
















 biology
biologySimilar presentations:
")








")
Основы управления произвольными движениями - основа обучения новому навыку
1.
Основы управленияпроизвольными
движениями – основа
обучения новому навыку.
2.
«... все бесконечное разнообразие внешних проявлениймозговой деятельности сводится окончательно к одному лишь
явлению — мышечному движению».
И.М. Сеченов
Общая схема управления движениями:
Решающий фактор поведения – это результат деятельности
Формирование групп нейронов для достижения результата (функциональная система)
Обработка сигналов,
поступающих из
внешней и внутренней
среды
(афферентный синтез)
Принятие
решения о целях
и задачах
действий
Создание
представления
об ожидаемом
результате и
формирование
конкретной
программы
движений
Анализ
полученного
результата и
внесение в
программу
поправок —
сенсорных
коррекций
3.
В процессах афферентного синтеза участвуют глубокиевнутренние процессы — побуждение к действию (мотивация) и
его замысел, извлекаются из памяти моторные следы (навыки) и
выученные тактические комбинации. У человека на их основе
создается определенный план и конкретная программа
движения. Эти процессы отражаются в изменениях электрической
активности мозга. На уровне спинного мозга процессы
преднастройки
отражаются
повышением
возбудимости
спиральных мотонейронов, в мышцах — повышением
чувствительности
проприорецепторов
скелетных
мышц.
Сенсорная информация о результате выполнения движения,
получаемая по каналам обратной связи, используется нервными
центрами для уточнения временных, пространственных и
силовых характеристик двигательных актов, внесения поправок в
команды. Такие поправки называются сенсорными коррекциями.
4.
Рис 1. Предположительная блок-схема прохождения управляющей информации при регуляции произвольных движений человека5.
Центральные аппараты и механизмы управления движениямиУ позвоночных центральные программы запускаются управляющими центрами, а у
беспозвоночных, как правило, командными нейронами, впервые выделенными
в 1968 г. К. Вирсма. Однако сигнал от командного нейрона или управляющего центра играет лишь
пусковую роль. Он не несет информации о том, как построить соответствующее движение.
Структура движения заложена во взаимосвязях мотонейронов, с которыми он связан.
Рассмотрим несколько примеров поведения у низших животных:
- рефлекс бегства у рака, легко вызываемый тактильным,
вибрационным или звуковым раздражением, реализуется через
возбуждение одного гигантского командного нейрона, связанного с
тремя двигательными мотонейронами
- в ответ на стимуляцию рыба уплывает, совершая быстрые и
повторяющиеся движения хвостом и головой; этот двигательный
акт запускается парой командных нейронов – маутнеровских
клеток, расположенных в продолговатом мозгу;
- у таракана есть командный нейрон, запускающий бег насекомого,
если его лапы касаются опоры, если же лапы находятся в воздухе
запускается команда полета.
6.
Ведущая роль в построении новых моторных программ у высших животных и человекапринадлежит передним отделам коры больших полушарий – премоторной и
префронтальной коре. Роль передних отделов коры больших полушарий показана в
работах А.С. Батуева, проводившего эксперименты с использованием
микроэлектородов, регистрирующих активность отдельных нейронов у обезьян. В
экспериментах были обнаружены три группы нейронов, последовательно вовлекаемых
в процесс осуществления условнорефлекторного двигательного навыка. Все три
популяции клеток были обнаружены в теменной и особенно в лобной коре.
1 группа клеток: реагирует на условные сигналы, поэтому их отнесли к
сенсорным нейронам;
2 группа клеток: реагирует на время действия сигнала;
3 группа клеток: реагирует в пусковой период.
Считают, что сенсорные нейроны первой группы передают информацию второй
группе, которые принадлежат кратковременной памяти. Третья же группа – это
нейроны моторных программ, они получают информацию от нейронов памяти
и запускают хорошо отработанную двигательную реакцию.
7.
Механизмы инициации движенияИнициация движения происходит следующим образом. В моторной коре строится
конечный и конкретный вариант моторного управления движением. Моторная кора
использует оба принципа управления: контроль через систему обратной афферетации и
через механизм центрального программирования. Это достигается тем, что к ней
сходятся сигналы от мышечной активности, от сенсомоторной, зрительной и других
отделов коры, которые и используются для моторного контроля коррекции движения.
Кроме того, к моторной коре приходят сигналы, связанные с программированием
движения из передних отделов коры и подкорки. Моторная кора получает также
информацию из структур, имеющих отношение к хранению двигательных программ.
Так, в опытах с регистрацией нейронной активности у обезьян было показано, что
при выполнении ими заученного движения активность нейронов зубчатого ядра
мозжечка на 10 мс опережает изменение активности нейрона в моторной коре,
которое предшествует появлению мышечного движения. В моторную кору поступают
также сигналы от базальных ганглиев – структура, которая ответственна за хранение
главным образом двигательных программ врожденного поведения (пищевого,
питьевого и др.), стереотипных движений. Клетки базальных ганглиев, так же как и
мозжечка, разряжаются до начала движений, совершаемых животными в ответ на
сигнал за 50 – 150 мс. В хвостатом ядре были найдены нейроны, у которых
возбуждение опережает условную двигательную пищевую и оборонительную реакции.
Эти реакции у нейронов появляются с выработкой условного рефлекса и исчезают с его
утешением. Сигналы от базальных ганглиев и мозжечка распространяются не только
вверх к коре, но и вниз, по нисходящему пути к спинному мозгу (экстрапирамидный
путь).
8.
Согласно работам шведского нейрофизиолога Г.Могенсона, все процессы управления движением
включают три блока и соответствующие им три фазы:
1) блок инициации движения, включающий
лимбическую систему с прилегающим ядром (n.
accumbens), и ассоциативную кору;
2) блок программирования движения, включающий
мозжечок, базальные ганглии, моторную кору,
таламус (как посредник между ними), а также
спинальные и стволовые генераторы;
3) исполнительный блок, охватывающий мотонейроны
и двигательные единицы.
9.
Рис 2: Схема инициации двигательного акта (по Г. Могенсону, 1977, сизменениями)
10.
В двигательной деятельности человека различают:• произвольные движения (сознательно управляемые целенаправленные действия)
• непроизвольные движения, котрые происходят без участия сознания и
представляют собой либо безусловные реакции, либо автоматизированные
двигательные навыки.
В основе управления произвольными движениями человека лежат два различных
физиологических механизма:
рефлекторное кольцевое регулирование
программное управление по механизму центральных команд.
Рефлекторное кольцевое регулирование:
Замкнутая система рефлекторного кольцевого
регулирования характерна для осуществления
различных форм двигательных действий и
позных реакций, не требующих быстрого
двигательного акта. Это позволяет нервным
центрам получать информацию о состоянии
мышц и результатах их действий по различным
афферентным путям и вносить поправки в
моторные команды по ходу действия.
11.
Программное управление по механизму центральных команд — этомеханизм регуляции движений, независимый от афферентных
проприоцептивных влияний. Такое управление используется в
случае выполнения кратковременных движений (прыжков, бросков,
ударов ), когда организм не успевает использовать информацию от
проприорецепторов мышц и других рецепторов. Вся программа
должна быть готова еще до начала двигательного акта. При этом
отсутствует замкнутое кольцо регуляции. Управление производится
по так называемой открытой петле, а активность во многих произвольно сокращающихся мышцах возникает раньше, чем регистрируется обратная афферентная импульсация. Например, при выполнении прыжковых движений электрическая активность в мышцах,
направленная на амортизацию удара, возникает раньше, чем происходит соприкосновение с опорой, т. е. она носит предупредительный
характер.
12.
Такие центральные программы создаются согласно сформированному в мозге(главным образом — в ассоциативной передне-лобной области коры) образу
двигательного действия и цели движения.
В дальнейшей конкретной разработке моторной программы принимают участие
мозжечок (латеральная область его коры) и базальные ядра (полосатое тело и
бледный шар). Информация от них поступает через таламус в моторную и
премоторную области коры и далее — к исполнительным центрам спинного мозга
и скелетным мышцам.
13.
Механизм кольцевого регулирования являетсяфилогенетически более древним и возникает раньше в
процессе индивидуального развития. Примерно к трем годам
достаточное развитие получают зрительные обратные связи,
осуществляющие текущий зрительно-моторный контроль, а с
5-6 лет происходит переход к текущему контролю движений с
участием проприоцептивных обратных связей. Этот механизм
достигает значительного совершенства к 7-9 летнему возрасту, после чего начинается переход к формированию
механизма центральных команд. К 10-11 годам повышение
скорости произвольных движений обеспечивается
достаточным развитием процессов предварительного
программирования их пространственных и временных
параметров. С этого возраста представлены оба механизма
управления произвольными движениями, дальнейшее
совершенствование которых продолжается вплоть до 17-19
лет.
14.
Роль ЦНС в регуляции движений:Спинной мозг осуществляет ряд элементарных двигательных рефлексов: рефлексы на
растяжение (миостатические и сухожильные рефлексы, например, коленный
рефлекс), кожные сгиба тельные рефлексы (например, защитный рефлекс отдергивания конечности при уколах, ожогах), разгиба тельные рефлексы (рефлекс
отталкивания от опоры, лежащий в основе стояния, ходьбы, бега), перекрестные
рефлексы и др.
Составной частью различных сложных двигательных
действий, как произвольных, так и непроизвольных,
часто являются ритмические рефлексы. Это одна из форм
древних и относительно простых рефлексов. Они
особенно выражены при выполнении циклической
работы, включаются в шагательные рефлексы. Основные
механизмы шагательных движений заложены в спинном
мозге. Специальные нейроны (спинальные
локомоторные генераторы) и многочисленные
взаимосвязи внутри спинного мозга обеспечивают
последовательную активность различных мышц
конечностей, согласование ритма и фаз движений,
приспособление движений к нагрузке на мышцы. В
среднем мозгу расположены нейроны «локомоторной
области», которые включают этот механизм и
регулируют мощность работы мышц, обеспечивая
примитивную форму локомоций — без ориентации в
пространстве.
15.
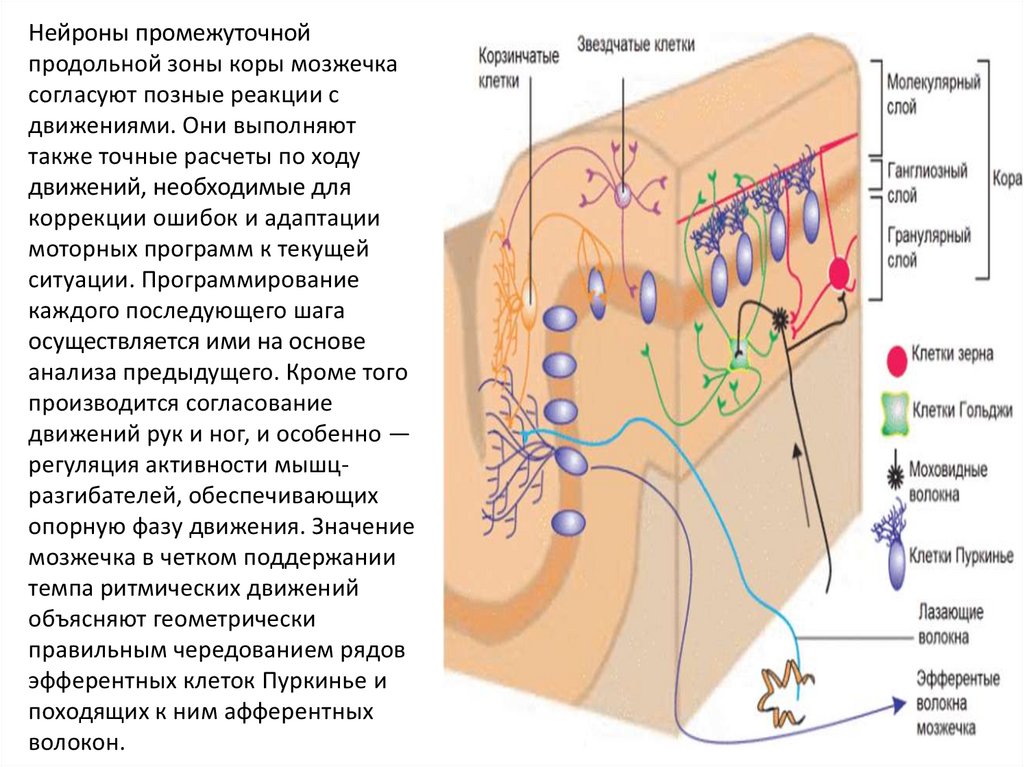
Нейроны промежуточнойпродольной зоны коры мозжечка
согласуют позные реакции с
движениями. Они выполняют
также точные расчеты по ходу
движений, необходимые для
коррекции ошибок и адаптации
моторных программ к текущей
ситуации. Программирование
каждого последующего шага
осуществляется ими на основе
анализа предыдущего. Кроме того
производится согласование
движений рук и ног, и особенно —
регуляция активности мышцразгибателей, обеспечивающих
опорную фазу движения. Значение
мозжечка в четком поддержании
темпа ритмических движений
объясняют геометрически
правильным чередованием рядов
эфферентных клеток Пуркинье и
походящих к ним афферентных
волокон.
16.
К управлению ритмическими движениями непосредственное отношение имеютактивирующие и угнетающие отделы ретикулярной формации, которые влияют на силу и
темп сокращения мышц, а также подкорковые ядра, которые организуют автоматическое
их протекание и содружественные движения конечностей. Включение древних форм
ритмических движений (циклоидных) в акт письма позволяет человеку перейти от
отдельного начертания букв к обычной письменной скорописи. То же самое происходит
при освоении акта ходьбы — с переходом от отдельных шагов к ритмической походке.
Плавность ритмических движений, четкое чередование реципрокных сокращений мышц
обеспечивают премоторные отделы коры.
Функцией комплекса различных корковых областей является определение
целесообразности локомоций, их смысла, ориентации в пространстве, перестройка
программ движений в различных ситуациях, включение ритмических движений как
составного элемента в сложные акты поведения. Об участии различных корковых
областей в регуляции циклических движений можно судить по появлению в их
электрической активности медленных потенциалов в темпе движения — «меченых
ритмов» ЭЭГ, а при редких движениях — по изменениям кривой, огибающей
амплитуду ЭЭГ.
17.
В высшей регуляции произвольных движений важнейшая роль принадлежитпередне-лобным областям (передним третичным полям). Здесь помимо обычных
вертикальных колонок нейронов существует принципиально новый тип
функциональной единицы — в форме замкнутого нейронного кольца. Циркуляция
импульсов в этой замкнутой системе обеспечивает кратковременную память. Она
сохраняет в коре возбуждение между временем прихода сенсорных сигналов и
формированием ответной эфферентной команды. Такой механизм служит основой
сенсомоторной интеграции при программировании движений, при осуществлении
зрительно-двигательных реакций.