

 нервной системы:")











 и клиновидный (Бурдаха)")



















 Наличие функциональных колонок.")











")









 biology
biologySimilar presentations:
 нервной системы")


")



")


Двигательная система
1. Двигательная система
2. Проявления двигательных функций у человека, их характеристика. Отделы соматической нервной системы, ее взаимоотношения с
другимисистемами.
3. Функции двигательной (соматической) нервной системы:
• 1.Управляет деятельностью опорнодвигательногоаппарата за счет: рефлекторных движений,
• включения врожденных и приобретенных двигательных
программ, произвольных движений.
• 2. Включает транспортное (ТО) и метаболическое
обеспечение (МО) двигательных функций.
3. Тонизирует деятельность психоэмоциональных
и надсегментарных вегетативных структур.
• 4. 2-е и 3-е обеспечивает тренирующий эффект
для поддержания здоровья.
• Отделы ДС: сенсорный, моторный.
4.
Организация движения:1.Побуждение к движению. Сигналы, побуждающие к
движению, поступают из ассоциативных и мотивационных
зон КБП и ЛС от:
внешних рецепторов (фоно-, фото-, вкусовых,
обонятельных, температурных, болевых, тактильных),
внутренних рецепторов (интерорецепторов).
2. Движение. Обеспечивает моторный отдел ДС.
Его функции:
1. Поддержание позы и равновесия, т.е. преодоление
внешних сил, в том числе силы тяжести, за счет
тонических сокращений медленных ДЕ, устойчивых
к утомлению.
5.
• 2.Собственно движения:• передвижение тела и его частей в пространстве,
• рабочие движения (профессиональные,
спортивные, бытовые навыки),
• коммуникации и передача эмоций
• (речь, мимика, жесты).
• Это обеспечивают ритмические сокращения
быстрых и медленных ДЕ.
• 3. Анализ движения.
6. Общий вид двигательной системы
Сигналы, побуждающие к движению поступают из ассоциативных имотивационных зон КБП и ЛС от рецепторов:
Внешний стимул
с рецепторов:
-фото
-фоно
-вкусовых
-обонятельных
-термо
-болевых
Проприорецепторы,
вестибулорецепторы,
тактильные,
висцеральные
рецепторы
моторный
отдел
поза движения
Внутренний стимул
с рецепторов
-проприо
-висцеро
-вестибуло
-термо
-болевых
неспецифические
эффекты
сенсорный
отдел
Коррекция движений
7.
Сенсорный отдел ДС(соматический анализатор,
кинестетический анализатор)
8. Сенсорный отдел ДС
• В результате получения сигналов, возникающихв процессе движения от проприорецепторов,
вестибулорецепторов, тактильных и
висцеральных рецепторов),
• обеспечивает:
• - контроль и коррекцию движений;
• -анализ положения и движения частей тела и
тела в пространстве.
9.
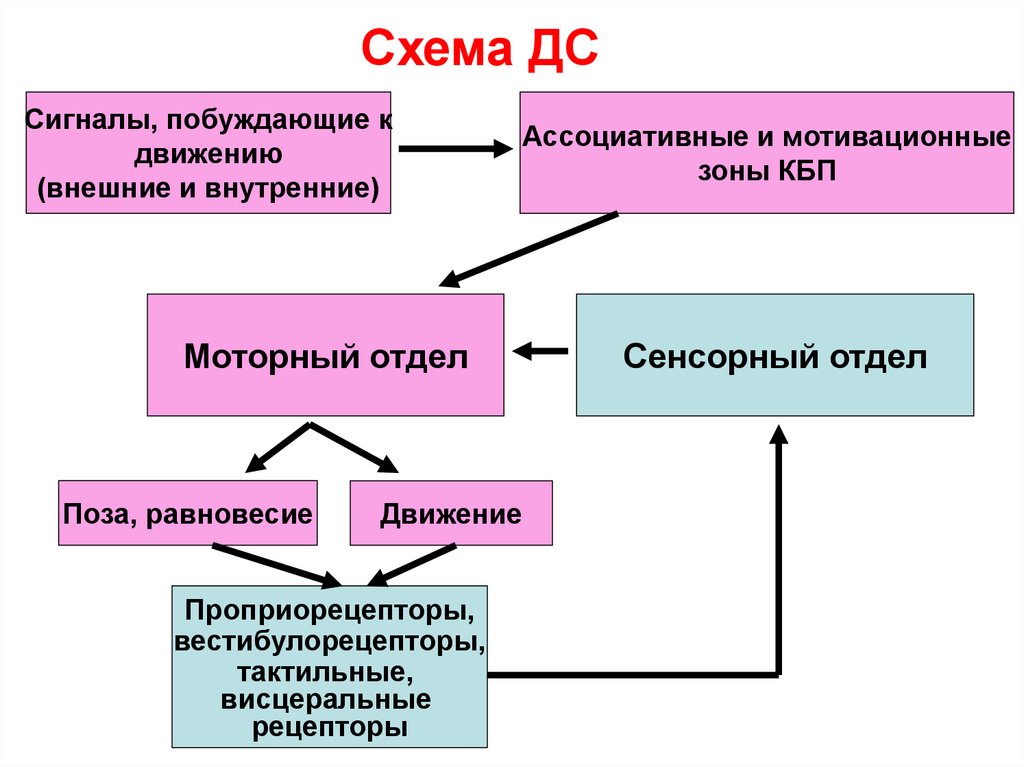
Схема ДССигналы, побуждающие к
движению
(внешние и внутренние)
Ассоциативные и мотивационные
зоны КБП
Моторный отдел
Поза, равновесие
Движение
Проприорецепторы,
вестибулорецепторы,
тактильные,
висцеральные
рецепторы
Сенсорный отдел
10.
Сенсорный отдел ДСРасположен в ЗЦИ КБП.
Представлен 2-мя анализаторами:
• кинестетическим (сомато-сенсорным),
• вестибулярным.
Кинестетический анализатор служит
для оценки положения и движения
конечностей и туловища,
имеет 3 отдела: рецепторный,
проводниковый и корковый (в ЗЦИ).
11.
Рецепторный отдел кинестетическогоанализатор – первичночувствующие
рецепторы:
1) проприорецепторы мышц, сухожилий и связок:
• рецепторы растяжения (мышечные веретена),
реагируют на растяжение мышц,
• рецепторы Гольджи – на сокращение мышц,
рецепторы связок и суставных сумок (тельца Пачини,
окончания Руффини), реагируют на движение в суставах.
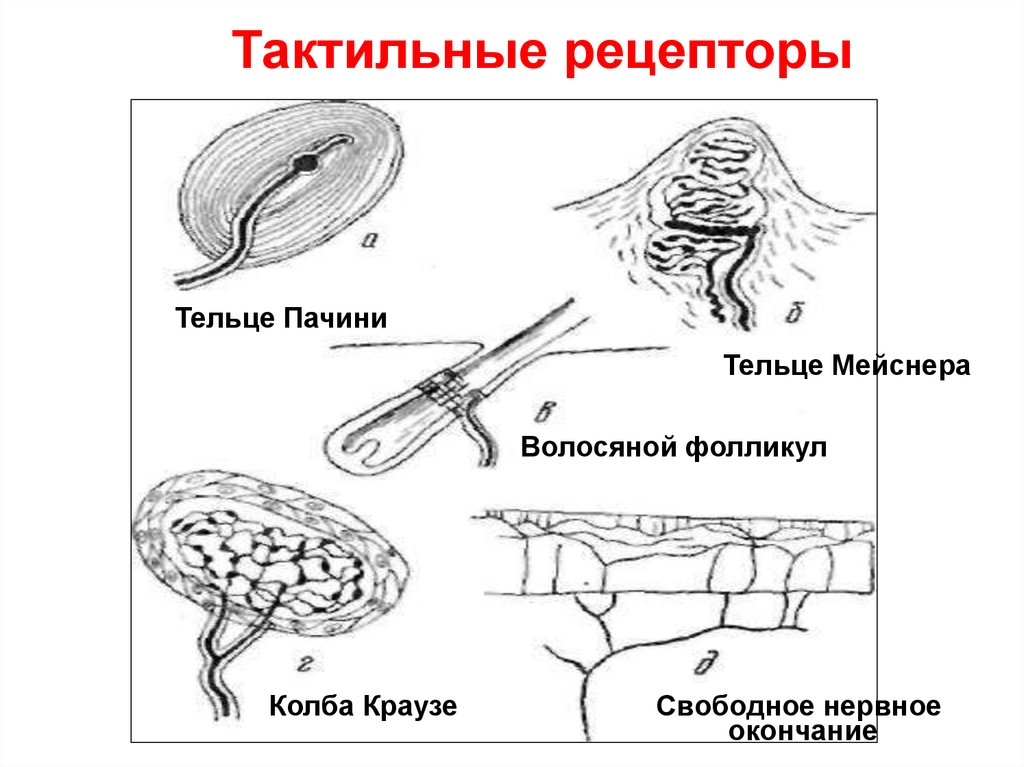
2) Тактильные высокодифференцированные
рецепторы кожи (тельца Мейснера, диски Меркеля, тельца
Фатер-Пачини).
12.
Тактильные рецепторыТельце Пачини
Тельце Мейснера
Волосяной фолликул
Колба Краузе
Свободное нервное
окончание
13.
Проприорецепторы:1) Мышечные веретена (рецептор
растяжения),
2) Рецепторы Гольджи
(возбуждаются при сокращении
мышцы).
14.
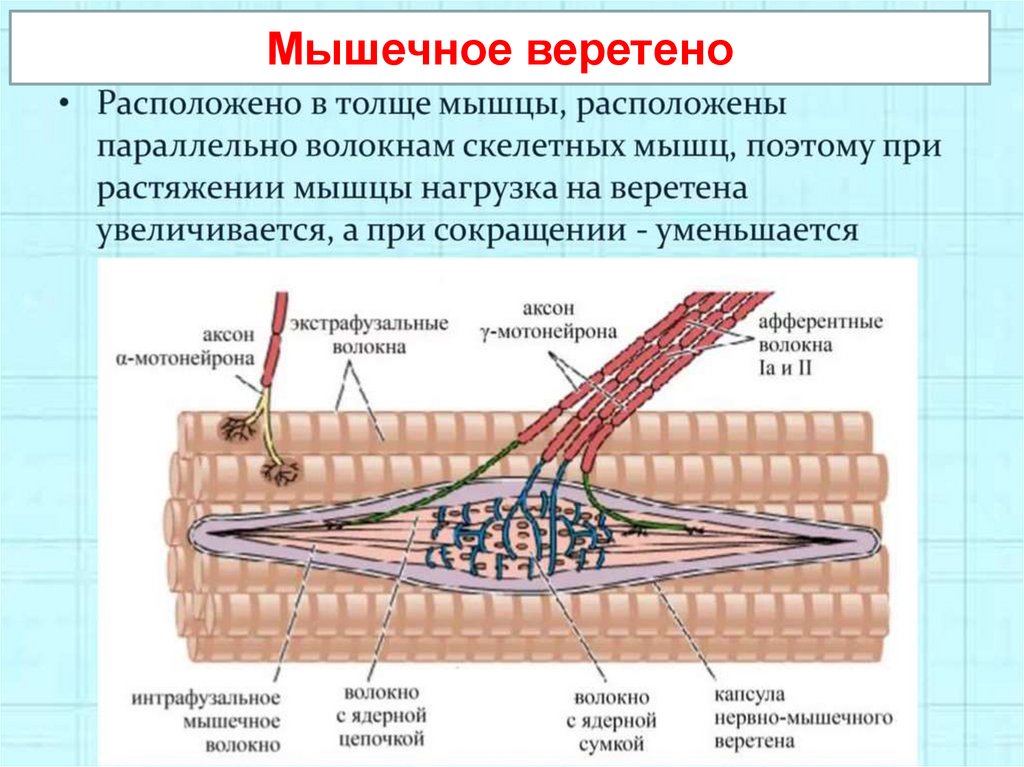
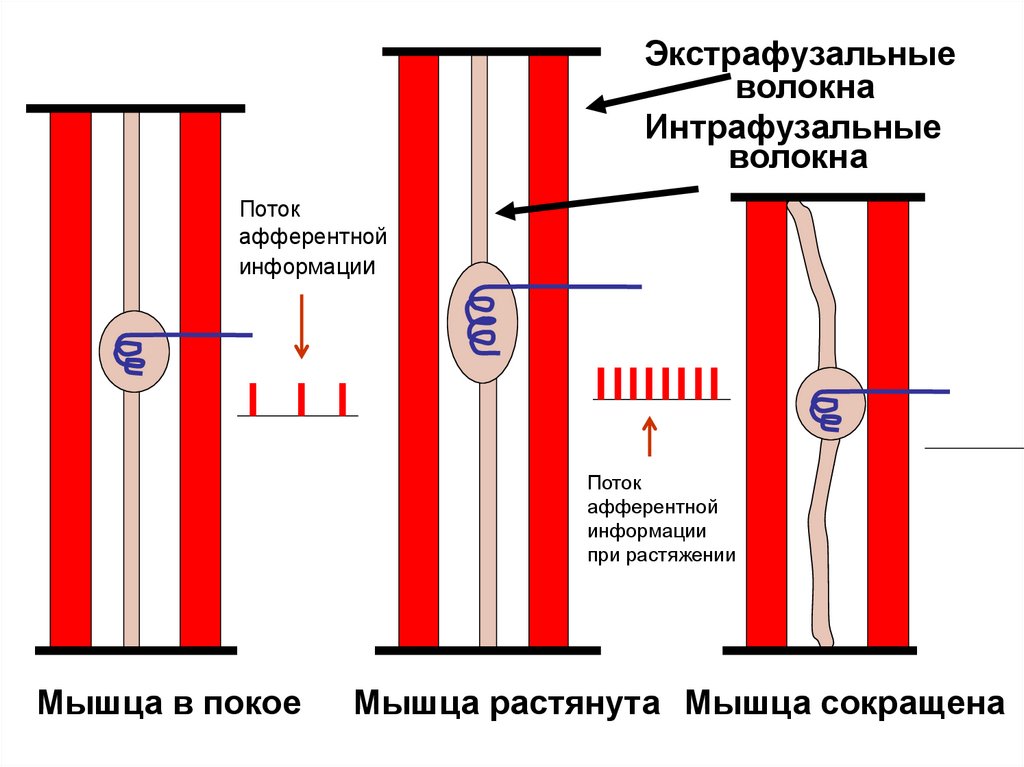
Мышечные веретена (2-3 мм) располагаются параллельно мышечнымволокнам (экстрафузальным волокнам),
возбуждаются при растяжении мышц.
Образованы интрафузальными
волокнами с ядерной сумкой (или
ядерной цепочкой), вокруг которых
спирально закручено окончание афферентного нейрона, реагирующее
на растяжение мышцы. Измеряют
длину мышцы.
15.
Мышечное веретено16.
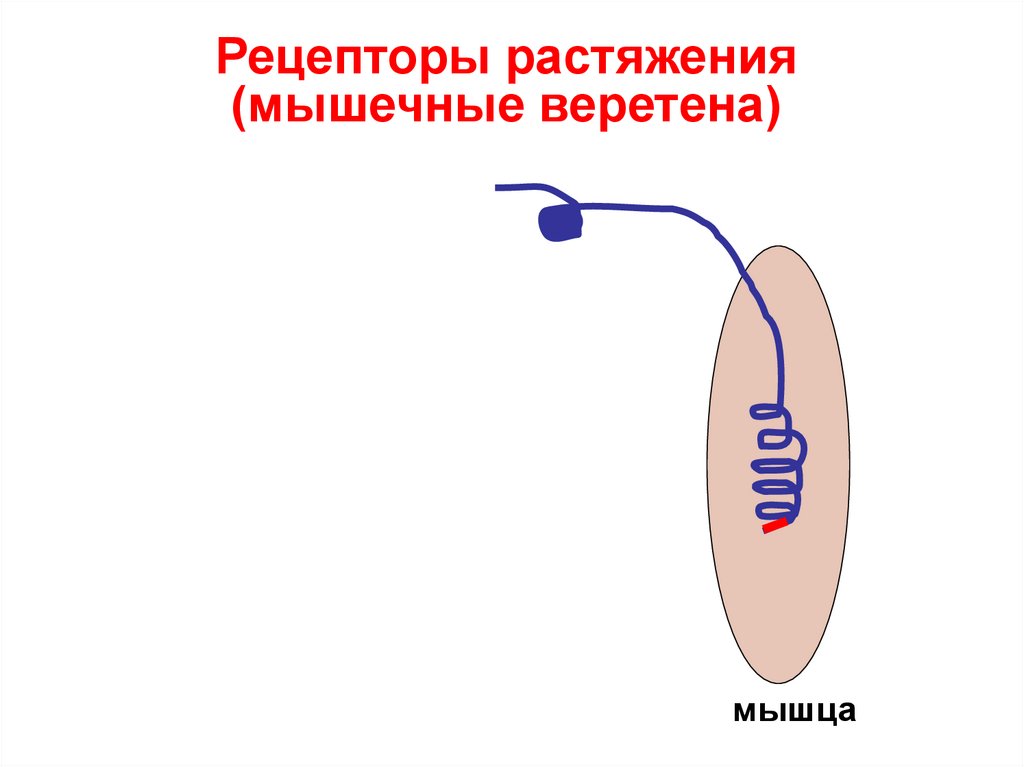
Рецепторы растяжения(мышечные веретена)
мышца
17.
Экстрафузальныеволокна
Интрафузальные
волокна
Поток
афферентной
информации
Поток
афферентной
информации
при растяжении
Мышца в покое
Мышца растянута Мышца сокращена
18.
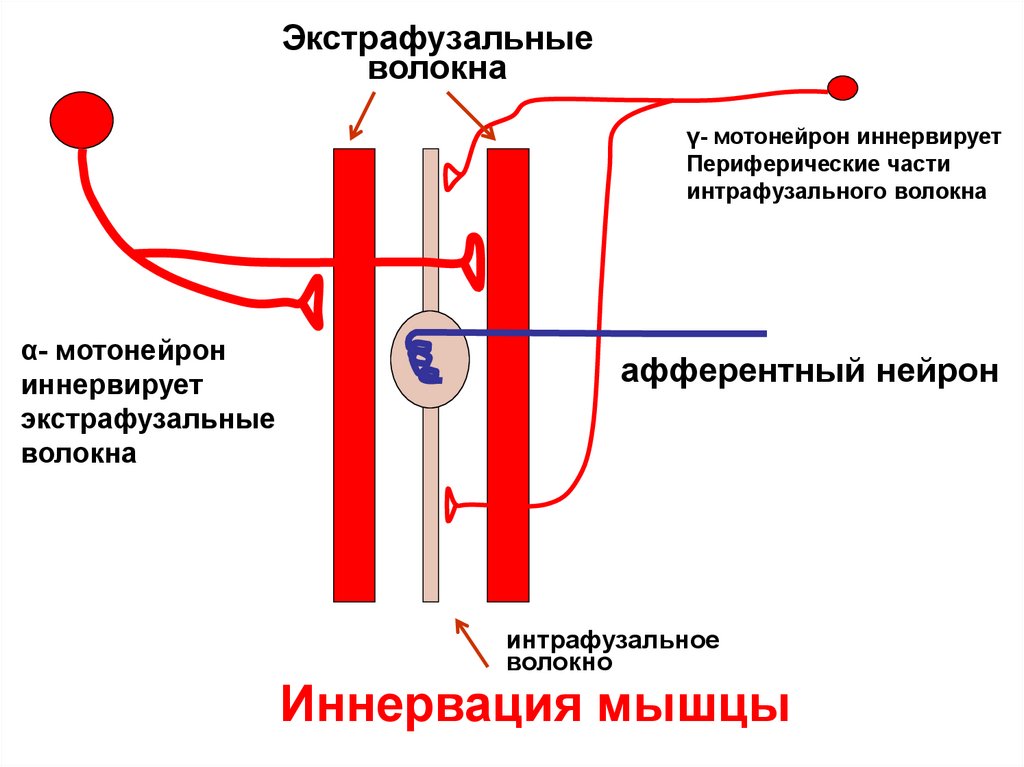
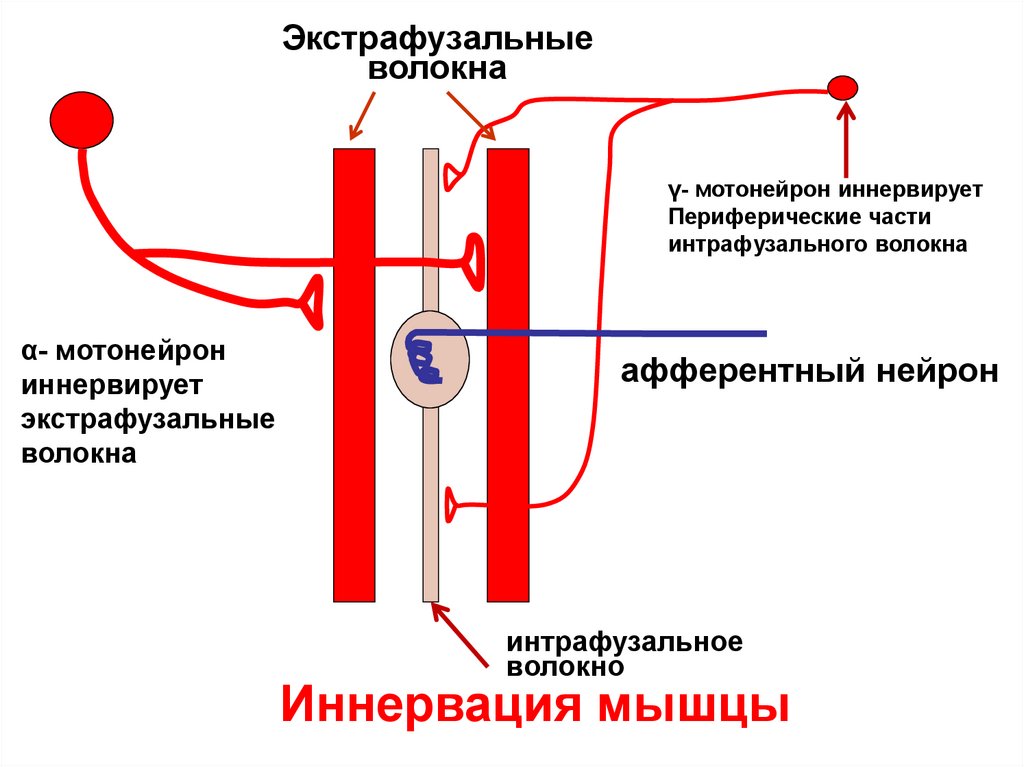
Экстрафузальныеволокна
γ- мотонейрон иннервирует
Периферические части
интрафузального волокна
α- мотонейрон
иннервирует
экстрафузальные
волокна
афферентный нейрон
интрафузальное
волокно
Иннервация мышцы
19.
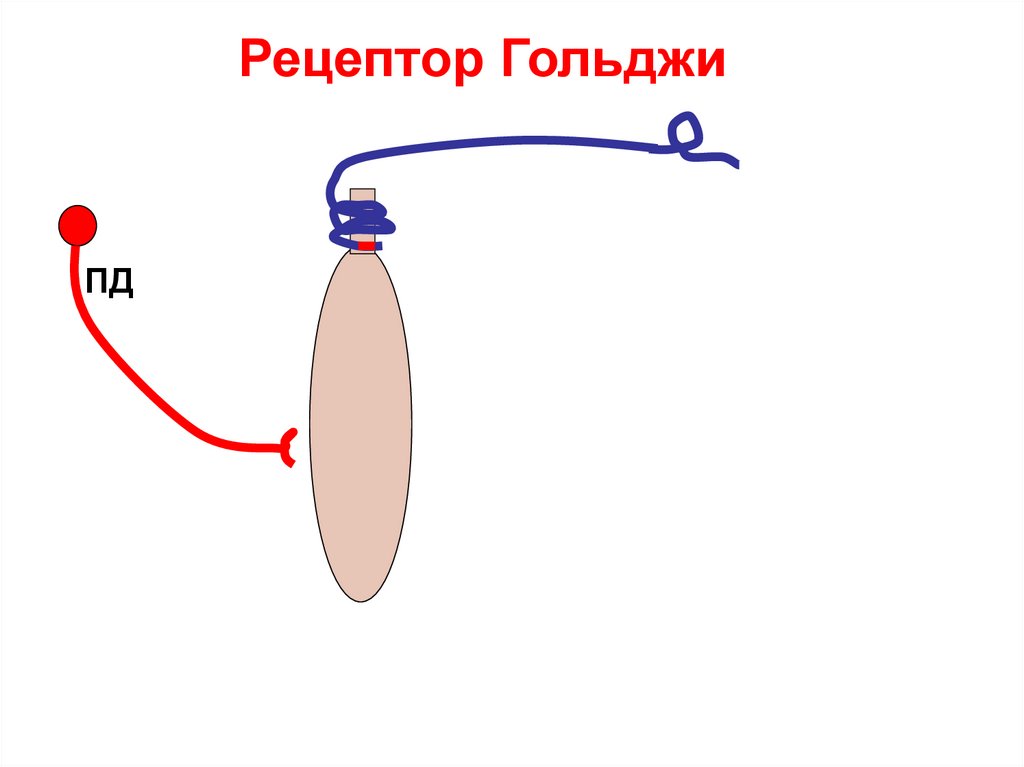
Рецепторы Гольджи•Группа сухожильных волокон, оплетенных дендритом чувствительного
•нейрона.
•Возбуждаются при сокращении
мышцы, когда сухожилие
натягивается,
•т.е. измеряют мышечное напряжение.
20.
Рецептор ГольджиПД
21. Афферентные пути соматической сенсорной системы
22. Тонкий (Голля) и клиновидный (Бурдаха)
• – проходит в задних столбахспинного мозга.
• По волокнам тонкого пучка
проводится возбуждение от
нижней части тела и нижних
конечностей.
• По волокнам клиновидного пучка
- от верхней части туловища и
верхних конечностей.
23.
• I нейрон – в спинальномганглии;
• II нейрон – в
продолговатом мозге в
соответствующих ядрах,
• затем после перекрестка
в латеральное ядро
таламуса,
• затем в
соматосенсорную зону
коры в ЗЦИ.
24.
• Скорость проведения 60 – 100м/с.• Обеспечивает кожно-механическую тонко
дифференцированную чувствительность,
• т. е. локализацию действия раздражителя,
изменение его во времени.
• Волокна этого пути дают коллатерали на
мотонейроны и интернейроны своего и
• вышележащего сегмента спинного мозга,
• образуя межсегментарные связи.
25.
Проводящий путь Голя и БурдахаКора БП
Таламус
Продолговатый мозг
Спинной мозг
26. Спиноталамический путь.
• Латеральный – проводит болевуюи температурную чувствительность.
• Вентральный – тактильную
чувствительность.
• Медленнопроводящий
• 1 – 30м/с.
• Дает информацию о качественной
природе раздражителей
27.
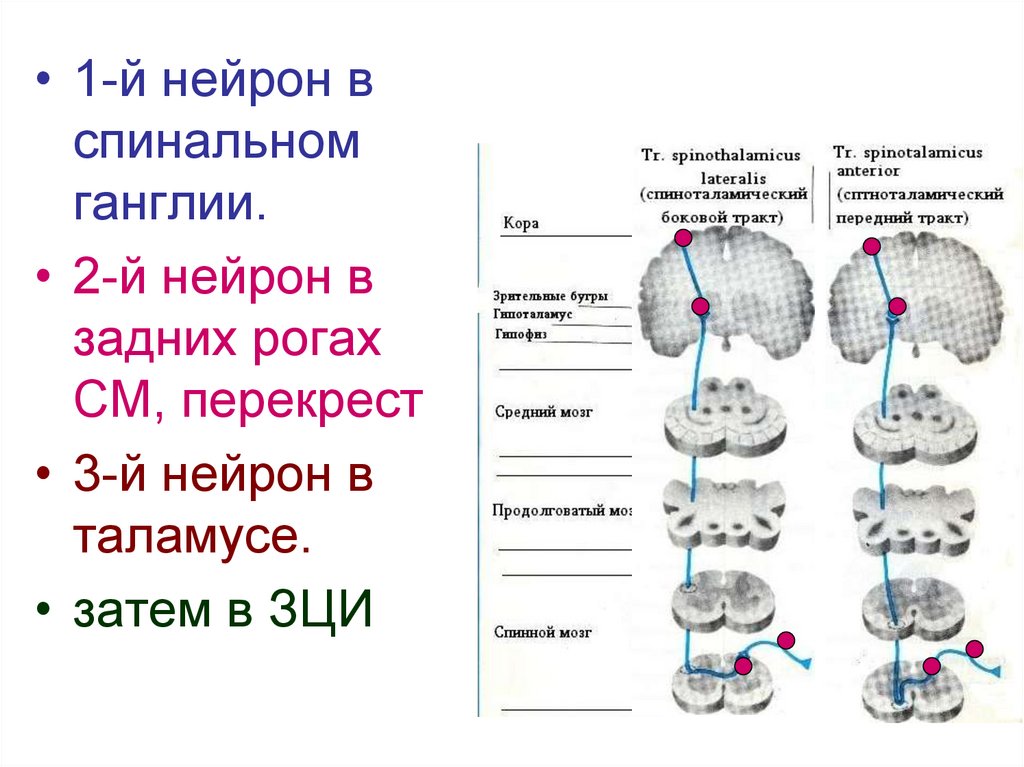
• 1-й нейрон вспинальном
ганглии.
• 2-й нейрон в
задних рогах
СМ, перекрест
• 3-й нейрон в
таламусе.
• затем в ЗЦИ
28. Спиномозжечковый путь.
• Скорость проведения – 110 – 120 м/с.• Обеспечивает позу и тонус мышц при движении.
Дорзальный – не перекрещенный Флексига.
• Передает информацию от мышц и связок
конечностей.
• Вентральный путь Говерса дважды
перекрещенный .
• Передает информацию от сухожильных, кожных
и висцерорецепторов через продолговатый мозг
и ножки мозга к коре мозжечка.
29.
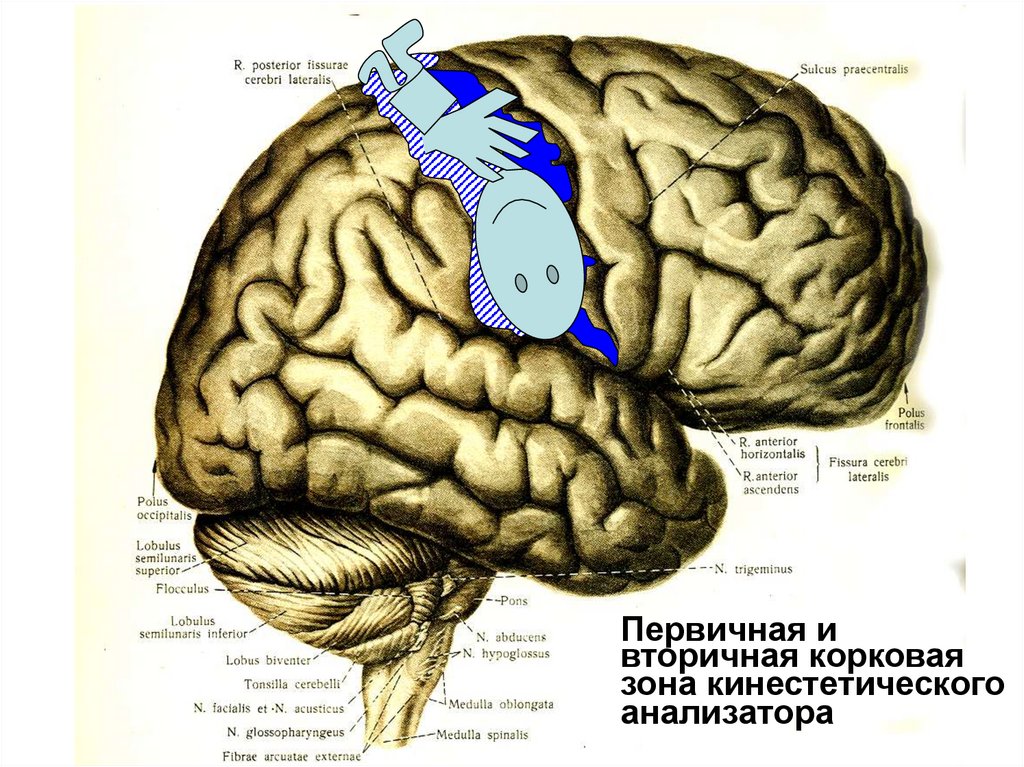
Корковый отдел кинестетическогоанализатра
Имеет первичную корковую проекционную зону,
расположеную в ЗЦИ.
Она имеет соматотопическую организацию
т.е.нейроны каждого отдела ЗЦИ получают и
анализируют информацию от мышц определенной
части тела:
от нижних конечностей – в верхне-медиальной
части ЗЦИ,
• от туловища – в верхней,
• от рук – в средней части,
• от шеи и головы – в средней и нижней.
30.
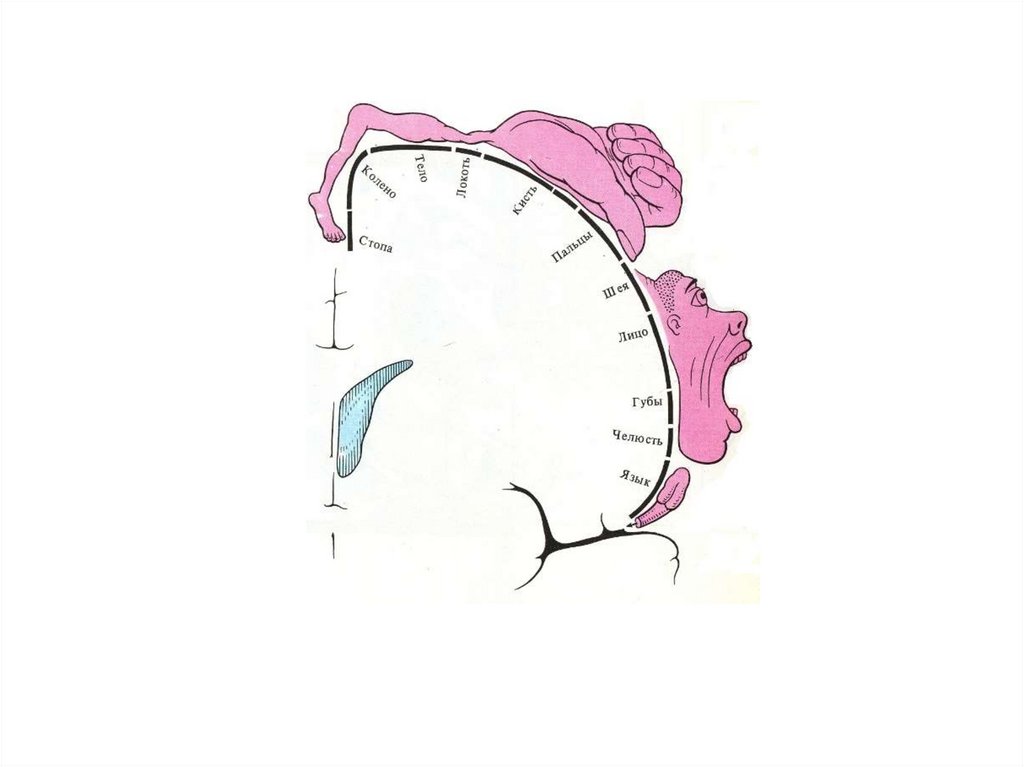
Первичная ивторичная корковая
зона кинестетического
анализатора
31. Центральные структуры сенсорного отдела.
• Кора.В каждом полушарии выделяются зоны соматической чувствительности –
соматосенсорные зоны (ССЗК).
Первая расположена в ЗЦИ. Здесь формируются ощущения
положения частей тела в пространстве, чувство мышечной радости,
усталости.
Вторая в сильвиевой борозде.– восприятие схемы тела, движения
тела в пространстве
Принципы организации ССЗК.
Нейроны 6-ти слоев коры объединены в сенсорные колонки, каждая
из которых получает информацию от проприорецепторов при
движении в определенном суставе.
1) Соматотопическая организация,
т. е. у различных рецептивных полей имеется определенная область
проекции в ЗЦИ.
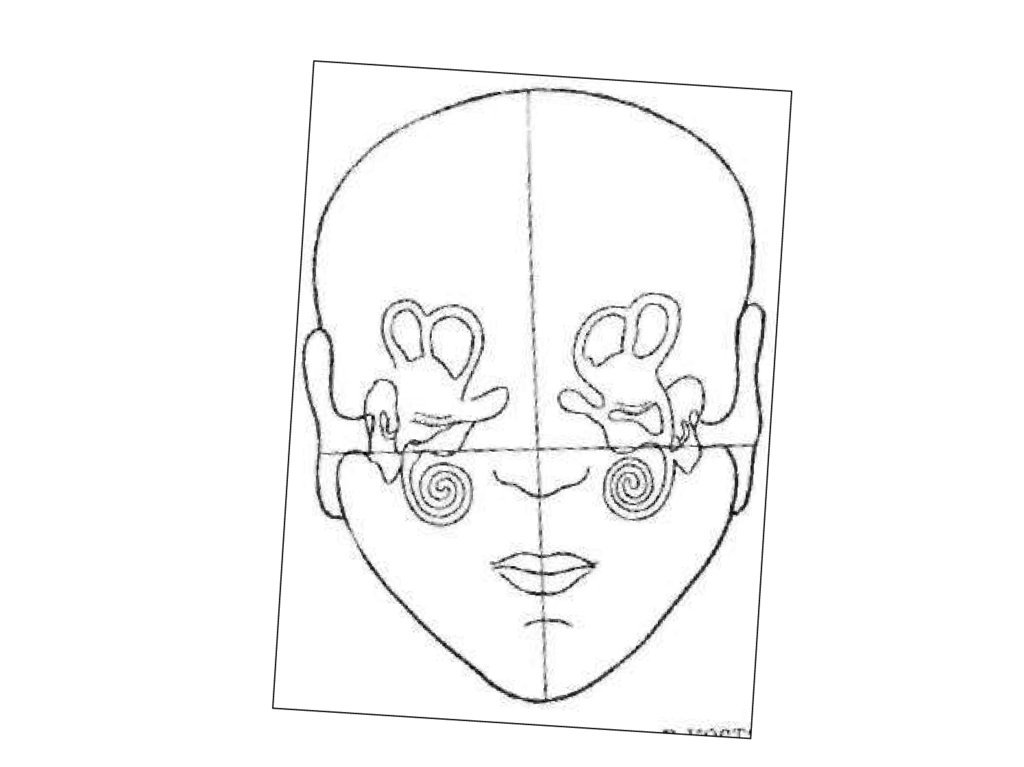
2) Сенсорный гомункулус,
т. е. различные рецептивные поля имеют различную площадь
представительства в ЗЦИ.
32. Роль соматосенсорной зоны коры
• Интеграция икритическая оценка
информации от
специфических ядер
таламуса.
• В этой же области
находится и
представительство
висцерорецепторов.
33.
Вестибулярный анализатор34.
35. Вестибулярный анализатор
• Воспринимает, анализирует и декодирует в ощущения ивосприятия сигналы, поступающие от вестибулорецепторов
преддверия и полукружных каналов, т.е. участвует в
пространственной ориентации головы и тела в покое и при
движении.
• Рецепторный отдел.
• Вторичночувствующие рецепторы двух отделов
вестибулярного аппарата.
• 1.Рецепторы преддверия
• (маточка и мешочек) реагируют на наклоны головы и
тела, тряску, прямолинейные движения с ускорением.
• Порог наклона – 1-20, ускорения – 2-20 см/с2.
36.
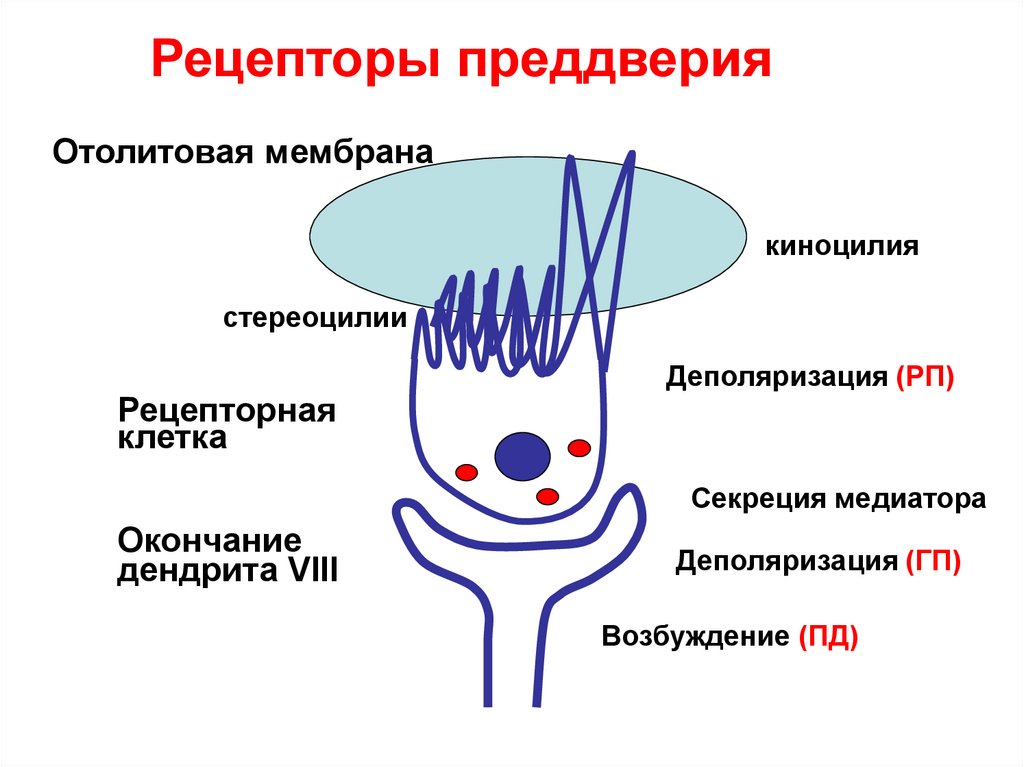
.• Рецепторы преддверия – волосковые
клетки, имеют длинный волосок
(киноцилию) и несколько коротких
(стереоцилии). Они погружены в
отолитовую мембрану, которая смещается
под действием силы тяжести при
наклонах головы.
• Смещение в сторону киноцилии
вызывает деполяризацию мембраны,
в сторону стереоцилий –
гиперполяризацию.
37.
Рецепторы преддверияОтолитовая мембрана
киноцилия
стереоцилии
Рецепторная
клетка
Деполяризация (РП)
Секреция медиатора
Окончание
дендрита VIII
Деполяризация (ГП)
Возбуждение (ПД)
38.
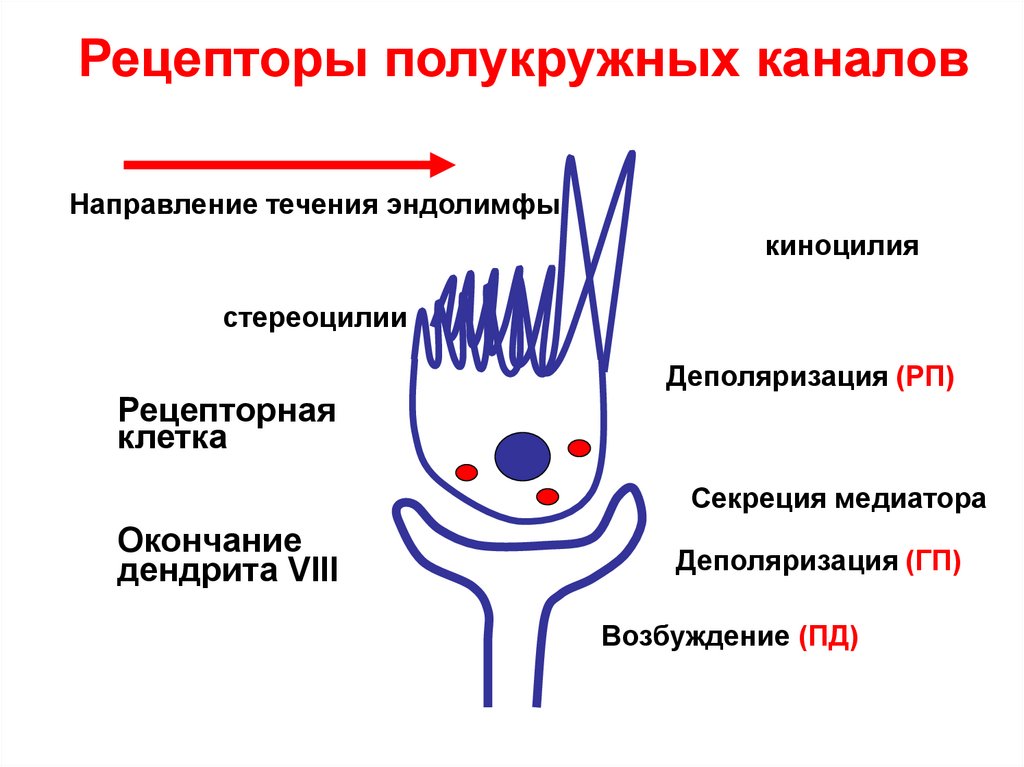
Рецепторы ампул полукружныхканалов расположены в ампулах 3-х
взаимно перпендикулярных каналов.
Адекватным раздражением для
рецепторов полукружных каналов
является вращение с ускорением.
Порог раздражения – 2-30/с2.
Возбуждаются также при согревании
или охлаждении наружного слухового прохода (калорическая проба).
39.
Рецепторы полукружных каналовНаправление течения эндолимфы
киноцилия
стереоцилии
Рецепторная
клетка
Деполяризация (РП)
Секреция медиатора
Окончание
дендрита VIII
Деполяризация (ГП)
Возбуждение (ПД)
40.
Проводниковый отдел1 нейрон – ганглий scarpae,
2 нейрон – нижнее, верхнее, медиальное, интерстициальное ядра
моста,
3 нейрон – таламус.
На уровне ядер моста возможно переключение на двигательные и вегетативные ядра РФ и ч/м нервов.
Возникают вестибуло-моторные и
вестибуло-вегетативные рефлексы.
41.
Вестибуло-моторные рефлексы:• нистагм глаз,
• изменение позы за счет перераспределения мышечного тонуса,
• нарушение координации движений:
а) при пальце-носовой пробе,
б) пробе с почерком,
• нарушение походки.
42.
Вестибуло-вегетативные рефлексы:• изменение ширины зрачка,
• изменение цвета кожи лица (сосудистая реакция),
• тахикардия,
• изменение тонуса и моторики ЖКТ
• и др.
43.
Корковый отделрасположен в нижней части ЗЦИ на
границе с медиальной областью
височной доли.
Формирует вестибуло-сенсорные
реакции:
• ощущения положения
головы и тела в пространстве,
• сенсо-моторные (головокружение)
• сенсо-вегетативные (тошнота)
ощущения.
44.
Проба Барани.Вращение в кресле Барани с закрытыми глазами, голова наклонена к
груди (10 вращений за 20 сек.).
Оценивают до и после вращения:
• вестибуло-моторные,
• вестибуло-вегетативные,
• вестибуло-сенсорные реакции.
45. Моторный отдел двигательной системы.
Моторная зона коры:I зона – в передней
центральной извилине.
II зона – в межполушарной
щели.
46.
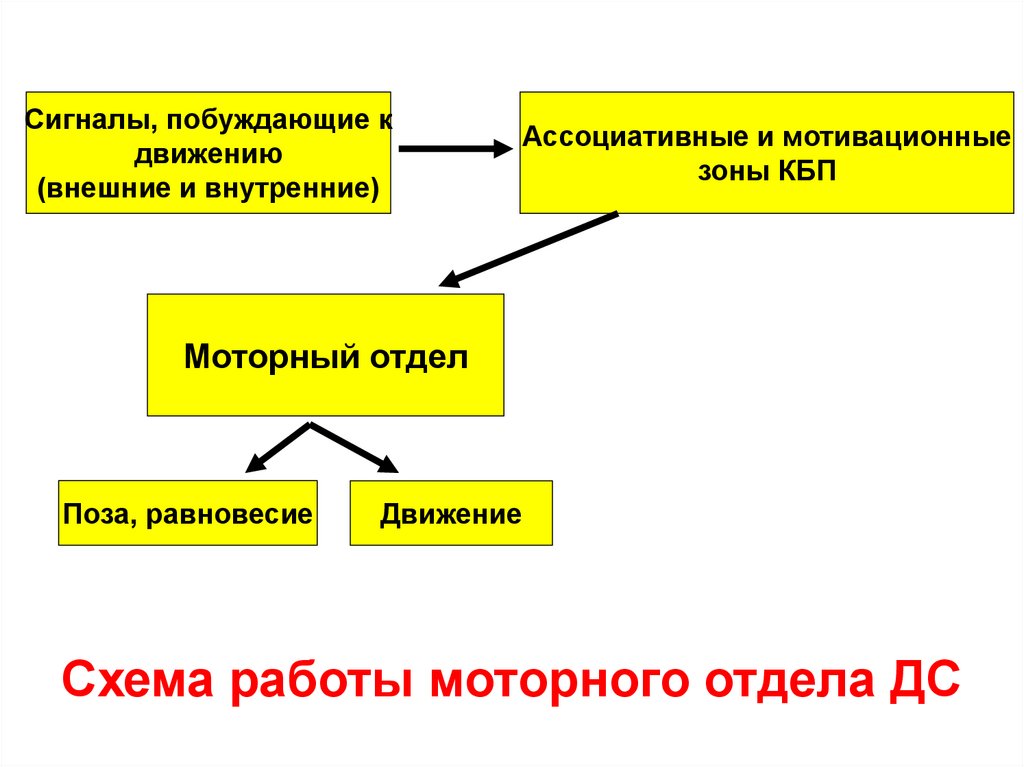
Сигналы, побуждающие кдвижению
(внешние и внутренние)
Ассоциативные и мотивационные
зоны КБП
Моторный отдел
Поза, равновесие
Движение
Схема работы моторного отдела ДС
47. Структурно – функциональная организация МЗК
• 1) Соматотопическая организация– каждая область тела имеет свою область
представительства в ПЦИ.
• 2) Моторный гомункулус площадь
представительства периферических
отделов в ПВИ зависит от выполняемой
функции.
Чем разнообразнее и точнее движения
выполняет группа мышц, тем больше ее
представительство в ПЦИ.
48. 3) Наличие функциональных колонок.
• ФК –это функциональноеобъединение вертикально
расположенных нейронов.
• Действуют одновременно,
активизируют все мышцы,
приводящие сустав в
определенное положение.
49.
50. Нейронная организация МЗК.
• Существует две точки зрения нароль пирамидных клеток МЗК.
• 1. Большие пирамидные клетки
иннервируют фазные
мотонейроны.
Малые – тонические.
• II . Большие – иннервируют α мотонейроны
Малые - γ -мотонейроны
51. Эфферентные связи коры.
52. Пирамидный путь
Коракортикобульбарный
кортикоспинальный Продолговатый мозг
• Быстрое включение
двигательной системы,
произвольная регуляция
тонуса и движений.
Спинной мозг
53. Экстрапирамидная система
Коррекция двигательныхпрограмм в ходе их выполнения:
• а) кортикоталамический –
коррекция позы и движений
• б) кортикоретикулярный:
• 1) ретикулярное ядро ПМ → СМ –
α - МН сгибателей
• 2) ретикулярное ядро ВМ → СМ –
• γ - МН разгибателей
54.
• в) к базальным ганглиям → СМ –стереотипные движения.
• г) к мозжечку (через ядра моста) –
коррекция движений.
• д) кортикорубральный → СМ
повышает тонус сгибателей,
координация тонуса разных групп
мышц.
• е) к нижней оливе →
вестибулоспинальный путь –
повышает тонус разгибателей.
55.
Экстрапирамидные пути связаны сцентрами, в которых записаны
двигательные программы
поддержания позы, равновесия, а
также программы сложных
автоматизированных движений как
врожденных, так и приобретенных, в
том числе - передвижения
в пространстве.
56. Роль структур экстрапирамидной системы
57.
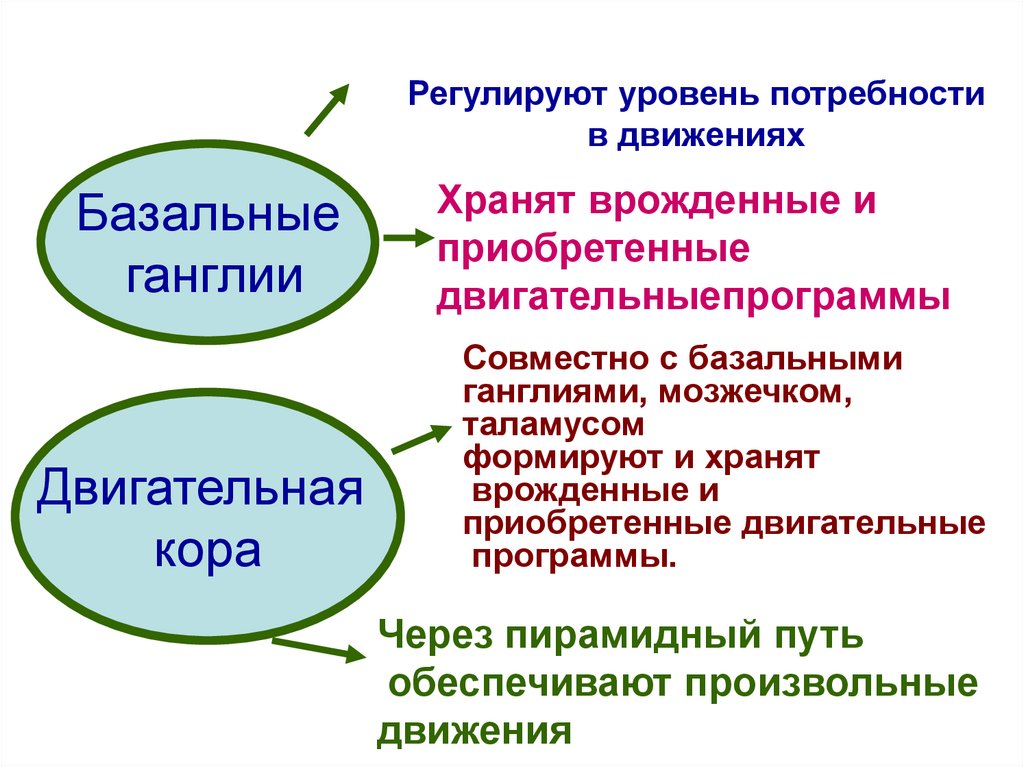
Регулируют уровень потребностив движениях
Базальные
ганглии
Двигательная
кора
Хранят врожденные и
приобретенные
двигательныепрограммы
Совместно с базальными
ганглиями, мозжечком,
таламусом
формируют и хранят
врожденные и
приобретенные двигательные
программы.
Через пирамидный путь
обеспечивают произвольные
движения
58.
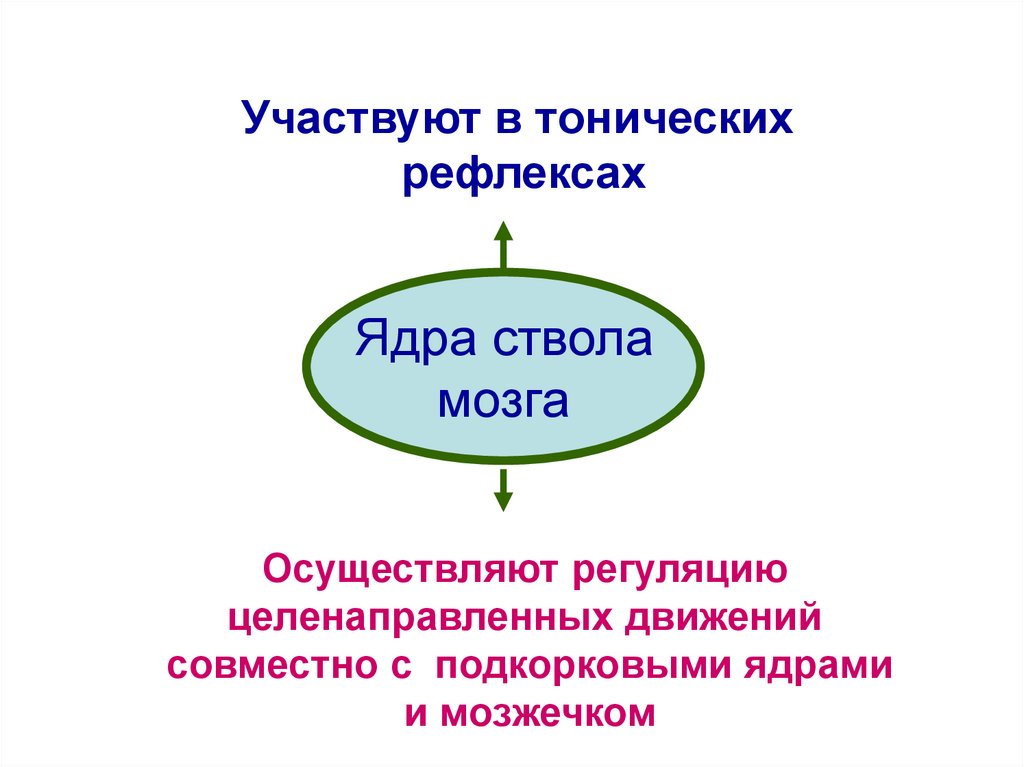
Участвуют в тоническихрефлексах
Ядра ствола
мозга
Осуществляют регуляцию
целенаправленных движений
совместно с подкорковыми ядрами
и мозжечком
59.
Контроль за осуществлениемтонических рефлексов ствола
мозга
Мозжечок
Контроль врожденных
двигательных
программ
и коррекция их по ходу
выполнения
Контроль за произвольными
движениями и приобретенными
двигательными программами.
Коррекция по ходу их выполнения.
60. Управление двигательными функциями на уровне нервного центра спинного мозга.
61.
• Нисходящие влияния моторной зоныкоры заканчиваются на мотонейронах
двигательных центров, расположенных в
спинном мозге, в двигательных ядрах
ЧМН.
• В то же время на уровне двигательных
центров происходят процессы регуляции
тонуса мышц и движений.
• Регуляция тонуса мышц. происходит с
участием проприорецепторов –
мышечного веретена (рецептор
растяжения) и рецептора Гольджи
(возбуждается при сокращении мышцы).
62.
• Мышечныеверетена
(2-3
мм)
образованы
интрафузальными волокнами с ядерной сумкой (или
ядерной
цепочкой).
Располагаются
параллельно
экстрафузальным волокнам.
• К веретену подходит волокно группы Аα, ветвится
внутри веретена. Ветви спирально обвивают
центральную часть каждого интрафузального
волокна.
• Возбуждаются при растяжении мышц. Измеряют длину
мышцы.
• Периферические части интрафузальных волокон
иннервированы γ- мотонейроном
• Экстрафузальные волокна иннервированы α- МН
63.
Экстрафузальныеволокна
γ- мотонейрон иннервирует
Периферические части
интрафузального волокна
α- мотонейрон
иннервирует
экстрафузальные
волокна
афферентный нейрон
интрафузальное
волокно
Иннервация мышцы
64. Функциональная характеристика мышечного веретена (МВ)
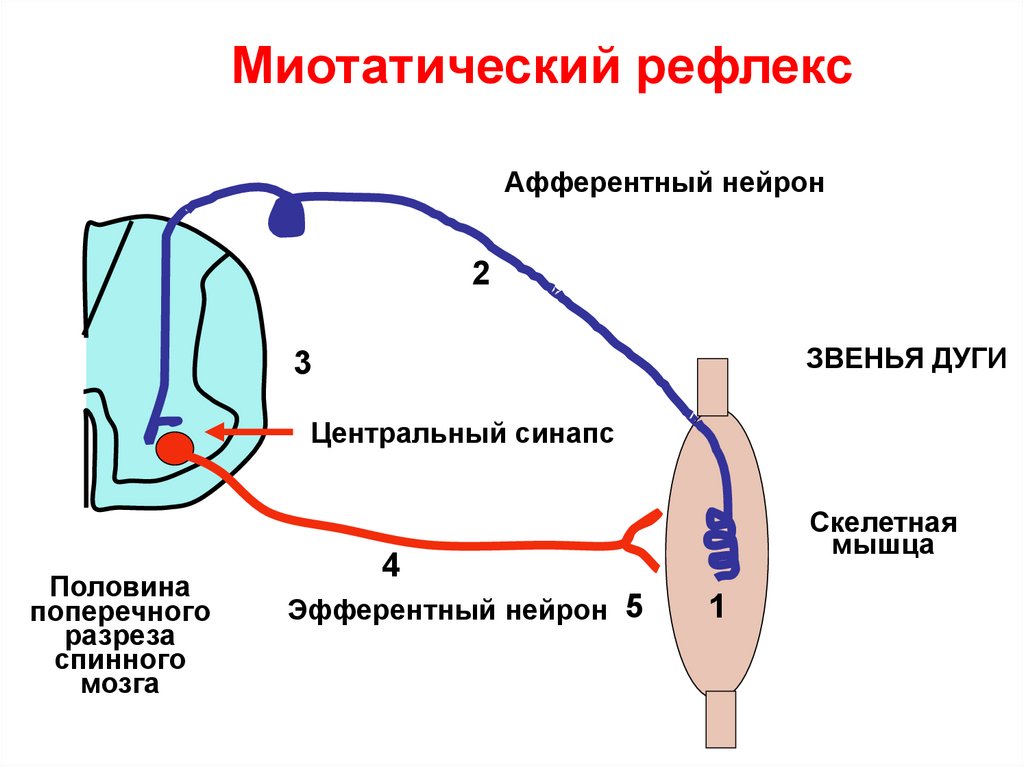
1.При растяжении мышцы.Возникают миотатические
МВ возбуждается рефлексы.
Поддерживают тонус
мышцы. Дуга рефлекса
моносинаптическая.
2.При сокращении периферических
частей мышечного веретена за счет
активации γ – мотонейрона.
65.
Миотатический рефлексАфферентный нейрон
2
ЗВЕНЬЯ ДУГИ
3
Центральный синапс
Половина
поперечного
разреза
спинного
мозга
Скелетная
мышца
4
Эфферентный нейрон 5
1
66. Регуляция тонуса мышцы с участием гамма- мотонейрона
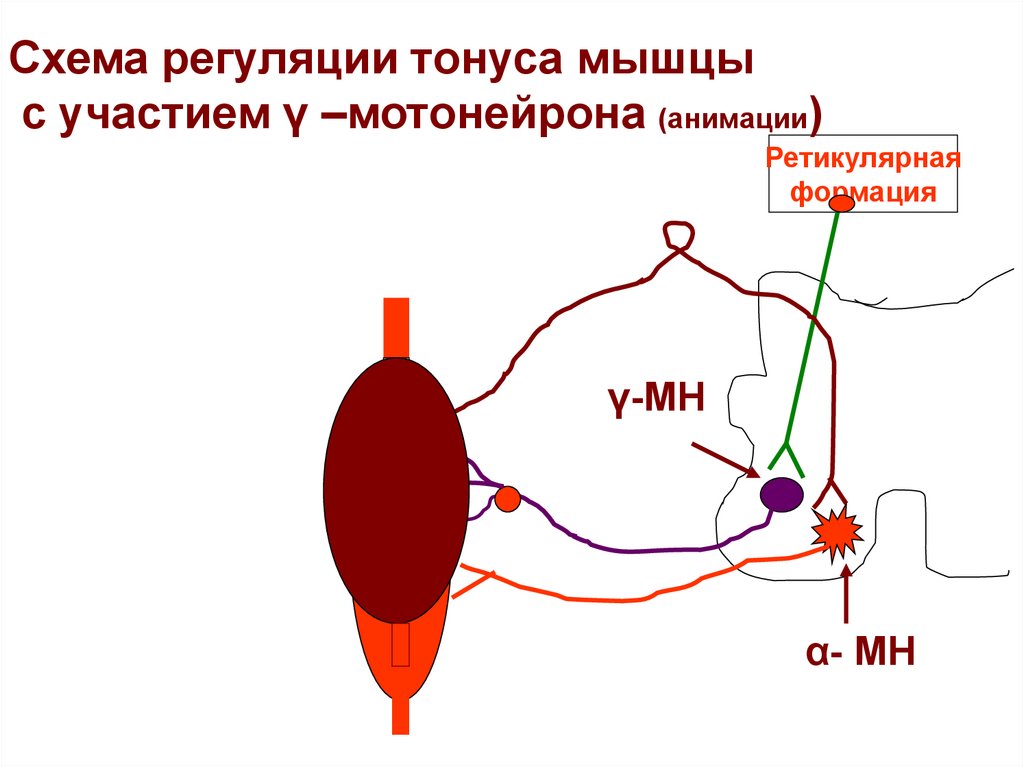
Регуляция тонуса мышцы с участием гаммамотонейрона• События развиваются в следующей последовательности:
• 1. Активация γ- мотонейрона (сигналами из Ретикулярной
формации, коры).
• 2. Сокращение периферических частей интрафузального
волокна.
• 3. При этом ядерная сумка интрафузального волокна
растягивается,
• 4. Афферентный сигнал достигает α -мотонейрона
передних рогов спинного мозга ( моносинаптическая дуга).
• 5. Экстрафузальные мышечные волокна сокращаются.
• Изменение функционального состояния МВ обеспечивает:
1) Поддержание фиксированного положения конечности
2) Автоматическое регулирование силы сокращения в
зависимости от нагрузки
67.
Супраспинальные влияния на гаммамотонейроныγ – мотонейрон.
Мышца в покое
Мышца в покое
68.
Схема регуляции тонуса мышцыс участием γ –мотонейрона (анимации)
Ретикулярная
формация
γ-МН
α- МН
69.
• 2. На уровне нервного центра• осуществляется контроль за
степенью сокращения
мышцы (с рецепторов
Гольджи).
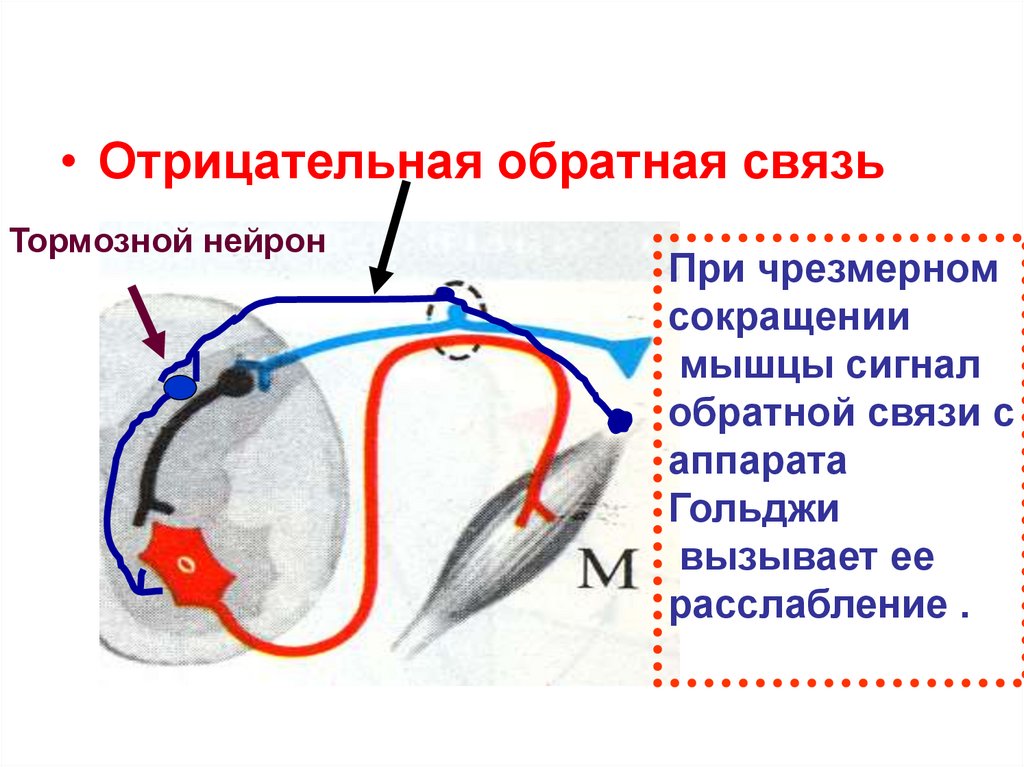
70. Сухожильный рецептор Гольджи
• Образовансухожильными нитями,
отходящими от 10
экстрафузальных
волокон.
• Окружен
соединительнотканной
капсулой.
• Контролирует степень
сокращения мышцы.
• Афферентный путь
Представлен волокнами
группы Аβ.
71.
• Отрицательная обратная связьТормозной нейрон
При чрезмерном
сокращении
мышцы сигнал
обратной связи с
аппарата
Гольджи
вызывает ее
расслабление .
72.
• 3.На уровне нервных центровобеспечиваются защитные
сгибательные рефлексы
• при раздражении рецепторов
кожи тактильных,
температурных, болевых
рецепторов.
73.
• 4. Реализация врожденныхдвигательных программ:
• а)перекрестный разгибательный
рефлекс;
• б) шагательный рефлекс
• В основе лежат реципрокные
отношения между центрами антагонистами
74.
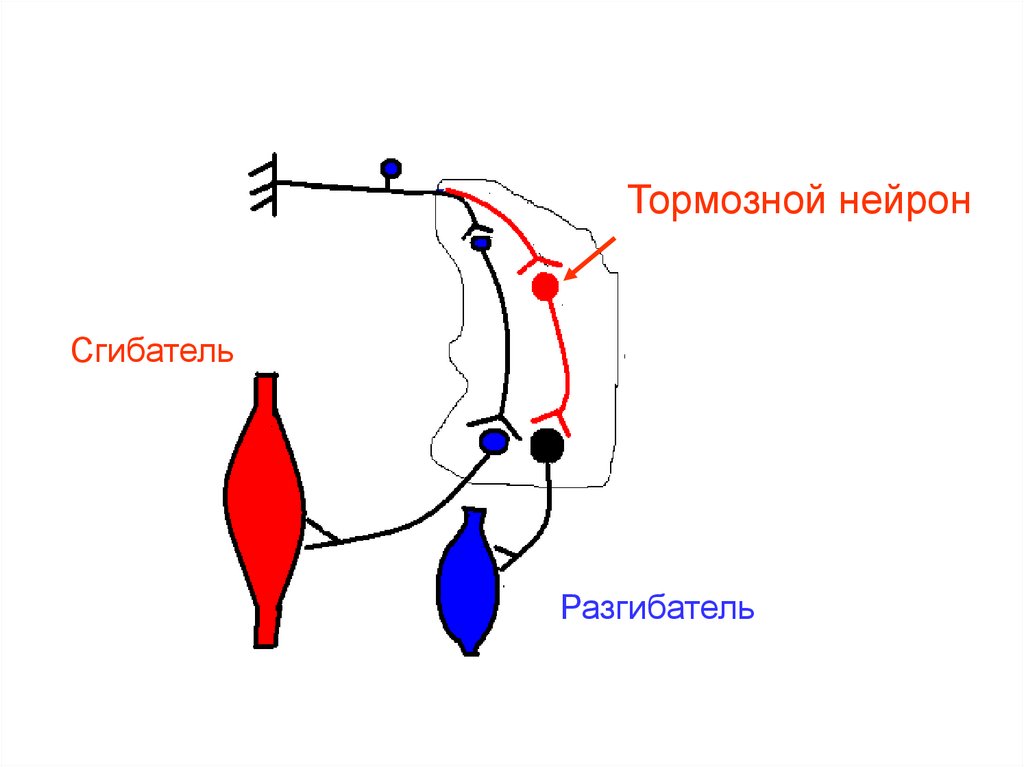
Виды программ:• программа реципрокного торможения
центров антагонистов своей стороны:
возбужден
сгибатель
заторможен
разгибатель
75.
Тормозной нейронСгибатель
Разгибатель
76.
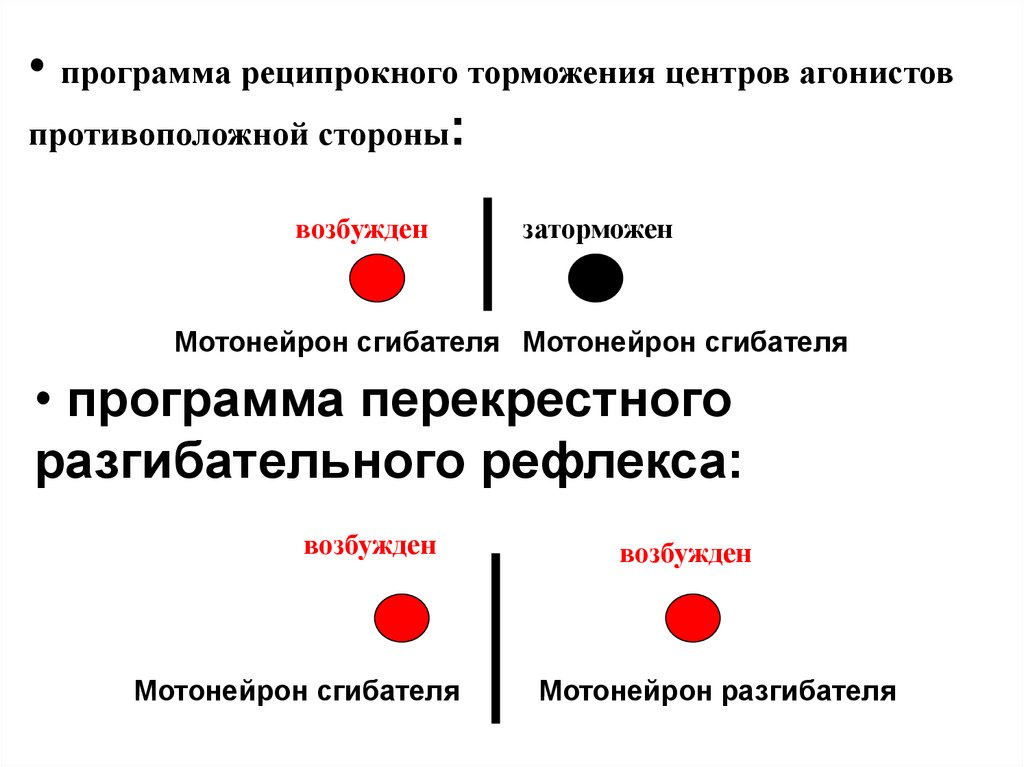
• программа реципрокного торможения центров агонистовпротивоположной стороны:
возбужден
заторможен
Мотонейрон сгибателя Мотонейрон сгибателя
• программа перекрестного
разгибательного рефлекса:
возбужден
Мотонейрон сгибателя
возбужден
Мотонейрон разгибателя
77.
• программа шагательного рефлекса:заторможен
возбужден
сг
разг
Мотонейрон сгибателя
руки
сг
разг
Мотонейрон сгибателя
ноги
разг
сг
Мотонейрон разгибателя
руки
разг
сг
Мотонейрон разгибателя
ноги
78.
Эти программы готовы к работена первом месяце после рождения:
погруженный в воду ребенок плавает. На основе
этих программ ползанье возникает у детей с 6-ти
мес.
Супраспинальный контроль
за программами в ходе их выполнения
осуществляют через гамма-мотонейроны кора БП,
РФ, мозжечок.
Если надо усилить сокращение, стимулируются
гамма-мотонейроны, если надо ослабить –
активность гамма-мотонейронов уменьшается.