


















 и его характеристика")



















 medicine
medicine biology
biologySimilar presentations:


")
")

")




ОФВТ. Вводная. Биопотенциалы
1. ОФВТ. Вводная. Биопотенциалы.
Берг М.Д.Учебная программа по нормальной физиологии включает
252 уч. часа, из них: лекции – 42 (21 лекция), практические и
семинарские занятия – 102, самоподготовка – 108 часов.
ОФВТ. Вводная.
Биопотенциалы.
Профессор Берг М.Д.
2. Кафедра нормальной физиологии, 103 года со дня основания
Организатор кафедры - профессорВериго Бронислав Фортунатович
(1860-1926)
Профессор Берг М.Д.
3.
Основатель кафедры – профессор Бронислав Фортунатович Вериго(1860-1926), ученик И.М.Сеченова. Он положил начало основному
направлению работы пермской школы физиологов - разработке положений
«учения о функциональном состоянии».
Б.Ф.Вериго приехал в Пермь в 1916 году уже состоявшимся ученым, доктором
наук с мировым именем, поработавшим в лучших физиологических
лабораториях Германии, Франции, Швейцарии.
Будучи студентом Петербургского университета, он выполнил работу, которая
поставила имя Вериго в ряд с крупнейшими европейскими физиологами.
Катодическая депрессия Вериго.
Профессор, ученик М.И. Сеченова. Известен в мировой литературе как автор
исследований по электрофизиологии, написал три тома «Основ общей
биологии».
С 1917 года работал в Перми (1917-1926). Несмотря на трудности первой
мировой и гражданской войн, организовал работу кафедры, приобрел в
Стокгольме современное научное и учебное оборудование. В пермский
период жизни подготовил капитальное руководство «Основы физиологии
человека и животных (1918), второе издание вышло в 1924 году.
Кафедра организована 26 декабря 1916 года.
Профессор Берг М.Д.
4.
Physys – природа, logos – учение.Физиология – наука о природе жизненных
процессов.
Мы изучаем физиологию человека.
ПРЕДМЕТ ФИЗИОЛОГИИ:
• функции отдельных клеток, органов и
систем, организма в целом,
• управление ими,
• модулирование (изменение свойств
объекта регуляции и систем регуляции),
• транспортно-метаболическое
обеспечение функций.
Профессор Берг М.Д.
5.
В компетенцию физиологии входят также:• возрастные особенности развития и
управления физиологическими системами и
организмом в целом (ОНТОГЕНЕЗ),
• физиология трудовой деятельности,
• космическая физиология,
• спортивная физиология и др.
Современная физиология изучает эти процессы на уровне клетки и внутриклеточных
структур.
Профессор Берг М.Д.
6.
Использует новые методы клеточной физиологии: наряду смикроэлектродным методом регистрации биопотенциалов
появился оптический метод регистрации потенциалов – на
основе оптогенетики и оптохимии.
Появилась возможность видеть идущие в клетке
биохимические реакции – серийная фемтосекундная
(10-15 с ) кристаллография (SFX), лазерные
нанотехнологии.
На органном и организменном уровне также
используются новые методы исследования: компьютерная
томография, функциональная ЭЭГ, магнитоэнцефалография (МЭГ), функциональная магниторезонансная
томография (МРТ), ультразвуковая допплерография и др.
Но мы на занятиях, конечно, будем пользоваться
традиционными методами классической физиологии
и методами клинической физиологии.
Профессор Берг М.Д.
7.
Метод магнитнорезонансной томографии (МРТ) в сочетаниис.математическим анализом.
Различия формируются после полового созревания.
Профессор Берг М.Д.
8.
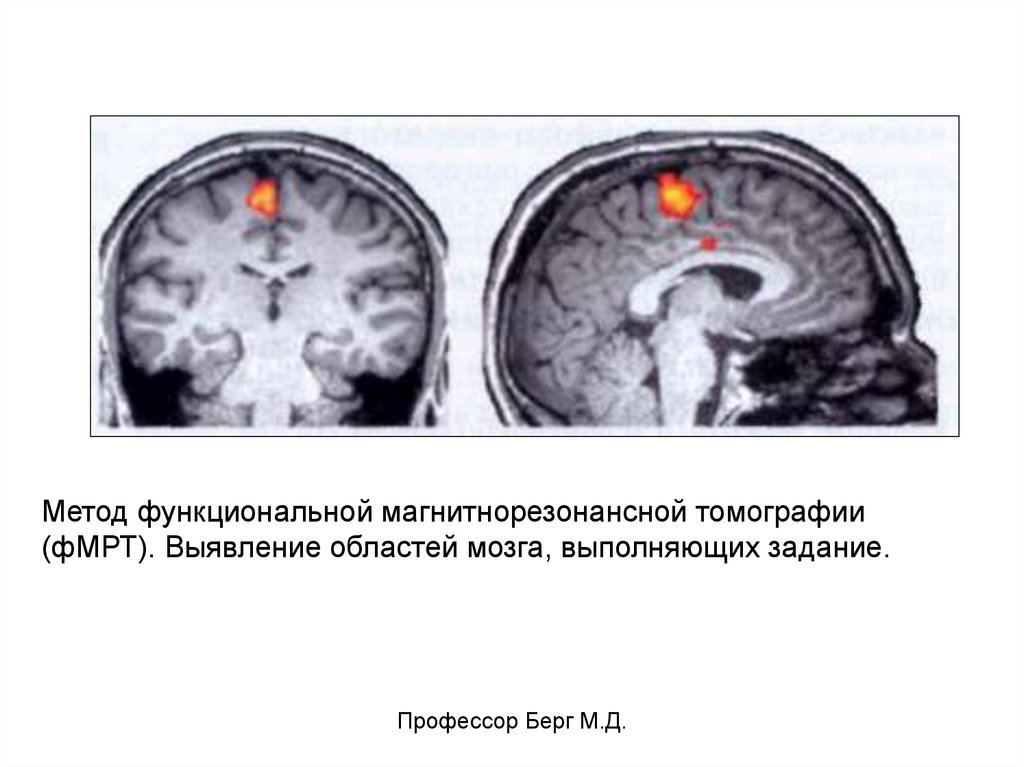
Метод функциональной магнитнорезонансной томографии(фМРТ). Выявление областей мозга, выполняющих задание.
Профессор Берг М.Д.
9.
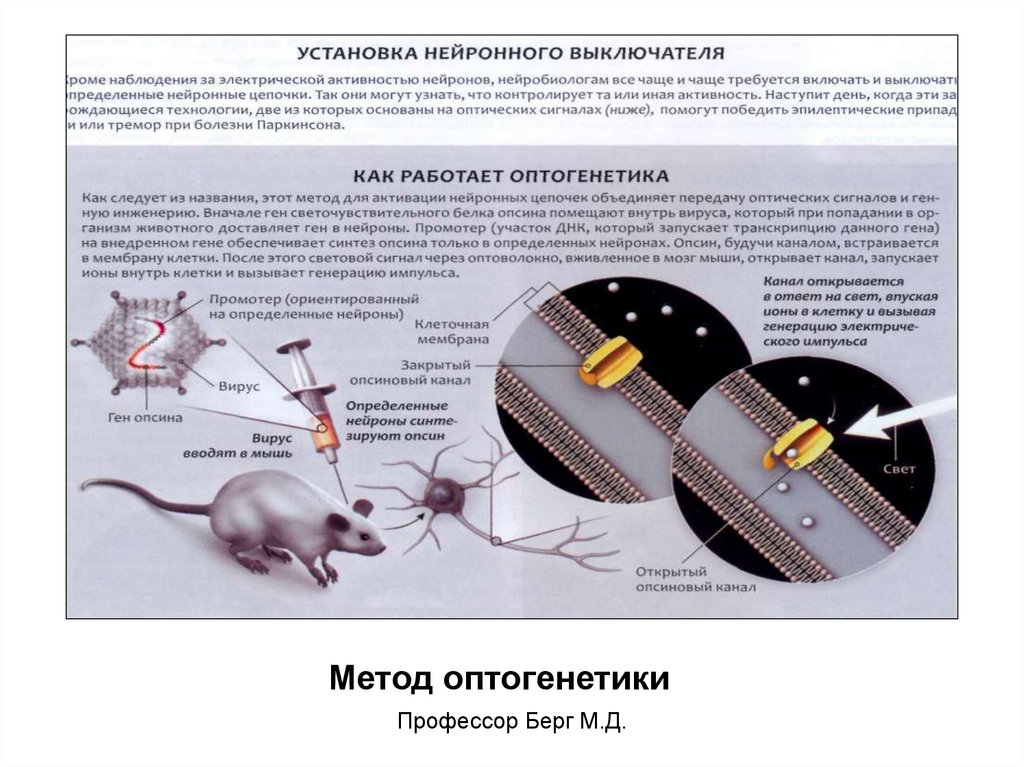
Метод оптогенетикиПрофессор Берг М.Д.
10. Физиологические свойства ткани как основа их деятельного состояния
Профессор Берг М.Д.11.
Наши возможности взаимодействовать сосредой и развиваться, отвечать на воздействия
(на раздражение) связаны с возбудимыми
тканями – нервной, мышечной (скелетная,
сердечная, гладкая) и железистой.
Нервная ткань, вместе с химическими веществами,
циркулирующими с кровью, выступают как системы
управления в организме (нервная и гуморальная
регуляция). Управление осуществляется через белки
мембраны клетки – хеморецепторы и связанные с ними
ионные каналы.
Мышечная и железистая ткани через свои хеморецепторы мембран формируют ответ ткани на управляющие
сигналы благодаря специфическим органеллам клетки.
Профессор Берг М.Д.
12.
Клетка, как самостоятельное государство, имеетмножество внутриклеточных структур и функций
ферментов цитоплазмы, которые активируются
хеморецепторами, расположенными в мембране
клетки. Хеморецепторы связанны с органеллами и
ферментными системами строго определенных
компартаментов клетки, что обеспечивает
целенаправленный ответ клетки.
Хеморецепторы мембран реагируют на сигналы
клеточного выживания, дифференцировки,
апоптоза (клеточной смерти), управления
пролиферацией, метаболизмом, сокращением,
секрецией и, конечно, возбуждением, т.е.
генерацией потенциала действия.
Профессор Берг М.Д.
13.
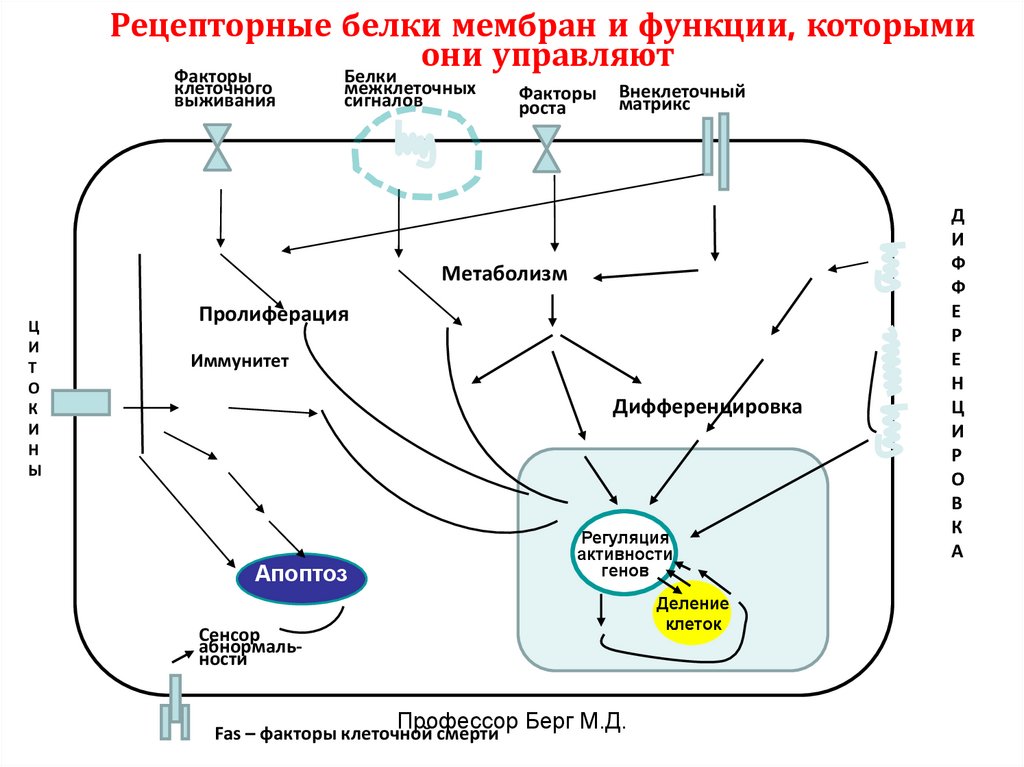
Рецепторные белки мембран и функции, которымиони управляют
Факторы
Белки
клеточного
выживания
межклеточных
сигналов
Факторы
роста
Внеклеточный
матрикс
Метаболизм
Ц
И
Т
О
К
И
Н
Ы
Пролиферация
Иммунитет
Дифференцировка
Регуляция
активности
генов
Апоптоз
Деление
клеток
Сенсор
абнормальности
Профессор Берг М.Д.
Fas – факторы клеточной смерти
Д
И
Ф
Ф
Е
Р
Е
Н
Ц
И
Р
О
В
К
А
14.
В возбудимых тканях большую роль играютбелки межклеточных сигналов, реагирующие
на медиаторы, гормоны, биологически
активные вещества.
Для выполнения функции группы
возбудимых клеток объединяются
в структурно-функциональные единицы, в
которых между клетками есть связующие
контакты, образованные белками
внеклеточного матрикса, и щелевидные
контакты (тесные контакты) для обмена
ионами и веществами. Включаются в работу
эти единицы сигналами от нейронов.
Профессор Берг М.Д.
15.
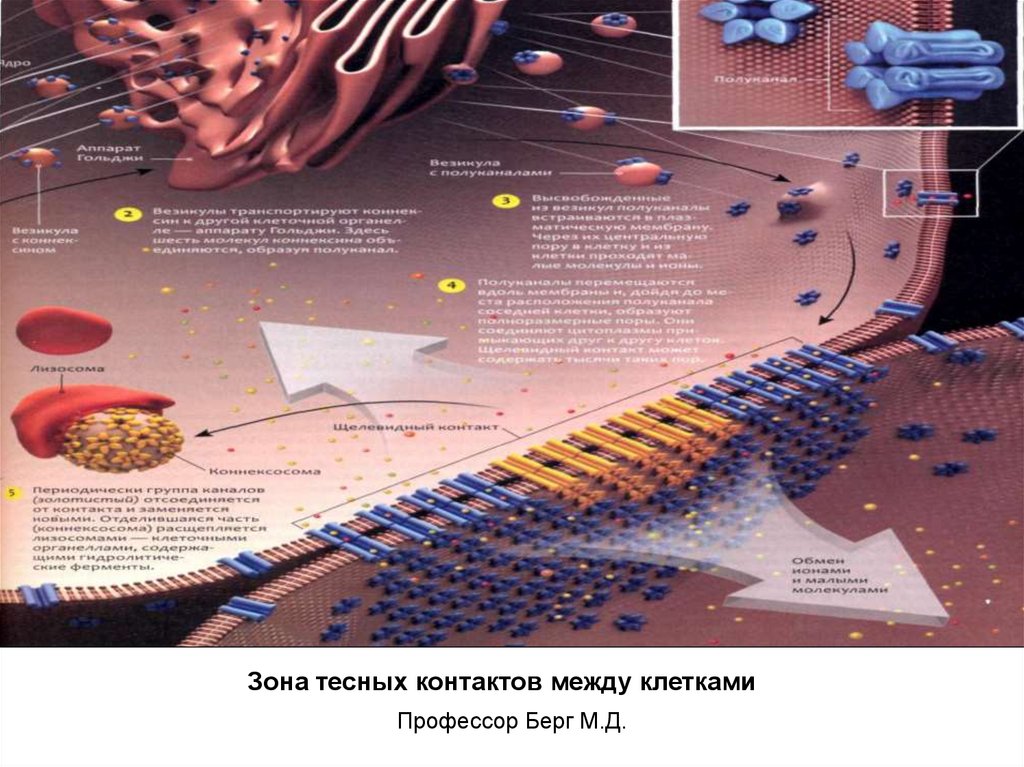
Зона тесных контактов между клеткамиПрофессор Берг М.Д.
16.
Возбудимые клетки характеризуютсяобщими физиологическими свойствами:
• возбудимостью,
• проводимостью,
а также индивидуальными:
- для мышц:
• сократимостью,
• автоматией,
- для железистых клеток:
• способностью к секреции.
Возбудимость – способность отвечать на
раздражение. Она измеряется:
• порогом раздражения (минимальная сила
раздражения, способная вызвать ответ),
• порогом деполяризации (мв),
Профессор Берг М.Д.
17.
а также показателями, которые используются,если раздражителем является постоянный ток:
• реобаза (мв),
• полезное время (мс),
• хронаксия (мс).
Проводимость – способность проводить
возбуждение.
Она измеряется скоростью проведения, м/с.
Нервные волокна – 1-120 м/с (3,6 – 430 км/час),
мышечные волокна – 0,5-4 м/с.
Сократимость – способность изменять длину
или напряжение мышцы (геометрию мышцы).
Измеряется силой (кг), величиной работы (кгм)
или мощности (кгм/мин).
Профессор Берг М.Д.
18.
Автоматия – способность самовозбуждатьсябез внешних воздействий, измеряется в им/мин.
От 1 до 200 в мин.
Секреция – способность синтезировать и
выделять вещества, измеряется в мл/мин.,
г/мин. От 0,1 до 10 мл/мин.
Возбудимость, проводимость и автоматия
связаны со строением и функцией клеточной
мембраны.
Сократимость – с функцией миофибрилл,
содержащих сократительные (актин и миозин) и
модуляторные (тропонин, тропомиозин) белки.
Секреция – с секреторным аппаратом клетки
(эндоплазматический ретикулум, аппарат
Гольджи, рибосомы) и мембраной.
Профессор Берг М.Д.
19.
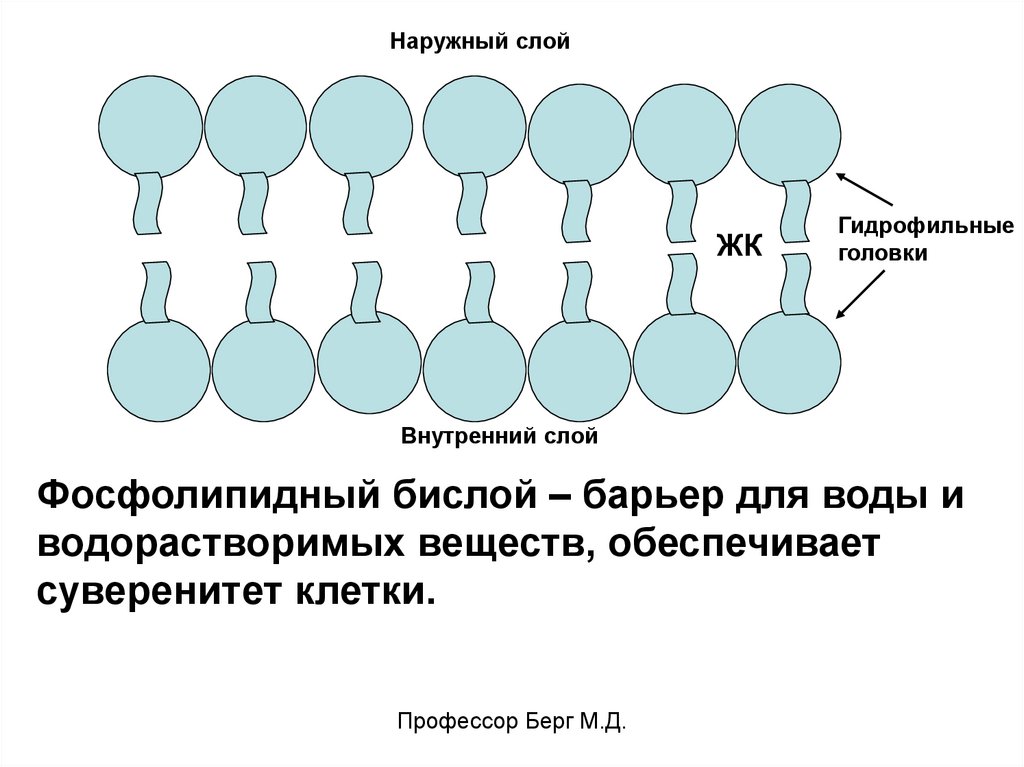
СТРОЕНИЕ И ФУНКЦИИ МЕМБРАНМембрана – оболочка клетки толщиной 6-12 нм.
Образована фосфолипидами и белками.
1. Бислой фосфолипидов.
Гидрофильные головки направлены в водную
среду (фазу) вне и внутри клетки.
Гидрофобные хвосты (жирные кислоты) – во
внутрь мембраны.
Профессор Берг М.Д.
20.
Наружный слойЖК
Гидрофильные
головки
Внутренний слой
Фосфолипидный бислой – барьер для воды и
водорастворимых веществ, обеспечивает
суверенитет клетки.
Профессор Берг М.Д.
21.
2) Белки – интегральные и плавающие (периферические).Функции:
• транспортная:
поры и каналы для пассивного транспорта веществ в клетку и из
клетки;
насосы – для активного переноса веществ.
• рецепторная (для приема и передачи информации в клетку),
• регуляторная (для управления внутриклеточными процессами).
Профессор Берг М.Д.
22. Plasmalemma
Общий вид клеточной мембраныПрофессор Берг М.Д.
23.
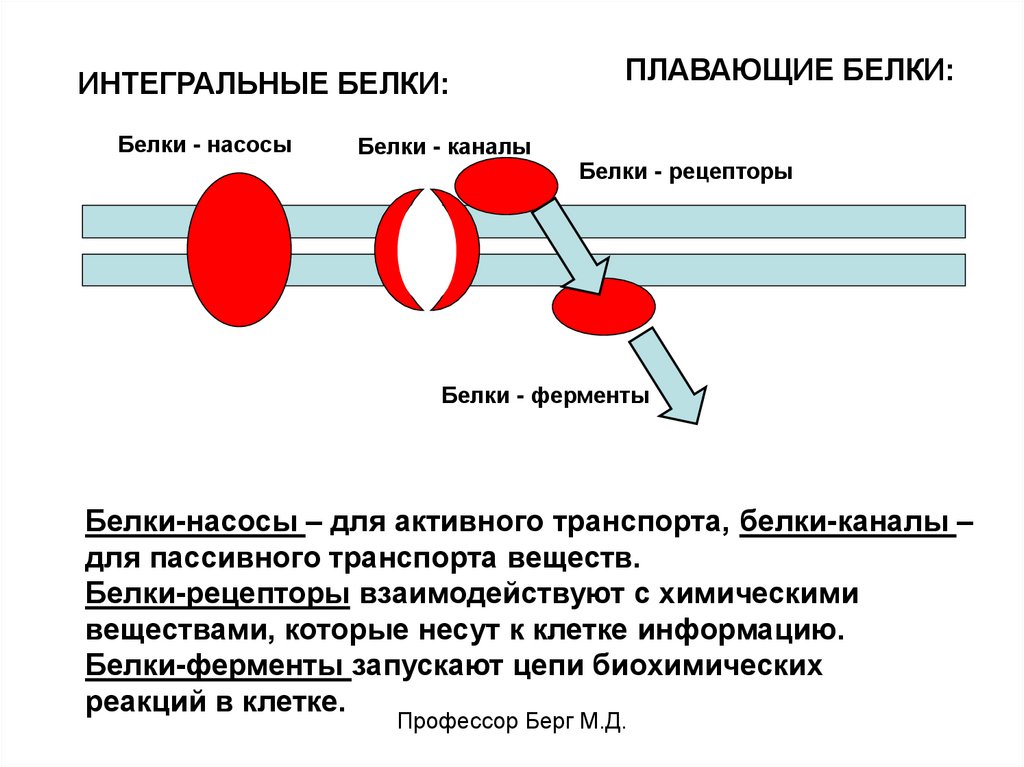
ИНТЕГРАЛЬНЫЕ БЕЛКИ:Белки - насосы
Белки - каналы
ПЛАВАЮЩИЕ БЕЛКИ:
Белки - рецепторы
Белки - ферменты
Белки-насосы – для активного транспорта, белки-каналы –
для пассивного транспорта веществ.
Белки-рецепторы взаимодействуют с химическими
веществами, которые несут к клетке информацию.
Белки-ферменты запускают цепи биохимических
реакций в клетке.
Профессор Берг М.Д.
24.
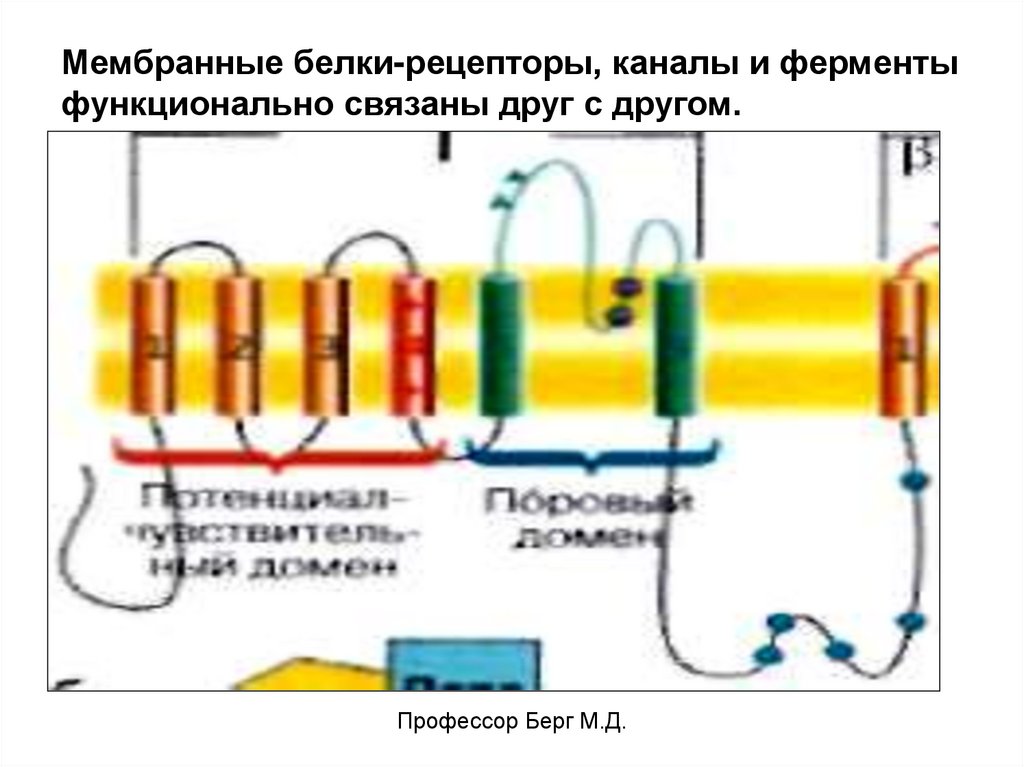
Мембранные белки-рецепторы, каналы и ферментыфункционально связаны друг с другом.
Профессор Берг М.Д.
25.
Ионные каналы необходимы дляформирования заряда мембраны, виды:
К+, Na+, Ca++, CL-.
2 состояния:
- открытые в покое (не управляемые) – К+, Na+,
- закрытые в покое (управляемые) – Na+, Са++,
К+, CL- .
Открываются управляемые каналы
двумя ключами:
химическим (хемозависимые, х/з ),
• электрическим (потенциалзависимые
каналы, п/з).
По скорости работы – быстрые (Na+ п/з) и
медленные (Na+ х/з).
Профессор Берг М.Д.
26.
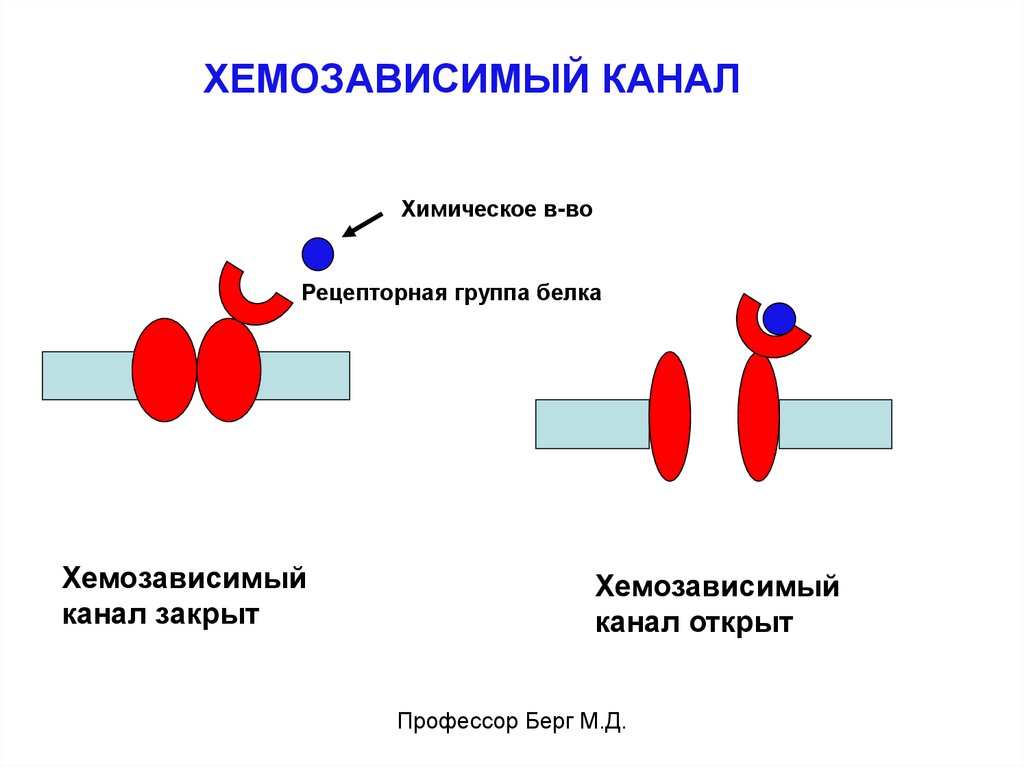
ХЕМОЗАВИСИМЫЙ КАНАЛХимическое в-во
Рецепторная группа белка
Хемозависимый
канал закрыт
Хемозависимый
канал открыт
Профессор Берг М.Д.
27.
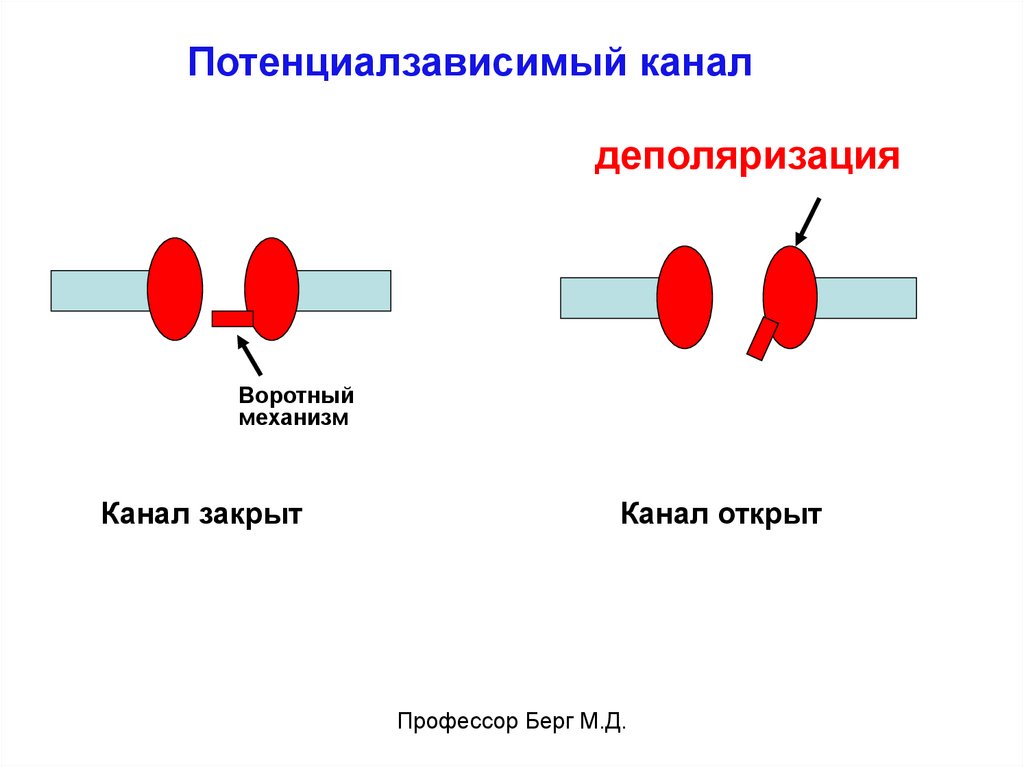
Потенциалзависимый каналдеполяризация
Воротный
механизм
Канал закрыт
Канал открыт
Профессор Берг М.Д.
28.
Ионные насосы – белки-переносчики.В плазматической мембране – это натрийкалиевый насос (Na+/K+-насос) и кальциевый
(Са++-насос).
Кроме транспорта ионов они могут
осуществлять котранспорт и антитранспорт
(антипорт) других веществ (глюкоза,
аминокислоты).
Для транспорта воды в мембране могут быть
специальные водные поры – белки
аквапорины (например, в почках).
Профессор Берг М.Д.
29.
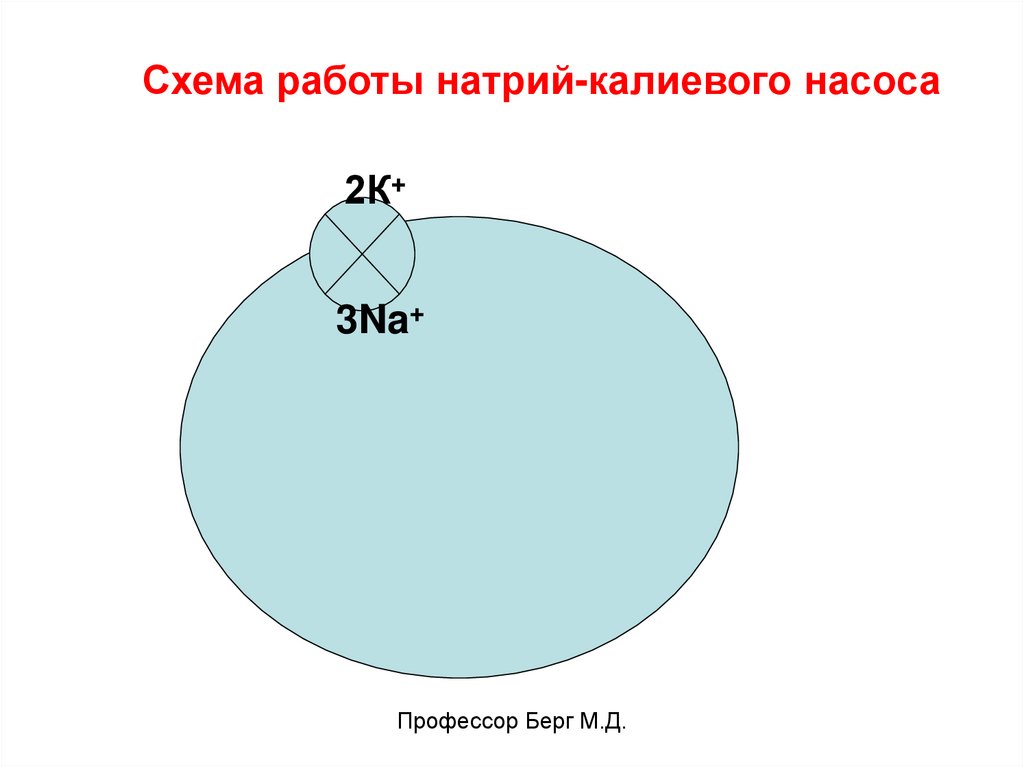
Схема работы натрий-калиевого насоса2К+
3Na+
Профессор Берг М.Д.
30. Потенциал покоя (ПП) и его характеристика
Профессор Берг М.Д.31.
В покое плазматическая мембрана возбудимыхклеток поляризована: снаружи она заряжена
«+», внутри – «-».
Эта поляризация называется потенциалом
покоя (ПП).
ПП – трансмембранная разница заряда
плазматической мембраны.
ПП = - 30-90 мв (в разных клетках).
ПП регистрируют путем введения внутриклеточного микроэлектрода.
Профессор Берг М.Д.
32.
Микроэлектрод вводят через мембрану в клетку ирегистрируют заряд внутри клетки
(микроэлектродная техника).
КCL
0,5 мкм
клетка
-
Профессор Берг М.Д.
Регистрирующий
прибор
(УБП, ОСЦГ)
33.
Возникновение ПП объясняетмембранно-ионная теория (БернштейнЧаговец-Ходжкин-Хаксли-Кац).
Причины возникновения ПП:
1) Разница концентраций ионов в клетке и вне
ее.
Концентрация К+ в клетке в 30 раз больше, чем
вне ее. Концентрация Na+ к клетке в 10 раз
меньше, чем вне ее.
2) Полупроницаемость мембраны (разная
проницаемость для ионов).
В покое в мембране открыто много ионных
каналов для К+ и небольшое количество – для
Na+.
Профессор Берг М.Д.
34.
В результате в покое происходит диффузияионов К+ через открытые неуправлемые
каналы из клетки наружу за счет разницы
концентраций. Одновременно – небольшая
диффузия ионов Na+ в клетку.
Профессор Берг М.Д.
35.
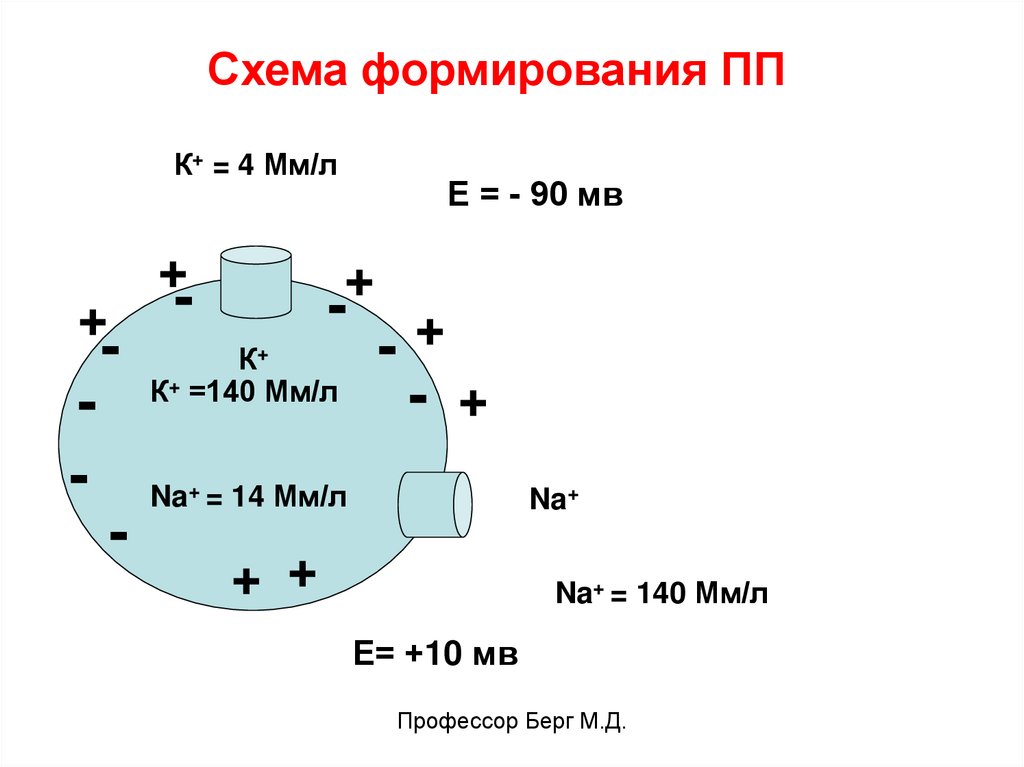
Схема формирования ППК+ = 4 Мм/л
+
+
-
-
Е = - 90 мв
+
-
К+
К+ =140 Мм/л
+
-
- +
Na+ = 14 Мм/л
Na+
+ +
Na+ = 140 Мм/л
Е= +10 мв
Профессор Берг М.Д.
36.
Ионы К+ заряжают наружную сторону мембраны «+», а во внутренней среде создаетсяизбыток отрицательных ионов (CL-, белки).
Калиевый поток создает потенциал:
Ек+ = -90 мв
Натриевый поток в клетку создает потенциал,
равный +10 мв (ЕNa+ = +10 мв).
ЕПП = -90 +10 = -80 мв
В разных тканях плотность К+ и Na+ каналов не
одинакова, поэтому величина ПП различается:
• скелетные мышцы – (-60-90) мв,
• нейроны – (-50-80) мв,
• сердце – (-85-90) мв,
• гладкие мышцы – (-30-70) мв,
• железистые клетки – -30 мв.
Профессор Берг М.Д.
37.
Для поддержания постоянства концентрацииионов в клетке (ионный гомеостаз) в мембране
существуют ионные насосы – Na+/К+-насос.
Он перекачивает в клетку 2 иона К+ и откачивает
из клетки 3 иона Na+. Работает против градиента
концентрации, использует энергию АТФ.
Кальциевый насос (гладкая мышца) выкачивает
ионы Са++ из клетки, тоже использует энергию
АТФ.
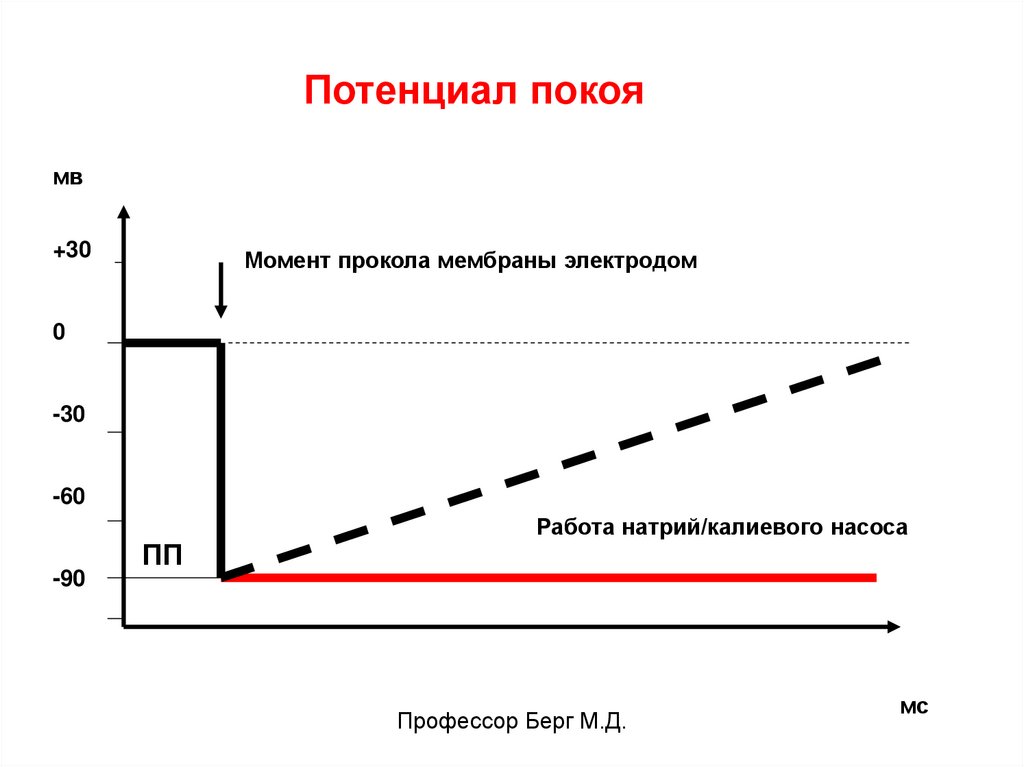
Ниже показана регистрация ПП микроэлектродным методом. В условиях работы натрий-калиевого насоса ПП сохраняет свою величину
(красная линия), при отключении насоса пунктирная линия.
Профессор Берг М.Д.
38.
Потенциал покоямв
+30
Момент прокола мембраны электродом
0
-30
-60
-90
ПП
Работа натрий/калиевого насоса
Профессор Берг М.Д.
мс
39.
Термины:• поляризация мембраны,
• деполяризация мембраны,
• гиперполяризация мембраны,
• реполяризация мембраны.
Профессор Берг М.Д.
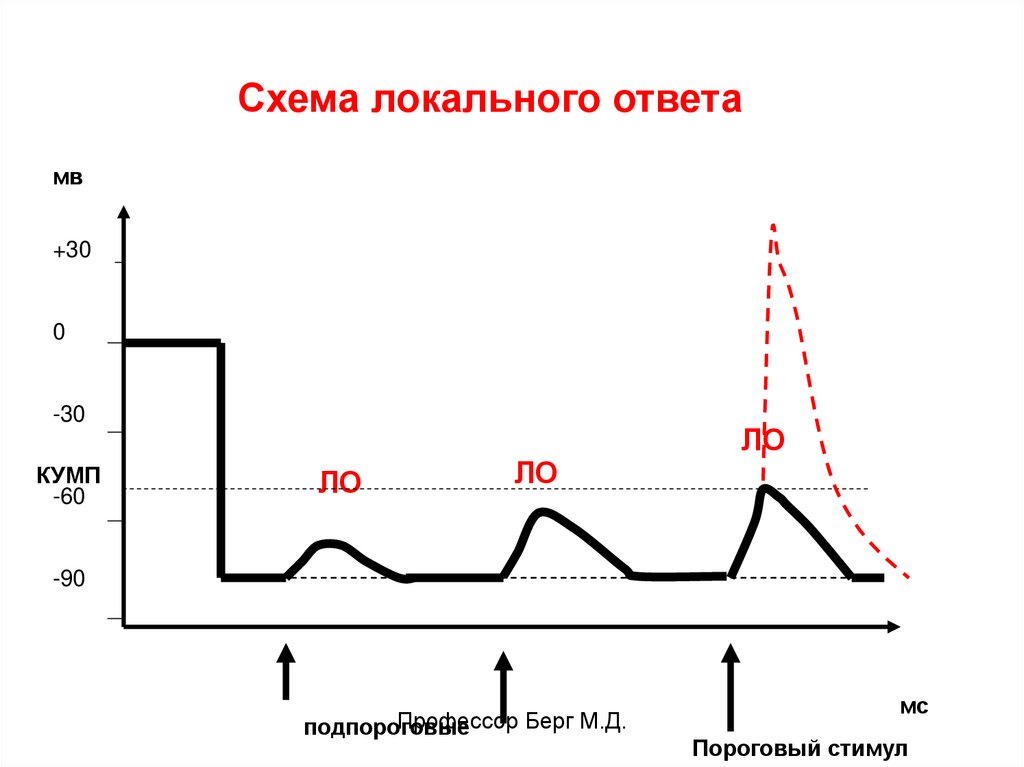
40. Ответ клетки на раздражение
Ответ клетки на раздражение зависит отсилы раздражения.
На подпороговые раздражения она дает
локальный ответ (ЛО), на пороговые и
сверхпороговые – распространяющийся
ответ в виде потенциала действия (ПД).
ЛО – местный, не распространяющийся
ответ, способный к суммированию, зависит
от величины подпорогового стимула.
Связан с открытием хемозависимых Na+каналов, не подчиняется закону «все или
ничего».
Профессор Берг М.Д.
41.
Схема локального ответамв
+30
0
-30
КУМП
-60
ЛО
ЛО
ЛО
-90
Профессор Берг М.Д.
подпороговые
мс
Пороговый стимул
42.
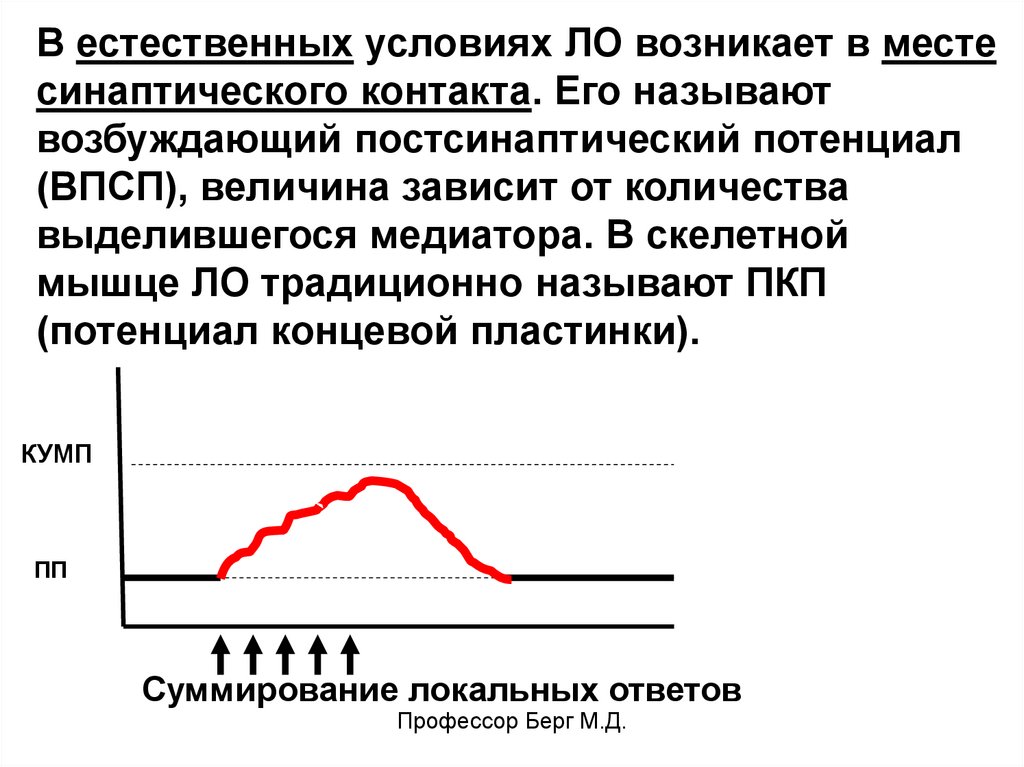
В естественных условиях ЛО возникает в местесинаптического контакта. Его называют
возбуждающий постсинаптический потенциал
(ВПСП), величина зависит от количества
выделившегося медиатора. В скелетной
мышце ЛО традиционно называют ПКП
(потенциал концевой пластинки).
КУМП
ПП
Суммирование локальных ответов
Профессор Берг М.Д.
43.
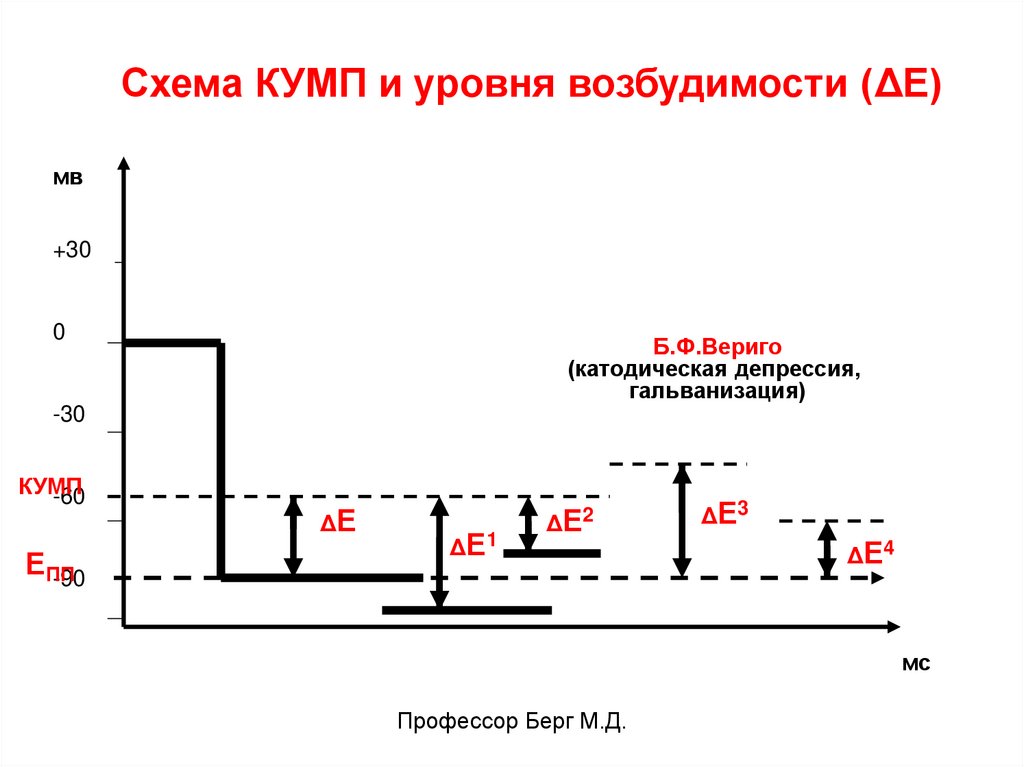
КУМП – уровень деполяризации мембраны, прикотором открываются потенциалзависимые
быстрые Na+-каналы, в результате возникает
возбуждение (ПД).
КУМП находится на уровне -30-60 мв.
Разница между ПП и КУМП называется порогом
деполяризации (ΔЕ). К этой величине
деполяризации чувствительны п/з ионные
каналы для Na+.
Он (ΔЕ) характеризует возбудимость клетки.
При изменении величины КУМП или ПП
изменяется величина ∆Е, т.е. уровень
возбудимости клетки.
Профессор Берг М.Д.
44.
Схема КУМП и уровня возбудимости (ΔЕ)мв
+30
0
Б.Ф.Вериго
(катодическая депрессия,
гальванизация)
-30
КУМП
-60
ЕПП
-90
ΔЕ
Δ Е1
Δ Е2
ΔЕ 3
Δ Е4
мс
Профессор Берг М.Д.
45. Ответ ткани на пороговые и сверхпороговые раздражения
Профессор Берг М.Д.46.
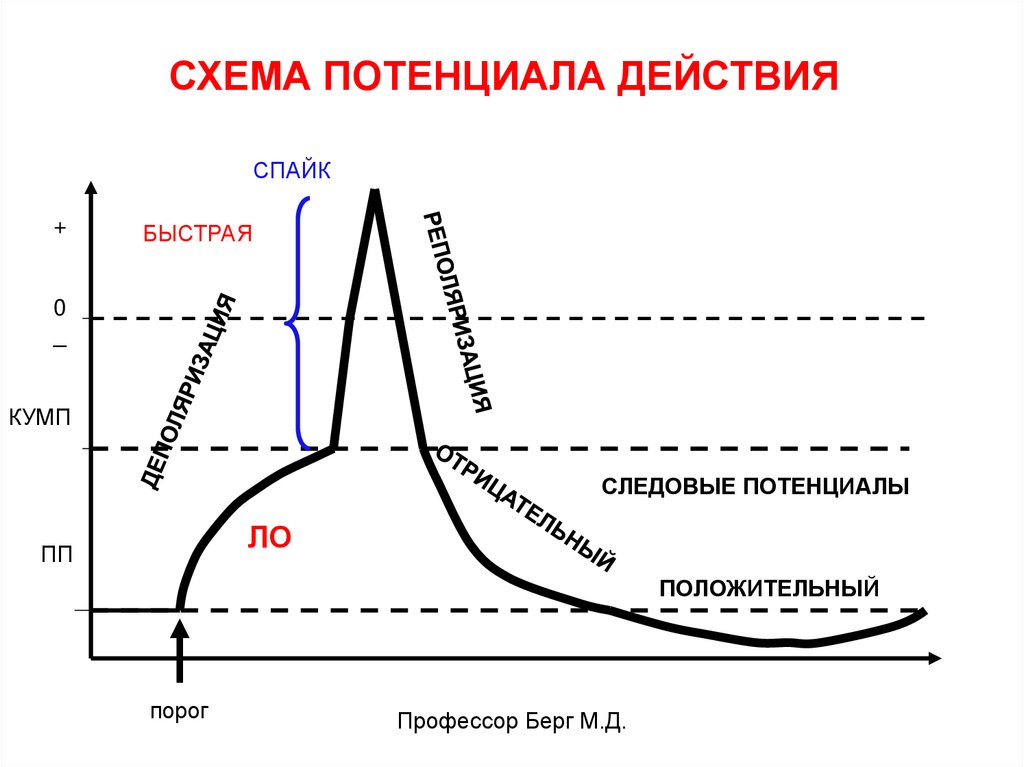
В ответ на пороговый или сверхпороговыйстимул возникает кратковременное изменение
заряда мембраны (от 0,003 до 0,3 с), которое
называют потенциалом действия (ПД).
Пороговый раздражитель вызывает
деполяризацию, достигающую КУМПа.
При этом открываются п/з Nа+ каналы, через
них в клетку лавинообразно входят ионы Na+.
Они вызывают неуправляемую
деполяризацию - ПД.
Профессор Берг М.Д.
47.
Деполяризация сначала достигает «0», а затеммембрана перезаряжается на противоположный
знак – «+» (инверсия заряда).
После этого заряд мембраны возвращается к
исходному уровню.
Восстановление заряда мембраны называется
реполяризацией, которая сначала идет быстро,
а потом – медленно (следовые потенциалы).
Профессор Берг М.Д.
48.
СХЕМА ПОТЕНЦИАЛА ДЕЙСТВИЯСПАЙК
+
БЫСТРАЯ
0
_
КУМП
СЛЕДОВЫЕ ПОТЕНЦИАЛЫ
ЛО
ПП
ПОЛОЖИТЕЛЬНЫЙ
порог
Профессор Берг М.Д.
49.
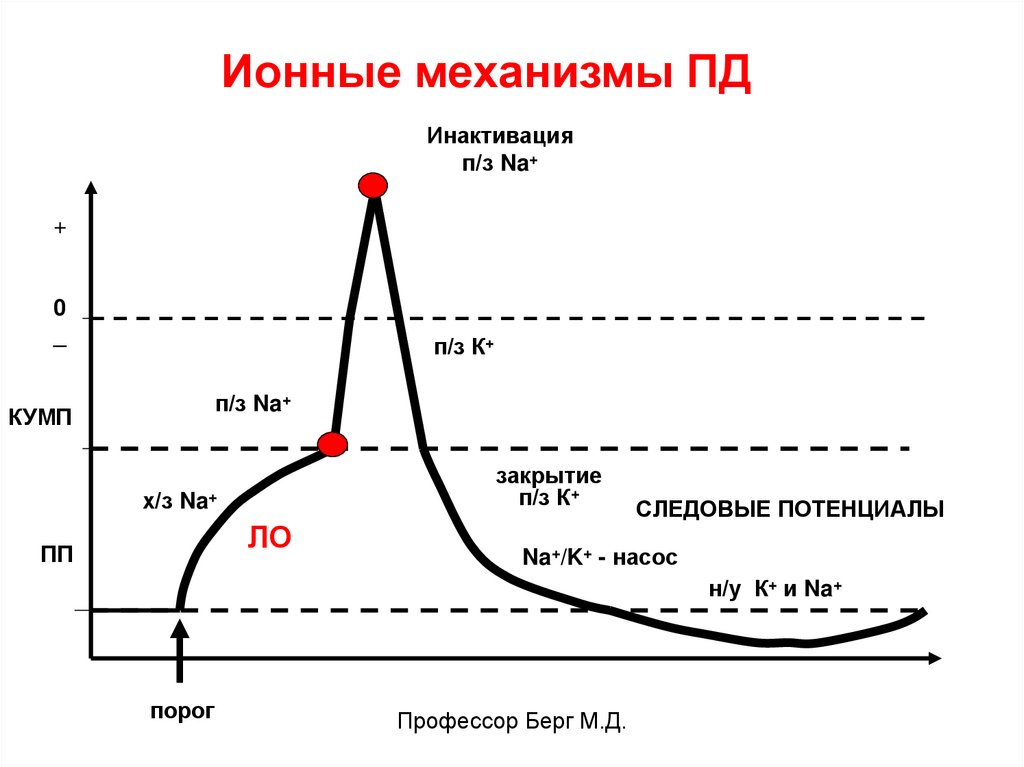
Ионные механизмы ПДИнактивация
п/з Na+
+
0
_
п/з К+
п/з Na+
КУМП
закрытие
п/з К+
х/з Na+
ЛО
ПП
СЛЕДОВЫЕ ПОТЕНЦИАЛЫ
Na+/K+ - насос
н/у К+ и Na+
порог
Профессор Берг М.Д.
50.
Свойства импульсного ответа:1. Возникает на пороговые и сверхпороговые
раздражения.
2. Подчиняется закону «всё или ничего».
3. Распространяется по мембране без
затухания.
Профессор Берг М.Д.
51.
ЛОКАЛЬНЫЙ И ИМПУЛЬСНЫЙ ОТВЕТмембрана
Профессор Берг М.Д.
52.
Закон «все или ничего»Импульсный ответ (ПД)
Сила раздражения
порог
Профессор Берг М.Д.
53.
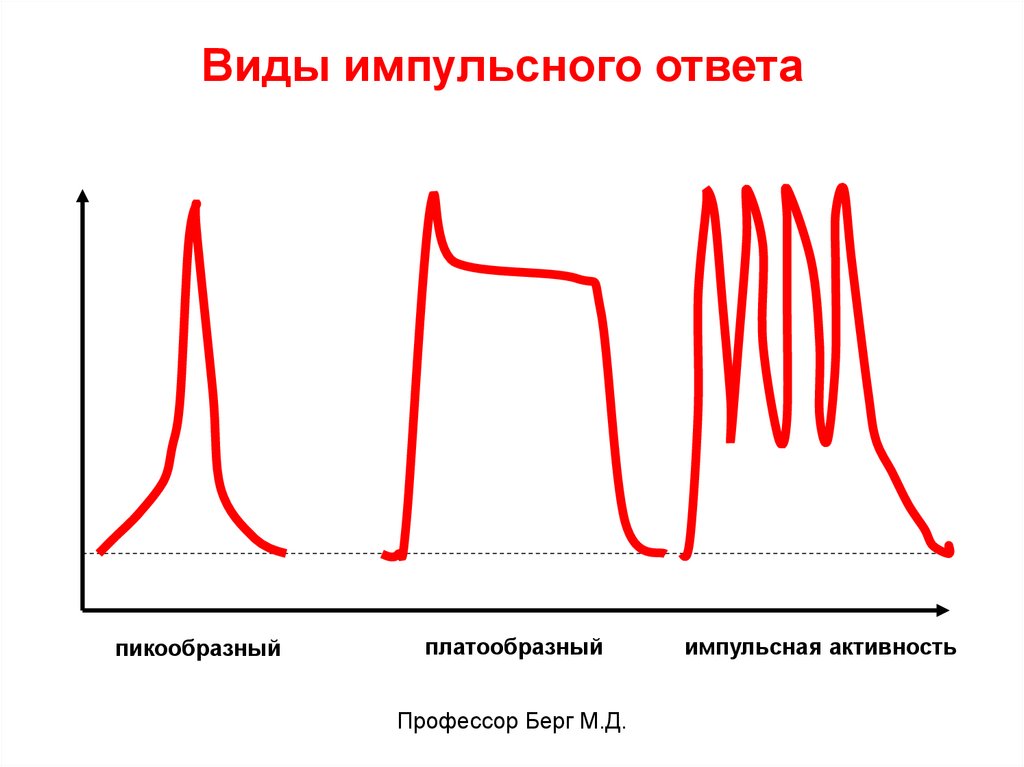
Виды импульсного ответапикообразный
платообразный
Профессор Берг М.Д.
импульсная активность
54.
Схема движения ионов Nа+ЛО
х/з
ПД
п/з
Na+
Na+
Профессор Берг М.Д.
55.
Методы регистрации ПД1. Внутриклеточное отведение микроэлектродом.
2. Внеклеточное отведение макроэлектродами:
• Электромиография
• Электронейрография
• Электрокардиография
• Электроэнцефалография
• Электрогастрография
Профессор Берг М.Д.
56.
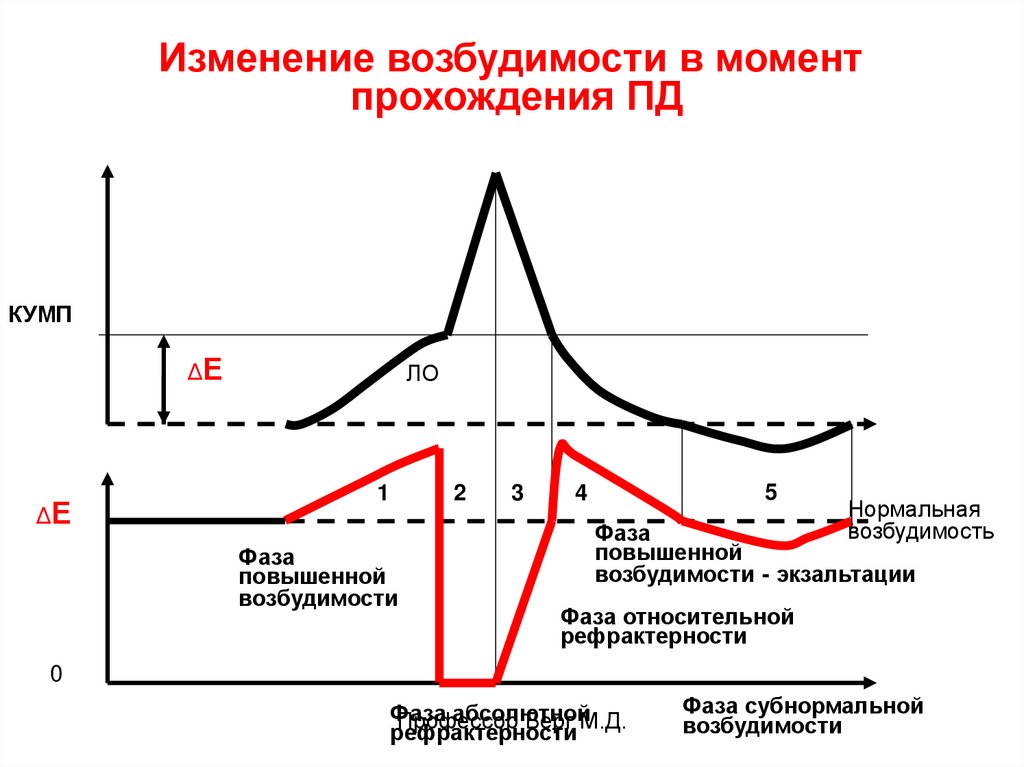
Изменение возбудимости при ПДВ период ЛО возбудимость повышена - 1.
В фазу деполяризации – снижена до 0
(абсолютная рефрактерность) - 2.
В фазу быстрой реполяризации – понижена
(относительная рефрактерность) - 3.
В период отрицательного следового
потенциала – повышена (фаза экзальтации) - 4,
В период положительного следового
потенциала – понижена (фаза субнормальной
возбудимости) - 5.
Профессор Берг М.Д.
57.
Изменение возбудимости в моментпрохождения ПД
КУМП
ΔЕ
ΔЕ
ЛО
1
2
Фаза
повышенной
возбудимости
3
5
4
Нормальная
возбудимость
Фаза
повышенной
возбудимости - экзальтации
Фаза относительной
рефрактерности
0
Фаза
абсолютной
Профессор
Берг М.Д.
рефрактерности
Фаза субнормальной
возбудимости
58.
1. В фазу повышенной возбудимости клеткаотвечает на подпороговые раздражения.
2. В фазу абсолютной рефрактерности не
реагирует даже на сверхпороговые
раздражения.
3. и 5. В фазу пониженной возбудимости
отвечает только на сверхпороговые
раздражения.
4. В фазу экзальтации отвечает на
подпороговые раздражения.
Профессор Берг М.Д.