидрофолиевой
кислоты (ДГФК)
ПАБК
+
-
Сульфаниламид
Дигидрофолиевая
кислота (ДГФК)
ТГФК – КоЕ
В Б/С пуринов и
пиримидинов
Биосинтез
ДНК, РНК у
бактерий
Для синтеза ДГФК микрооорганизмы берут из окружающей
среды парааминобензойную кислоту (ПАБК).
Сульфаниламидные антибактериальные препараты имеют
сходное строение с ПАБК и являются конкурентными
ингибиторами (антиметаболитами) в синтезе бактериями ДГФК.
Микробная клетка использует сульфаниламиды
(псевдосубстраты) вместо ПАБК и нарушается синтез ДГФК и
тетрагидрофолиевой кислоты (ТГФК ), фактора роста бактерий.
Создается бактериостатический эффект.
12/ Ж.В
13.
5. Действие конкурентных обратимых ингибиторов на примере ХЭ.5.1. Локализация и биологическая роль холинэстераз.
Различают два типа холинэстераз (ХЭ): ацетилхолинэстеразу (АХЭ, истинная ХЭ,
холинэстераза I, КФ 3.1.1.7) и бутирилхолинэстеразу (БуХЭ, сывороточная ХЭ,
псевдохолинэстераза, холинэстераза II, КФ 3.1.1.8). Ферменты являются простыми
белками, относятся к классу гидролаз и подклассу эстераз.
АХЭ содержится преимущественно в эритроцитах, нервно-мышечных синапсах
скелетной мускулатуры, центральной нервной системе.
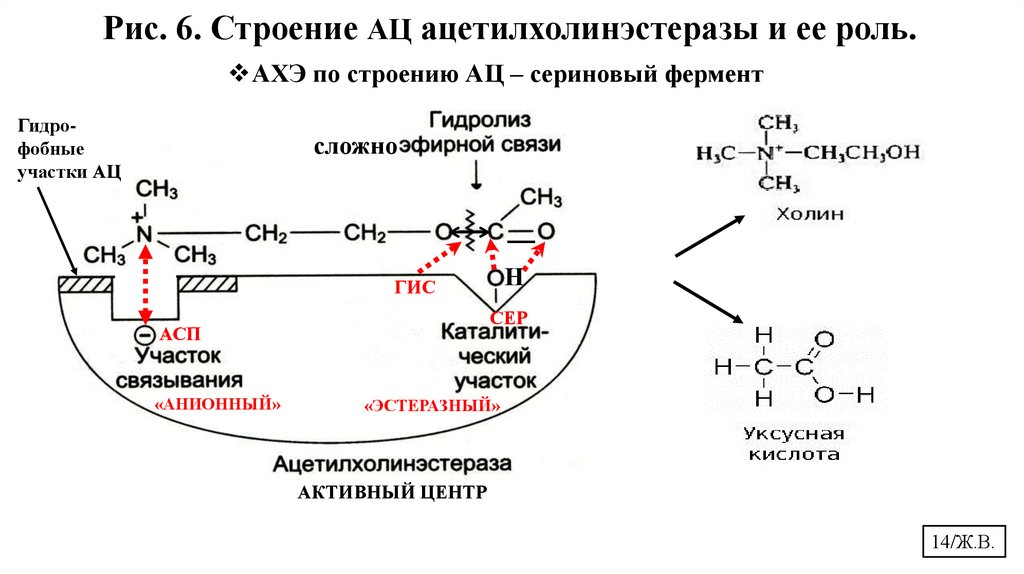
АХЭ катализирует расщепление ацетилхолина на одноатомный азотсодержащий
спирт холин и уксусную кислоту. (Рис.6.)
Ацетилхолин участвует в передаче нервных импульсов в качестве химического
медиатора в синапсах центральной и периферической нервной системы, выделяясь в
синаптическую щель в момент проведения импульса.
13/Ж.В.
14.
Рис. 6. Строение АЦ ацетилхолинэстеразы и ее роль.АХЭ по строению АЦ – сериновый фермент
Гидрофобные
участки АЦ
сложно
Н
ГИС
СЕР
АСП
«АНИОННЫЙ»
«ЭСТЕРАЗНЫЙ»
АКТИВНЫЙ ЦЕНТР
14/Ж.В.
15.
5.1. Локализация и биологическая роль холинэстераз.АХЭ играет ключевую роль в процессах нейрогуморальной и синаптической
передачи: в холинэргических синапсах АХЭ катализирует гидролиз
ацетилхолина и тем самым прекращает воздействие этого медиатора на
холинорецептор, отвечающий за проницаемость постсинаптической мембраны
для ионов.
Его ферментативное расщепление необходимо для подготовки
постсинаптической мембраны к проведению следующего нервного импульса.
АХЭ нервной ткани связана с наружной стороной постсинаптической
мембраны, состоит из однородных гликопротеиновых субъединиц, чаще из
четырех.
Система «ацетилхолин–холинэстераза» участвует в регуляции возбудимости и
сократимости гладкой и сердечной мышц, в осуществлении процессов
активного и пассивного переноса ионов через мембраны.
15/Ж.В.
16.
5.1.Локализация и биологическая роль холинэстераз.
Сывороточная ХЭ (псевдохолинэстераза) находится в сыворотке (плазме) крови,
печени, гладкой мускулатуре, поджелудочной железе, слизистой оболочке кишечника,
легких и других органах.
Сывороточная ХЭ синтезируется в печени и выходит в кровь, поэтому ее содержание
в крови отражает белоксинтезирующую функцию печени. Возможно, что постоянное
присутствие чрезвычайно активной ХЭ в плазме крови является защитным
механизмом и предотвращает распространение по тканям ацетилхолина при его
попадании в кровяное русло.
Показано, что в небольших количествах ХЭ содержится и в центральной нервной
системе. Существует предположение, что роль ХЭ нервной системы сводится к защите
АХЭ от угнетающего действия таких эфиров холина, которые практически не
разрушаются АХЭ, но могут образоваться в организме (пропионилхолин,
бутирилхолин, валерилхолин и др.), а ХЭ их разрушает.
Возможно также, что в отдельных участках мозга могут создаваться ненормально
высокие концентрации ацетилхолина, избыток которого тормозит АХЭ, но не влияет
на активность ХЭ, которая его разрушает.
16/ Ж.В.
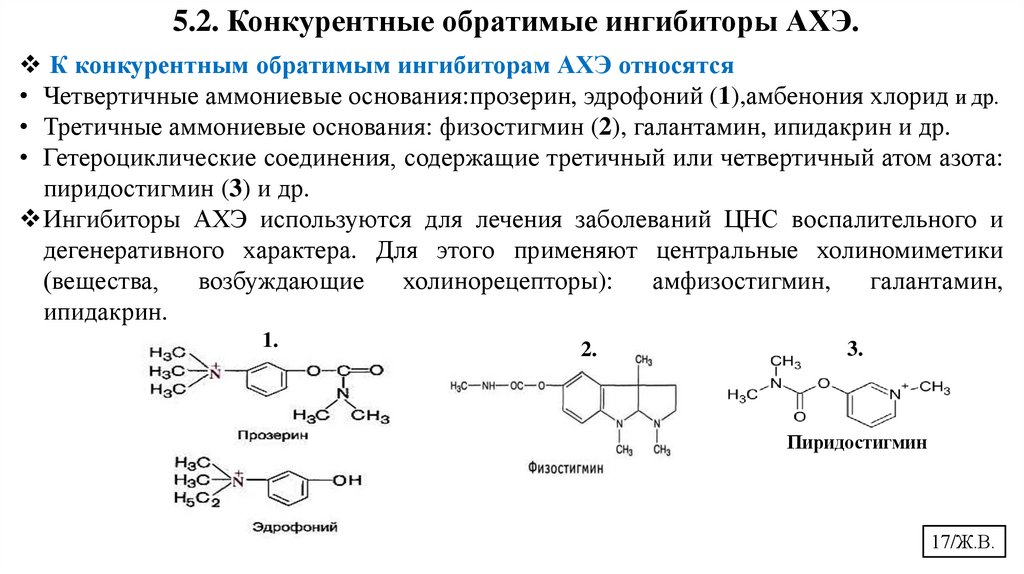
17.
5.2. Конкурентные обратимые ингибиторы АХЭ.К конкурентным обратимым ингибиторам АХЭ относятся
• Четвертичные аммониевые основания:прозерин, эдрофоний (1),амбенония хлорид и др.
• Третичные аммониевые основания: физостигмин (2), галантамин, ипидакрин и др.
• Гетероциклические соединения, содержащие третичный или четвертичный атом азота:
пиридостигмин (3) и др.
Ингибиторы АХЭ используются для лечения заболеваний ЦНС воспалительного и
дегенеративного характера. Для этого применяют центральные холиномиметики
(вещества,
возбуждающие
холинорецепторы):
амфизостигмин,
галантамин,
ипидакрин.
1.
2.
3.
Пиридостигмин
17/Ж.В.
18.
5.3. Лекарственные препараты, как конкурентные обратимыеингибиторы АХЭ.
При миастении (повышенная утомляемость и слабость скелетных мышц
в результате нарушения нервно-мышечной передачи) применяют
ингибиторы
холинэстеразы:
неостигмина
метилсульфат,
пиридостигмина бромид, амбенония хлорид.
Эти лекарственные средства при миастении увеличивают объем
движений, мышечную силу, двигательную активность, жизненную
емкость легких и др.
Ингибиторы холинэстеразы применяют при атонии гладких мышц
пищевода, кишечника, мочевого пузыря и др.
Также ингибиторы холинэстеразы применяют для устранения
миопаралитического действия антидеполяризующих миорелаксантов,
вводят внутривенно галантамин, неостигмина метилсульфат.
Активируют ацетилхолинэстеразу ионы кальция и магния.
18/Ж.В.
19.
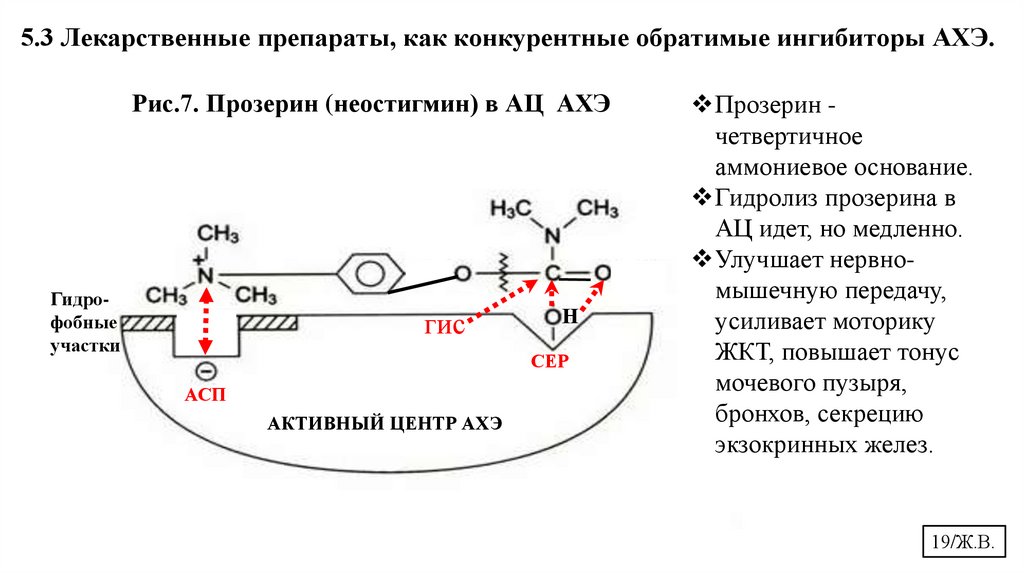
5.3 Лекарственные препараты, как конкурентные обратимые ингибиторы АХЭ.Рис.7. Прозерин (неостигмин) в АЦ АХЭ
Гидрофобные
участки
гис
Н
СЕР
АСП
АКТИВНЫЙ ЦЕНТР АХЭ
Прозерин четвертичное
аммониевое основание.
Гидролиз прозерина в
АЦ идет, но медленно.
Улучшает нервномышечную передачу,
усиливает моторику
ЖКТ, повышает тонус
мочевого пузыря,
бронхов, секрецию
экзокринных желез.
19/Ж.В.
20.
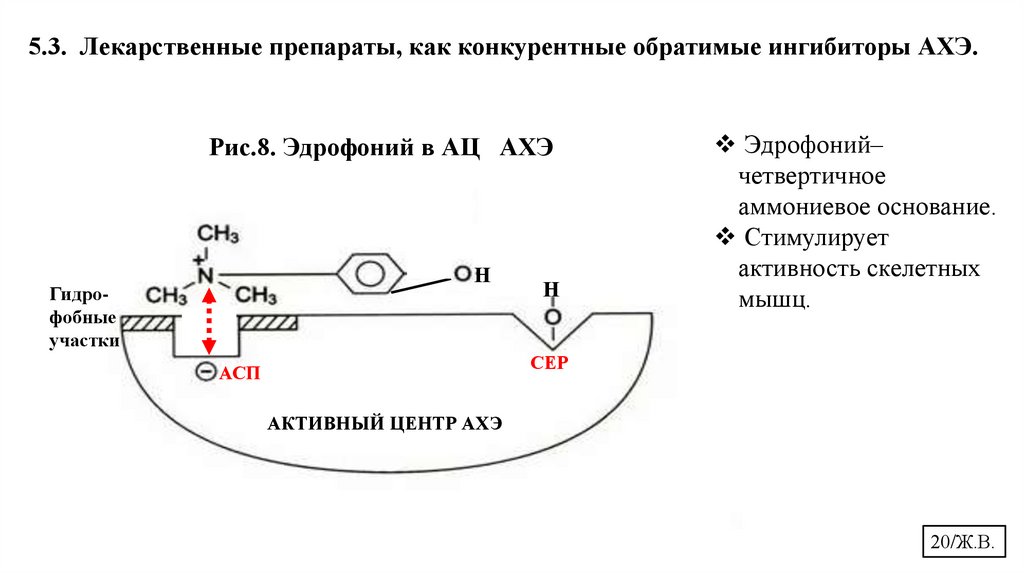
5.3. Лекарственные препараты, как конкурентные обратимые ингибиторы АХЭ.Рис.8. Эдрофоний в АЦ АХЭ
Н
Гидрофобные
участки
Н
Эдрофоний–
четвертичное
аммониевое основание.
Стимулирует
активность скелетных
мышц.
СЕР
АСП
АКТИВНЫЙ ЦЕНТР АХЭ
20/Ж.В.
21.
6. Небратимое ингибирование активности ферментовИнгибитор (I) связывается с ферментом (Е) прочными ковалентными
связями.
6.1. Необратимое специфическое ингибирование активности ферментов.
На примере серинового фермента - АХЭ
К специфическим необратимым ингибиторам АХЭ относятся
фосфорорганические соединения (ФОС) – эфиры кислот фосфора
(фосфорной, пирофосфорной, фосфористой, фосфоновой, фосфиновой,
тио- и дитиофосфорной, тиофосфористой) и их сернистые, азотистые и
другие производные.
21/Ж.В.
22.
6.1. Необратимое специфическое ингибирование активности АХЭФОС применяются как инсектициды (метафос, меркаптофос, хлорофос,
карбофос, фосдрин, лептофос и др.)
ФОС применяются как лекарственные препараты (хлорофтальм,
фосфакол, армин и т. д.)
Наиболее токсичные представители группы ФОС – это боевые
отравляющие вещества нервно-паралитического действия: зоман, табун,
зарин
22/Ж.В.
23.
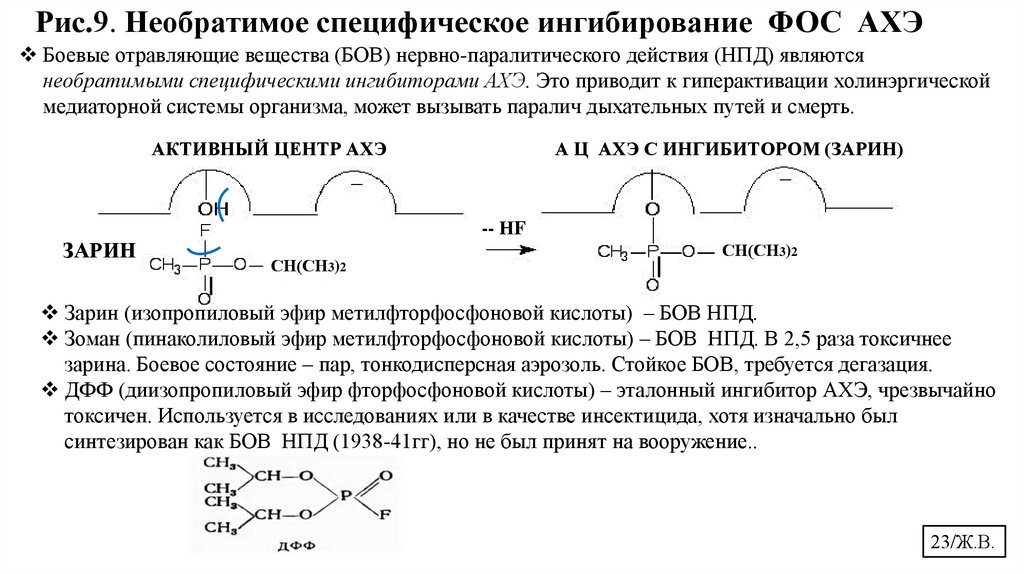
Рис.9. Необратимое специфическое ингибирование ФОС АХЭБоевые отравляющие вещества (БОВ) нервно-паралитического действия (НПД) являются
необратимыми специфическими ингибиторами АХЭ. Это приводит к гиперактивации холинэргической
медиаторной системы организма, может вызывать паралич дыхательных путей и смерть.
АКТИВНЫЙ ЦЕНТР АХЭ
А Ц АХЭ С ИНГИБИТОРОМ (ЗАРИН)
-- НF
ЗАРИН
СН(СН3)2
СН(СН3)2
Зарин (изопропиловый эфир метилфторфосфоновой кислоты) – БОВ НПД.
Зоман (пинаколиловый эфир метилфторфосфоновой кислоты) – БОВ НПД. В 2,5 раза токсичнее
зарина. Боевое состояние – пар, тонкодисперсная аэрозоль. Стойкое БОВ, требуется дегазация.
ДФФ (диизопропиловый эфир фторфосфоновой кислоты) – эталонный ингибитор АХЭ, чрезвычайно
токсичен. Используется в исследованиях или в качестве инсектицида, хотя изначально был
синтезирован как БОВ НПД (1938-41гг), но не был принят на вооружение..
23/Ж.В.
24.
6.1.1. Антидоты / реактиваторы АХЭАнтагонисты ФОС, способные восстанавливать активность холинэстераз путем
дефосфорилирования молекулы инактивированного фермента, получили название
реактиваторов холинэстераз.
По характеру своего фармакологического действия они являются антидотами ФОС.
Восстанавливая активность холинэстераз в органах, функция которых была
нарушена в результате прекращения гидролиза синаптического ацетилхолина,
реактиваторы холинэстераз способствуют восстановлению синаптической передачи.
К реактиваторам холинэстераз принадлежат оксимы (RR1C = N–OН),
• содержащие четвертичные ониевые группы (пралидоксим йодид, пралидоксим
хлорид, дипироксим, тримедоксим, аллоксим, токсогонин и др.);
• оксимы, не содержащие четвертичных ониевых групп (моноизонитрозоацетон,
изонитрозин, диэтиксим и др.). При проведении специфической антидотной терапии
ведется постоянный контроль активности холинэстеразы.
Проведение мониторинга ХЭ обязательно для людей, работа которых связана с
производством отравляющих веществ, получением и использованием инсектицидов,
пестицидов и лекарственных препаратов – ингибиторов фермента.
24/Ж.В.
25.
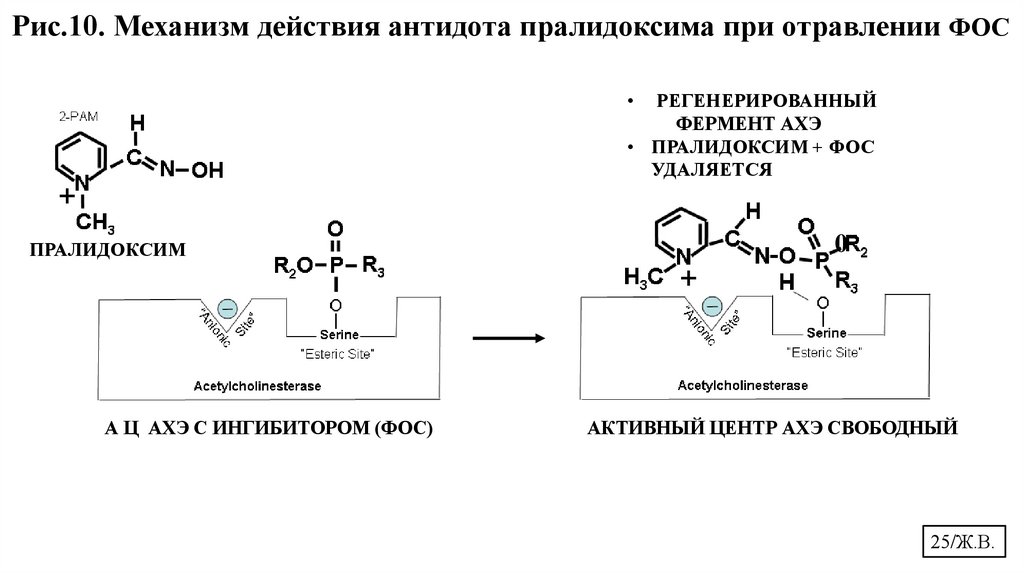
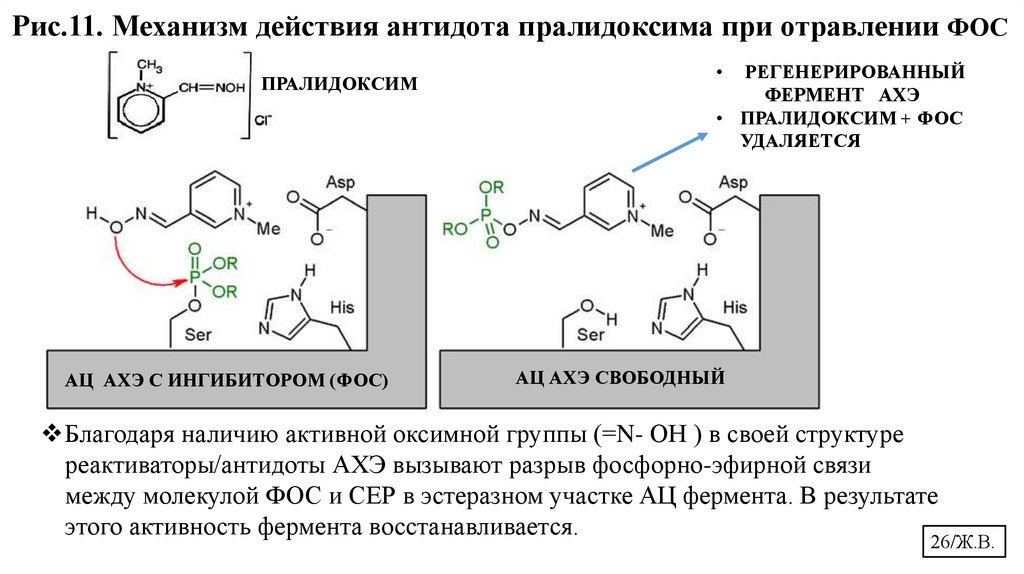
Рис.10. Механизм действия антидота пралидоксима при отравлении ФОСРЕГЕНЕРИРОВАННЫЙ
ФЕРМЕНТ АХЭ
• ПРАЛИДОКСИМ + ФОС
УДАЛЯЕТСЯ
+
0
ПРАЛИДОКСИМ
+
А Ц АХЭ С ИНГИБИТОРОМ (ФОС)
АКТИВНЫЙ ЦЕНТР АХЭ СВОБОДНЫЙ
25/Ж.В.
26.
Рис.11. Механизм действия антидота пралидоксима при отравлении ФОСПРАЛИДОКСИМ
АЦ АХЭ С ИНГИБИТОРОМ (ФОС)
РЕГЕНЕРИРОВАННЫЙ
ФЕРМЕНТ АХЭ
• ПРАЛИДОКСИМ + ФОС
УДАЛЯЕТСЯ
АЦ АХЭ СВОБОДНЫЙ
Благодаря наличию активной оксимной группы (=N- ОН ) в своей структуре
реактиваторы/антидоты АХЭ вызывают разрыв фосфорно-эфирной связи
между молекулой ФОС и СЕР в эстеразном участке АЦ фермента. В результате
этого активность фермента восстанавливается.
26/Ж.В.
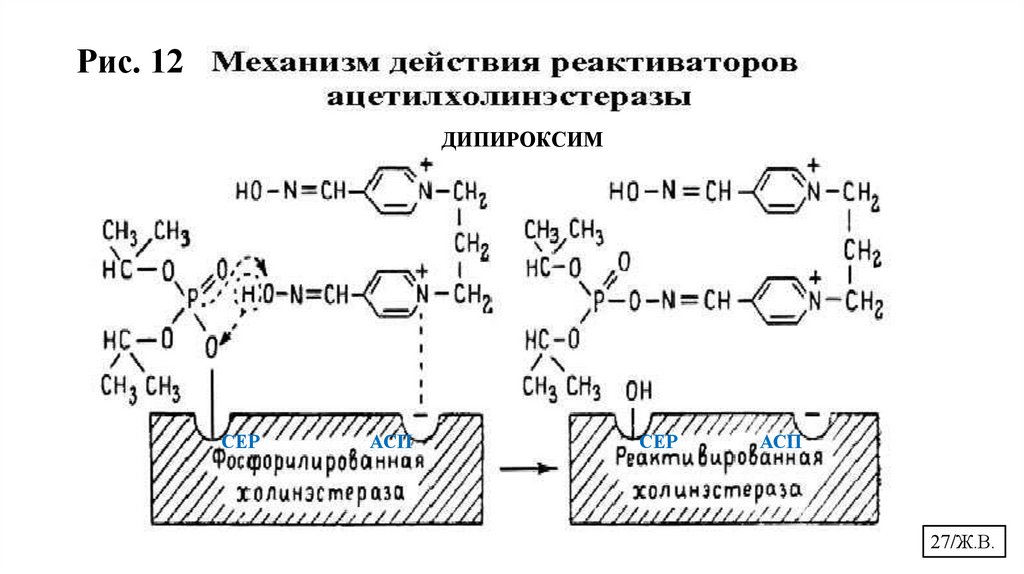
27.
Рис. 12ДИПИРОКСИМ
СЕР
АСП
СЕР
АСП
27/Ж.В.
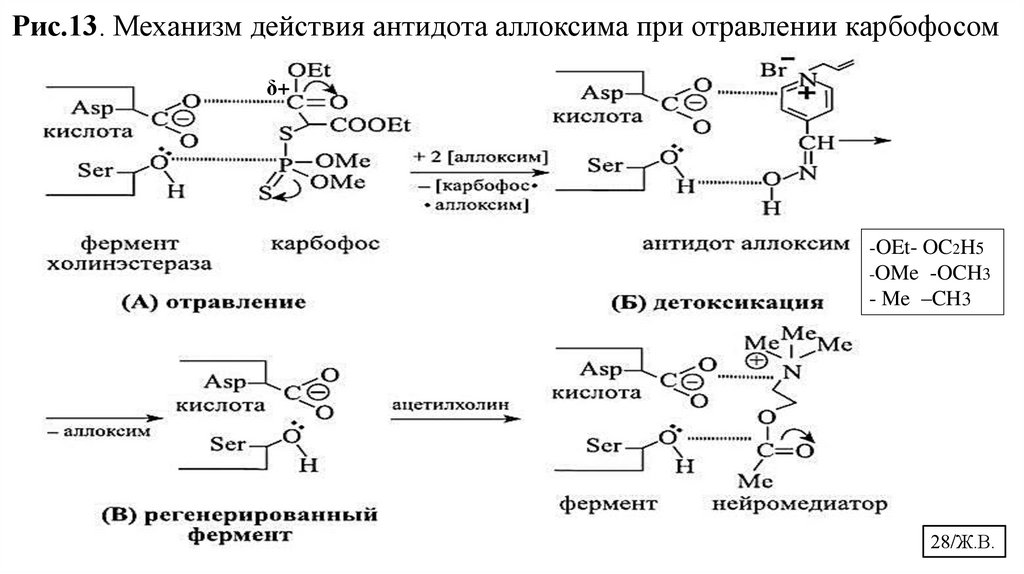
28.
Рис.13. Механизм действия антидота аллоксима при отравлении карбофосомδ+
-OEt- OC2H5
-OMe -OCH3
- Me –CH3
28/Ж.В.
29.
6.2. Необратимое неспецифическое ингибирование ферментов.На примере тиоловых ферментов - дегидрогеназ
Дегидрогеназы относятся к группе «тиоловых ферментов», каталитическое
действие которых обусловлено сульфгидрильными (тиоловыми) НS-группами,
входящими в состав активного центра молекул фермента или ответственными за
поддержание каталитически активной конформации.
Ингибиторами дегидрогеназ являются вещества, блокирующие HS-группы.
К ним относятся окисляющие, меркаптидобразующие, алкилирующие вещества.
Например:
• пероксид водорода (окислитель)
• хлорид ртути и другие соли тяжелых металлов (соли As, Сd, Pb, Ag, Cu –
меркаптидобразующие агенты)
• йодуксусная кислота (алкилирующий агент).
Окислители, тяжелые металлы, алкилирующие агенты могут взаимодействовать с
–SН группами любых белков и ферментов, поэтому относятся к неспецифическим
ингибиторам.
29/Ж.В.
30.
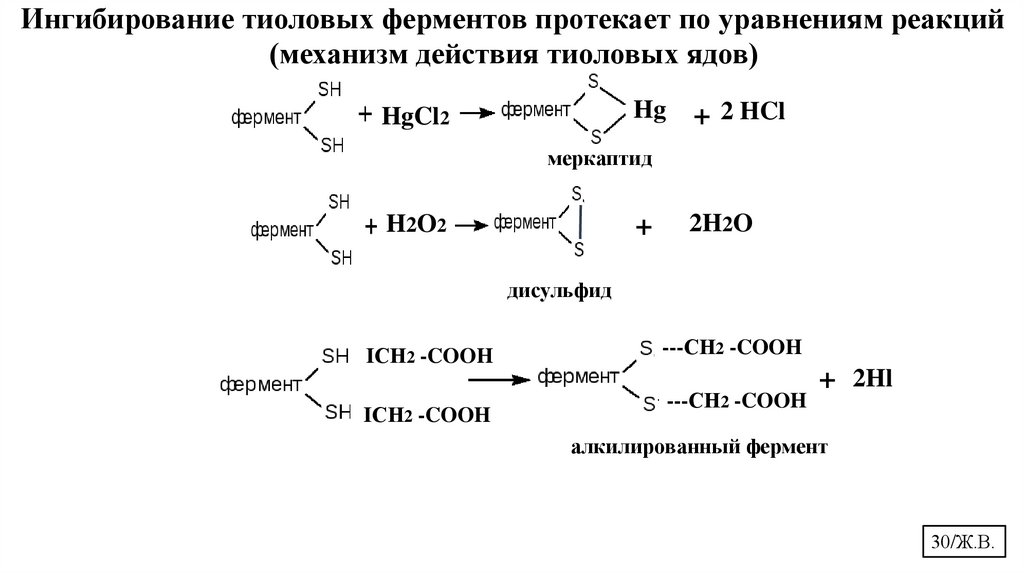
Ингибирование тиоловых ферментов протекает по уравнениям реакций(механизм действия тиоловых ядов)
Hg
HgCl2
+ 2 HCl
меркаптид
++
H 2O2
2H2O
дисульфид
IСН2 -COOH
IСН2 -COOH
---СН2 -COOH
---СН2 -COOH
+ 2Hl
алкилированный фермент
30/Ж.В.
31.
6.2.1.Антидоты для тиоловых ферментов (дегидрогеназ)Необратимые неспецифические ингибиторы тиоловых
ферментов называются «тиоловые яды». К ним относятся:
меркаптообразующие агенты, окислители и алкилирующие агенты.
Антидоты
• Низкомолекулярные (ди)тиолсодержащие вещества для защиты и
реактивации ферментов от действия тяжелых металлов ( БАЛ,
унитиол, дигидролипоевая кислота, димеркаптоянтарная кислота);
пенициламин (монотиоловая группа антидотов);
• Антиоксиданты для защиты от действия окислителей (аскорбиновая
кислота, напр., при отравлении перманганатом калия).
31/ЖВ.
32.
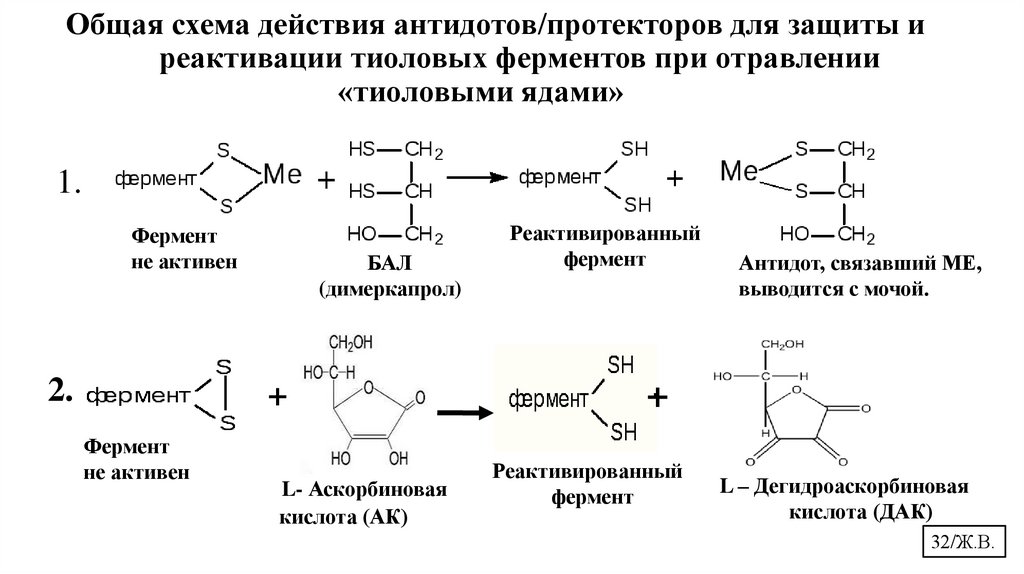
Общая схема действия антидотов/протекторов для защиты иреактивации тиоловых ферментов при отравлении
«тиоловыми ядами»
1.
Фермент
не активен
2.
БАЛ
(димеркапрол)
+
Фермент
не активен
L- Аскорбиновая
кислота (АК)
Реактивированный
фермент
Антидот, связавший МЕ,
выводится с мочой.
+
Реактивированный
фермент
L – Дегидроаскорбиновая
кислота (ДАК)
32/Ж.В.
33.
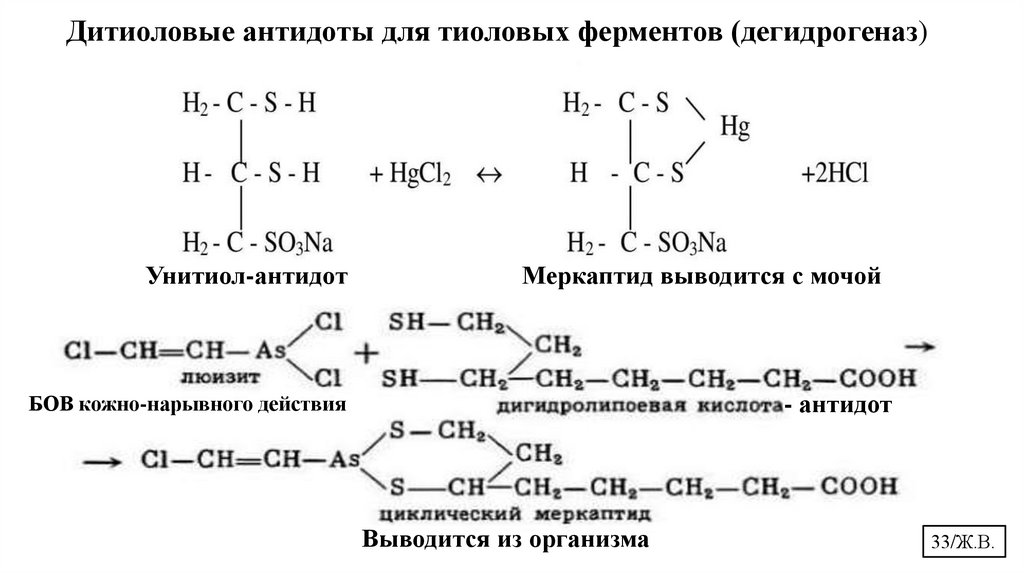
Дитиоловые антидоты для тиоловых ферментов (дегидрогеназ)Унитиол-антидот
Меркаптид выводится с мочой
БОВ кожно-нарывного действия
- антидот
Выводится из организма
33/Ж.В.
34.
Антидоты для тиоловых ферментов (дегидрогеназ)Димеркаптоянтарная кислота
(эффективна при острых
отравлениях мышьяком)
Пенициламин (группа
монотиолов)
Действие тиоловых ядов на человека (гл. образом металлов) проявляется при
профессиональных контактах в промышленности, в разных сферах хозяйственной
деятельности (в чистом виде, в сплавах, в составе органических и неорганических
соединений) и постоянно возрастает в результате антропогенного загрязнения ими
окружающей среды.
34/Ж.В.
35.
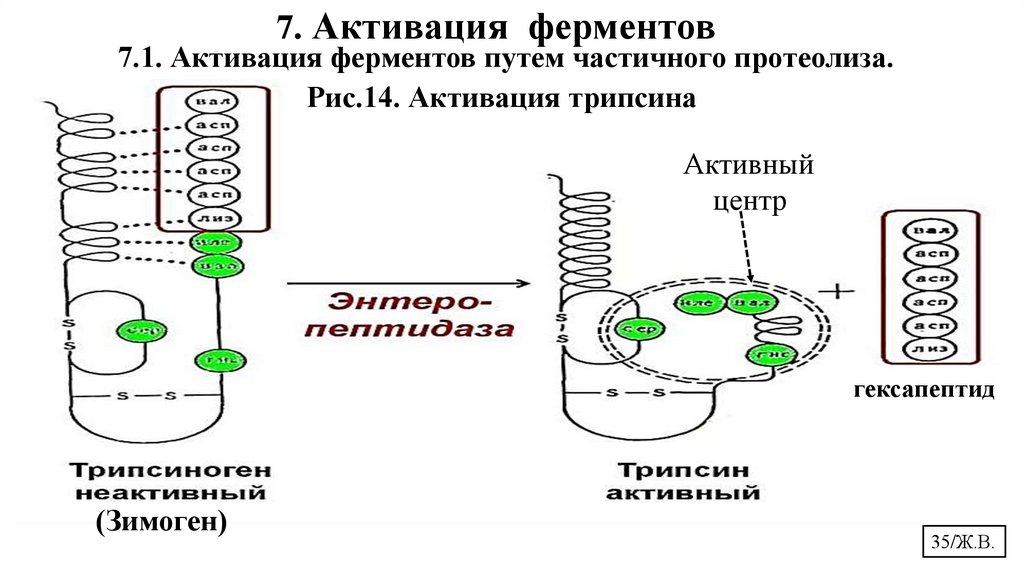
7. Активация ферментов7.1. Активация ферментов путем частичного протеолиза.
Рис.14. Активация трипсина
.
Активный
центр
гексапептид
(Зимоген)
35/Ж.В.
36.
7.1.Активация ферментов путем частичного (ограниченного) протеолиза.Отщепление пептида вызывает необратимые конформационные
перестройки в молекуле фермента, которые приводят к образованию
3-й структуры и формированию АЦ.
Частичный протеолиз лежит в основе активации:
• протеолитических ферментов ЖКТ
• матриксных металлопротеиназ, участвующих в катаболизме белков
межклеточного матрикса соединительных тканей (коллагеназы и др.)
• белков свёртывающей системы крови
• белков системы комплемента
• гормонов белковой природы
• белков, участвующих в минерализации эмали зуба
• белков калликреин-кининовой и ренин-ангиотензивной системы,
участвующих в регуляции сосудистого тонуса и т.д.
36/Ж.В.
37.
7.2. Активация ферментов за счет действия антидотов / протекторов.(См. предыдущие слайды)
Вещества, способные защищать ферменты от инактивации (напр.
«тиоловыми ядами») или возвращать ферментам уже утраченную
активность называются протекторами.
В качестве естественных протекторов в организме могут выступать
трипептид глутатион (Гл-SH), витаминоподобное вещество липоевая
кислота, цистеин, аскорбиновая кислота и др.)
Антидоты и реактиваторы, введенные в организм при отравлениях
тиоловыми ядами, ФОС и др., не только возвращают активность
ферментам, но и защищают активные группы неповрежденных
ферментов от действия ингибиторов (активация путем защиты).
37/Ж.В.
38.
7.3. Активация ферментов ионами металлов• Стабилизируют каталитически активную конформацию фермента.
Ме образуют тройные мостиковые комплексы: Ме-Е-S; Е-S-Ме;
Е-Ме-S;
• Сближают Е и S, способствуют их взаимодействию путем
формирования координационных связей (возможно вызывают
напряжение связей в S в ходе процесса);
• Удерживают реагирующие группы в требуемой трехмерной
ориентации;
• Участвуют в ковалентном катализе;
• Положительный заряд иона может стабилизировать переходное
состояние за счет электростатических взаимодействий.
• Активаторами могут быть: Mg2+, Mn2+, Ca2+ и др.
38/Ж.В.
39.
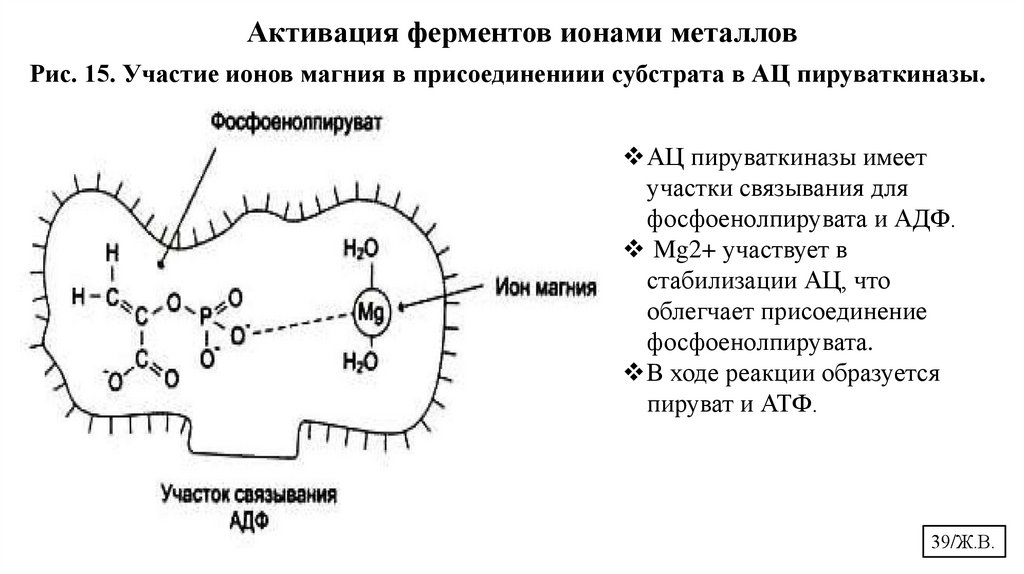
Активация ферментов ионами металловРис. 15. Участие ионов магния в присоединениии субстрата в АЦ пируваткиназы.
АЦ пируваткиназы имеет
участки связывания для
фосфоенолпирувата и АДФ.
Mg2+ участвует в
стабилизации АЦ, что
облегчает присоединение
фосфоенолпирувата.
В ходе реакции образуется
пируват и АТФ.
39/Ж.В.
40.
8. РЕГУЛЯЦИЯ АКТИВНОСТИ ФЕРМЕНТОВБыстрая
Медленная
Изменение активности «имеющихся в
наличии» молекул ферментов
Изменение количества молекул
ферментов
8.1. Медленная регуляция
• Усиление биосинтеза белков- ферментов- это индукция синтеза.
• Индуцибельные ферменты-это ферменты, которые вырабатываются в необходимом
количестве только в присутствии определенных индукторов. Индукторами часто
являются субстраты данного фермента, гормоны, лекарственные препараты,
ксенобиотики и др. К числу индуцибельных ферментов, синтез которых стимулируют
субстраты, относится цитохром Р 450
• Конститутивные ферменты – это определенная часть ферментов, вырабатывающаяся
в организме человека постоянно, скорость синтеза которых избирательно не
регулируется и не зависит от наличия соответствующих субстратов.
• Снижение биосинтеза белков-ферментов – это репрессия синтеза.
40/Ж.В.
41.
8.2. Быстрая регуляцияОсновные способы быстрой регуляции активности ферментов
• Аллостерическая регуляция
• Регуляция путем ковалентной модификации
(фосфорилирования/дефосфорилирования) молекулы фермента
• Регуляция с помощью белок-белковых взаимодействий
8.2.1.Аллостерическая регуляция
Аллостерические ферменты всегда имеют четвертичную структуру и состоят из
нескольких субъединиц (протомеров).
• В одних протомерах находятся регуляторные (аллостерические) центры, в которые
встраиваются аллостерические эффекторы ( конечные продукты метаболических
путей, АТФ, вторичные посредники цАМФ, цГМФ и др. Рис. 16.)
Такие субъединицы называются регуляторными.
• В других субъединицах- протомерах находятся активные центры для осуществления
химической реакции. Эти протомеры называются каталитическими. (Рис.17)
41/Ж.В.
42.
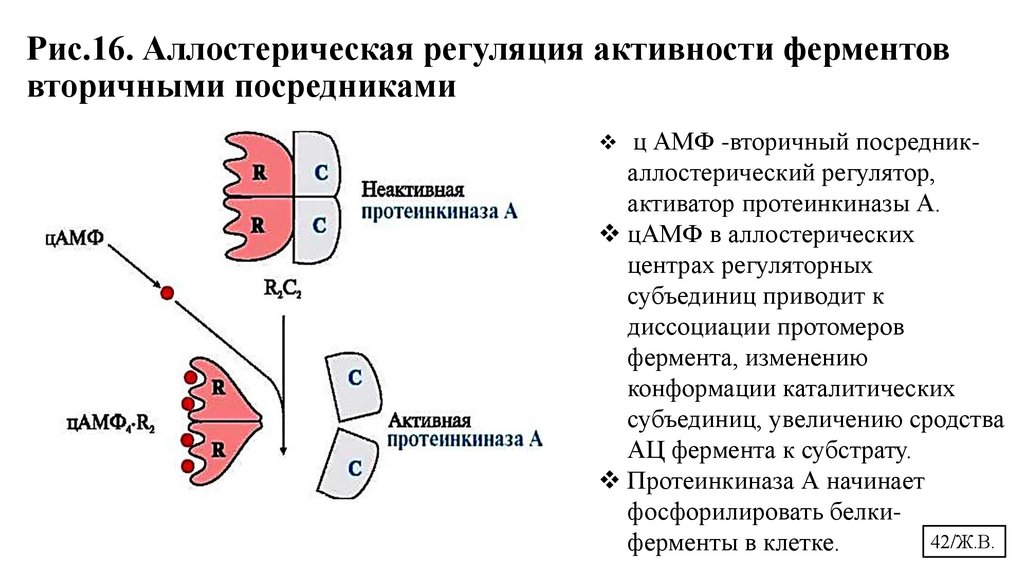
Рис.16. Аллостерическая регуляция активности ферментоввторичными посредниками
ц АМФ -вторичный посредник-
аллостерический регулятор,
активатор протеинкиназы А.
цАМФ в аллостерических
центрах регуляторных
субъединиц приводит к
диссоциации протомеров
фермента, изменению
конформации каталитических
субъединиц, увеличению сродства
АЦ фермента к субстрату.
Протеинкиназа А начинает
фосфорилировать белки42/Ж.В.
ферменты в клетке.
43.
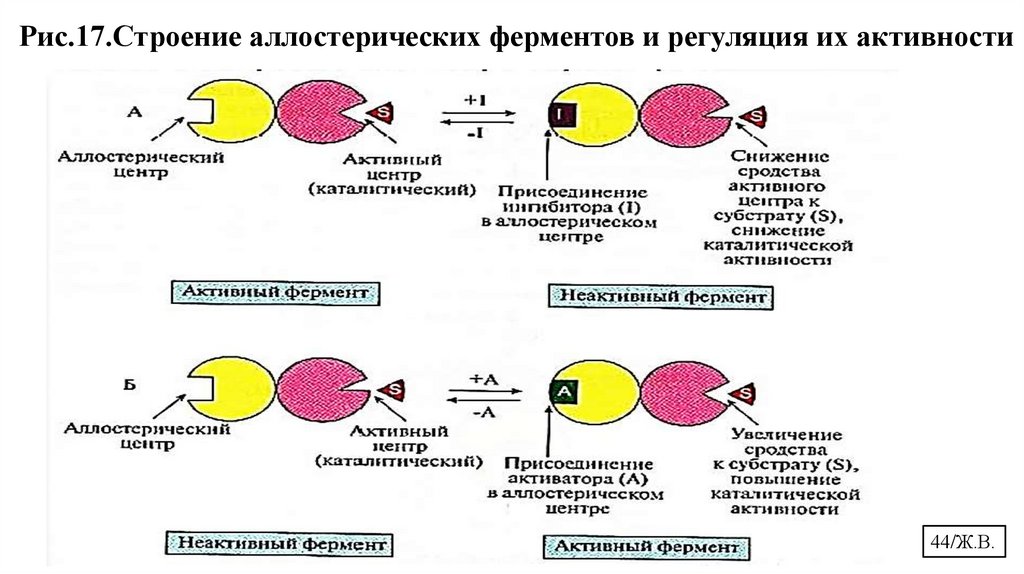
8.2.1. Аллостерическая регуляция• Аллостерические эффекторы связываются с аллостерическими центрами
силами слабого взаимодействия.
• Положительные эффекторы активируют фермент, отрицательные –
ингибируют (Рис. 17).
• Аллостерические ферменты обладают свойством кооперативности.
• Аллостерические ферменты катализируют ключевые реакции
метаболических путей.
• Аллостерическая регуляция – обратима.
• Аллостерические центры проявляют различную специфичность к
лигандам.
Аллостерическая регуляция активности первого фермента
43/Ж.В.
метаболического пути конечным метаболитом (продуктом) по
принципу отрицательной обратной связи (ретроингибирование)- это
распространенный способ регуляции обмена веществ в клетке (Рис.18)
44.
Рис.17.Строение аллостерических ферментов и регуляция их активности44/Ж.В.
45.
Рис. 18. Аллостерическая регуляция активности ферментовпринципу отрицательной обратной связи (ретроингибирование)
-
Например:
• ГЕМ ингибирует аминолевулинатсинтазу (процесс синтеза гема);
• L –изолейцин ингибирует треониндегидратазу (процесс
образования L-изолейцина из L-треонина) и др.
45/Ж.В.
46.
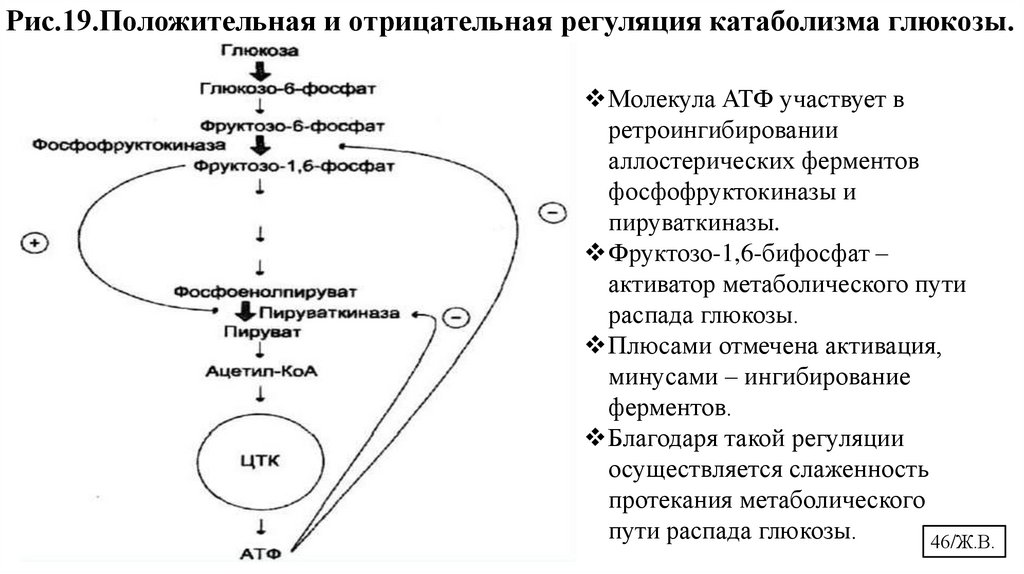
Рис.19.Положительная и отрицательная регуляция катаболизма глюкозы.Молекула АТФ участвует в
ретроингибировании
аллостерических ферментов
фосфофруктокиназы и
пируваткиназы.
Фруктозо-1,6-бифосфат –
активатор метаболического пути
распада глюкозы.
Плюсами отмечена активация,
минусами – ингибирование
ферментов.
Благодаря такой регуляции
осуществляется слаженность
протекания метаболического
пути распада глюкозы.
46/Ж.В.
47.
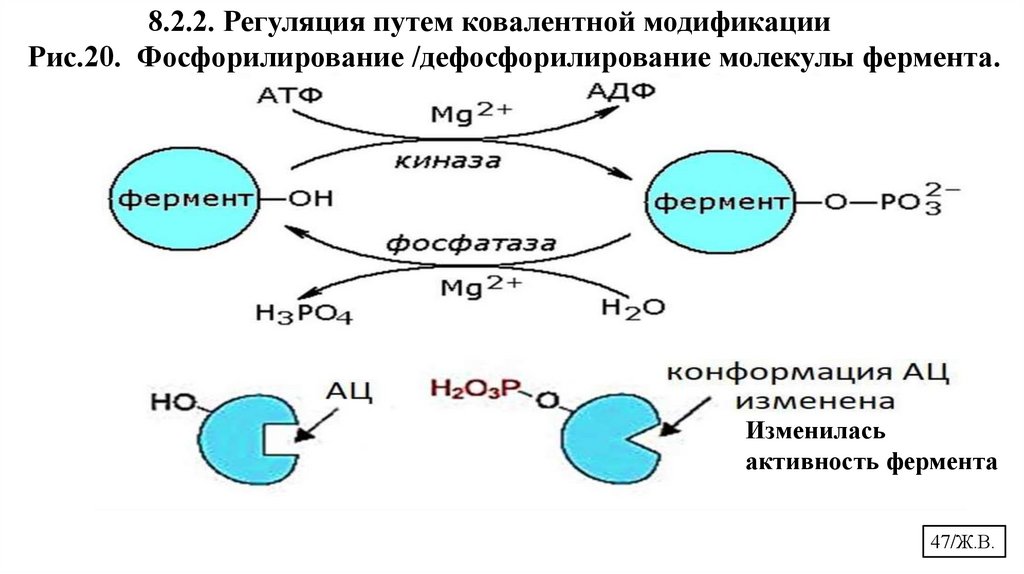
8.2.2. Регуляция путем ковалентной модификацииРис.20. Фосфорилирование /дефосфорилирование молекулы фермента.
Изменилась
активность фермента
47/Ж.В.
48.
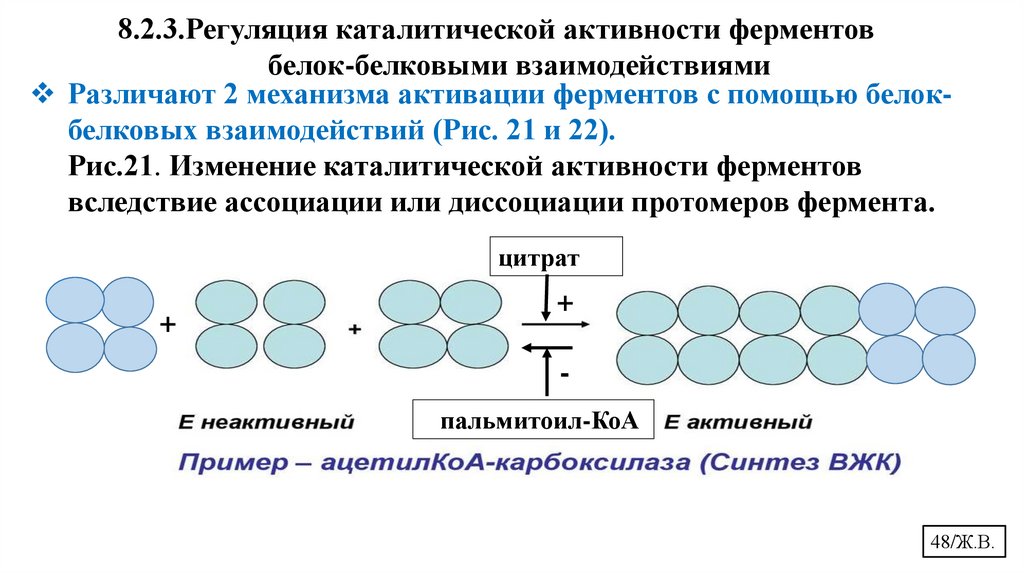
8.2.3.Регуляция каталитической активности ферментовбелок-белковыми взаимодействиями
Различают 2 механизма активации ферментов с помощью белокбелковых взаимодействий (Рис. 21 и 22).
Рис.21. Изменение каталитической активности ферментов
вследствие ассоциации или диссоциации протомеров фермента.
цитрат
+
+
пальмитоил-КоА
48/Ж.В.
49.
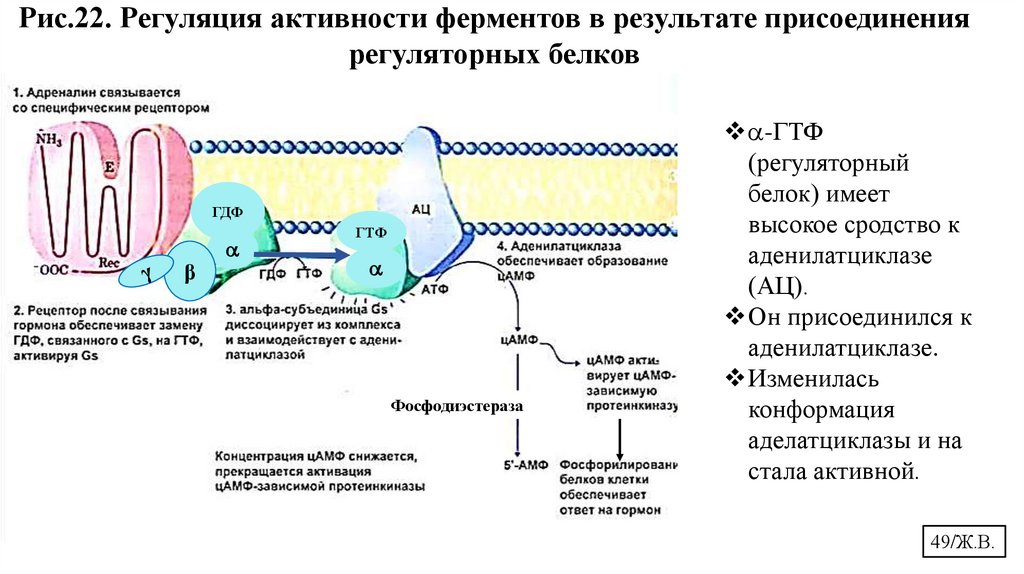
Рис.22. Регуляция активности ферментов в результате присоединениярегуляторных белков
ГДФ
ГДФ
β
ГТФ
Фосфодиэстераза
-ГТФ
(регуляторный
белок) имеет
высокое сродство к
аденилатциклазе
(АЦ).
Он присоединился к
аденилатциклазе.
Изменилась
конформация
аделатциклазы и на
стала активной.
49/Ж.В.
50.
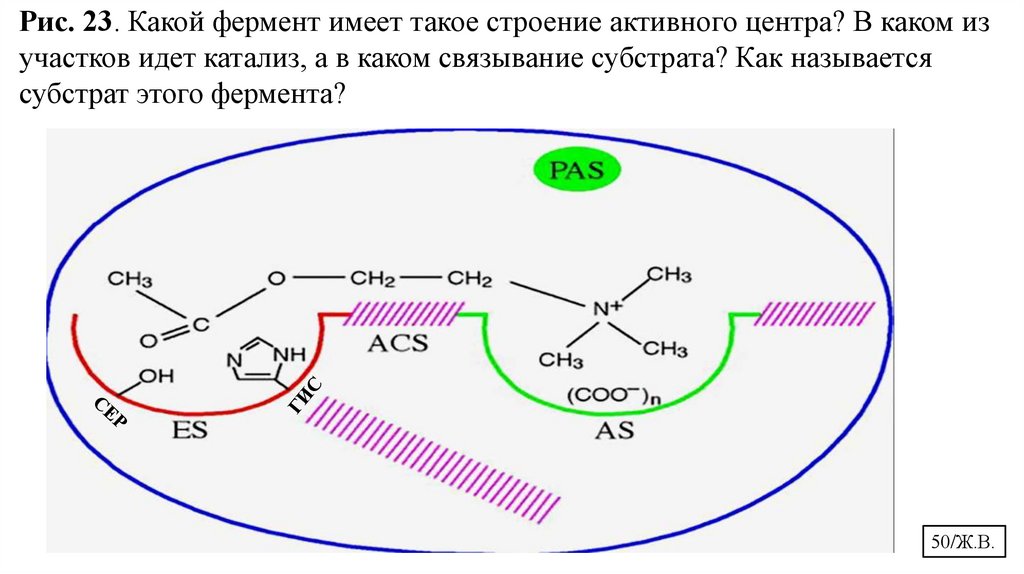
Рис. 23. Какой фермент имеет такое строение активного центра? В каком изучастков идет катализ, а в каком связывание субстрата? Как называется
субстрат этого фермента?
50/Ж.В.
51.
Список литературы1. Биохимия : учебник / под ред. Е.С. Северина. – М. : ГЭОТАР-Медиа, 2011. – С. 100–118.
2. Биохимия [Электронный ресурс] : учебник / под ред. Е. С. Северина. – 5-е изд., испр. и доп. – М. :
ГЭОТАР-Медиа, 2015. – http://www.studmedlib.ru / book / ISBN9785970433126.html (разд. 2, п. VI–IX).
3. Биологическая химия. Биохимия полости рта : учебник / Т.П. Вавилова, А.Е. Медведев. – М. :
ГЭОТАР-Медиа, 2014. – С. 138–154.
4. Биологическая химия. Биохимия полости рта [Электронный ресурс]: учебник / Т.П. Вавилова, А.Е.
Медведев. – М. : ГЭОТАР-Медиа, 2014. – http://www.studmedlib.ru / book / ISBN9785970430392.html (ч. II,
гл. 5).
5. Белки. Ферменты. Витамины : учебно-методическое пособие к практическим и лабораторным
занятиям по биологической химии – биохимии полости рта для студентов стоматологического факультета
/ под ред. Ж.В. Антоновой, Р.Н. Павловой, В.А. Дадали. – СПб. : Изд-во СЗГМУ им. И.И. Мечникова,
2019. – С. 98–108.
6. Березов, Т.Т. Биологическая химия / Т.Т. Березов, Б.Ф. Коровкин. – М. : Медицина, 1998. – С. 145–158,
165–168, 638–641.
51/Ж.В.
52.
СПАСИБО ЗА ВНИМАНИЕ!ПРОЙДИТЕ ТЕСТИРОВАНИЕ.
52/Ж.В.