




















































































































 biology
biologySimilar presentations:


 размножение")







Цитология растений. Вегетативные органы растений
1.
Ботаника. 1 семестр2.
ОглавлениеI.
Цитология растений
II.
Ткани растений
III. Вегетативные органы растений
IV. Генеративные органы растений
3.
I. Цитология растенийОбщая характеристика растительной клетки
Цитоплазма
Пластиды
Строение и функции ядра
Вакуоль
Клеточная оболочка
Цитоплазматические включения
4.
Общая характеристика растительнойклетки
Клетка представляет собой основную структурную и функциональную
единицу всех живых существ и обладает всеми признаками живого:
ростом, обменом веществ и энергией с окружающей средой,
делением, раздражимостью, наследственностью и др.
Среди растений есть виды, представленные одной клеткой (некоторые
виды водорослей), однако большинство является многоклеточными
организмами. Строение клеток разнообразно и зависит от
выполняемых ими функций.
По степени сложности внутренней организации клетки можно
разделить на 2 типа: прокариотические и эукариотические.
5.
6.
ЦитоплазмаЦитоплазма — основная часть протопласта клетки, в которой проходят все
процессы клеточного обмена веществ, кроме синтеза нуклеиновых кислот,
происходящего в ядре.
Основу цитоплазмы, ее бесструктурный матрикс называют гиалоплазмой.
Гиалоплазма — бесцветная коллоидная система, обладающая
ферментативной активностью и обеспечивающая взаимодействие всех
органелл цитоплазмы. Гиалоплазму пронизывают микротрубочки и
микрофиламенты, совокупность которых составляет цитоскелет растительной
клетки. Цитоскелет влияет на перемещение внутриклеточных структур и
определяет форму растущей клетки.
Микротрубочки — надмолекулярные агрегаты длиной в несколько микронов с
упорядоченным расположением молекул белка тубулина. Способны к
самосборке и самораспаду. Принимают участие во внутриклеточном
транспорте веществ, а также в формировании жгутиков, ресничек,
ахроматинового веретена деления.
Микрофиламенты — способные сокращаться нити белка актина. Образование
и распад микротрубочек и микрофиламентов вызывают обратимые переходы
участков цитоплазмы из золя в гель.
7.
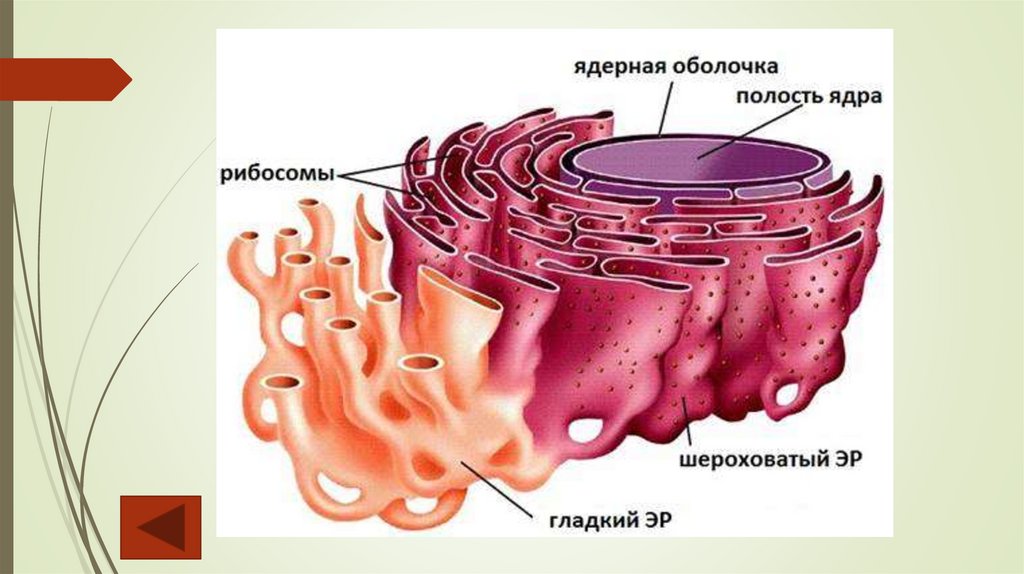
РибосомыРибосомы — универсальные органеллы, содержащиеся во всех
клетках. Функция рибосом — биосинтез белка. Каждая рибосома
состоит из двух субъединиц — большой и малой. В состав рибосомы
ядерных организмов входят четыре молекулы рибосомальной РНК
(рРНК) и белки (около 100 видов). Образование субъединиц рибосом
происходит в ядре. Покидая ядро, они поступают в цитоплазму, где на
молекуле информационной РНК (иРНК) происходит их сборка в
рибосому.
8.
9.
Эндоплазматический ретикулумЭндоплазматический ретикулум (ЭР), или эндоплазматическая сеть, —
непрерывно изменяющаяся трехмерная система субмикроскопических
цистерн, канальцев и пузырьков, отделенных от гиалоплазмы элементарной
мембраной и заполненных бесструктурной энхилемой.
Канальцы ЭР непосредственно переходят в наружную мембрану ядерной
оболочки, благодаря чему осуществляется связь ядра с цитоплазмой. Канальцы
ЭР, переходящие из одной клетки в другую и обеспечивающие связь между
ними, называют плазмодесмами.
Длинные канальцы с гладкой поверхностью (агранулярный, или гладки, ЭР)
принимают участие в синтезе углеводов, жиров, эфирных масел, смол,
каучука, стероидных гормонов, а также в накоплении и выведении ядовитых
веществ. Цистерны, короткие канальцы и пузырьки, на поверхности которых
располагаются рибосомы, представляют собой гранулярный, или
шероховатый, ЭР. Его главная функция — транспорт и накопление белков,
синтезированных рибосомами. Агранулярный ЭР в клетках обычно развит
слабо, но в клетках выделительных тканей он выражен очень хорошо.
Таким образом, эндоплазматическая сеть обеспечивает транспорт веществ
как внутри клетки, так и между клетками.
10.
11.
Аппарат ГольджиАппарат Гольджи состоит из отдельных диктиосом (телец Гольджи) и
пузырьков Гольджи. Диктиосомы представляют собой стопки плоских
круглых цистерн (пять — семь, иногда до 20), отделенных от гиалоплазмы
одной мембраной и заполненных матриксом. По краям цистерны
переходят в состоящую из трубочек сеть, от которой отчленяются пузырьки
Гольджи.
Диктиосомы имеют два полюса. С одной их стороны (образующей)
происходит постоянное образование новых цистерн из канальцев ЭР; с
другой (секретирующей) — старые цистерны распадаются на пузырьки
Гольджи, которые направляются к плазмалемме или тонопласту — двум
пограничным мембранам клетки. В цистернах диктиосом завершаются
многие обменные реакции, проходящие в клетке. В них, в частности,
накапливаются, конденсируются и упаковываются для транспортировки
вещества, которые необходимо удалить из цитоплазмы.
Одна из важнейших функций аппарата Гольджи — синтез полисахаридов
(пектинов, гемицеллюлоз, слизей), идущих на построение клеточной
стенки.
12.
13.
ЛизосомыЛизосомы — одномембранные органеллы округлой формы, в
матриксе которых содержатся гидролитические ферменты —
гидролазы, способные расщеплять органические вещества, в том
числе и биополимеры.
В отличие от грибов и животных в клетках растений лизосомы
встречаются реже. В растительной клетке гидролитические ферменты
могут находиться в разных ее органеллах и зонах, образуя так
называемый лизосомный клеточный компартмент.
Основная функция лизосом — внутриклеточное переваривание,
автолиз: разрушение отдельных участков цитоплазмы собственной
клетки, заканчивающееся образованием на их месте
цитоплазматической вакуоли. Лизосомы очищают клетку от уже не
работающих органелл.
14.
15.
ПероксисомыПероксисомы (микротельца) встречаются в большинстве типов клеток
растений и грибов. Они представляют собой мелкие (0,2—1,5 мкм)
одномембранные органеллы сферической формы. Их плотный
матрикс состоит в основном из окислительно-восстановительных
ферментов.
Функции определяются типом клеток, в которых они находятся.
Например, при прорастании семян пероксисомы, находящиеся в
клетках запасающих тканей, обеспечивают превращение жирных
масел в сахара; в клетках же фотосинтезирующих тканей в них
проходят реакции светового дыхания — поглощение кислорода,
выделение двуокиси углерода, синтез аминокислот.
16.
17.
МитохондрииМитохондрии — постоянно перемещающиеся органеллы округлой,
цилиндрической или нитевидной формы. Относительно крупный размер
(длина — до 10 мкм, диаметр — 0,2-1 мкм) позволяет видеть их в световой
микроскоп. Число, расположение, форма и размеры митохондрий
постоянно меняются.
Совокупность митохондрий клетки называют хондриомом.
Внутренняя мембрана митохондрии образует выступающие в ее матрикс
трубковидные выросты — кристы, что увеличивает внутреннюю активную
поверхность органеллы. В матриксе находятся кольцевые молекулы
митохондриальной дезоксирибонуклеиновой кислоты (ДНК),
специфические иРНК и тРНК и рибосомы прокариотического типа,
отличающиеся от цитоплазматических меньшими размерами. Это
позволяет митохондриям самостоятельно синтезировать белки для своих
мембран.
Митохондрии — энергетический центр клетки. На поверхности внутренней
мембраны, в матриксе и межмембранном пространстве идут процессы
внутриклеточного дыхания — окисление органических веществ кислородом
воздуха до диоксида углерода и воды.
18.
19.
ПластидыПластиды встречаются только в растительных клетках. Выделяют три типа пластид
(хлоро–, лейко– и хромопласты), которые отличаются друг от друга составом
пигментов (цветом), строением и выполняемыми функциями.
Хлоропласты имеют зеленый цвет и встречаются во всех зеленых органах растения
(листьях, стеблях, незрелых плодах). Они содержат зеленый пигмент хлорофилл,
который находится в хлоропластах в нескольких формах. Кроме хлорофилла в них
содержатся пигменты, относящиеся к группе каротиноидов, в частности желтый
(ксантофилл) и оранжевый (каротин), но обычно они маскируются хлорофиллом.
Хлоропласты, как правило, имеют линзовидную форму и сложное строение.
Снаружи они ограничены оболочкой, состоящей из двух мембран. У хлоропластов,
особенно высших растений, значительно развиты внутренние мембранные
поверхности, имеющие форму плоских мешочков, называемых тилакоидами
(ламеллами). На их мембранах находится хлорофилл. Тилакоиды могут
располагаться одиночно, но чаще собраны в стопочки – граны. Внутренняя среда
пластид называется стромой. В строме хлоропластов всегда встречаются
пластоглобулы – включения жирных масел, в которых растворены каротиноиды, а
также рибосомы, светлые зоны с нитями ДНК, а в некоторых случаях – крахмальные
зерна, белковые кристаллы.
Основная функция хлоропластов – фотосинтез. Кроме того, в них, как и в
митохондриях, происходит процесс образования АТФ из АДФ, который называется
фотофосфорилированием. Хлоропласты способны также к синтезу и разрушению
полисахаридов (крахмала), некоторых липидов, аминокислот, собственного белка.
20.
21.
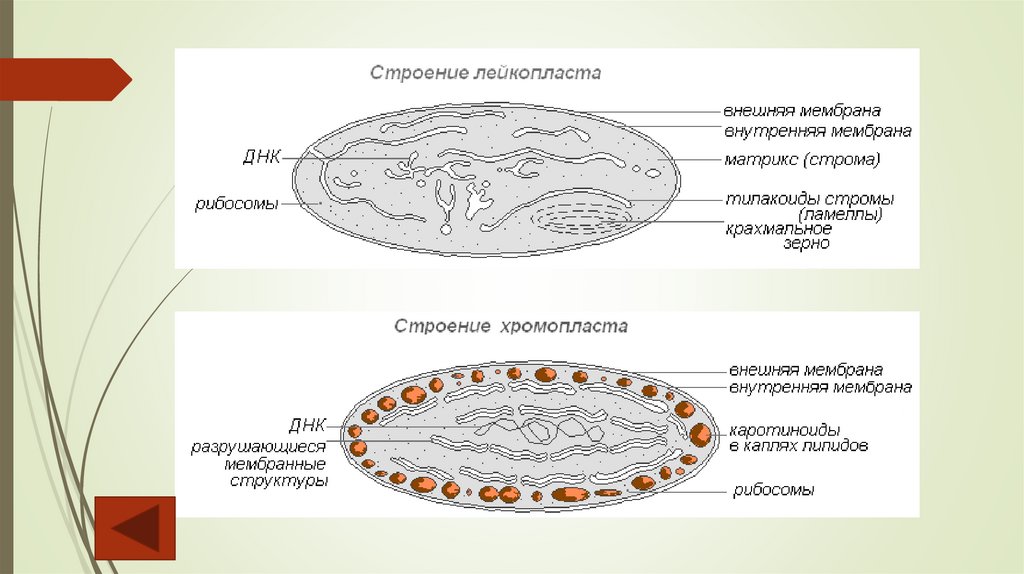
Лейкопласты – бесцветные мелкие пластиды, встречающиеся в запасающих органахрастений (клубнях, корневищах, семенах и т. д.). Для лейкопластов характерно слабое
развитие внутренней системы мембран, представленной одиночными тилакоидами,
иногда трубочками и пузырьками. Остальные компоненты лейкопластов (оболочка,
строма, рибосомы, ДНК, пластоглобулы) сходны с описанными для хлоропластов.
Основная функция лейкопластов – синтез и накопление запасных питательных веществ, в
первую очередь крахмала, иногда белков. Лейкопласты, накапливающие крахмал,
называют амилопластами, белок – протеопластами, жирные масла – олеопластами.
Пластиды, окрашенные в желтый, оранжевый, красный цвета, носят название
хромопластов. Их можно встретить в лепестках (лютик, одуванчик, тюльпан), корнеплодах
(морковь), зрелых плодах (томат, роза, рябина, хурма) и осенних листьях. Яркий цвет
хромопластов обусловлен наличием каротиноидов, растворенных в пластоглобулах.
Внутренняя система мембран в данном типе пластид, как правило, отсутствует.
Хромопласты имеют косвенное биологическое значение: яркая окраска лепестков и
плодов привлекает опылителей и распространителей плодов.
В молодых, меристематических клетках имеются пропластиды – органеллы, окруженные
двумя мембранами и способные передвигаться подобно амебам. В онтогенезе, в
зависимости от типа ткани и условий среды пропластиды могут развиваться в хлоропласты
(на свету) или лейкопласты (чаще – без света, за исключением лейкопластов в
эпидермисе).
22.
23.
Схема взаимопревращениепластид
24.
Строение и функции ядраЯдро — центральная органелла клетки, регулирующая все процессы
ее жизнедеятельности. В ядре находится и воспроизводится
наследственная информация, зашифрованная в хромосомах и
определяющая все признаки не только клетки, но и всего организма в
целом. Ядро контролирует работу всех органелл клетки, определяет
интенсивность и направление проходящего в ней обмена веществ.
Строение ядра одинаково у всех ядерных организмов. Оно состоит из
ядерной оболочки, ядерного сока (кариолимфы, или нуклеоплазмы),
ядрышка и хроматина.
25.
Ядерная оболочка и кариолимфаЯдерная оболочка состоит из двух мембран, между которыми находится
перинуклеарное пространство, заполненное матриксом. Внешняя
мембрана ядерной оболочки, к которой часто прикрепляются рибосомы,
соединена с канальцами ЭР, благодаря чему ядро оказывается связанным
не только с цитоплазмой, но и с другими клетками. Ядерная оболочка не
сплошная, а прерывистая — в ней есть ядерные поры. Эти поры могут
открываться и закрываться, регулируя связь между ядром и цитоплазмой
(кариолимфой и гиалоплазмой). Число открытых пор зависит от
интенсивности процессов синтеза, происходящих в клетке.
Ядерный сок (кариолимфа, нуклеоплазма) — активный компонент ядра, в
котором осуществляется деятельность остальных компонентов ядра. Это
прозрачный коллоидный раствор, содержащий ферменты, необходимые
для синтеза всех трех видов PHК, а также для образования субъединиц
рибосом.
26.
ЯдрышкоЯдрышко — плотное сферическое тельце, обычно образующееся в
области вторичной перетяжки спутничных хромосом. В ядре содержится от
одного до нескольких ядрышек диаметром 1—3 мкм. Размеры ядрышек
зависят от интенсивности процесса биосинтеза белков.
Главная функция ядрышка — синтез рРНК. Соединяясь с белками,
поступающими из гиалоплазмы, рРНК образует субъединицы рибосом,
которые через поры в ядерной оболочке перемещаются в цитоплазму, где
из них на молекулах и PH К происходит самосборка рибосом.
Таким образом, ядрышки играют немаловажную роль в биосинтезе белков
клетки. При делении клетки, когда хроматин трансформируется в
хромосомы, ядрышки распадаются, а после окончания деления
формируются вновь.
27.
ХроматинВажнейшая часть ядра — хроматин, который представляет собой
молекулы ДНК, заключенные в белковые футляры. Скручивание этих
структур (дезоксирибонуклеопротеидов) в спираль позволяет
размещаться в ядре очень длинным (до 2 см) молекулам ДНК.
Хроматин представляет собой деспирализованные и гидратированные
хромосомы, сохраняющие свою индивидуальность.
Таким образом, хроматин — это особая форма существования
хромосом, их функционально активная форма. Хромосомы можно
наблюдать в клетке лишь во время ее деления, когда происходит
спирализация нитей (фибрилл) хроматина, в результате чего
хромосомы утолщаются, укорачиваются и становятся хорошо
заметными.
28.
29.
ВакуольВакуоли содержатся почти во всех растительных клетках. Они представляют
собой полости, заполненные клеточным соком и ограниченные от
цитоплазмы мембраной – тонопластом. Для большинства зрелых клеток
растений характерна центральная вакуоль. Она, как правило, настолько
крупна (70–90 % объема клетки), что протопласт со всеми органеллами
располагается в виде очень тонкого постенного слоя.
Клеточный сок, содержащийся в вакуоли, представляет собой водный
раствор различных веществ, являющихся продуктами жизнедеятельности
протопласта. В его состав могут входить углеводы (сахара и
полисахариды), белки, органические кислоты и их соли, минеральные
ионы, алкалоиды, гликозиды, танины и другие растворимые в воде
соединения.
Вакуоли в растительных клетках выполняют две основные функции:
накопление запасных веществ, отходов и поддержание тургора.
30.
31.
Клеточная оболочкаКлеточная оболочка – структурное образование на периферии клетки,
придающее ей прочность, сохраняющее ее форму и защищающее
протопласт. Оболочка, как правило, бесцветна и прозрачна, легко пропускает
солнечный свет. По ней могут передвигаться вода и растворенные
низкомолекулярные вещества. Оболочки соседних клеток соединены
пектиновыми веществами, образующими срединную пластинку.
Скелетным веществом оболочки клеток высших растений является целлюлоза.
Молекулы целлюлозы, представляющие собой очень длинные цепи, собраны
по нескольку десятков в группы – микрофибриллы. В них молекулы
располагаются параллельно друг другу и «сшиты» многочисленными
водородными связями. Они обладают эластичностью, высокой прочностью и
создают структурный каркас оболочки, а также погружены в ее аморфный
матрикс, состоящий в основном из гемицеллюлоз и пектиновых веществ.
Различают первичную и вторичную клеточные оболочки. Меристематические и
молодые растущие клетки, реже клетки постоянных тканей, имеют первичную
оболочку, тонкую, богатую пектином и гемицеллюлозой. Вторичная клеточная
оболочка образуется по достижении клеткой окончательного размера и
накладывается слоями на первичную со стороны протопласта.
32.
ПлазмодесмыПлазмодесмы присущи только растительным клеткам. Они
представляют собой тонкие цитоплазматические тяжи, соединяющие
соседние клетки. В одной клетке может содержаться от нескольких
сотен до десятков тысяч плазмодесм.
Стенки канала плазмодесмы выстланы плазмалеммой, непрерывной
по отношению к плазмалемме смежных клеток. По центру канала
проходит мембранный цилиндр – центральный стержень
плазмодесмы, соединенный с мембраной ЭПР. Между центральным
стержнем и плазмалеммой в канале находится гиалоплазма.
Плазмодесмы выполняют функцию межклеточного транспорта
веществ.
33.
34.
ПорыПорами называют неутолщенные места оболочки (углубления), на которых
отсутствует вторичная оболочка. Они содержат тончайшие отверстия, через
которые проходят плазмодесмы.
По форме порового канала различают простые и окаймленные поры. У
простых диаметр канала приблизительно одинаков на всем протяжении от
полости клетки до первичной оболочки и канал имеет форму узкого
цилиндра.
У окаймленных (трахеальные элементы) канал суживается в процессе
отложения вторичной оболочки; поэтому внутреннее отверстие поры,
выходящее в полость клетки, значительно уже, чем наружное, упирающееся в
первичную оболочку.
В смежных клетках поры располагаются напротив друг друга. Это облегчает
транспорт воды и растворенных веществ от клетки к клетке. Общие поры
имеют вид канала, разделенного перегородкой из срединной пластинки и
первичными оболочками (замыкающая пленка поры).
35.
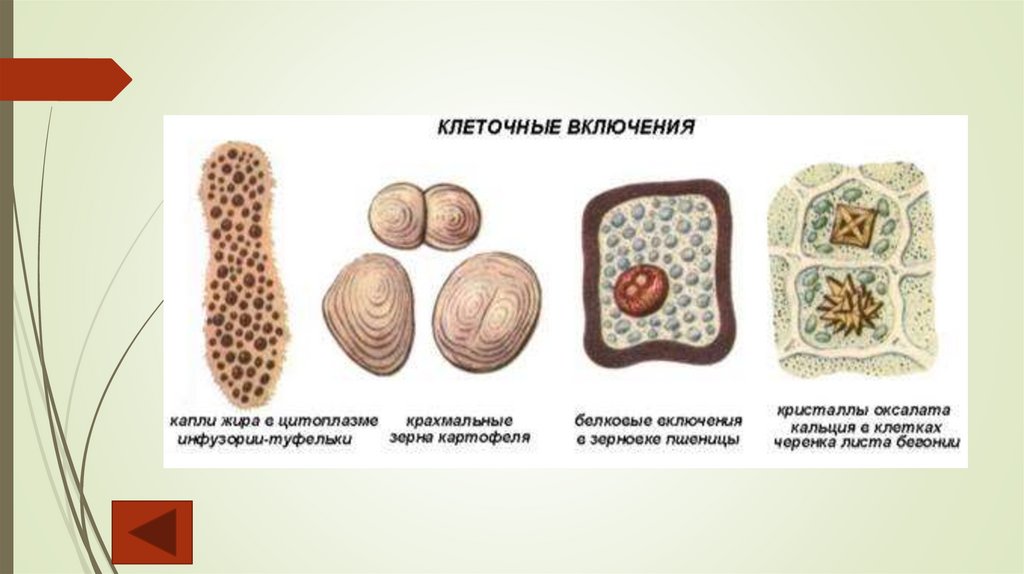
Цитоплазматические включенияВключения – это локальная концентрация некоторых продуктов обмена в
определенных местах клетки.
Крахмальные зерна образуются только в строме пластид живых клеток. В
хлоропластах на свету откладываются зерна ассимиляционного (первичного)
крахмала. Значительно большего объема достигают зерна запасного (вторичного)
крахмала, откладывающиеся в лейкопластах (амилопластах). Различают простые,
полусложные и сложные зерна.
Липидные капли накапливаются в гиалоплазме. Наиболее богаты ими семена и
плоды, где они могут быть преобладающим по объему компонентом протопласта.
Запасные белки чаще всего откладываются в вакуолях в виде зерен округлой или
овальной формы, называемых алейроновыми. Бывают простыми и сложными
(кристаллиты, глобоиды).
Кристаллы оксалата кальция – конечные продукты обмена; откладываются обычно в
вакуолях. По форме различают одиночные кристаллы, друзы (шаровидные
образования, состоящие из многих мелких сросшихся кристаллов), рафиды (мелкие
игольчатые кристаллы, собранные в пучки), кристаллический песок.
36.
37.
Ткани растенийПонятие о ткани. Классификация растительных тканей
Образовательные ткани
Покровные ткани
Всасывающие ткани
Ассимиляционные ткани
Запасающие ткани
Аэренхима
Механические ткани
Проводящие ткани
Выделительные ткани
38.
Понятие о ткани. Классификациярастительных тканей
Ткани – это устойчивые, т.е. закономерно повторяющиеся, группы
клеток, сходные по происхождению, строению и приспособленные к
выполнению одной или нескольких функций. Ткань называется простой,
если все ее клетки одинаковы по форме и функциям (паренхима,
склеренхима). Сложные ткани (покровные, проводящие) состоят из
клеток, неодинаковых по форме, внутреннему строению и функциям,
но связанных общим происхождением.
Все ткани растений можно разделить на две неравные по объему
группы: недифференцированные образовательные ткани, или
меристемы, и дифференцированные, или постоянные ткани.
По анатомо-физиологическому принципу выделяют образовательные,
ассимиляционные, запасающие, воздухоносные, покровные,
всасывающие, секреторные, механические и проводящие ткани.
39.
Образовательные тканиУ взрослых растений образование новых клеток приурочено к
определенным участкам – меристемам. Важная особенность
меристем состоит в том, что одни ее клетки (инициальные) способны
делиться неограниченное число раз, обеспечивая непрерывное
нарастание массы растения; другие клетки, являющиеся
производными от инициалей, делятся только ограниченное количество
раз и затем переходят к специализации.
Меристемы состоят из плотно расположенных мелких клеток с
большими ядрами и тонкими оболочками. По местоположению
меристемы можно разделить на апикальные, краевые, латеральные,
интеркалярные и раневые.
40.
Апикальные (верхушечные) располагаются на верхушке побегов и на кончике всехмолодых корешков и обеспечивают рост растения в длину. Топографически и
онтогенетически с апикальной меристемой побега связано образование краевой
(маргинальной) меристемы, формирующей пластинку листа.
Латеральные (боковые) способствуют росту растения в толщину и располагаются
параллельно боковой поверхности того органа, в котором они находятся.
Первичные латеральные меристемы (прокамбий, перицикл) возникают
непосредственно под апексами и являются их производными. Вторичные
меристемы (камбий и феллоген) образуются из тканей первичных меристем или из
клеток постоянных тканей в процессе упрощения их структуры и приобретения
свойств меристемы.
Интеркалярные (вставочные) располагаются обычно у основания междоузлий и
обеспечивают рост растения в длину. Они имеют временный характер и
превращаются в постоянные ткани.
Раневые (травматические) возникают в любой части растения при поранениях.
Клетки постоянных тканей, окружающие повреждение, дедифференцируются,
приобретают способность к делению и образуют раневую ткань - каллус. Клетки
каллуса постепенно превращаются в клетки постоянной ткани (раневой пробки).
41.
42.
Покровные тканиПокровные ткани защищают внутренние ткани растений от прямого влияния факторов внешней
среды, регулируют испарение и газообмен.
Эпидермис является сложной первичной покровной тканью и располагается на поверхности листьев
и молодых стеблей. Основные клетки эпидермиса относительно неспециализированы и слагают
основную массу ткани. Это живые, бесцветные, плотно прилегающие друг к другу клетки. Боковые
стенки основных клеток часто бывают извилистыми, что повышает прочность их сцепления. Наружные
стенки эпидермальных клеток наиболее утолщены и покрыты кутикулой – гидрофобным веществом,
препятствующим излишнему испарению воды. Поверх кутикулы обычно откладывается воск,
придающий поверхности органа сизоватый оттенок. У некоторых растений (например, у хвойных)
оболочки основных клеток одревесневают, а у хвощей – окремневают.
Устьица являются высокоспециализированными эпидермальными клетками, выполняющими
функцию газообмена и транспирации. У большинства наземных растений устьица располагаются
на нижней стороне листа. Они представляют собой отверстия, ограниченные двумя замыкающими
клетками. Рядом часто находятся побочные клетки, отличающиеся от основных клеток эпидермиса
размерами и формой и участвующие в движении устьиц. Замыкающие и побочные клетки
составляют устьичный аппарат.
Трихомы (волоски) представляют собой одно- и многоклеточные выросты эпидермиса. Если в
образовании выроста задействованы ткани, лежащие под эпидермисом, такие выросты называют
эмергенцами (шипы розы, волоски хмеля).
Трихомы можно разделить на кроющие и железистые. Кроющие волоски – обычно мертвые
образования, заполненные воздухом и покрывающие стебли и листья многих ксерофитов.
Железистые волоски – живые структуры, выделяющие смолы, сахара, эфирные масла, слизи. К
основным функциям трихом относятся защита органов растений от перегрева, выведение
токсичных солей из тканей листа, а также механическая и химическая защита от насекомых.
43.
44.
Перидерма. Продолжительность жизни эпидермиса различна у разных видов и их органов.Например, на листьях и стеблях травянистых растений эпидермис сохраняется до конца их
жизни. В стеблях и корнях, которые разрастаются в толщину путем вторичного роста, возникает
вторичная покровная ткань – феллема (пробка). Вместе с феллогеном (пробковым камбием)
и феллодермой она входит в состав перидермы, относимой в последнее время к особой
анатомо-топографической зоне.
Феллоген представлен меристематической тканью, формирующей перидерму. На срезе он
выглядит как слой, состоящий из прямоугольных клеток, уплощенных по радиусу органа.
Феллоген внутрь откладывает клетки феллодермы, снаружи – пробки. Феллодерма
представлена одним или несколькими слоями радиально расположенных паренхимных живых
клеток, изнутри примыкающих к феллогену, и выполняет функцию его питания. Молодые клетки
феллемы (пробки), отложенные феллогеном, имеют тонкие оболочки. Затем возникают
вторичные оболочки, содержащие ламеллы суберина и воска, вследствие чего их клеточная
стенка опробковевает, теряет живое содержимое и заполняется воздухом. Пробка защищает
растение от потери влаги, резких колебаний температуры, механических повреждений,
микроорганизмов.
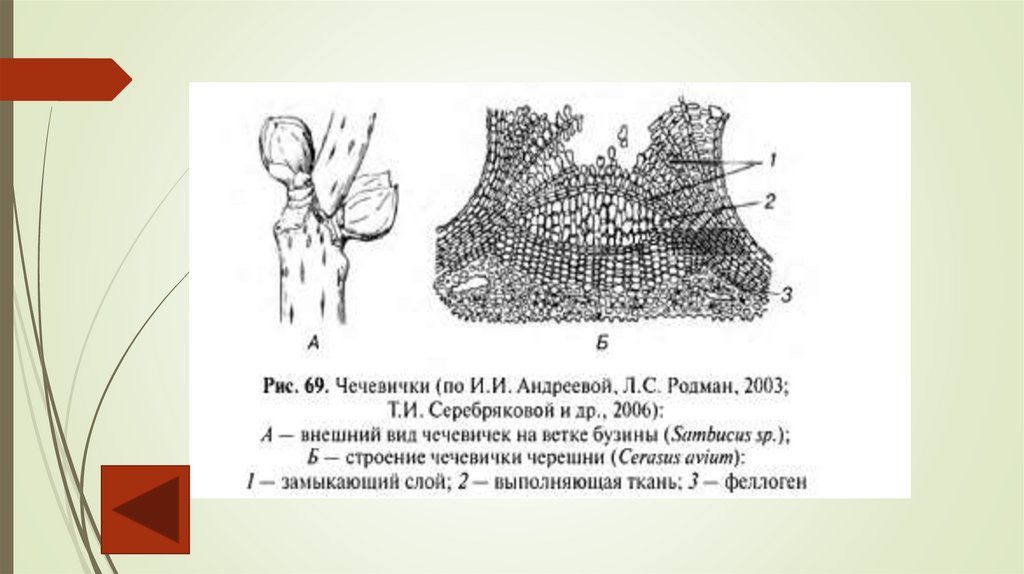
Лежащие под пробкой живые ткани испытывают потребность в газообмене. Поэтому в
перидерме формируются чечевички – участки, через которые происходит газообмен. На
поверхности молодых побегов деревьев и кустарников просматриваются бугорки.
Выполняющая чечевичку ткань образуется у побегов первого года еще до появления сплошного
слоя феллогена в результате деления паренхимных клеток, лежащих под устьичным
аппаратом. В последующие годы выполняющую ткань продуцирует и дополняет ее новыми
слоями феллоген. С наступлением холодов феллоген откладывает под выполняющей тканью
замыкающий слой из опробковевших клеток, который разрывается весной под напором новых
клеток.
45.
46.
Корка (ритидом) приходит насмену гладкой перидерме у
некоторых древесных
растений и состоит из
чередующихся слоев
перидермы и мертвой
паренхимы, т. е. имеет
сложный гистологический
состав. Она предохраняет
растение от механических
повреждений, резких
колебаний температуры,
пожаров.
47.
Всасывающие тканиВсасывающие ткани обеспечивают поступление в растение воды и
растворенных в ней веществ. Они различны по структуре и
распространению среди растений. Наиболее типична ризодерма –
наружный слой клеток молодых корешков с корневыми волосками. Через
него происходит всасывание воды и минеральных веществ. Остальные типы
всасывающих тканей, как правило, связаны с определенными условиями
или приурочены к какому-то таксону. Всасывающая ткань гаусторий
(присосок) хорошо развита у растений-паразитов (заразиха, повилика),
губчатая ткань веламен – на воздушных корнях орхидных. Поглощающие
ткани развиваются в прорастающих семенах (например, на щитке у
зародышей злаков) и в водопоглощающих волосках листьев. У некоторых
водных растений известны гидропоты – клетки или группы клеток на
поверхности листьев, избирательно поглощающие растворенные в воде
вещества.
48.
Ассимиляционные тканиГлавная функция ассимиляционных тканей — фотосинтез. Поэтому их еще называют
фотосинтезирующими паренхимами. Располагаются они в листьях и в стеблях молодых
побегов под прозрачной эпидермой, а также в цветках и плодах. Фотосинтезирующую
паренхиму можно встретить и в воздушных корнях растений-эпифитов (орхидей), и корнях
водных растений, т.е. корнях, которые развиваются на свету. Именно эти ткани придают
зеленую окраску листьям и стеблям. Ассимиляционную паренхиму стебля называют
хлоренхимой, листа — мезофиллом. В верхней части листовой пластинки обычно
располагается столбчатый, или палисадный, мезофилл, состоящий из плотно сомкнутых
клеток, вытянутых перпендикулярно поверхности листа и располагающихся в один или
несколько рядов; в нижней части пластинки сосредоточен губчатый мезофилл,
отличающийся более или менее изодиаметрическими клетками и большим числом
крупных межклетников.
Фотосинтезирующие паренхимы первичны по происхождению и имеют простое строение.
Они состоят из относительно однородных паренхимных клеток с тонкими стенками. В
постенном слое цитоплазмы много хлоропластов, общий объем которых может достигать
80% объема протопласта. В зависимости от условий освещенности хлоропластаы могут
перемешаться в цитоплазме, занимая наиболее оптимальное положение (недостаточное
или избыточное освещение тормозит процесс фотосинтеза). Иногда увеличение
поверхности постенного слоя цитоплазмы, а следовательно, и числа хлоропластов в
клетках мезофилла листа достигается образованием вдающихся внутрь клетки складок ее
стенки. Это можно наблюдать у многолетних игловидных листьев (хвоинок) сосны. Такой
мезофилл называют складчатым.
49.
Запасающие тканиК накоплению запасных веществ способны все живые клетки растения. Когда же
запасающая функция является для клеток ткани основной, говорят о запасающих тканях
(запасающих паренхимах). Запасающие паренхимы могут быть первичными и вторичными
по происхождению. Они состоят из живых тонкостенных изодиаметрических клеток,
особенности строения которых зависят от типа запасных веществ. Если это крахмал, то
клетки содержат много лейкопластов (амилопластов, или крахмальных зерен)', если сахара
или инулин, то в клетках есть крупная вакуоль, а если белок — много мелких вакуолей,
образующих со временем алейроновые зерна. В качестве запасного вещества в стенках
клеток может откладываться гемицеллюлоза (у семян финиковой пальмы).
У растений-суккулентов, обитающих в засушливых местах (агава, алоэ, кактусы), в клетках
запасающей паренхимы накапливается вода. В этом случае в вакуолях водоносных клеток
часто содержатся слизи, обладающие высокой водоудерживающей способностью.
В запасающих тканях растений накапливаются вещества, широко используемые человеком.
Пищевые сортовые растения обычно отличаются очень хорошо развитыми запасающими
паренхимами, находящимися в разных их органах. В запасающей паренхиме корнеплодов
моркови, редьки, репы, свеклы, корневых клубней батата, кочанов капусты, луковиц лука,
стеблей сахарного тростника накапливаются сахара; в побеговых клубнях стахиса и
топинамбура — инулин, в таких же клубнях картофеля, зерновках кукурузы, пшеницы, риса и
других культивируемых злаков — крахмал. Благодаря усилиям селекционеров в запасающих
тканях семянок масличных сортов подсолнечника содержится более 50% масла — намного
больше, чем у их дикорастущих родственников.
50.
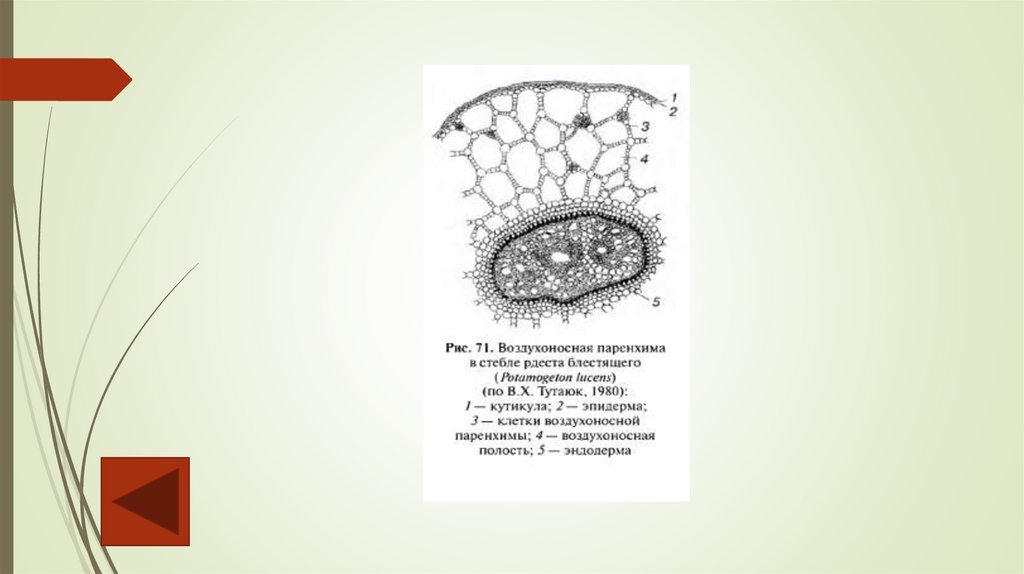
АэренхимаВ растениях довольно часто встречается ткань с большим числом очень крупных
межклетников — аэренхима (рис. 71). Соединяясь между собой, ее межклетники
образуют единую вентиляционную сеть. Поэтому эту ткань иногда называют
вентиляционной. Газовый состав межклетников значительно отличается от состава
атмосферного воздуха, так как клетки в процессе своей жизнедеятельности
выделяют в межклетники одни газы и поглощают другие. От условий обитания и
особенностей строения того или иного растения зависит характер циркуляции
газов по межклетникам, обеспечивающий нормальную жизнедеятельность
растительного организма. Проходя по всему телу растения от корней до листьев,
аэренхима выполняет вентиляционные и отчасти дыхательные функции,
обеспечивая близлежащие ткани растения кислородом. Газы в аэренхиме
перемещаются только путем диффузии. Аэренхиму можно рассматривать как
модификацию паренхимы. Однако форма ее клеток очень разнообразна. Они
могут быть округлыми (цветоножка кубышки), звездчатыми (стебель ситника) и др.
Аэренхима — первичная ткань. Ее клетки имеют тонкие первичные стенки,
покрытые со стороны межклетников слизью, уменьшающей транспирацию.
Вакуоли крупные, цитоплазма занимает постенное положение и содержит много
мелких лейкопластов.
Аэренхима хорошо развита в органах водных и болотных растений: в
цветоножках кубышки, стеблях белокрыльника, пушицы, рдеста, ситника, корнях
камыша. У водных растений аэренхима выполняет не только вентиляционные
функции. Воздушные полости в стеблях и листьях уменьшают удельный вес
растений, позволяя им свободно плавать в воде.
51.
52.
Механические тканиМеханические ткани обусловливают прочность растения. Стенки клеток, слагающих эти ткани,
утолщены. Механические ткани чаще всего выполняют свое назначение в сочетании с остальными
тканями растения, образуя их арматуру. К данному типу тканей относятся колленхима и склеренхима.
Колленхима – это механическая ткань, являющаяся первичной и служащая для укрепления молодых
стеблей и листьев во время роста. Клетки колленхимы живые, с неравномерно утолщенными
неодревесневшими стенками, вследствие чего они способны растягиваться при росте органа. В
зависимости от характера утолщения стенок различают уголковый (оболочка утолщается в углах, где
сходятся 3–5 клеток), пластинчатый (тангенциальные стенки утолщаются сплошными параллельными
слоями) и рыхлый (утолщение оболочек происходит на участках, примыкающих к межклетникам) типы
колленхимы.
Склеренхима состоит из мертвых клеток с равномерно утолщенными и, как правило,
лигнифицированными оболочками. Ее слагают два типа клеток: склеренхимные волокна и склереиды.
Склеренхимные волокна образованы мертвыми прозенхимными клетками с острыми концами и
толстыми оболочками, имеющими простые поры.
Склереиды представляют собой мертвые клетки разнообразной формы с очень толстыми оболочками,
пронизанными поровыми каналами. Клеточные стенки склереид одревесневают, в них откладываются
известь, кремнезем, суберин, вследствие чего протопласт отмирает. Встречаются в плодах, листьях,
стеблях, где располагаются поодиночке или группами (например, в мякоти плода груши). Склереиды,
располагающиеся плотно, без межклетников, образуют косточки плодов сливы, вишни, абрикоса,
скорлупу грецкого ореха.
Наряду с волокнами и склереидами, составляющими склеренхиму, в проводящей ткани высших
растений имеются клетки, также специализирующиеся на выполнении опорной функции. Это
древесинные (волокна либриформа) и лубяные волокна. Они отличаются от волокон типичной склеренхимы происхождением; поэтому рассматриваются как структурные элементы тех тканей, в которых
образовались.
53.
Проводящие тканиПроводящие ткани выполняют функцию транспортировки по растению
питательных веществ. Они образуют в теле растения непрерывную разветвленную
систему, соединяющую все его органы. Ткань, по которой передвигаются вода и
растворенные в ней минеральные вещества, называется ксилемой. Транспорт
продуктов ассимиляции осуществляет второй тип проводящей ткани – флоэма.
Ксилема, так же, как и флоэма, является сложной тканью и включает три типа
клеток: трахеальные элементы, механические волокна и клетки паренхимы.
Трахеальные элементы (трахеиды, сосуды) – это мертвые клетки вытянутой
формы с неравномерно утолщенными лигнифицированными оболочками,
пронизанными порами. Одревеснение оболочек происходило постепенно и
способствовало укреплению стенок водопроводящих элементов. У примитивных
организмов на тонкостенных оболочках сначала появлялись кольчатые, затем
спиральные утолщения и возникали кольчатые и спиральные трахеальные
элементы.
В процессе эволюции одревеснение распространилось почти на всю оболочку,
но в ней сохранились тонкостенные участки (поры), имеющие округлую или
продолговатую форму. Так возникли точечные и лестничные трахеальные
элементы, являющиеся разновидностями порового типа утолщения. Трахеиды
являются основными водопроводящими элементами плаунов, хвощей,
папоротников, голосеменных растений. Первичная клеточная оболочка на
клеточных оболочках у них не нарушена; поэтому передвижение воды
осуществляется путем фильтрации через поры.
54.
Сосуды характерны для покрытосеменных растений. Членики сосудов располагаются одинпод другим, образуя длинную полую трубку. Основное отличие сосудов от трахеид состоит в
том, что их поперечная перегородка имеет сквозные отверстия (перфорации), вследствие
чего значительно увеличивается скорость передвижения воды.
Членики сосудов возникают из живых клеток, которые имеют тонкие оболочки и растут в
длину и ширину. Затем начинает откладываться вторичная оболочка (не откладывается в
местах образования пор и перфораций). Поперечные стенки члеников сосудов в местах
перфораций растворяются, начинается проведение воды.
Сосуды являются важнейшим эволюционным приобретением растений. Они начали
появляться в независимых эволюционных группах (у селягинеллы, орляка, эфедры) и
окончательно закрепились у покрытосеменных, явившись важным фактором их
процветания и приспособления к сухопутным условиям. Скорость передвижения воды по
сосудам у некоторых высоких деревьев может достигать 8 м/ч (в среднем – 1 м/ч).
Древесинные волокна (волокна либриформа) выполняют опорную и защитную функции для
трахеальных элементов и паренхимы. Они эволюционно возникли из трахеид, их
преобразование шло в направлении потери проводящей функции, преобразования
окаймленных пор в простые и повышения механической прочности. Древесинная
паренхима часто окружает трахеальные элементы. Она регулирует поступление и
скорость движения растворов и запасает питательные вещества. Собранные в
горизонтальные полосы участки паренхимных клеток образуют так называемые
древесинные лучи, передающие растворы в радиальном направлении. Рассеянная среди
трахеальных элементов паренхима, в виде вертикальных тяжей тянущаяся вдоль осевых
органов, называется древесинной, или тяжевой. Клетки паренхимы могут образовывать выросты в полость сосудов – тиллы. Тиллообразование усиливает механическую прочность
центральной части стволов деревьев.
55.
Флоэма – это ткань сосудистых растений, проводящая органическиевещества. Первичная флоэма, которую подразделяют на протофлоэму и
метафлоэму, дифференцируется из прокамбия, вторичная (луб) является
производной камбия. В состав флоэмы входят ситовидные элементы, клеткиспутницы, лубяные волокна и клетки паренхимы.
Ситовидные элементы – это живые прозенхимные клетки, выполняющие
проводящую функцию. На их стенках находятся ситовидные поля – участки
клеточной оболочки, пронизанные многочисленными отверстиями, через
которые посредством плазмодесм сообщаются протопласты соседних
ситовидных элементов. Различают два типа ситовидных элементов:
ситовидные клетки (длинные с заостренными концами, ситовидные поля – по
продольным стенкам, лишены клеток-спутниц, ядро уменьшено или
фрагментирует) и ситовидные трубки (состоят из коротких члеников,
расположенных друг над другом, ситовидные поля – на поперечных стенках,
образующих ситовидную пластинку, имеют клетки-спутницы и в зрелом
состоянии лишены ядра). Ситовидные трубки функционально более
совершенны, чем ситовидные клетки, и характерны для покрытосеменных
растений.
56.
57.
Выделительные тканиСекреторными (выделительными) тканями принято называть структуры, выделяющие терпены,
полисахариды, соли, воду и другие вещества. Иногда эти вещества представляют собой
конечные продукты обмена, иногда – выполняют функцию защиты от насекомых, от поедания
животными, предохраняют от загнивания древесины. Различают наружные и внутренние
выделительные ткани.
Перечислим наружные секреторные ткани.
1. Железистые волоски представляют собой трихомы, или эмергенцы (производные
эпидермиса и лежащих под ним тканей), и являются обычно многоклеточными структурами.
Выполняют выделительную и защитную функцию.
2. Гидатоды выделяют воду и соли на поверхность листа из его внутренних частей. Этот
процесс называют гуттацией. Гидатоды обычно располагаются по краю листа. Гуттация
происходит при временном избыточном поступлении воды и затрудненной транспирации
(по утрам – у земляники, манжетки).
3. Нектарники выделяют сахаристую жидкость, привлекающую насекомых. Обычно
располагаются в цветках.
4. Осмофоры – желёзки, продуцирующие аромат у многих растений. Выделяют летучий
секрет, представленный в основном эфирными маслами. Служат для привлечения
насекомых-опылителей.
5. Пищеварительные желёзки встречаются у насекомоядных растений и выделяют
пищеварительные ферменты, кислоты и другие вещества, с помощью которых
перевариваются пойманные животные.
58.
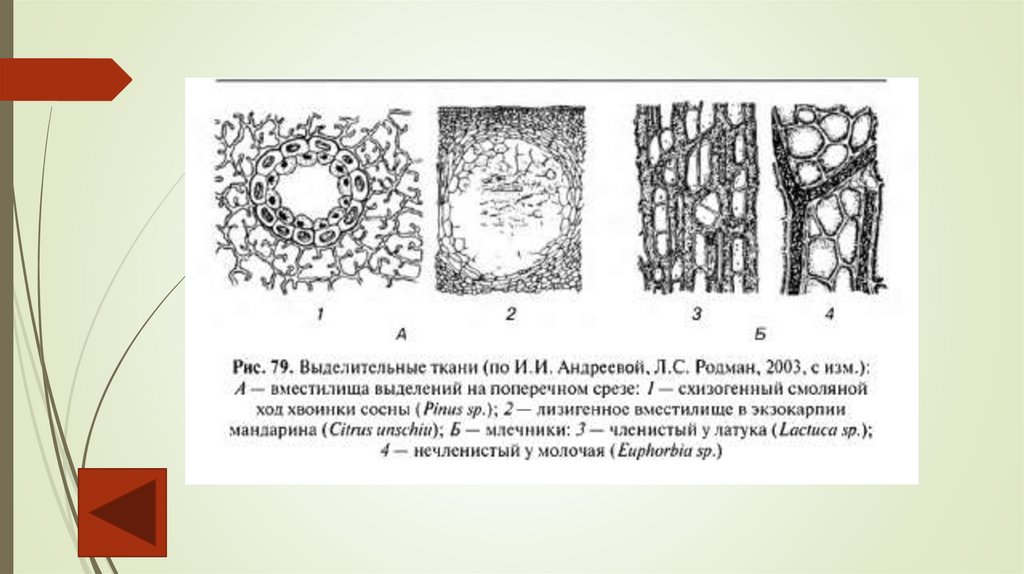
Перечислим внутренние секреторные ткани.1. Секреторные идиобласты – выделительные клетки, рассеянные среди других
тканей и накапливающие различные вещества (оксалат кальция, терпены, слизи,
танины). Оболочка выделительных клеток утолщается, в ней для изоляции ядовитого
секрета от окружающих клеток откладывается суберин. Эфирномасляные
идиобласты встречаются у лавровых, перечных.
2. Вместилища выделений встречаются у растений разных систематических групп,
они разнообразны по форме, величине, происхождению. По происхождению
выделяют лизигенные и схизогенные вместилища.
Лизигенные возникают в результате растворения группы клеток, обособившихся
внутри какой-либо ткани. Их деление приводит к образованию небольшого очага
мелких клеток, вырабатывающих секрет. Впоследствии оболочки растворяются , и на
их месте формируется полость, заполненная секретом. Лизигенные вместилища
встречаются в кожуре цитрусовых, листьях эвкалипта.
Схизогенные возникают из межклетников при отделении клеток друг от друга. При
этом клетки, прилегающие к вместилищу, становятся эпителиальными, т. е.
вырабатывают и выделяют в полость экскреторное вещество. Эпителий изолирует
секрет от живых тканей. Схизогенные вместилища хорошо развиты у хвойных
(смоляные ходы), некоторых сложноцветных, зонтичных.
3. Млечники – клетки или ряды клеток, содержащие в вакуолях млечный сок (латекс).
Он может содержать смолы, каучук, эфирные масла, алкалоиды. Млечники бывают
двух типов: членистые и нечленистые.
59.
60.
Вегетативные органы растенийКорень и корневая система
Побег и система побегов
61.
Корень и корневая системаОпределение и функции корня
Зоны молодого корневого окончания
Корневая система
62.
Определение и функции корняКорень — основной вегетативный орган растения, выполняющий в типичном случае
функцию почвенного питания. Корень — осевой орган, обладающий радиальной
симметрией и неопределенно долго нарастающий в длину благодаря деятельности
апикальной меристемы. От побега он морфологически отличается тем, что на нем
никогда не возникают листья, а апикальная меристема всегда прикрыта чехликом.
Кроме главной функции поглощения веществ из почвы, корни выполняют и другие
функции:
а)
корни укрепляют («заякоривают») растения в почве, делают возможным
вертикальный рост и вынесение побегов вверх;
б)
в корнях синтезируются различные вещества (многие аминокислоты, гормоны,
алкалоиды и пр.), которые затем передвигаются в другие органы растения;
в)
в корнях могут откладываться запасные вещества;
г)
корни взаимодействуют с корнями других растений, микроорганизмами,
грибами, обитающими в почве.
63.
Зоны молодого корневого окончанияНежные клетки апикальной меристемы, словно живым наперстком, всегда
прикрыты чехликом. Лишь в редчайших случаях (у некоторых водных растений и
паразитов) чехлик отсутствует. Чехлик состоит из живых клеток и постоянно
обновляется: по мере того, как с его поверхности слущиваются старые клетки, на
смену им, изнутри, апикальная меристема образует новые молодые клетки.
Наружные клетки отслаиваются от поверхности чехлика, еще будучи живыми. Они
продуцируют обильную слизь, которая облегчает продвижение. Центральная
(осевая) часть чехлика составляет так называемую колумеллу (колонку), в клетках
которой содержится много крахмальных зерен. Предполагают, что эти зерна
служат статолитами (греч. статос — стоящий; литое — камень), т. е. способны
перемещаться в клетке при изменении положения кончика корня в пространстве,
благодаря чему корень изгибается и растет в прежнем направлении.
Под чехликом находится зона деления, клетки которой имеют характер
меристемы. Почти все делящиеся клетки сосредоточены в этой зоне, имеющей
длину менее 1 мм. Зону деления часто можно отличить на живом корешке по
желтоватой окраске, зависящей от того, что клетки меристемы заполнены
цитоплазмой и не имеют заметных вакуолей.
64.
Зоны молодого корневого окончанияВслед за этой зоной корень остается гладким, но становится светлым и как бы
прозрачным. Это — зона растяжения. В этой зоне клетки сильно увеличиваются
в продольном направлении (вдоль оси корня), однако клеточные деления уже
почти отсутствуют и объем корня увеличивается за счет общего обводнения
клеток и появления больших вакуолей. Растущий корень упругим кончиком
(находящимся в состоянии тургора) с большой силой проталкивается между
частичками почвы. Протяженность зоны растяжения обычно невелика и не
превышает нескольких миллиметров.
Клетки, оказавшиеся в конце зоны растяжения, резко прекращают растяжение
и уже более не смещаются относительно частиц почвы. Теперь на ризодерме
возникают многочисленные корневые волоски, плотно охватывающие частицы
почвы и как бы срастающиеся с ними. Зону корня, несущую волоски, называют
зоной поглощения, или всасывания. Прямые опыты показали, что подавляющую
массу воды и солей молодые корни воспринимают именно в этой зоне. Зона
поглощения имеет протяжение до нескольких сантиметров.
65.
66.
Корневая системаКорневая система – это совокупность всех корней одной особи. В состав корневых систем
могут входить корни различной морфологической природы. Среди них различают:
1) главный корень, развивающийся из зародышевого корешка;
2) боковые корни, всегда возникающие на корне и образующиеся обычно в зоне
проведения из перицикла (т. е. они не мешают корню расти);
3) придаточные корни, возникающие на стеблях, листьях из меристем, сердцевинных лучей
и других тканей. Образуются при черенковании, поранениях.
Выделяют такие типы коренных систем:
аллоризная (стержневая), составленная из хорошо развитого главного и боковых корней.
Такой тип корневой системы характерен для голосеменных и покрытосеменных двудольных
растений;
первично гоморизная (плауны, хвощи, папоротники), представленная только придаточными
корнями;
вторично гоморизная (мочковатая), при которой главный корень закладывается, но рано
отмирает или вообще не развивается, а корневая система представлена придаточными
корнями. Характерна для однодольных растений.
67.
68.
Побег и система побеговПобег
Строение и деятельность апикальной меристемы побега
Морфологическое строение стебля
Первичное строение стебля
Стелярная теория
Типы вторичных утолщений стебля
Лист и его функции
Морфологическое строение листа
Анатомия листа
69.
ПобегПобег – это основной орган семенных растений, выполняющий
функцию воздушного питания. Состоит из оси (стебля) с
расположенными на ней (нем) листьями и почками.
В отличие от корня, побег расчленен на междоузлия и узлы с одним или
несколькими листьями, прикрепленными к каждому узлу. Угол между
стеблем и листом в месте его отхождения называют листовой пазухой.
Междоузлия могут быть длинными, и тогда побег называют удлиненным;
если же междоузлия короткие, побег называют укороченным.
Побег, развивающийся из зародышевого стебелька, называют главным.
В узлах расположены боковые почки, из которых формируются
боковые побеги. Образуется система побегов.
70.
Строение и деятельность апикальноймеристемы побега
Нарастание побега осуществляется за счет апикальных меристем. Апекс побега отличается от
апекса корня отсутствием чехлика и наличием листовых зачатков (примордиев). Гладким остается
лишь кончик апекса, который называется конусом нарастания.
В апексе побега имеются инициальные клетки, делящиеся непрерывно. У многих споровых
имеется лишь одна инициаль тетраэдрической формы. У семенных растений апекс, как правило,
представлен массивом инициальных клеток. В апексе цветковых растений выделяют два участка
инициалей. На дистальном конце апекса хорошо выделяются два (реже 1–4) слоя клеток,
делящихся антиклинально (перпендикулярно поверхности) и обеспечивающих рост апекса в
поверхность. Эта зона инициальных клеток называется туникой. Несколько слоев клеток, лежащих
под туникой, делящихся во всех направлениях и увеличивающих объем апекса, называют
корпусом.
Там, где должен возникнуть листовой зачаток, деление клеток усиливается, они становятся
периклинальными; стираются границы между внутренним слоем туники и корпусом. Эта
меристема называется периферической (область инициального кольца). В центре оси апекса
обособляется стержневая меристема, образующая сердцевину.
Теорию «туники и корпуса» сформулировал немецкий ботаник А.Шмидт (1924). Из наружного слоя
туники возникает протодерма, образующая впоследствии эпидермис. На уровне первых листовых
зачатков (в пределах периферической зоны) между бугорком и осью закладывается прокамбий,
дающий начало первичным проводящим тканям. Основная меристема, образующая ткани
первичной коры, формируется либо из внутреннего слоя туники, либо из наружных слоев корпуса.
71.
Морфологическое строение стебляСтебель – это осевая часть побега растений, состоящая из узлов и междоузлий и имеющая
неограниченный рост. Рост стебля в длину осуществляется за счет деятельности
верхушечных и вставочных меристем. Стебель несет на себе листья, почки, генеративные
органы и выполняет главным образом опорную, проводящую, листонесущую функции.
Иногда стебель функционирует как запасающий орган, служит для прикрепления к опоре,
размножения и т. д.
По направлению и способу роста стебли чаще бывают прямостоячими, реже
приподнимающимися (клевер луговой), ползучими (клевер ползучий, костяника, земляника),
стелющимися (огурец, кабачок), вьющимися (горец вьюнковый), цепляющимися (хмель,
подмаренник цепкий), лазящими (горох) и т. д.
Стебель обычно имеет более или менее цилиндрическую форму и в поперечном сечении
бывает округлым. Однако некоторые семейства или отдельные виды растений отличаются
другой формой стебля, что является их систематическим признаком. Например, для
семейства губоцветных характерен четырехгранный стебель, для осоковых – трехгранный, а
для злаков - полый в междоузлиях стебель-соломина.
У древесных растений стебли многолетние. Главный стебель дерева называется стволом,
стебли кустарников – стволиками. У многолетних травянистых растений стебли, как
правило, живут один год.
72.
Первичное строение стебляВ стебле, имеющем первичное строение, различают эпидермис,
первичную кору и стелу (центральный цилиндр). Эпидермис
располагается снаружи. В состав первичной коры могут входить
ассимиляционная, механическая, запасающая, выделительная ткани,
аэренхима.
Стела имеет более сложное строение. Первичные проводящие ткани
обычно собраны в один или несколько проводящих пучков различного
типа. Расположение первичных проводящих элементов зависит от
заложения и функционирования прокамбия. Если он закладывается
участками, возникает пучковый тип, если сплошным слоем –
непучковый.
В центре стебля обычно находится сердцевина или воздушная полость
(например, у тыквы, злаков).
73.
Стелярная теорияСтела – совокупность первичных проводящих пучков вместе с заключенными между ними прочими
тканями и примыкающим к коре перициклом.
В 1886 году Ф. ван Тигем и А.Дулио впервые ввели понятие «стела», заложив основы стелярной теории
– учения о типах строения и закономерностях эволюции центрального цилиндра всех высших
растений.
Наиболее древний и примитивный тип стелы – протостела – присущ первым наземным растениям
(риниофитам). Последующая эволюция стелы была связана с увеличением поверхности
проводящих тканей.
Из протостелы развилась актиностела, ксилема которой на поперечном срезе имеет вид звезды.
Данный тип стелы также характерен для примитивных растений (плауновидных, вымерших
хвощевидных). Переход от протостелы к актиностеле связан с возникновением пучков, идущих в
боковые органы; кроме того, в актиностеле проводящие ткани имеют большую поверхность
соприкосновения с окружающими живыми тканями, что способствует лучшему проведению
веществ. Результат дальнейшей специализации актиностелы – появление плектостелы, характерной
для осевых органов плауновидных. В плектостеле ксилема представлена отдельными участками,
расположенными параллельно или радиально друг к другу.
В процессе эволюции протостела дала также начало сифоностеле. Сифоностела имеет трубчатое
строение и обладает сердцевиной. Возникновение сифоностелы обеспечило существование
более крупных растений, т. к. расположение ксилемы на периферии сделало стебли более
прочными. Возникновение сердцевины увеличило поверхность соприкосновения проводящих тканей
с живыми клетками, а также создало возможность запасания питательных веществ. Известны два
типа сифоностелы: эктофлойная (флоэма охватывает ксилему снаружи) и амфифлойная
(флоэма окружает ксилему снаружи и изнутри).
74.
75.
Стелярная теорияДальнейшее эволюционное усложнение стелы связано с развитием крупных
боковых органов (вай, листьев). В местах их отхождения от стебля в стеле, в
области узлов, образуются большие листовые лакуны (прорывы), заполненные
паренхимой. Вследствие этого сифоностела паренхимными полосами
(радиальными лучами), идущими от сердцевины к коре в радиальном
направлении, рассекается на отдельные пучки, сохраняющие общее
положение по цилиндрической поверхности (на поперечном срезе – кольцом).
Возникают диктиостела, артростела и эустела.
Диктиостела (характерна для папоротников, например для орляка, щитовника,
кочедыжника) имеет вид сетчатой трубки. Она возникает из амфифлойной
сифоностелы вследствие образования большого количества листовых
прорывов. Эустела встречается у семенных растений и составлена из
коллатеральных открытых пучков, расположенных по кругу. Артростела
(характерна для хвощей) является разновидностью эустелы, представлена
закрытыми пучками, расположенными вокруг центральной полости стелы и
соединяющимися в узлах.
Последнее звено в эволюции стелы стебля представляет собой атактостела
однодольных растений. Ей присущи закрытые коллатеральные пучки, сложная
картина прохождения пучков по стеблю (дугообразные индивидуальные
листовые следы) с их разбросанным расположением по всему поперечному
сечению стебля
.
76.
Типы вторичных утолщений стебляДля большинства двудольных и голосеменных растений характерно вторичное утолщение
стеблей. Это изменение связано с деятельностью камбия и феллогена. Возникновение
камбия и интенсивность его работы неодинаковы у различных растений. Различают
следующие типы вторичных утолщений.
1. При пучковом типе в обособленных прокамбиальных пучках сначала возникает пучковый
камбий. Затем между его разобщенными прослойками появляются перемычки
межпучкового камбия. Пучковый камбий начинает откладывать проводящие элементы, а
межпучковый – паренхиму или механические элементы, вследствие чего проводящие пучки
остаются хорошо различимыми (Aristolochia-tip).
2. При переходном типе в прокамбиальных пучках возникает пучковый камбий, затем между
проводящими пучками появляются перемычки межпучкового камбия. После этого камбий на
всем протяжении начинает откладывать проводящие элементы (Ricinus-tip). У некоторых
растений с широкими первичными лучами отдельные участки межпучкового камбия
образуют более мелкие вторичные проводящие пучки (Helianthus-tip).
3. При непучковом типе камбий закладывается как непрерывный слой (кольцо) из сплошного
прокамбия и начинает откладывать на всем протяжении проводящие элементы (Saponariatip). У многих древесных растений первичные пучки тесно сближены, поэтому при вторичном
утолщении возникают слои проводящих элементов, разделенные радиальными лучами (Tiliatip).
77.
Лист и его функцииЛист – это один из основных органов высших растений, занимающий боковое
положение на стебле и выполняющий функции фотосинтеза, газообмена и
транспирации.
Первые листовые органы – семядоли – возникают в результате дифференциации
меристематического тела предзародыша. Все последующие листья возникают
из экзогенных выростов (листовых примордиев) на апексе побега. Большинство
листьев имеют более или менее плоскую форму и дорсовентральное строение,
выражающееся в различии его верхней и нижней сторон. Различают листья:
1) бифациальные (двусторонние) – листья типичного строения, у которых верхняя
и нижняя стороны различаются морфологически (характер жилок, опушение) и
анатомически;
2) эквифациальные – листья, обе стороны которых имеют одинаковое
морфологическое и анатомическое строение (злаки);
3) унифациальные (с одной поверхностью) – листья либо округлые в сечении
(лук), либо уплощенные с боков (гладиолус, ирис), у которых вся поверхность
пластинки соответствует лишь нижней стороне.
78.
Морфологическое строение листаЛист состоит из листовой пластинки и черешка. Место прикрепления листа к стеблю
называют основанием. Листья, имеющие черешок, называют черешковыми. Листья без
черешка называют сидячими.
У большинства однодольных и некоторых двудольных (зонтичные) основание листа
разрастается, охватывая узел целиком, и образует трубку, называемую влагалищем листа.
У некоторых видов растений (представители семейств бобовых, розоцветных и др.) у
основания листа возникают парные выросты (прилистники), защищающие лист на ранних
стадиях его развития.
Листья располагаются на стебле в определенном порядке. Он отражает симметрию в
структуре побега. Различают четыре типа размещения листьев: спиральное, двурядное,
супротивное и мутовчатое. Спиральное (очередное) листорасположение – это
расположение листьев по спирали; при этом от каждого узла стебля отходит один лист.
Двурядное (супротивно-очередное) листорасположение характеризуется тем, что листья
располагаются по одному в каждом узле, но обязательно на противоположной стороне
оси. При супротивном листорасположении листья сидят на каждом узле попарно, один
против другого, при мутовчатом – на одном узле располагаются три листа и более. Обычно
листья располагаются так, чтобы обеспечить наименьшую взаимную затеняемость. Это
явление получило название листовой мозаики.
Листья морфологически очень разнообразны; поэтому нет признака, позволяющего их
классифицировать. Существующие классификации искусственны, т.к. основаны только на
внешнем сходстве.
79.
Различают простые и сложные листья. Простые состоят из однойпластинки. Сложные имеют общий черешок (рахис), к
которому прикреплены на черешочках несколько листовых
пластинок, называемых листочками сложного листа. Сложные
листья могут быть тройчато-, пальчато- и перистосложными.
Перистосложные листья, имеющие на конце оси непарный
листочек, называют непарноперистосложными, а листья с осью,
переходящей в усик или шипик, – парноперистосложными.
Если общий черешок разветвлен, образуются
многократносложные листья: дваждыперистосложные,
триждыперистосложные и т.д.
80.
81.
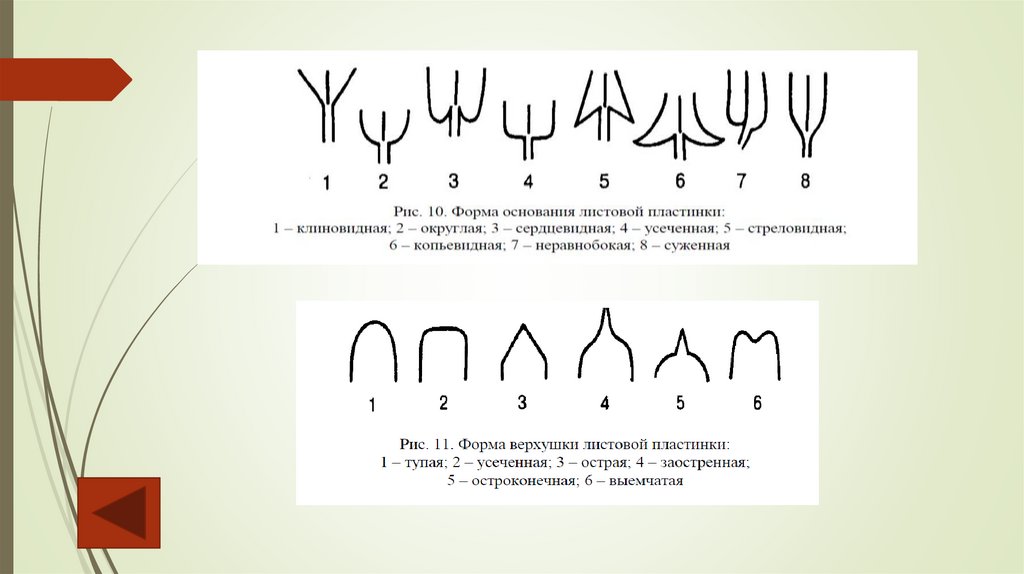
Форма, рассечение листовых пластинок простыхлистьев и листочков сложного листа могут быть
весьма разнообразными. Форма основания
листовой пластинки бывает клиновидной, округлой,
сердцевидной, усеченной, стреловидной,
копьевидной и т.д. Верхушка листа может быть тупой,
усеченной, острой, заостренной, остроконечной,
выемчатой.
82.
83.
Форма листовой пластинки определяется соотношением ее длины иширины и положением наиболее широкой ее части.
84.
Край листа имеет вырезки разной глубины. В тех случаях, если они незаходят глубже чем на 1/ 4 ширины полупластинки, лист называют цельным,
а его край – изрезанным. Край листовой пластинки может быть зубчатым,
пильчатым, двоякопильчатым, городчатым, выемчатым, волнистым. Листья
с цельными краями называют цельнокрайними.
85.
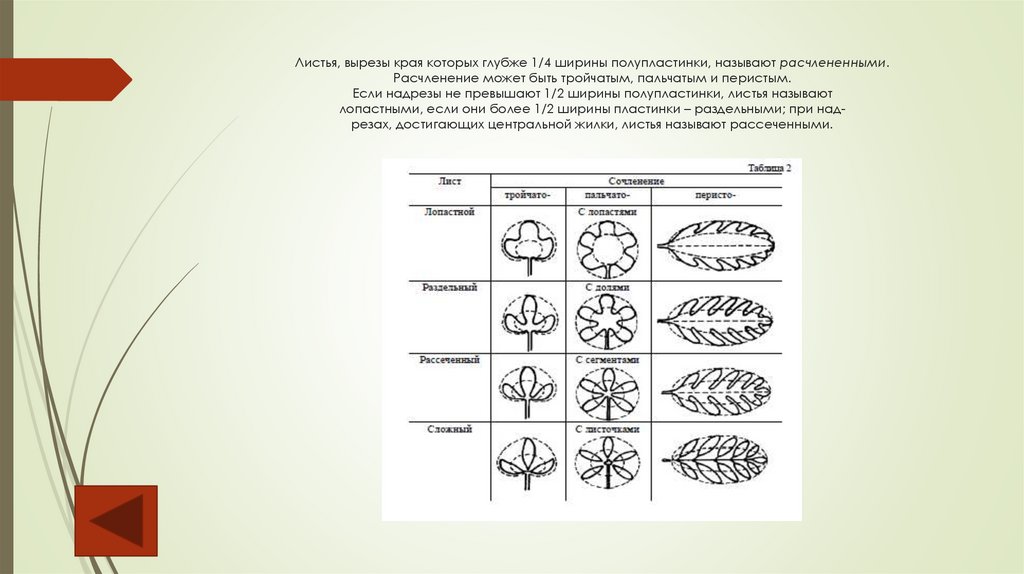
Листья, вырезы края которых глубже 1/4 ширины полупластинки, называют расчлененными.Расчленение может быть тройчатым, пальчатым и перистым.
Если надрезы не превышают 1/2 ширины полупластинки, листья называют
лопастными, если они более 1/2 ширины пластинки – раздельными; при надрезах, достигающих центральной жилки, листья называют рассеченными.
86.
Одним из важных морфологических признаков листа является типжилкования. Жилками условно называют проводящие пучки листа
вместе с сопровождающими тканями, которые хорошо заметны с
нижней стороны. Листья с дихотомическим жилкованием редко
встречаются у современных растений. Основные типы жилкования у
двудольных растений – сетчатое с перистым расположением
основных жилок (имеется главная жилка, от нее отходят
соединяющиеся, более тонкие и многократно ветвящиеся боковые
жилки) и сетчатое с пальчатым расположением основных жилок
(они в виде лучей отходят из одной точки в основании листовой
пластинки). Однодольным растениям свойственно дуговидное и
параллельное жилкование.
87.
Анатомия листаАнатомическое строение листа определяется его главной функцией – фотосинтезом.
Поэтому основной тканью листа является ассимиляционная, или мезофилл. Остальные
ткани листа обеспечивают работу мезофилла и поддерживают связь с окружающей
средой.
С верхней и нижней стороны лист покрыт эпидермисом, выполняющим функции
защиты, газообмена и транспирации. Обычно эпидермис однослойный, бесцветный.
Нижний и верхний эпидермис часто отличаются по опушению, характеру жилкования,
цвету, расположению устьиц и другим признакам. На верхнем эпидермисе могут
откладываться слои кутикулы и воска, снижающие испарение воды. Устьица у
большинства наземных растений расположены с нижней стороны.
Под эпидермисом находится мезофилл, представленный несколькими слоями
ассимиляционной ткани, в котором происходят основные физиологические
процессы: фотосинтез и дыхание растений. У дорсовентральных листьев мезофилл
дифференцирован на столбчатый (палисадный) и губчатый. Столбчатый
располагается в один или несколько слоев под верхним эпидермисом. Его клетки
плотно сомкнуты, имеют вытянутую форму и содержат около 75% хлоропластов листа.
Основная функция столбчатого мезофилла – фотосинтез. Губчатый мезофилл
расположен под столбчатым (с нижней стороны). Он представлен округлыми
клетками с большими межклетниками и выполняет функцию газообмена. Для
унифациальных листьев характерен однородный мезофилл.
88.
В листе проводящие ткани формируют закрытые коллатеральные пучки, причем ксилема всегдаповернута к верхней стороне, а флоэма – к нижней. Флоэму и ксилему в крупных проводящих
пучках окружают клетки склеренхимы, а в мелких – обкладочные клетки, контролирующие
проведение веществ. Мезофилл листа густо пронизан жилками, образующими единую
проводящую систему, которая связана с проводящей системой стебля.
Роль арматурных тканей в листе выполняют склеренхимные волокна, отдельные склереиды и тяжи
колленхимы, существенно повышающие прочность данного органа.
По морфологическому и анатомическому строению листа различаются светолюбивые
(гелиофиты) и тенелюбивые (сциофиты) растения.
У тенелюбивых тонкие листья с широкой листовой пластинкой располагаются горизонтально и
имеют темно-зеленую окраску (высокая концентрация хлорофилла). Клетки эпидермиса
крупные, с тонкими наружными стенками, кутикула развита слабо. Устьица крупные, лежат
неглубоко. Столбчатый мезофилл либо однослойный (его клетки имеют трапециевидную
форму), либо не выражен. Хлоропласты крупные, занимают постенное положение. У
тенелюбивых растений механические и проводящие ткани развиты слабо.
Листья многих светолюбивых растений имеют толстую, жесткую пластинку с хорошо развитой
многослойной столбчатой паренхимой, часто сильно рассеченной. Если лист освещается с двух
сторон, то столбчатая паренхима располагается под верхним и нижним эпидермисом.
Эпидермис состоит из мелких толстостенных клеток и покрыт мощным слоем кутикулы. Для
листьев характерны большое количество устьиц погруженного типа, густая сеть жилок, хорошо
развитые механические ткани или запасающая воду паренхима.
89.
Генеративные органы растенийЦветок
Морфологическая характеристика соцветий
Классификация соцветий
Опыление растений
Строение семени
Прорастание семян
Строение и классификации плодов
Распространение плодов и семян
90.
ЦветокМорфологические типы цветков
Строение околоцветника
Андроцей
Строение гинецея
Семязачаток
Двойное оплодотворение и его значение
Типы отклонений от нормального оплодотворения у цветковых растений
Формула и диаграмма цветка
91.
Морфологические типы цветковЦветок – это высокоспециализированный орган полового и бесполого размножения
покрытосеменных растений. В цветках происходят процессы микро– и мегаспорогенеза, микро–
и мегагаметогенеза, опыления, оплодотворения, образования плода и семени. Абсолютное
большинство цветковых растений имеют обоеполые цветки.
Цветок состоит из стерильной и фертильной частей. К стерильной части относится околоцветник,
к фертильной – андроцей и гинецей. Цветоножка представляет собой междоузлие под цветком и
соединяет его со стеблем. Цветки, не имеющие цветоножек, называют сидячими. На цветоножке
могут располагаться листочки, называемые прицветниками. Цветоложе – это расширенная часть
цветоножки, к которой прикрепляются все части цветка. Чаще всего оно бывает плоским, реже –
выпуклым (лютик, малина) или вогнутым.
Части цветка могут размещаться на цветоложе по спирали (спиральное расположение) или по
кругу (циклическое расположение). Иногда может быть смешанное (гемициклическое)
расположение, когда, например, части околоцветника располагаются по кругу, а тычинки и
пестики – по спирали.
Для некоторых групп ветроопыляемых растений характерны раздельнополые цветки, состоящие
из одних тычинок (мужские, или тычиночные) или из одних пестиков (женские, или пестичные).
Такие цветки обычно имеют редуцированный околоцветник или лишены его. Раздельнополые
цветки могут развиваться на одной и той же особи. Такое растение называется однодомным.
Если цветки образуются на разных особях, растение называется двудомным.
В зависимости от симметрии различают цветки актиноморфные (правильные, многосторонне
симметричные), билатерально-симметричные (с двумя осями симметрии), зигоморфные (с
одной осью симметрии) и асимметричные. Симметрия цветка определяется в основном
структурой околоцветника.
92.
Строение околоцветникаОколоцветник бывает простым (гомохламидным) и двойным (гетерохламидным). В случае отсутствия
околоцветника цветок называют ахламидным. Двойной околоцветник дифференцирован на чашечку
и венчик.
Чашечка в основном выполняет защитную функцию, состоит из чашелистиков, окрашенных обычно в
зеленый цвет. Они могут быть свободными или сросшимися (как у бобовых) и образуют спайную или
колокольчатую чашечку, на верхушке которой находятся зубцы. В некоторых группах чашечка
редуцируется (зонтичные) или видоизменяется в волоски, щетинки (сложноцветные).
Предполагается, что чашечка произошла от листьев верховой формации, о чем свидетельствуют
морфологическое и анатомическое сходство, одинаковое число проводящих пучков. Кроме того,
у некоторых растений, например пиона, наблюдается постепенный переход от листьев верховой
формации к чашелистикам.
Венчик имеет разнообразную окраску и по размерам обычно значительно превосходит чашечку. Он
состоит из лепестков. У некоторых растений (гвоздика, мыльнянка) верхняя (расширенная) часть
лепестка называется пластинкой, или отгибом, нижняя (суженная) – ноготком. Лепестки венчика могут
быть одинаковыми или различаться между собой. У многих растений венчик является
сростнолепестным. При этом различают трубку венчика и отгиб, который обычно заканчивается
зубцами, или лопастями. Место перехода трубки в отгиб называется зевом.
Предполагается, что венчик произошел от видоизмененных тычинок, о чем свидетельствуют яркая
окраска и сходство анатомического строения (один проводящий пучок). У представителей
семейства кувшинковых отмечается постепенный переход от тычинок к лепесткам.
Простой околоцветник состоит из одинаковых листочков и характерен для представителей
однодольных растений и некоторых двудольных (например ветреницы, купальницы). Он чаще
бывает венчиковидным (ярко окрашенным), иногда – чашечковидным (зеленым).
93.
94.
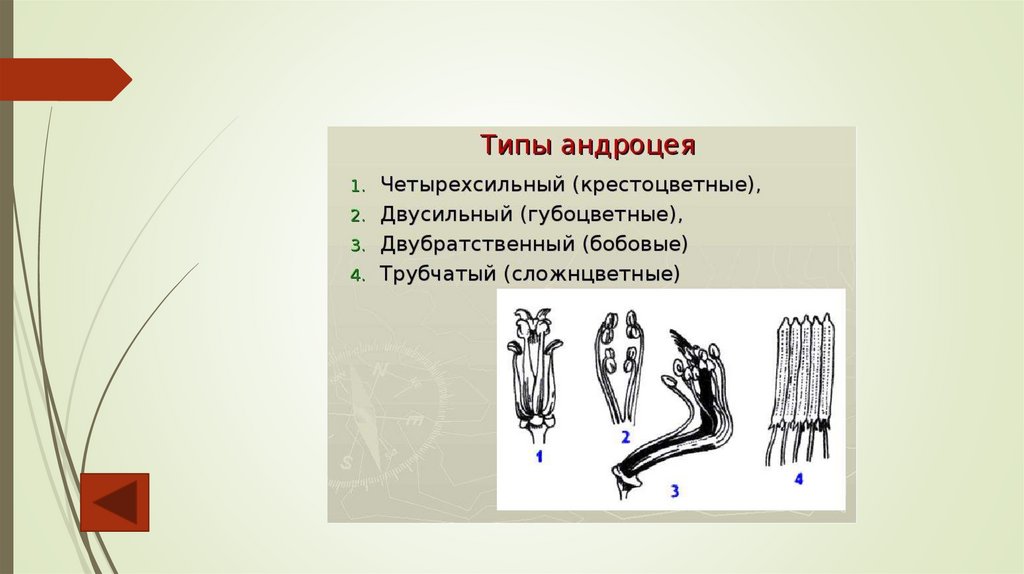
АндроцейАндроцей – это совокупность тычинок в цветке. Их количество варьирует от одного
(орхидные) до нескольких сотен. Тычинки могут быть свободными или способны
срастаться, могут иметь неодинаковую длину. Тычинка состоит из тычиночной нити и
пыльника. Прикрепление тычиночной нити к пыльнику также может быть
разнообразным. Пыльник имеет две половинки (теки), отделенные друг от друга
связником. Каждая тека содержит по два пыльцевых гнезда, являющихся гомологами
микроспорангиев.
На поперечном срезе пыльника можно увидеть следующие ткани. Снаружи
располагается эпидермис, за ним следует эндотеций, клетки которого часто имеют
разнообразные утолщения, способствующие вскрыванию пыльника. Под эндотецием
располагаются 1–3 средних слоя, состоящих из паренхимных клеток. Самый
внутренний слой стенки пыльника (тапетум) снабжает спорогенную ткань
питательными веществами, однако после образования микроспор его клетки, как
правило, разрушаются.
В пыльцевом гнезде находится спорогенная ткань, формирующая микроспоры
(пыльцевые зерна), причем при их образовании происходит процесс мейоза.
Пыльцевое зерно имеет две оболочки: наружную (экзину) и внутреннюю (интину). В
нем развивается мужской гаметофит, который очень редуцирован и состоит из двух
клеток: сифоногенной и генеративной. Генеративная заменяет антеридий и дает
начало двум спермиям.
Многие ученые считают, что тычинки произошли от микроспорофиллов голосеменных
растений (листоподобных структур, эволюционировавших параллельно с листом) в
результате значительной редукции последних. В процессе дальнейшей эволюции
цветка произошла дифференциация тычинок на тычиночную нить и пыльник.
95.
96.
Строение гинецеяГинецей – это совокупность плодолистиков, образующих один или несколько
пестиков.
Плодолистик состоит из стерильной и фертильной (семязачатков) частей. У
примитивных растений гинецей состоит из несросшихся плодолистиков. Такой тип
называют апокарпным. Плодолистик может быть плоским, а также с не полностью
или полностью сращенными краями.
По современным представлениям, плодолистик произошел от мегаспорофиллов
древних голосеменных растений. Вначале он был несомкнутым, в дальнейшем
произошло полное срастание краев плодолистика, а семязачатки, расположенные
внутри, получили бóльшую защиту.
В процессе эволюции плодолистики срослись и образовали ценокарпный гинецей
трех основных типов (рис. 14). В результате срастания замкнутых плодолистиков
своими боковыми поверхностями формируется синкарпный гинецей. Паракарпный
гинецей возникает в процессе размыкания отдельных плодолистиков и срастания их
между собой краями, а лизикарпный – в результате разрушения боковых стенок
синкарпного гинецея. Количество плодолистиков, образовавших ценокарпный
гинецей, можно определить по числу столбиков на одной завязи, количеству лопастей
рыльца, числу гнезд завязи.
Пестик дифференцируется на завязь, стилодий (оттянутая верхушка плодолистика) и
рыльце. Сросшиеся стилодии нескольких плодолистиков называют столбиком. В
зависимости от типа цветоложа завязь может занимать разное положение по
отношению к другим органам цветка. Верхняя завязь не срастается с
околоцветником, свободно располагается на цветоложе, а части околоцветника и
тычинки прикрепляются под ней. Цветок в этом случае называют подпестичным.
Нижняя завязь полностью срастается с вогнутым цветоложем, основанием частей
околоцветника и тычинок. Она находится как бы под околоцветником и тычинками.
97.
98.
Место прикрепления семязачатка к завязи называют плацентой, арасположение плацент в завязи – плацентацией. В зависимости от того, как
располагаются плаценты внутри завязи, различают следующие типы
плацентаций:
1) ламинальная, т. е. семязачатки прикреплены ко всей внутренней
поверхности завязи (магнолия, дегенерия). Является самым
примитивным типом плацентации;
2) сутуральная (краевая), т. е. семязачатки расположены вдоль шва.
• Краевая – расположение семязачатков в 2 ряда вдоль брюшного шва.
Такой тип плацентации характерен для апокарпного гинецея.
• Центрально-угловая – синкарпный гинецей.
• Париетальная (постенная) – паракарпный гинецей.
• Центрально-осевая (колончатая) – лизикарпный гинецей.
Внутри завязи располагаются семязачатки, число которых может
варьировать от одного до нескольких миллионов.
99.
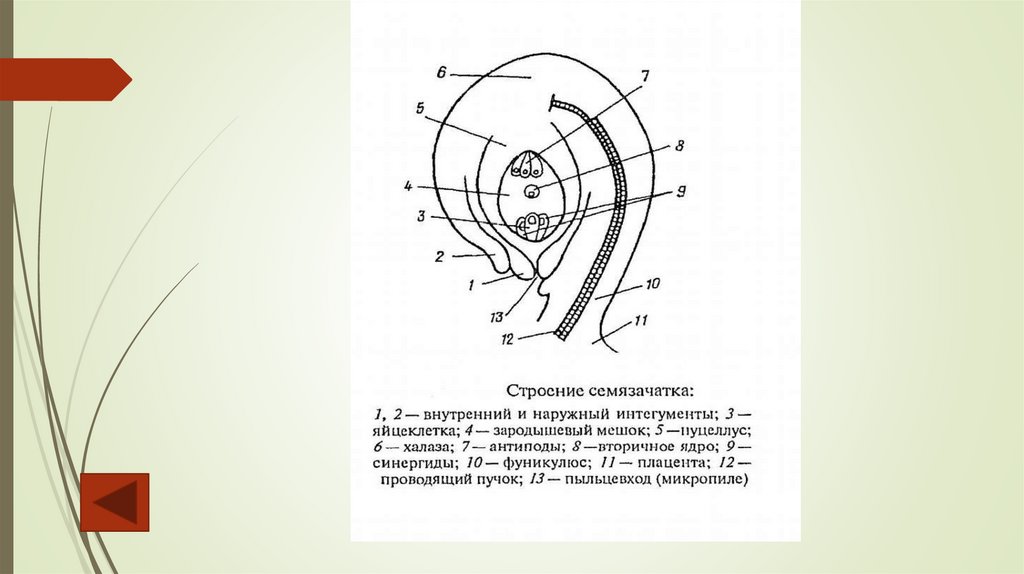
СемязачатокСемязачаток – многоклеточное образование семенных растений, из которого
развивается семя. Семязачаток состоит из нуцеллуса (мегаспорангия), окруженного
одним или двумя интегументами (покровами), которые на верхушке не смыкаются,
образуя узкий канал – микропиле. Через этот канал пыльцевая трубка проникает к
зародышевому мешку. Основание нуцеллуса, от которого отходят интегументы,
называется халазой. Семязачаток прикрепляется к плаценте с помощью фуникулюса
(семяножки). Место прикрепления семязачатка к семяножке называют рубчиком.
Семязачаток возникает на плаценте в виде небольшого бугорка. После достижения им
определенной величины у его основания возникают интегументы в виде одного-двух
кольцевых валиков, а в апексе нуцеллуса вычленяются одна или несколько
археспориальных клеток. Далее одна из археспориальных клеток (материнская клетка
мегаспор) делится путем мейоза и образует четыре гаплоидные мегаспоры. Этот
процесс носит название мегаспорогенеза. Дальнейшее развитие обычно получает
только одна (чаще нижняя) мегаспора, которая дает начало женскому гаметофиту
(зародышевому мешку), а три другие мегаспоры дегенерируют.
Выделено 16 типов зародышевых мешков. Наиболее распространен зародышевый
мешок Polygonum-типа, при формировании которого в мегаспоре трижды происходит
митоз. Образуется восемь свободных ядер; по три ядра отходят к разным полюсам
клетки, покрытые сначала лишь мембраной, а позднее и тонкой оболочкой. Три верхние
клетки называют яйцевым аппаратом. Средняя из них становится яйцеклеткой, две по
бокам – синергидами. Три клетки, расположенные на халазальном полюсе, – антиподы.
Два ядра, оставшихся в центральной части зародышевого мешка, называют
полярными. До или после проникновения пыльцевой трубки в зародышевый мешок они
сливаются во вторичное (диплоидное) ядро.
100.
101.
Опыление растенийОпылением называется перенос пыльцы на рыльце пестика.
Различают два основных типа опыления: автогамию, или самоопыление, при
котором опыление происходит в пределах одного цветка, и аллогамию
(перекрестное опыление) – перенос пыльцы с одного цветка на другой. Если
цветки находятся на одном растении, перекрестное опыление называют
гейтоногамией, если на разных – ксеногамией. Гейтоногамия генетически
эквивалентна автогамии, однако требует тех или иных агентов для переноса
пыльцы.
Перекрестное опыление встречается в природе гораздо чаще и обеспечивает
генетически более разнообразное потомство. Самоопыление является
резервной возможностью опыления в том случае, когда по каким-то причинам
(погодные условия, отсутствие опылителя и др.) не произошло перекрестное
опыление. В обоеполых цветках есть различные приспособления,
препятствующие самоопылению. Обычно таким приспособлением служит
дихогамия – разновременность созревания тычинок и пестика (более раннее
созревание пыльцы называют протерандрией, рыльца – протерогинией).
У некоторых видов дихогамия сопровождается гетеростилией
(разностолбчатостью): у одних цветков тычинки имеют короткие тычиночные
нити, а столбик пестика длинный, у других – наоборот. Кроме того,
приспособлениями к перекрестному опылению являются самостерильность
(неспособность пыльцы прорастать на рыльце пестика того же цветка),
раздельнополость цветков, двудомность растений.
102.
Агентами перекрестного опыления чаще всего выступают насекомые и ветер.Опыление с помощью насекомых называется энтомофилией. Растения привлекают
насекомых прежде всего с помощью пыльцы и нектара, непосредственно
используемых насекомыми в качестве пищи; поэтому их называют первичными
аттрактантами. У многих специализированных энтомофильных растений имеются
нектарники. У ряда растений они развиваются внутри длинных мешковидных выростов
(шпорцев), развивающихся из лепестков или листочков околоцветника.
Окраска, форма и размеры цветков, а также запах служат для насекомых
«указателями» на присутствие в цветках необходимых для них субстанций (нектара и
пыльцы), однако сами по себе они не используются насекомыми. Их называют
вторичными аттрактантами. Для цветков энтомофильных растений характерны
венчики, окрашенные в яркие цвета (красный, розовый, сиреневый, фиолетовый,
синий).
Анемофилия – это опыление растений, осуществляемое с помощью ветра. К
анемофильным растениям относятся голосеменные растения, многие
покрытосеменные (береза, тополь, ива, осоки, злаки). У видов, специализированных на
опылении ветром, цветки обычно мелкие, не имеют яркоокрашенного венчика;
околоцветник у них невзрачный, чешуевидный или пленчатый, иногда почти полностью
редуцирован. Пыльца легкая, мелкая, гладкая, образующаяся в больших количествах.
Рыльце пестика, часто разветвленное на тонкие лопасти, хорошо улавливает
разносимую ветром пыльцу. Цветки часто собраны в соцветия. Многие анемофильные
растения – двудомные (осина, крапива двудомная). Лесные анемофилы цветут весной,
до распускания листьев.
Кроме того, существуют еще другие способы опыления: орнитофилия (опыление с
помощью птиц), хироптерофилия (опыление летучими мышами) и гидрофилия.
Орнитофильные цветки обычно красные, имеют прочную структуру и выделяют
большое количество нектара. Цветки, опыляющиеся с помощью летучих мышей,
раскрываются ночью; они обычно белые и обладают сильным неприятным запахом. В
них, как правило, содержится большое количество пыльцы и нектара. Опыление с
помощью воды (гидрофилия) характерно для водных растений (роголистник, наяда,
элодея), у которых цветки погружены в воду или соцветия находятся над водой.
103.
Двойное оплодотворение и егозначение
Пыльцевое зерно, попав на рыльце пестика, начинает прорастать. При этом
его содержимое выпячивается через поры экзины и образует пыльцевую
трубку. Между опылением и оплодотворением проходит различное время.
Ядро клетки пыльцевой трубки и два спермия находятся на растущем конце
трубки. Достигнув завязи, пыльцевая трубка направляется к семязачатку и
проникает в него чаще всего через микропиле (порогамия), в редких случаях
– через халазу (халазогамия). Дальше трубка растет по направлению к
зародышевому мешку. Оболочка пыльцевой трубки растворяется – и
спермии оказываются внутри зародышевого мешка. Один из них сливается с
яйцеклеткой, другой – с диплоидной центральной клеткой. В результате
образуются диплоидная зигота и триплоидная клетка, которая дает начало
эндосперму. Происходит так называемое двойное оплодотворение –
характерный процесс покрытосеменных растений. Из оплодотворенного
семязачатка впоследствии формируется семя. Типичный способ
оплодотворения называется амфимиксисом.
104.
Типы отклонений от нормальногооплодотворения у цветковых растений
1. Апомиксис – формирование зародыша из неоплодотворенных элементов
зародышевого мешка или семязачатка. Различают следующие его типы:
а) партеногенез – развитие зародыша из неоплодотворенной яйцеклетки с
редукционным делением материнской клетки (1n) или без него (2n);
б) апогамия – развитие зародыша (1n или 2n) из антипод или синергид;
в) апоспория – утеря мегаспорогенеза и развитие зародышевого мешка из
вегетативной клетки нуцеллуса или интегумента с последующим врастанием
внутрь зародышевого мешка.
2. Полиэмбриония (многозародышевость):
а) истинная – развитие в одном зародышевом мешке нескольких зародышей из
одной зиготы в результате ее неправильного деления или расщепления ее
верхушечной клетки;
б) ложная– образование зародыша либо в результате развития в семязачатке не
одной, а нескольких мегаспор (лилия, манжетка), либо в результате развития
дополнительных зародышевых мешков из клеток нуцеллуса или интегумента.
105.
Формула и диаграмма цветкаДля краткого условного выражения строения цветков применяют формулы. При
их составлении учитывают симметрию цветка, число кругов в нем, а также число
членов в каждом круге, срастание частей цветка и положение пестиков (верхняя
или нижняя завязь).
Формула цветка составляется следующим образом. Актиноморфный
(правильный) цветок обозначается звездочкой (∗), зигоморфный – стрелкой (↑ или
↓), асимметричный –
. Простой околоцветник обозначается буквой P, чашечка
– K (Ca), венчик – C (Co), андроцей – A, гинецей – G. У каждой буквы внизу
ставится индекс, указывающий на число членов данной части цветка (например
С5, А5) . Если частей членов много, неопределенное число, то ставят знак
бесконечности ∞ . Если данные части цветка располагаются не в одном, а в двух
кругах, то у буквы ставят две цифры, соединенные знаком «+» (например P3+3).
При срастании каких-либо частей цветка цифра, указывающая на их число,
заключается в скобки (K(5)). Верхняя завязь отмечается чертой под цифрой,
обозначающей число плодолистиков, которые образуют пестик (G(3) ), нижняя
завязь – чертой над цифрой (G(3) ); средняя (полунижняя) – сбоку.
Диаграмма представляет собой схематическую проекцию цветка на плоскость,
перпендикулярную оси цветка. При изображении диаграммы цветка чашелистики
указываются в виде скобки с килем на спинке, лепестки – круглой скобкой. Для
тычинок дается поперечный срез через пыльник (при большом числе тычинок
возможно упрощенное изображение в виде овала), для гинецея – поперечный
разрез завязи с плацентацией семязачатков. Диаграмма отражает также
срастание органов, наличие нектарников, дисков и т.д.
106.
Морфологическая характеристикасоцветий
Соцветие – это система видоизмененных побегов, несущих цветки. Образование соцветий является высокой
специализацией к опылению. Как при энтомофилии, так и при анемофилии вероятность опыления цветков в
соцветиях возрастает. К тому же цветки в соцветиях распускаются не одновременно, а последовательно, что
удлиняет период возможного опыления. В соцветиях повышается гарантия сохранения завязи при повреждении
насекомыми и устойчивость к неблагоприятным факторам.
Для описания и морфологической характеристики соцветий используют четыре группы признаков: 1) характер
олиственности; 2) порядок ветвления побегов; 3) способ их нарастания; 4) деятельность апикальной меристемы.
1. По наличию и характеру листьев на осях соцветия делят на фрондозные, брактеозные и эбрактеозные.
На осях фрондозных соцветий имеются хорошо развитые зеленые прицветники (фуксия, фиалка
трехцветная). Брактеозными называют соцветия, прицветники которых представлены чешуевидными
листьями верховой формации – брактеями (ландыш, сирень). У эбрактеозных (голых) соцветий прицветники
вообще редуцированы (пастушья сумка).
2. В зависимости от степени разветвленности выделяют простые и сложные соцветия. У простых на главной
оси располагаются одиночные цветки, а у сложных – частные соцветия, т. е. ветвление достигает двух и
более порядков.
3. Способ ветвления осей может быть моноподиальным и симподиальным. В случае моноподиального
ветвления каждая ось формируется за счет деятельности одной верхушечной меристемы. Соцветия с
такими осями называют моноподиальными, или рацемозными. Если оси ветвятся симподиально и являются
составными, соцветия относят к симподиальным, или цимозным.
4. В зависимости от особенностей функционирования апикальной меристемы выделяют открытые и
закрытые соцветия. В открытых соцветиях (бокоцветных, неопределенных) апикальная меристема не
формирует цветки и обладает неограниченным ростом. Цветки возникают на боковых осях и распускаются
снизу вверх (ландыш, черемуха). В закрытых соцветиях (верхоцветных, определенных) апикальная
меристема расходуется на образование верхушечного цветка (чистотел, барбарис), а цветки
распускаются сверху вниз.
107.
Классификация соцветийВ основу общей морфологической классификации соцветий положены два признака: способ
ветвления осей и степень их разветвленности. Соответственно выделяют рацемозные,
цимозные, составные соцветия и тирсы.
Рацемозные соцветия могут быть простыми (цветки сидят непосредственно на главной оси
соцветия) и сложными (цветки сидят на разветвлениях главной оси соцветий). К простым
моноподиальным соцветиям относятся следующие: кисть, цветки которой расположены на
удлиненной оси, имеют цветоножки (черемуха);
колос, сходный с кистью, но цветки сидячие (подорожник);
початок – колос с толстой, мясистой осью (кукуруза);
головка, сходная с кистью, но главная ось сильно укорочена, расширена, цветки сидячие
или на коротких цветоножках (клевер);
щиток, у которого на укороченной оси развиваются нижние цветки на более длинных
цветоножках, а верхние – на более коротких, благодаря чему все цветки расположены
почти в одной плоскости (спирея); зонтик, у которого главная ось сильно укорочена,
цветоножки выходят из ее верхушки и имеют одинаковую длину (проломник);
корзинка, сидячие цветки которой расположены на очень уплощенном и расширенном
конце укороченной главной оси (сложноцветные).
108.
109.
К сложным моноподиальным соцветиям относятся такие:сложный колос, на главной оси которого сидят элементарные
колоски (пшеница);
двойная кисть, на главной оси которой сидят пазушные простые
кисти (донник);
метелка, отличающаяся от двойной кисти более обильным
ветвлением, а также тем, что в нижней части она более ветвистая,
чем в верхней (сирень);
сложный зонтик, главная ось которого укорочена, от нее отходит
несколько осей, несущих на конце простые зонтики (укроп,
борщевик).
110.
111.
Цимозные соцветия характеризуются симподиальным ветвлением, относятся к сложным закрытым ипредставлены следующими типами:
• монохазий (ось первого порядка заканчивается цветком). Под цветком закладывается одна ось
второго порядка, которая перерастает ось первого порядка и также заканчивается цветком,
ниже закладывается следующая ось. В зависимости от порядка заложения боковых осей
различают завиток и извилину. В завитке все цветки направлены в одну сторону (незабудка). В
извилине боковые оси с цветком отходят поочередно в противоположные стороны (гладиолус);
• дихазий (ось первого порядка несет на верхушке цветок, под ним образуются, обычно
супротивно, две боковые оси, также заканчивающиеся цветками). Под цветком на боковых осях
могут закладываться по две оси третьего порядка и т.д. (герань, гвоздика);
• плейохазий (из каждой оси, несущей верхушечный цветок, выходит более двух ветвей,
перерастающих главную ось).
К сложным относятся также составные (агрегатные) соцветия и тирсы. У составных характер
ветвления главной и боковых осей различен. Например, если главная ось ветвится по типу кисти, а
боковые частные соцветия представлены корзинками, то такое соцветие называется кистью
корзинок. Наиболее распространенными типами составных соцветий являются такие: щиток
корзинок, кисть корзинок, кисть зонтиков, метелка колосков и т.д.
Тирс – это соцветие, имеющее моноподиально ветвящуюся главную ось, несущую боковые цимозные
соцветия (монохазии и дихазии). К тирсам относятся сережки березы, имеющие гибкую главную ось.
112.
113.
Строение семениСемя развивается из семязачатка после оплодотворения (в случае
апомиксиса – без оплодотворения). Снаружи оно покрыто семенной
кожурой, образованной из интегумента и выполняющей защитную
функцию. Эндосперм, возникший из триплоидного ядра, содержит
запасные вещества, питающие зародыш при прорастании. У некоторых
растений запасающую функцию может выполнять перисперм,
образовавшийся из нуцеллуса. Из оплодотворенной яйцеклетки
развивается зародыш.
Семена многих растений имеют придатки в виде сочных, мясистых, часто
окрашенных выростов, богатых питательными веществами. Если эти
выросты развиваются из семяножки (пассифлора, бересклет, мускатный
орех), их называют присемянниками, или ариллусами, если из
интегументов (хохлатка, клещевина) – карункулами, или ариллоидами.
Они служат для привлечения животных, участвующих в распространении
семян.
114.
Семенная кожура (спермодерма) выполняет защитную функцию. На кожуре естьнебольшое отверстие – микропиле, способствующее проникновению первых порций воды в
начале набухания, и рубчик – место прикрепления семени к семяножке. Степень развития,
твердость семенной кожуры определяются характером околоплодника: при
невскрывающихся твердых околоплодниках она тонкая (вишня, дуб, сложноцветные); в
противоположных случаях кожура твердая (виноград, калина). У граната – сочная семенная
кожура.
Эндосперм возникает из триплоидного ядра, которое после оплодотворения начинает
делиться первым. По характеру развития различают три основных типа эндосперма:
• ядерный (нуклеарный), т. е. сначала образуется большое количество ядер, затем вокруг
них формируются оболочки;
• клеточный (целлюлярный), т. е. каждое деление ядра сопровождается цитокинезом;
• гелобиальный (промежуточный), т. е. после первого деления зародышевый мешок делится
на две части: микропилярную (большую) и халазальную (маленькую). В них происходит
свободное деление ядер, а впоследствии возникают клеточные стенки.
В эндосперме запасаются крахмал, масла, белки. В покоящемся семени эндосперм
твердый. При прорастании вещества эндосперма гидролизируются под действием
ферментов и поглощаются зародышем; 85% покрытосеменных растений имеют эндосперм
(магнолиевые, лилейные, пальмы), 15% – не имеют (бобовые).
115.
Перисперм – запасающая ткань, характерная для некоторых растений (перец, кувшинка,звездчатка, свекла) и возникающая из нуцеллуса (2n).
По наличию запасающих тканей выделяют следующие типы семян:
• с эндоспермом (клещевина, злаки, пасленовые);
• с эндоспермом и периспермом (перец, кувшинка);
• с периспермом и без эндосперма (звездчатка, куколь, свекла);
• без эндосперма и перисперма (бобовые, орхидные).
Зародыш возникает из оплодотворенной яйцеклетки и состоит из меристематических
тканей. Гетеротрофен. Часто расчленен. Зародыш представлен осью и семядольными
листьями (два – у двудольных, один – у однодольных). Семядоли выполняют
выделительную, запасающую, всасывающую функции.
На оси зародыша у некоторых растений формируется почечка с зачатками настоящих
листьев. С другой стороны расположен корешок с корневым чехликом. Часть оси, к
которой прикрепляются семядоли, называют семядольным узлом. Ниже семядолей на оси
располагается гипокотиль (подсемядольное колено), выше – эпикотиль.
116.
В семени злаков эндосперм занимает значительный объем, т. к. в немоткладываются запасные вещества. Он дифференцирован на два слоя.
Наружный – алейроновый слой, в котором откладываются белки. Он
расположен сразу под семенной кожурой. Ближе к центру находятся
клетки с крахмальными зернами. Зародыш злаков состоит из одной
семядоли, зародышевого корешка, зародышевого стебелька и почечки.
Единственная семядоля (щиток) прилежит к хорошо развитому
эндосперму. В центре почечки хорошо заметен конус нарастания стебля,
прикрытый примордиями листьев. Наружный колпачковидный лист,
окружающий почечку, называется колеоптилем. Зародышевый корешок
окружен специальным многослойным чехлом (колеоризой), которая при
прорастании набухает и развивает на поверхности всасывающие
волоски. Иногда на стороне, противоположной щитку, образуется
чешуевидный вырост – эпибласт. Он расценивается некоторыми
учеными как остаток второй семядоли.
117.
118.
Прорастание семянДля прорастания семян необходимы определенные условия:
• наличие воды (ткани семян обезвожены и содержат 10–15% влаги);
• доступ воздуха (обеспечивает процессы дыхания);
• оптимальная температура (+25–35°);
• для некоторых семян – свет (табак, салат).
Однако не всегда семена прорастают при наличии всех благоприятных условий. Часто наблюдается глубокий
покой семян, причины которого могут быть различны:
• недоразвитый зародыш (на дозревание семени женьшеня требуется три года), обуславливающий так
называемый морфологический покой семян;
• водонепроницаемые (бобовые) или твердые (косточковые) покровы, механически препятствующие
прорастанию;
• наличие веществ-ингибиторов, химически тормозящих прорастание (ясень);
• пониженная газопроницаемость внутренних слоев покровов (злаки, табак).
Последние три причины обуславливают органический покой семян. Покой семян характерен для многих семян
умеренной зоны и является приспособлением к перенесению неблагоприятных условий (зима). Затем покровы
разрушаются, перегнивают, ингибиторы прорастания вымываются, зародыш дозревает и семя прорастает.
Для ускорения процесса проращивания семян используют ряд специальных приемов. С помощью
стратификации – выдерживания семян при низких положительных температурах – ускоряют прорастание
плодовых, лесных деревьев. При помощи скарификации – перетирания, пропускания через металлические щетки,
действия кислот – разрушают покровы «твердокаменных» семян (косточковые, клевер, шиповник).
119.
По скорости прорастания и сохранению всхожести можно выделить следующие типы семян:• с длительным, глубоким покоем и длительно сохраняемой всхожестью (многие древесные
растения, травы);
• прорастающие сразу или через некоторое время после опадения и сохраняющие всхожесть в
течение нескольких лет (культурные злаки, овощные культуры);
• прорастающие сразу и быстро теряющие всхожесть (ива, тополь);
• прорастающие на материнском растении (авиценния).
Процессу прорастания предшествует набухание, связанное с поглощением большого
количества воды и обводнением тканей семени. При этом кожура обычно разрывается.
Начинается активная ферментативная деятельность, связанная с мобилизацией запасных
веществ, которые начинают поглощаться зародышем. Зародыш увеличивается в размерах. Из
разрыва кожуры или микропиле выходит корешок, укрепляющий растение в почве, и начинает
самостоятельно всасывать воду (гипокотиль подталкивает корешок в почву). Далее возможны
два типа прорастания семян:
• эпигенное (надземное) наблюдается у растений с небольшими семядолями (однодольные,
некоторые двудольные); при этом сильно удлиняется гипокотиль, выносящий семядольные листья
на поверхность. Вслед за семядольными появляются первые настоящие листья;
• гипогенное (подземное) встречается у растений с массивными семядолями, выполняющими
запасающую функцию (бобовые). При этом сильно удлиняется эпикотиль, семядоли остаются в
земле, а на поверхность выносятся только первые настоящие листья. Такие растения обычно
имеют хорошо сформированную почечку.
120.
121.
Строение и классификация плодовПлод – характерный орган покрытосеменных растений. Его функция
заключается в защите и распространении семян. Образуется обычно из
стенок завязи. Иногда в образовании плода могут принимать участие
цветоложе, цветоножка, части околоцветника.
Разнообразие плодов определяется четырьмя группами признаков:
• строение околоплодника;
• способ вскрывания;
• строение гинецея;
• особенности, связанные с распространением;
122.
Околоплодник (перикарпий) представляет собой разросшуюся стенку завязи. Иногда в его образованиипринимают участие и другие части цветка. У некоторых растений (вишня, слива) перикарпий четко
дифференцирован на три зоны: экзокарпий (кожица), мезокарпий (съедобная мясистая часть) и эндокарпий
(косточка, окружающая семя).
В зависимости от консистенции околоплодника плоды делят на сухие (боб, коробочка, зерновка) и сочные
(костянка, ягода).
По типу вскрывания плодов различают:
невскрывающиеся плоды (околоплодник разрушается под влиянием механического воздействия,
деятельности микроорганизмов);
вскрывающиеся (сухие, многосемянные плоды). Вскрывание может происходить:
• продольными щелями (по брюшному шву или спинной жилке);
• разрывом по перегородкам (септицидные плоды);
• разрывом по гнездам (локулоцидные плоды).
Плоды могут вскрываться створками полностью или не полностью: зубчиками (гвоздичные), дырочками (мак),
крышечкой (белена).
Распадающиеся плоды эволюционно более продвинуты. Выделяют две их группы:
а) дробные распадаются продольно в плоскости срастания плодолистиков. При этом образуются
замкнутые односемянные мерикарпии (зонтичные, губоцветные, бурачниковые)
б) членистые распадаются поперечно в плоскостях, перпендикулярных продольной оси плодолистика. При
этом образуются замкнутые членики с поперечными ложными перегородками (растения районов с
засушливым климатом).
123.
Строение гинецея. Существует классификация, основанная на взаимосвязиплодолистиков друг с другом и другими частями цветка. При этом различают
простые, сборные плоды и соплодия. Простые плоды развиваются из одного пестика,
представленного одним или несколькими сросшимися плодолистиками (фасоль,
тюльпан). Сборные образуются из нескольких свободных плодолистиков, каждый из
которых формирует пестик (малина, земляника, ветреница). Соплодие развивается
из целого соцветия, опадающего целиком (ананас, инжир).
По типу гинецея различают апо–, син–, пара– и лизикарпные плоды.
Хотя морфология плода является устойчивым для вида признаком, у некоторых видов
на одном растении могут развиваться плоды, различающиеся по строению. Это
явление называется гетерокарпией.
У многих растений также хорошо выражена гетероспермия – разносемянность.
Например, у мари белой формируются семена трех типов: крупные светлокоричневые, более мелкие черные и самые мелкие – блестящие черные. Эти семена
прорастают не одновременно, а группами. Если по каким-то причинам не
прорастают крупные семена, прорастают средние, а мелкие остаются в покое.
Гетерокарпия и гетероспермия встречаются только в высокоразвитых семействах и
часто характерны для сорных растений, отличающихся высокой экологической
пластичностью и жизнеспособностью.
124.
125.
Распространение плодов и семянЕдиницами распространения могут служить разные части растений,
называемые диаспорами. Карпологические диаспоры представляют
собой семена, высыпавшиеся из вскрывшегося плода, целые
невскрывающиеся сухие и сочные плоды, части плодов – мерикарпии,
отдельные членики.
Единой классификации способов распространения семян и плодов нет;
обычно указывают главный фактор, обеспечивающий этот процесс.
1. Автохория – наиболее простой способ распространения,
осуществляемый без посредников. При этом происходит либо активное
разбрасывание семян при вскрывании с помощью особых структур
(недотрога, бешеный огурец), либо самопроизвольное опадение под
действием собственного веса (барохория). Барохория характерна для
плодов пальм, каштана, дуба.
2. Баллистохория – разбрасывание семян с помощью раскачивающихся
вегетативных органов (гвоздичные, колокольчиковые).
126.
3. Анемохория – распространение с помощью ветра. Анемохорные виды характеризуются высокойсеменной продуктивностью.
• Мелкие семена орхидей, грушанок распространяются слабыми конвекционными потоками.
• Специальные приспособления – летучки, волоски, крылатки – облегчают парение семян и плодов в
воздухе. Иногда такую функцию выполняет сохраняющийся при плоде прицветный лист (липа).
Семена-летучки характерны для ивы, тополя, кипрея. Плоды-летучки образуются у одуванчика, осота,
прострела. Плоды-крылатки имеются у таких растений, как береза, ольха, хмель, щавель. Иногда
летательные приспособления помогают не только передвижению плодов с помощью ветра, но и
зарыванию их в землю, как это наблюдается у ковыля, аистника.
• Особый случай – растения «перекати-поле», распространенные в степях или пустынях. При
созревании семян отламываются от подземной части и в виде шариков или комков перегоняются
ветром на большие пространства, постепенно рассеивая семена (верблюжья колючка, клоповник,
качим).
4. Гидрохория – распространение с помощью воды. Семена защищены от смачивания и, как правило,
обладают плавучестью. У многих водных и болотных растений (кувшинка, частуха, осоки) плоды
имеют специальные воздухоносные приспособления, позволяющие им держаться на поверхности
воды некоторое время и переноситься с помощью водных течений и ветра. Уникальный пример –
плоды тропической энтады, которые разносятся морскими течениями и даже обнаружены в
северных морях.
У некоторых растений (вероника, очиток едкий) плоды приспособлены к вымыванию из них семян
дождевой водой.
127.
5. Зоохория – распространение с помощью животных (птиц, млекопитающих, насекомых и др. групп).Она осуществляется тремя способами.
• Эндозоохория, т. е. животные поедают диаспоры, не переваривая семена, которые проходят через
пищеварительный тракт и выводятся наружу.
Обычно это сочные плоды, семена которых защищены от переваривания либо каменистым
эндокарпием (косточковые), либо твердой семенной кожурой.
• Синзоохория, т. е. растаскивание диаспор и отложение их про запас. Так распространяются
ореховидные диаспоры, которые животные уносят в гнезда или в специальные «кладовые» для запаса
кормов. Агентами синзоохории являются птицы (кедровки, сойки, дятлы) и грызуны (белки, бурундуки,
мыши).
Прогрессивной формой синзоохории является мирмекохория. Муравьи распространяют семена
растений с мясистыми придатками – элайосомами (разновидность ариллусов – фиалки, ожика,
чистотел). Элайосомы богаты маслами и другими питательными веществами. Выделяют летучие
вещества, привлекающие насекомых.
• Эпизоохория – случайный перенос диаспор, снабженных различными прицепками, крючками,
цепляющимися за шерсть животных (репейничек, лопух, липучка).
6. Антропохория – распространение диаспор с помощью человека. Так, благодаря
трансконтитентальным первозкам в Европу были завезены бодяк, элодея, овсюг, а в Америку –
подорожник. С каждым годом растет количество заносных сорных растений, составляющих мощную
конкуренцию культурным.