










.")








")





 и")



")




")


")
")












 medicine
medicine biology
biologySimilar presentations:








")

Нормальная физиология. Лекция 1
1. Лекции по нормальной физиологии
Лектордоцент кафедры нормальной
физиологии
Тютюнщикова Валентина Дмитриевна
2.
• Кафедра нормальной физиологии ПГМУ организована 26декабря 1916 года
• Основатель кафедры – профессор Бронислав
Фортунатович Вериго (1860-1926), ученик И.М.Сеченова. С
1917 по 1926 год работал в Перми. Б.Ф. Вериго приехал в
Пермь уже состоявшимся ученым, доктором наук с мировым
именем, поработавшим в лучших физиологических
лабораториях Германии, Франции, Швейцарии
• Несмотря на трудности первой мировой и гражданской войн,
организовал работу кафедры, приобрел в Стокгольме
современное научное и учебное оборудование Он положил
начало основному направлению работы пермской школы
физиологов - разработке «учения о функциональном
состоянии».
3.
• Будучи студентом Петербургского университета, онвыполнил работу, которая поставила имя Вериго в
ряд с крупнейшими европейскими физиологами.
Вериго Б.Ф. обнаружил катодическую депрессию,
названную его именем, открыл, эффект Вериго по
насыщению крови О2 в зависимости от напряжения
СО2.
• Вериго Б.Ф. Известен в мировой литературе как
автор исследований по электрофизиологии, написал
три тома «Основ общей биологии».
• В пермский период жизни подготовил капитальное
руководство «Основы физиологии человека и
животных» (1918). Второе издание вышло в 1924
году.
4. Физиология
• – наука о функциях и процессах в организмеили его составляющих (системах, органах,
клетках) и механизмах их регуляции,
• обеспечивающих жизнедеятельность при
взаимодействии с окружающей средой.
• Роль физиологии:
• Формирует понимание и дает знания о:
• - методах оценки уровня здоровья
• - способах поддержания уровня здоровья
• - об условиях, необходимых для
обеспечения здоровья.
5.
Области ФизиологииОбщая
Прикладная
Частная
Эволюционная
Сравнительная
6. Прикладная физиология
клиническаяэкологическая
авиационная
космическая
возрастная
труда
психофизиология
7. Методы исследований в физиологии
• наблюдение,• эксперимент острый и хронический
(метод экстирпации, трансплантации,
фистульный метод др.),
• функциональные нагрузки,
• клинико-физиологические,
• лабораторные,
• аналитическое исследование,
• математическое моделирование
8.
Использует новые методы клеточной физиологии: наряду смикроэлектродным методом регистрации биопотенциалов
появился оптический метод регистрации потенциалов – на
основе оптогенетики и оптохимии.
Появилась возможность видеть идущие в клетке
биохимические реакции – серийная фемтосекундная
(10-15 с ) кристаллография (SFX), лазерные
нанотехнологии.
На органном и организменном уровне также
используются новые методы исследования: компьютерная
томография, функциональная ЭЭГ, магнитоэнцефалография (МЭГ), функциональная магниторезонансная
томография (МРТ), ультразвуковая допплерография и др.
Но мы на занятиях, конечно, будем пользоваться
традиционными методами классической физиологии
и методами клинической физиологии.
Профессор Берг М.Д.
9.
Оптогенетика — методика исследования работы нервных клеток,основанная на внедрении в их мембрану специальных белков — опсинов,
которые встраиваются в мембрану клетки и активируются светом
(отсюда «опто»). Такие белки (опсины) есть у большинства животных
в сетчатке глаз, а также у некоторых растений, например у зеленых
водорослей.
Если на мозг воздействовать светом с определенной длиной волны, то те
нейроны, которые имеют такие каналы, будут активироваться или, наоборот,
не смогут генерировать потенциалы действия.
Для экспрессии каналов используются методы генной инженерии, для
последующей активации либо ингибирования нейронов и нервных
сетей используются лазеры, оптоволокно и другая оптическая аппаратура.
Оптогенетика появилась в 2005 году, а первым использованным опсином
стал канальный родопсин 2 (англ. channelrhodopsin-2, ChR2.)
Преимущество оптогенетических методов перед
традиционными электрофизиологическими методами изучения нервных
сетей и воздействия на них состоит в возможности высокоселективной
активации либо подавления конкретных нейрональных связей. Эта
селективность открывает новые возможности в терапии болезни
Паркинсона, депрессии, тревожности и эпилепсии.
10. Термины физиологии
11.
• Термины физиологии• Функция – специфическая деятельность систем
или органа.
• Процесс – последовательная смена явлений или
состояний в развитии какого-либо действия.
• Или совокупность последовательных действий,
направленных на достижение определенного
результата.
• Система в физиологии – совокупность
органов или тканей, связанных общей
функцией.
• Норма – это пределы оптимального
функционирования живой системы.
12. Биоэлектрическая активность возбудимых тканей
13. Физиологические свойства тканей как основа их деятельного состояния ( понятие, критерии оценки, последствия изменений).
14. Возбудимые ткани
Возбудимые ткани способны реагировать на раздражение генерациейпотенциала действия.
К возбудимым тканям относятся:
- нервная:
1)воспринимает раздражители путём генерации нервных
импульсов; 2) передаёт эти импульсы к эффектору, осуществляя
связь организма с окружающей средой;
3) обеспечивает взаимодействие тканей, органов и систем организма и их
регуляцию.
-мышечная: 1) обеспечивает перемещение организма в пространстве , 2)
движение органов внутри организма (сердце, язык, кишечник и др.)
-железистая:
1)
обеспечивает
регуляцию
деятельности
внутренних органов посредством гормонов, выделяемых эндокринными
клетками непосредственно в кровь либо диффундирующих через
межклеточное пространство в соседние клетки.
15. Свойства возбудимых тканей
ОбщиеВозбудимость
Проводимость
Специфические
Для мышечной
Сократимость
Для сердечной, гладкой
мышц, некоторых нейронов
Автоматия
16. ХАРАКТЕРИСТИКА СВОЙСТВ ВОЗБУДИМЫХ ТКАНЕЙ
1.Возбудимость - способность отвечать на раздражениегенерацией потенциала действия (ПД).
• Критерии возбудимости
• Порог
раздражения.
Минимальная
сила
раздражителя, cпособная вызвать возбуждение
(зависимость обратная).
• Для электрических раздражителей:
• Реобаза. Минимальная сила тока или напряжения,
способная вызвать возбуждение (определение дано
Лапиком).
• Полезное время. Наименьшее время, в течение
которого должен действовать раздражающий стимул
для возникновения возбуждения (характеризует
скорость возникновения возбуждения).
17.
• Хронаксия . Наименьшее время, в течениекоторого электрический ток, равный удвоенной
реобазе, должен действовать на ткань, чтобы
вызвать возбуждение (характеризует скорость
возникновения возбуждения).
• При повышении возбудимости может
появиться ответ на ранее подпороговые
раздражители.
• При снижении возбудимости на прежние по
силе раздражители ответа не будет. Нужно
увеличивать силу раздражения.
18.
2.ПРОВОДИМОСТЬ– способность ткани проводить возбуждение от
места возникновения к другим участкам или
структурам.
• Критерии проводимости
- скорость распространения возбуждения:
В нервных волокнах - 1 – 120 м/с
В мышечных волокнах -0,5-4 м/с
• При изменении проводимости
нарушается распространение возбуждения
в нейронных сетях, рефлекторных дугах,
от одной части органа к другой.
19.
• 3. СОКРАТИМОСТЬ способность ткани менятьгеометрию.
Критерии сократимости - изменение длины или тонуса
мышцы. При сокращении мышц можно измерить развиваемое
усилие (динамометрия, кг).
При изменении сократимости возникает нарушение позы,
движений, моторной активности гладкомышечных внутренних
органов.
4.АВТОМАТИЯ
• Способность ткани самопроизвольно возбуждаться без
внешних раздражителей.
• Критерии автоматии – частота спонтанных возбуждений.
Изменение автоматии - Нарушение работы сердца,
моторной активности гладкомышечных внутренних органов
20. Биоэлектрические явления в возбудимых тканях
• Развиваются на мембране клеток.• Представлены потенциалом покоя (ПП) и
потенциалом действия (ПД).
• Теория, объясняющая существование
биоэлектрических явлений в организме
называется мембранно-ионной.
• Авторы этой теории: В. Ю. Чаговец – 1896 г.,
Бернштейн 1902-1903 гг., Ходжкин, Хаксли, Кац.
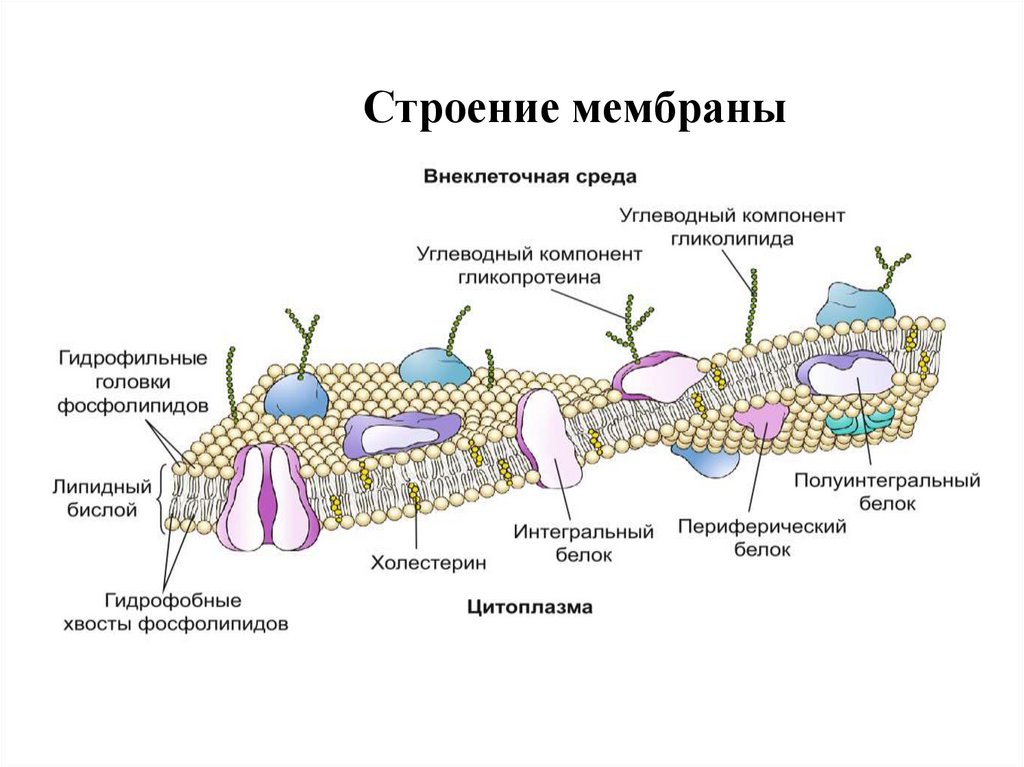
21. Строение мембраны
гидрофильныеучастки молекул
фосфолипидов
гидрофобные
концы молекул
находятся внутри
бислоя
22.
Строение мембраны23. Строение мембраны (жидкостно-мозаичная модель)
Строение мембраны(жидкостномозаичная модель)
Непрерывный
бислой липидов .
Гидрофобные концы
молекул находятся
внутри бислоя
гидрофильные
участки молекул
фосфолипидов
Изнутри мембрана
выстлана слоем
гиалоплазмы.
Толщина – 6-10 нм. Липидов около 40%,
белков около 60%.
На внешней стороне углеводы в виде
гликопротеидов или гликолипидов и
образуют гликокаликс. Углеводы участвуют
в рецепции БАВ, регуляции иммунитета.
24. Белки мембраны
• Интегральные. Это большинство белков, прочносвязаны с фосфолипидами. Встроены в
фосфолипидный матрикс клеточной мембраны.
• Периферические белки, прикрепленные к
поверхности мембраны.
• По функции делятся на:
• Белки-насосы – для активного транспорта, белки-каналы –
для пассивного транспорта веществ.
• Белки-рецепторы взаимодействуют с химическими
• веществами, которые несут к клетке информацию.
• Белки-ферменты запускают цепи биохимических
• реакций в клетке.
25. Роль мембраны
• 1.Регуляторная, рецепторная функция• за счет рецепции внеклеточных биологически активных
веществ осуществляют регуляцию внутриклеточного
содержимого и внутриклеточных реакций. Это приводит к
изменению активности ферментных систем мембраны и
запуску
механизмов
вторичных
«месенджеров»
(«посредников»).
• 2.Барьерная функция связана с наличием
бислоя липидов, обеспечивающего избирательную
проницаемость для жирорастворимых молекул и
почти полную непроницаемость для ионов.
26.
• 3. Транспортная функция мембраны• связана с работой селективных белковых каналов, по
которым ионы перемещаются через мембрану (К+, Na+, Ca++,
CL-.)
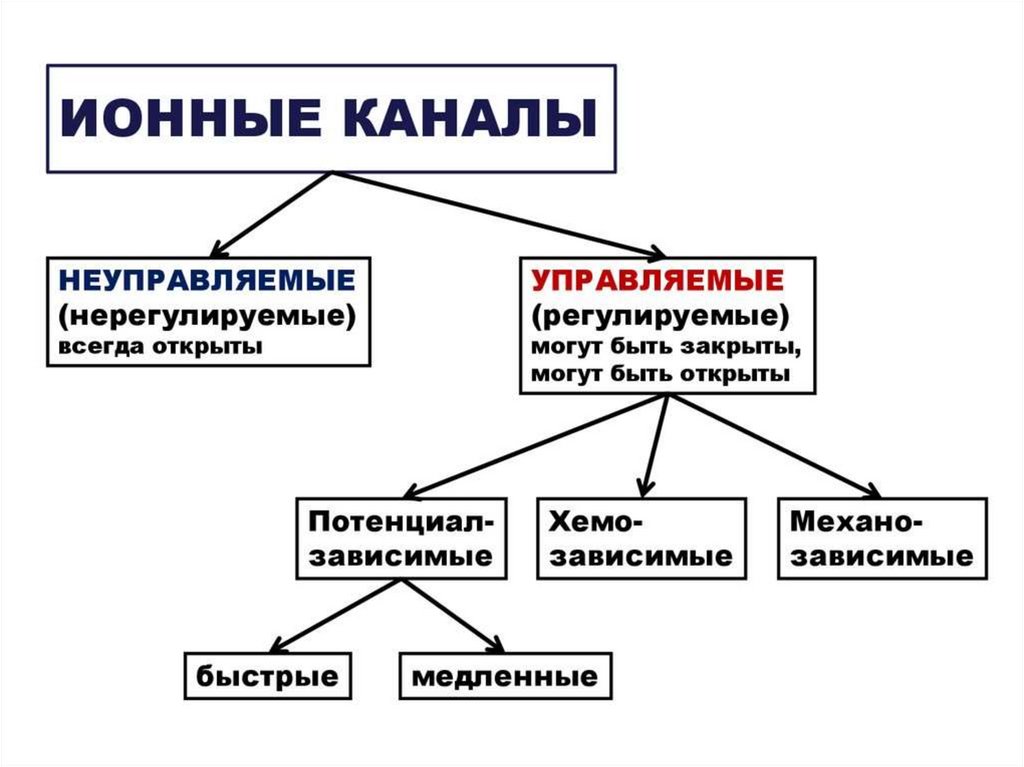
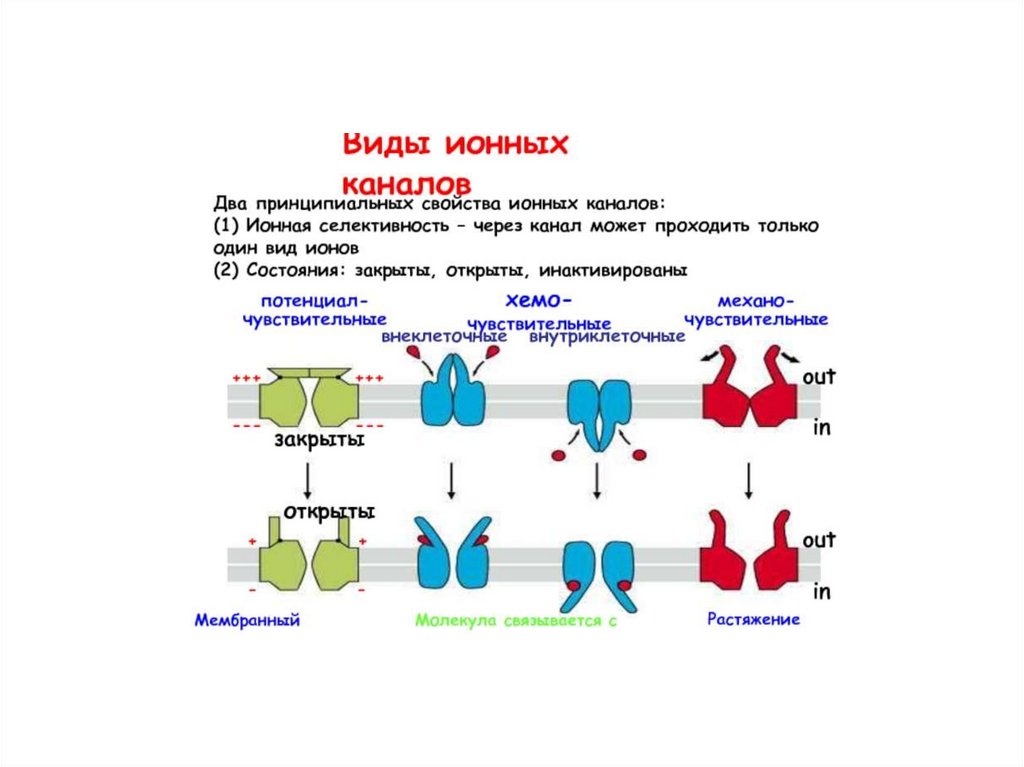
• Виды ионных каналов.
• 1.По состоянию
• - открытые в покое (не управляемые) – К+, Na+,
• - закрытые в покое (управляемые) – Na+, Са++, К+, CL. 2.Управляемые каналы по способу открытия различают:
а) хемозависимые открываются при воздействии на
хеморецепторы мембраны клетки физиологически активных
веществ (нейромедиаторов, гормонов и т. д).
б) потенциалзависимые открываются при определенном
уровне потенциала мембраны.
27.
•3.По скорости инактивации, т.е. закрывания:a) быстроинактивирующиеся, т.е. быстро переходящие в
закрытое состояние. Они обеспечивают быстро нарастающее
снижение МП и такое же быстрое восстановление.
б) медленноинактирующиеся. Их открывание вызывает
медленное снижение МП и медленное его восстановление
4.По избирательности:
a) Селективные, т.е. специфические. Эти каналы проницаемы
для строго определенных ионов (К+, Na+, Ca++, CL-.)
б) Малоселективные, неспецифические, не имеющие
определенной ионной избирательности. Их в мембране
небольшое количество.
28.
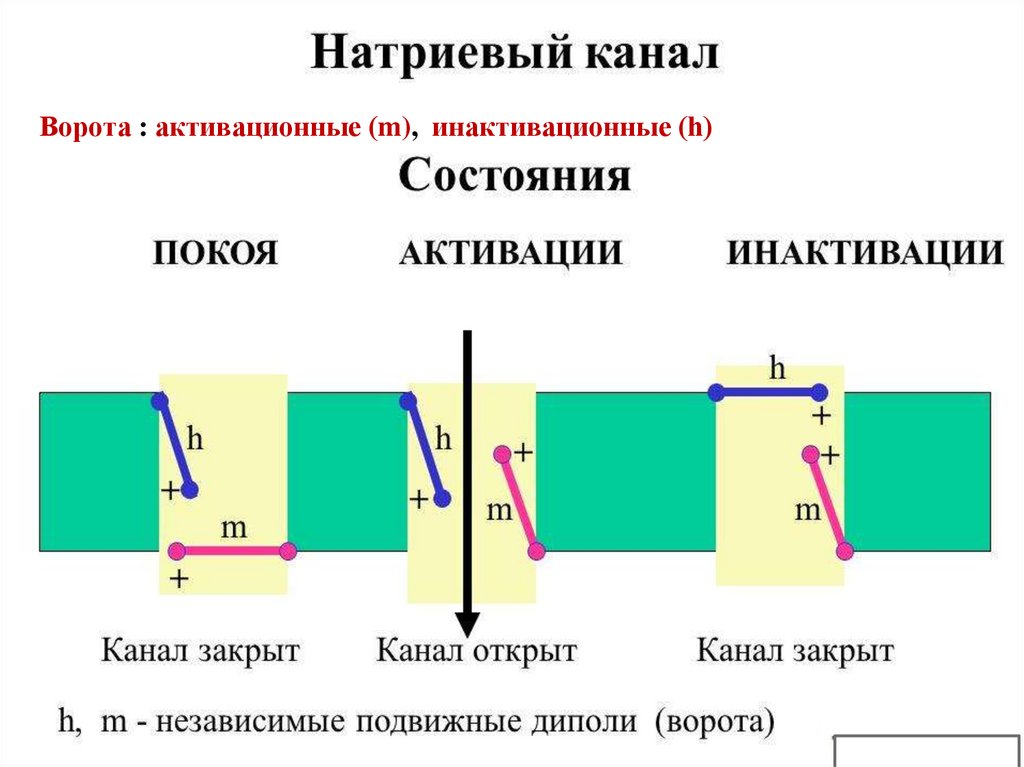
29. Строение селективных ионных каналов.
• 1.Селективный фильтр, расположенный в устье каналаобеспечивает прохождение через канал строго
определенных ионов.
• 2.Активационные ворота. У потенциалзависимых
каналов имеется сенсор, который открывает их на
определенном уровне МП. У хемочувствительных- при
действии соответствующего БАВ.
• 3. Инактивационные ворота. Обеспечивают
закрывание канала и прекращение проведения ионов
по каналу на определенном уровне МП
• Неспецифические ионные каналы не имеют ворот.
30. Селективные ионные каналы могут находиться в трех состояниях, которые определяются положением активационных (m) и
инактивационных (h) ворот:• 1.Закрытом, когда активационные закрыты, а
инактивационные открыты.
• 2.Активированном, и те и другие ворота
открыты.
• 3.Инактивированном, активационные ворота
открыты, а инактивационные закрыты
• Открытие каналов приводит к пассивному
транспорту ионов по градиенту концентрации и,
как следствие, изменению заряда мембраны.
31.
Ворота : активационные (m), инактивационные (h)32.
33.
34. Роль ионов в биоэлектрических процессах
• Биоэлектрические явления ввозбудимых тканях связаны с
перемещением ионов через
мембрану.
35. Потенциал покоя
• В покое плазматическая мембрана возбудимыхклеток поляризована: снаружи она заряжена «+»,
внутри – «-». Эта поляризация называется
потенциалом покоя (ПП).
• ПП – трансмембранная разница заряда
плазматической мембраны.
• ПП = - 30-90 мв (в разных клетках).
36.
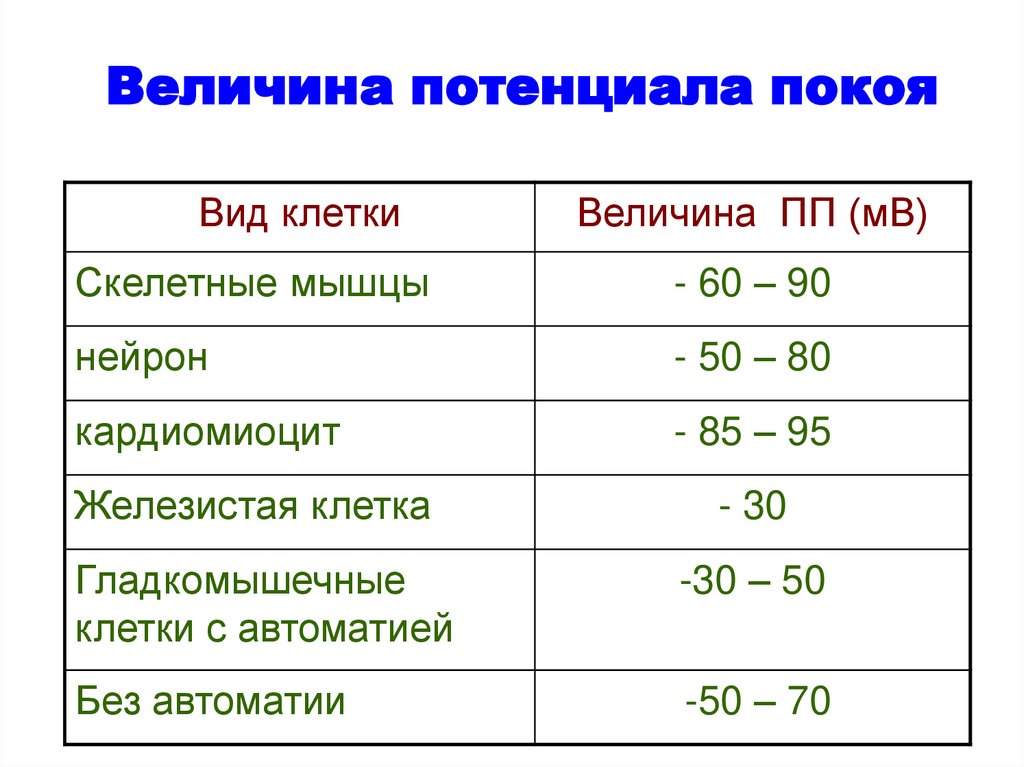
Величина потенциала покояВид клетки
Величина ПП (мВ)
Скелетные мышцы
- 60 – 90
нейрон
- 50 – 80
кардиомиоцит
- 85 – 95
Железистая клетка
- 30
Гладкомышечные
клетки с автоматией
-30 – 50
Без автоматии
-50 – 70
37. Происхождение потенциала покоя (мембранно-ионная теория)
ПП в основном создается выходящими ионами К+ .
В покое в мембране открыты каналы для К+ и закрыты
практически все каналы для Na+.
Существуют ионные градиенты:
Внутри клетки ионов К+ в 30 – 50 раз больше, чем
снаружи. Na+ внутри находится в основном в связанном
состоянии или находится в компартментах, т.е.
иммобилизован. Концентрация Na+ к клетке в 10 раз
меньше, чем вне ее.
38.
• Ионы К+ пассивно, по градиенту концентраций, черезоткрытые каналы выходят из клетки и создают
положительный заряд наружной стороны мембраны
клетки.
• Внутри остаются крупные анионы (CL-, белки).
• которые не могут пройти через каналы для иона К+ и
создают отрицательный заряд внутренней стороны
мембраны.
• Калиевый поток создает потенциал:Ек+ = -90 мв (знак «-»
заряд внутренней стороны мембраны),
• Но мембрана несколько проницаема для Na+.
• Натриевый поток в клетку создает потенциал, равный
+10 мв (ЕNa+ = +10 мв) и снижает внутреннюю
отрицательность, создаваемую выходящим К
• Таким образом: ЕПП = ЕК - ЕNa
• ЕПП = -90 - (+10 )= -80 мв
39.
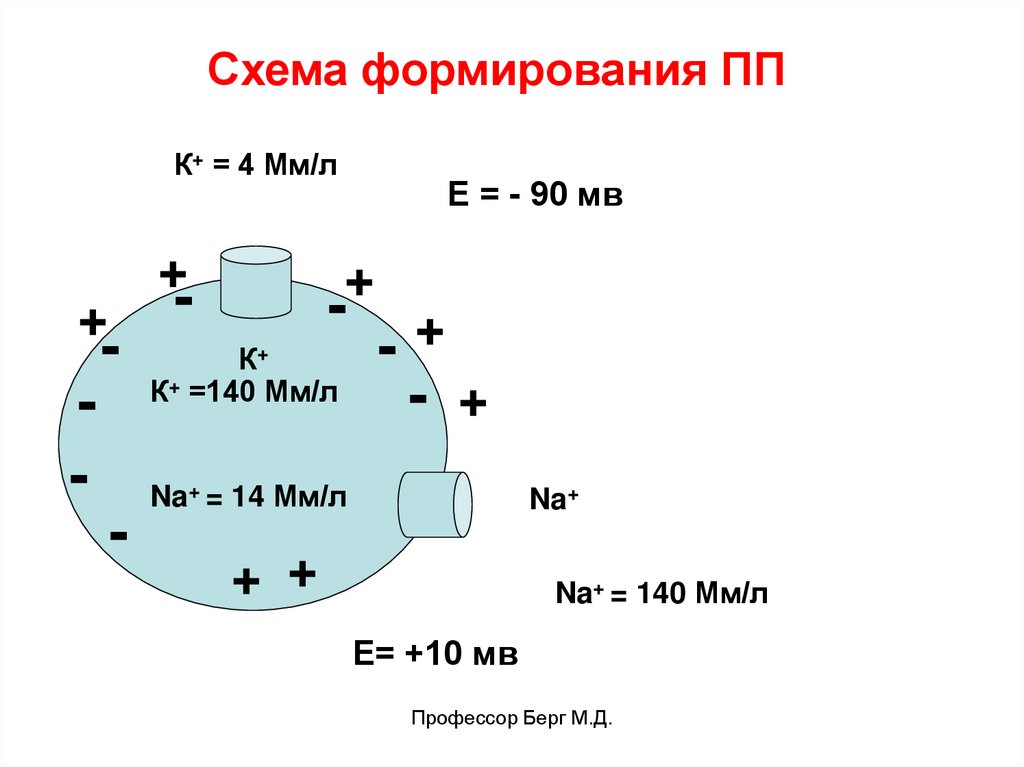
Схема формирования ППК+ = 4 Мм/л
+
+
-
-
Е = - 90 мв
+
-
К+
К+ =140 Мм/л
+
-
- +
Na+ = 14 Мм/л
Na+
+ +
Na+ = 140 Мм/л
Е= +10 мв
Профессор Берг М.Д.
40. K – Na насос
• Несмотря на непрерывное перемещениеионов через мембрану, сохраняется
разная концентрация Na+ К+ снаружи и
внутри клетки. Поддержание ионных
градиентов осуществляет натрий калиевый насос.
• Ионный насос удаляет из клетки Na+ и
возвращает вышедший К+.
• Переносчиком для Na и К является мембранная
Nа – К зависимая АТФ – аза ( фермент,
расщепляющий АТФ).
41.
• Переносчик с внутренней стороны мембраныклетки присоединяет 3 иона натрия, а с
наружной 2 иона калия.
• При этом начинается расщепление АТФ.
Выделенная энергия используется для переноса
ионов из меньшей концентрации в большую ( Na
из клетки, K в клетку
• т.е. против градиента концентраций (активный
транспорт ионов).
• Удаление из клетки больше катионов, чем в нее
возвращается поддерживает отрицательность
внутренней стороны мембраны (электрогенный
насос).
42.
Схема работы натрий-калиевого насоса2К+
3Na+
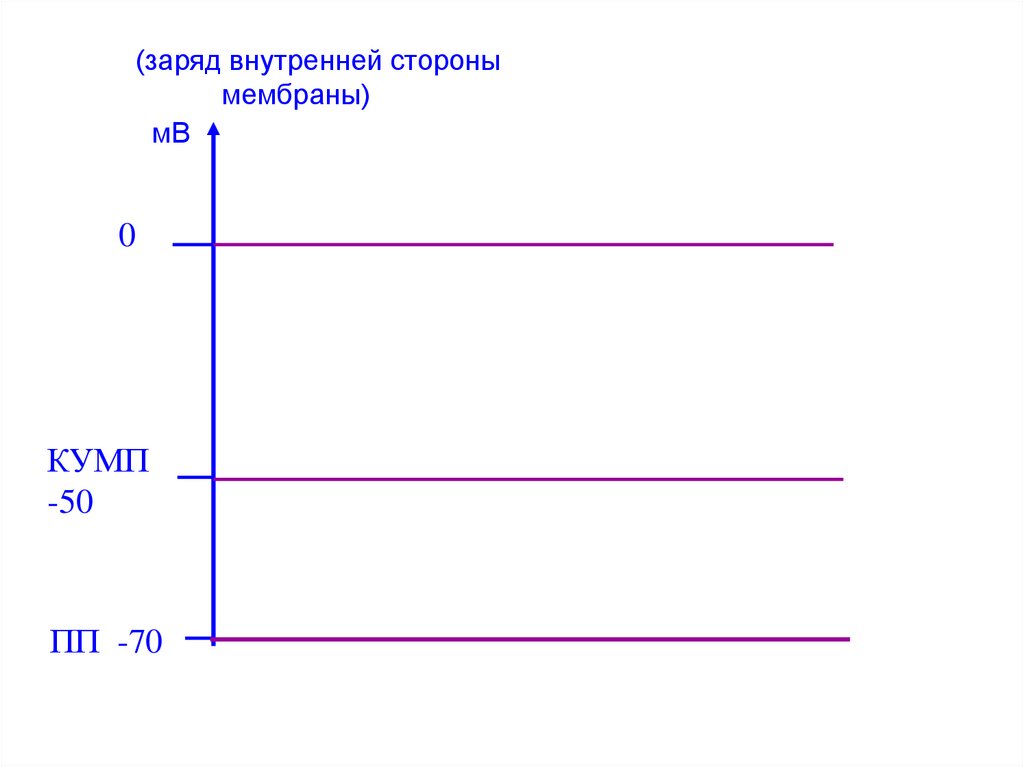
43. Критический уровень мембранного потенциала (КУМП)
• – значение ПП, при достижениикоторого открываются
потенциалзависимые каналы для
натрия и возникает ПД.
Величина КУМП меньше ПП на
20-25 мВ.
44.
(заряд внутренней сторонымембраны)
мВ
0
КУМП
-50
ПП -70
45. Изменения потенциала покоя
Снижение внутреннейотрицательности деполяризация;
Повышение гиперполяризация.
Величина порога раздражения
и возбудимость зависит от
разности (ΔЕ) между
величинами мембранного
потенциала (Ео) и
критического уровня
деполяризации ( Ек).
При уменьшении ΔЕ
возбудимость повышается,
•при увеличении возбудимость
снижается.
МВ
0
Ек
ΔЕ
Ео
КУМП
деполяризация
ПП
гиперполяризация
46.
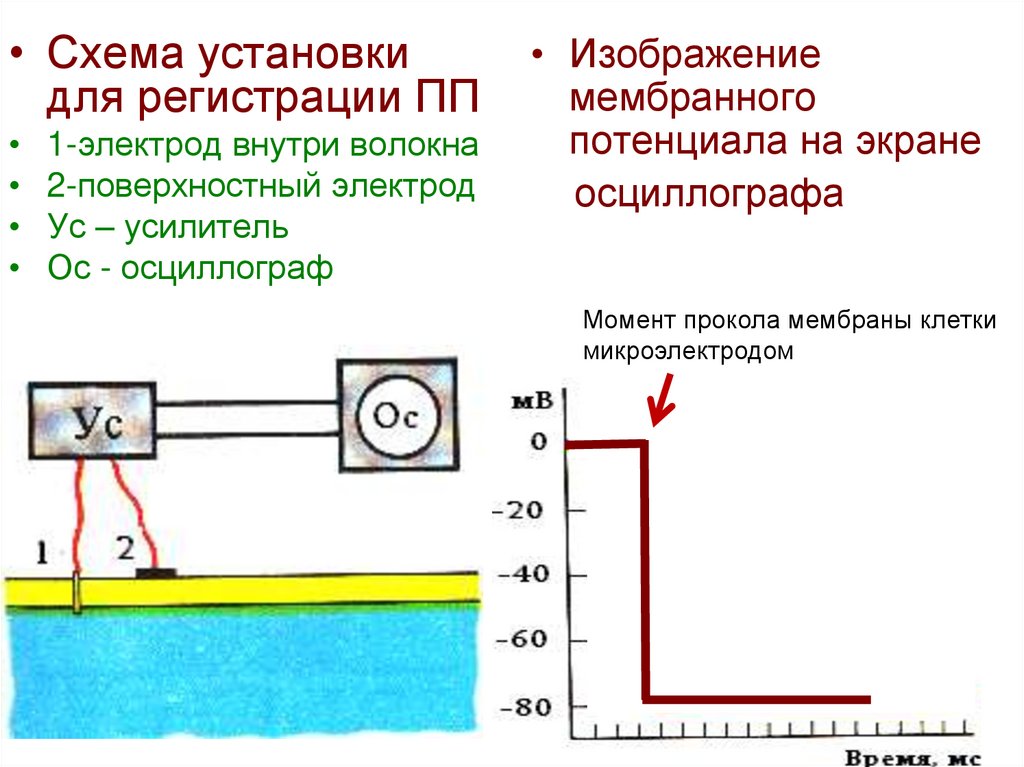
• Схема установкидля регистрации ПП
1-электрод внутри волокна
2-поверхностный электрод
Ус – усилитель
Ос - осциллограф
• Изображение
мембранного
потенциала на экране
осциллографа
Момент прокола мембраны клетки
микроэлектродом
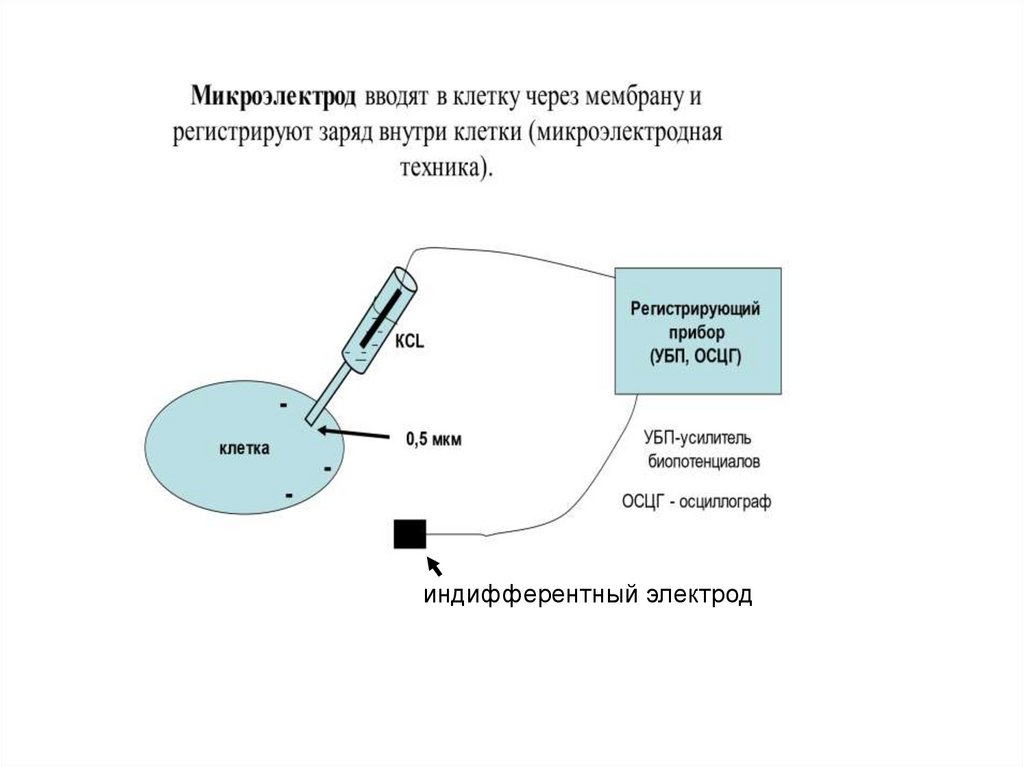
47.
индифферентный электрод48. Виды ответов возбудимой ткани на различные по силе раздражения
1.Локальный ответ2. Потенциал действия
49. Локальный ответ (ЛО)
• Возникает в локальном участке при действиихимических или электрических стимулов силой 30
– 90% от пороговой силы раздражения
(подпороговый раздражитель).
• Под влиянием подпорогового раздражения
открываются хемочувствительные каналы для Na
(иногда Са). Возникает деполяризация мембраны, но
не достигнув КУМП быстро сменяется
реполяризацией.
• При высокой частоте подпороговых
раздражений возможна суммация ЛО
50.
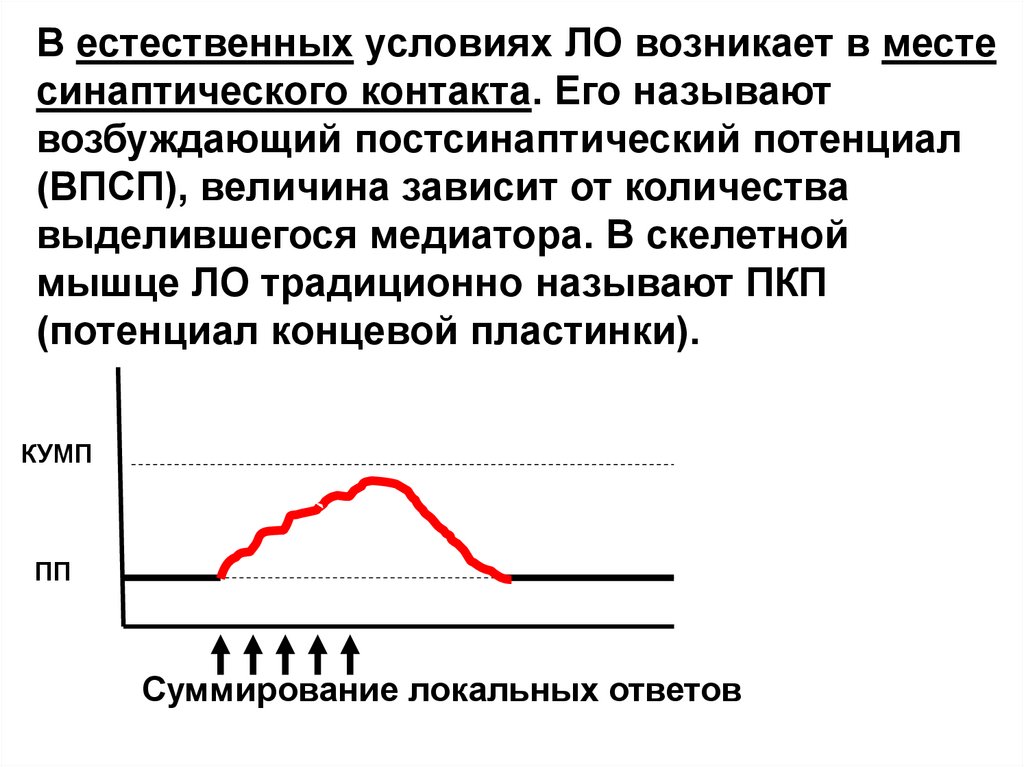
В естественных условиях ЛО возникает в местесинаптического контакта. Его называют
возбуждающий постсинаптический потенциал
(ВПСП), величина зависит от количества
выделившегося медиатора. В скелетной
мышце ЛО традиционно называют ПКП
(потенциал концевой пластинки).
КУМП
ПП
Суммирование локальных ответов
51. Свойства локального ответа (ЛО)
Не распространяетсяЗависит от силы раздражения
Способен к суммации
Является предфазой ПД
52.
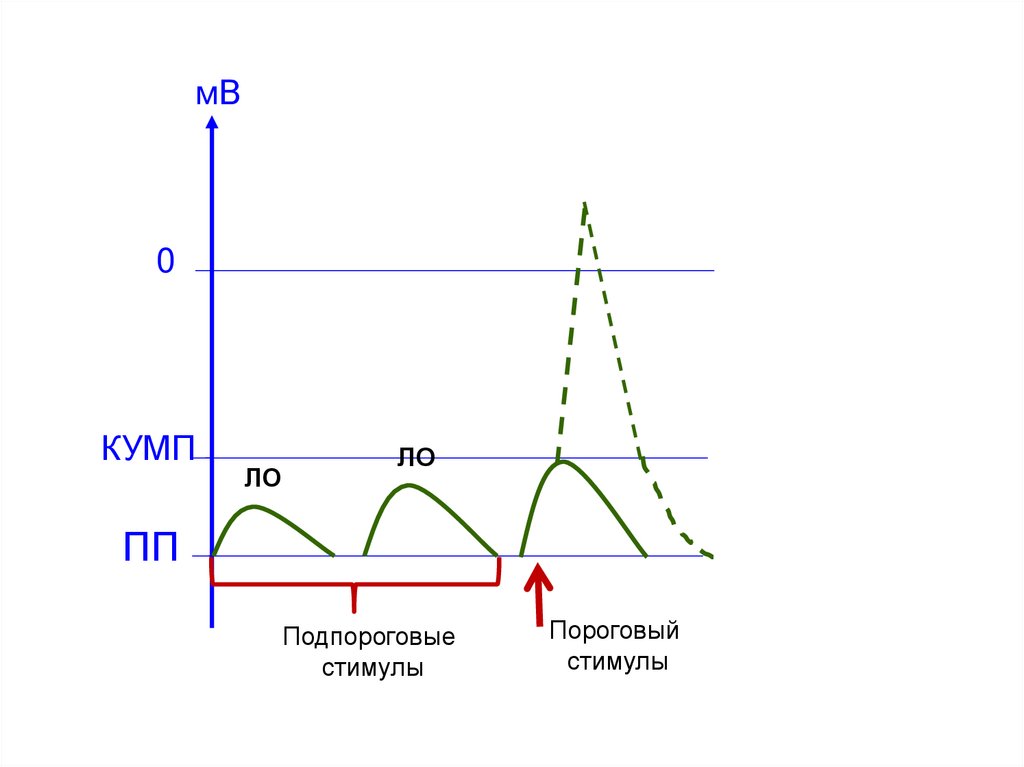
мВ0
КУМП
ЛО
ЛО
ПП
Подпороговые
стимулы
Пороговый
стимулы
53.
• Потенциал действия - это быстрыеколебания ПП под влиянием порогового или
сверхпорогового раздражения.
• Условия возникновения ПД :
• пороговая сила раздражения
• достаточная длительность раздражения
• достаточная скорость нарастания
раздражения.
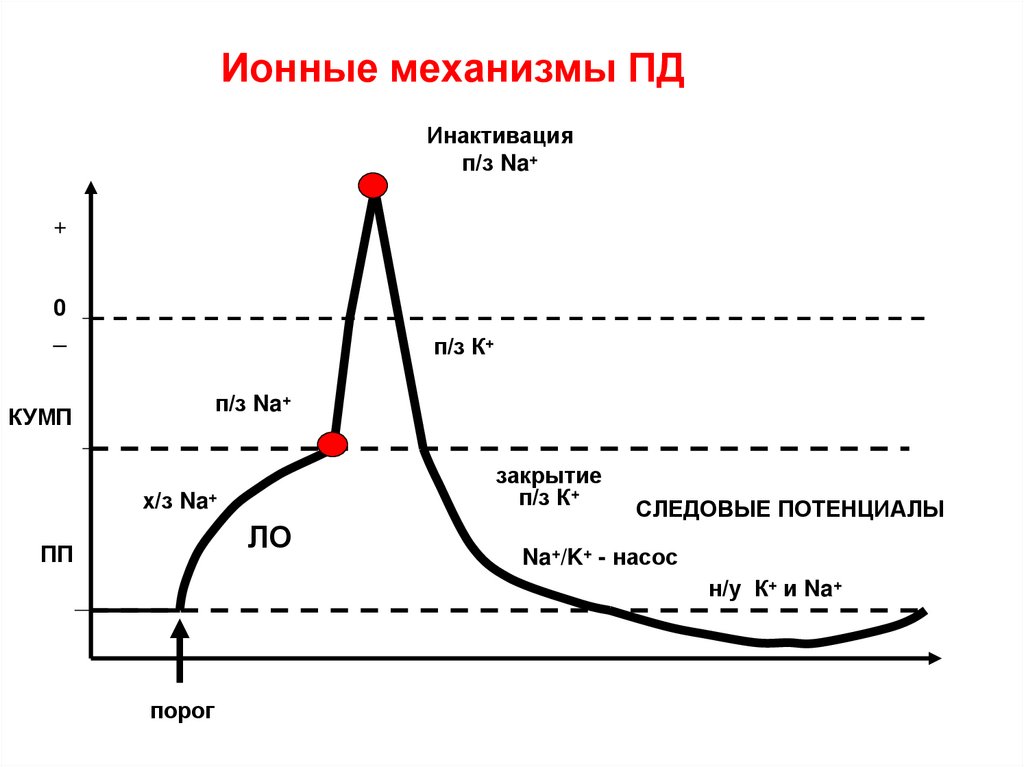
54. Ионный механизм ПД
• Под влиянием порогового раздражения в мембранеклетки открываются хемочувствительные каналы
для Na+ . Медленный ток Na + внутрь клетки
снижает ПП до КУМП.
• С этого момента в мембране открываются быстрые
потенциалзависимые каналы для Na+ и Na+
лавиной входит внутрь клетки. Внутренняя
отрицательность снижается до 0, а затем возникает
перезарядка мембраны (внутри +, снаружи-).
Происходит деполяризация мембраны
• При достижении величины перезарядки +10 +40 мВ
( в зависимости от вида клеток) каналы для натрия
закрываются (происходит натриевая инактивация)
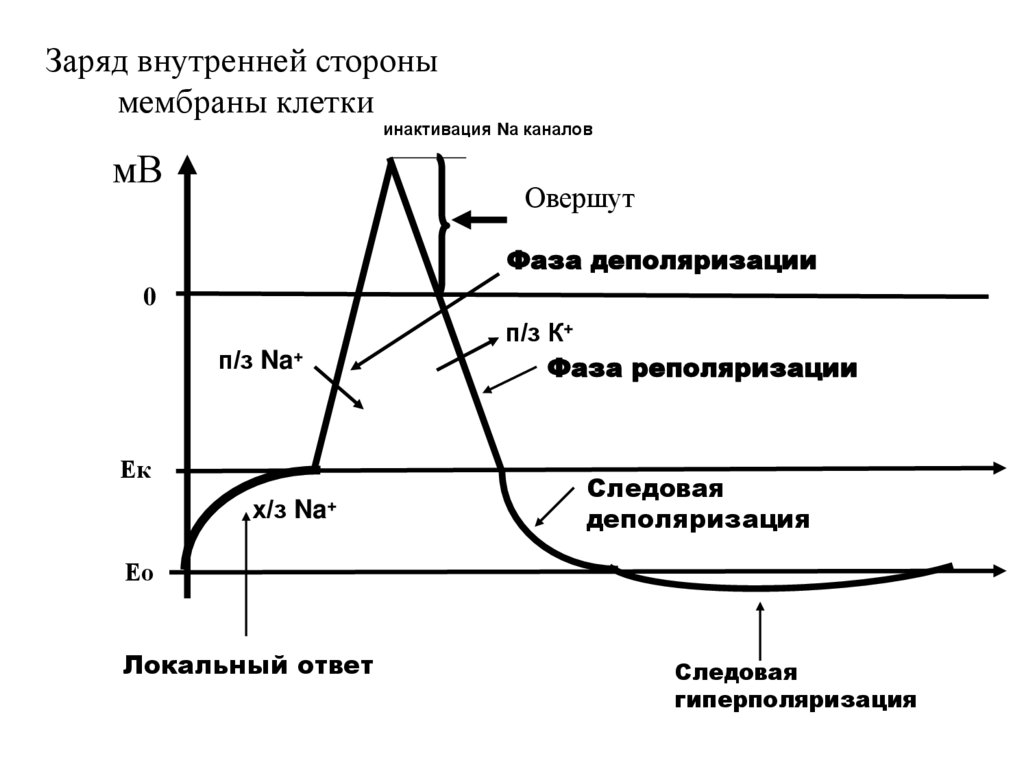
• и начинается фаза реполяризации.
55.
• Реполяризация идет сначала быстро,потом медленно и могут наблюдаться
следовые потенциалы.
• Следовая деполяризация отражает
инерционность закрытия натриевых
каналов.
• Следовая гиперполяризация связана с
работой в мембране электрогенного
ионного насоса.
56. Элементы потенциала действия
57.
Заряд внутренней сторонымембраны клетки
инактивация Nа каналов
мВ
Овершут
Фаза деполяризации
0
п/з Na+
Ек
х/з Na+
п/з К+
Фаза реполяризации
Следовая
деполяризация
Ео
Локальный ответ
Следовая
гиперполяризация
58.
Ионные механизмы ПДИнактивация
п/з Na+
+
0
_
п/з К+
п/з Na+
КУМП
закрытие
п/з К+
х/з Na+
ЛО
ПП
СЛЕДОВЫЕ ПОТЕНЦИАЛЫ
Na+/K+ - насос
н/у К+ и Na+
порог
59.
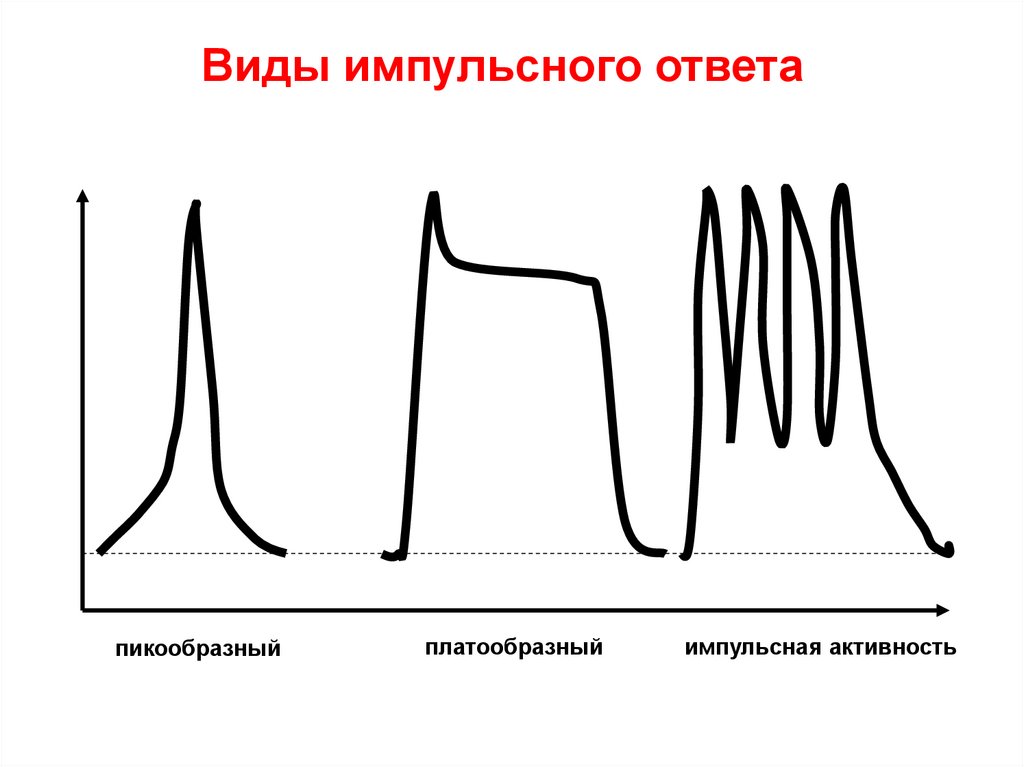
Виды импульсного ответапикообразный
платообразный
импульсная активность
60. Формы ПД
• ПикообразныйЗаряд внутренней стороны
мембраны клетки
мВ
Овершут
Фаза деполяризации
0
Na
K
Фаза реполяризации
Следовая
деполяризация
Ек
Ео
Локальный ответ
Следовая
гиперполяризация
61. Платообразный ПД кардиомиоцита желудочков
мВ0
-60 Ек
-90 Ео
Са2+
Na+
К+
62. Свойства потенциала действия
РаспространяетсяПроводится без
затухания
Не суммируется
в одиночной
структуре
Подчиняется
Закону «Все или
ничего»
63. Изменение возбудимости при возбуждении
0Ек
Ео
возбудимость
Исходный
уровень
0
1
2
3
4
5
Изменение
возбудимости
во времени
64.
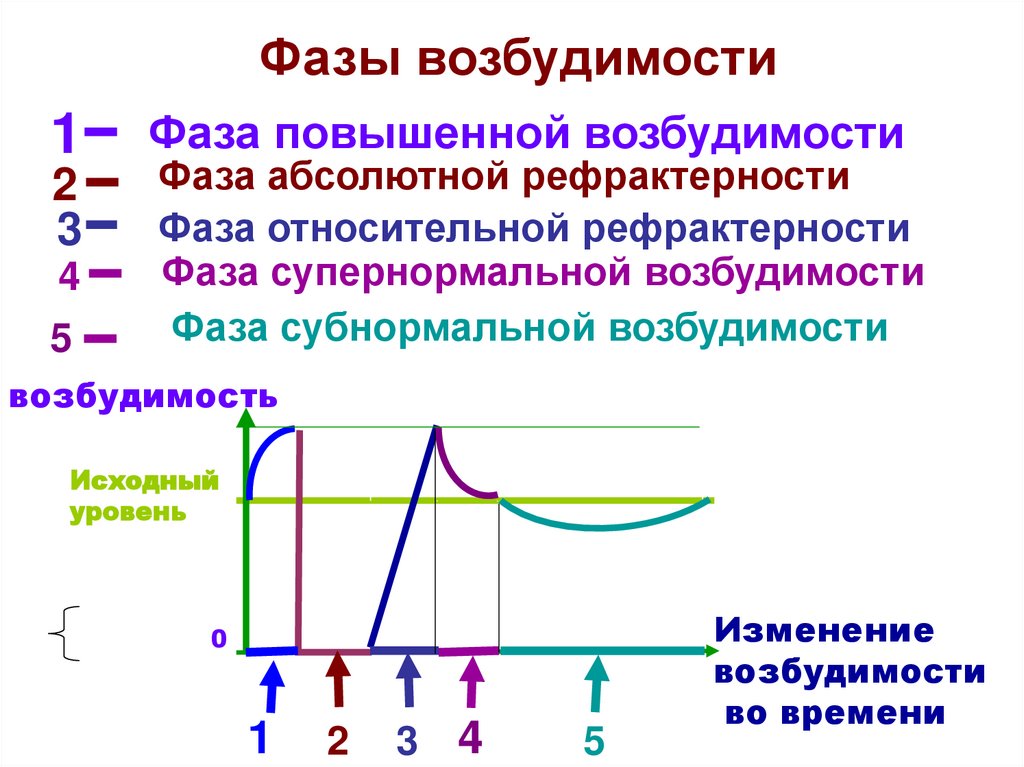
Фазы возбудимости1
2
3
4
5
Фаза повышенной возбудимости
Фаза абсолютной рефрактерности
Фаза относительной рефрактерности
Фаза супернормальной возбудимости
Фаза субнормальной возбудимости
возбудимость
Исходный
уровень
0
1
2
3
4
5
Изменение
возбудимости
во времени
65.
Кодированиеинформации
в организме
66.
• Информация — любая совокупностьсигналов, сведений (данных),
которые какая-либо система
воспринимает из окружающей среды
(входная информация),
• выдает в окружающую среду (
исходящая информация )
• или сохраняется внутри
определенной системы (внутренняя
информация).
67. Кодирование информации в организме - это
перевод информации в систему условныхсигналов
Уровни кодирования:
рецептор
В виде рецепторного потенциала - РП
проводящие
пути
В виде потенциалов действия ПД
отделы ЦНС
кора
Перекодирование информации
Декодирование информации
68. Этапы кодирования информации:
Информацияклетка
Изменение
потенциала
покоя
Пикообразный
Импульсный
ответ
Локальный
ответ
Платообразный