




































































































 biology
biologySimilar presentations:



")






Введение в генетику. Взаимодействие генов (лекция № 4)
1.
Лекция № 4Введение в генетику.
Взаимодействие генов.
Профессор
Смирнов Сергей Николаевич
1
2.
План1. Предмет, задачи, методы, этапы развития, основные
термины генетики.
2. Принцип гибридологического анализа. Законы Менделя, их
цитологические основы. Закон «чистоты гамет». Анализирующее
скрещивание. Летальные гены.
3. Доминантный и рецессивный типы наследования признаков
человека. Промежуточный характер наследования.
4. Взаимодействие аллельных и неаллельных генов.
Полигенное наследование признаков.
5. Первичная и вторичная плейотропия.
6. Множественные аллели.
7. Наследование групп крови человека по системам АВ0 и MN.
Наследование резус-фактора. Резус-конфликт.
8. Иммуногенетика: предмет и задачи. Тканевая и видовая
специфичность белков. Антигенные свойства белков.
2
3.
1.Предмет, задачи, методы, этапы развития,основные термины генетики.
Генетика - это наука о наследственности и
изменчивости.
Наследственность - свойство живых организмов
передавать следующим поколениям особенности
морфофункциональной организации и
индивидуального развития.
Изменчивость - свойство живых организмов,
заключающееся в изменении генов и их проявлений в
процессе развития организмов.
3
4.
Перед генетикой стоят следующие задачи:Теоретические:
а. Изучение проблемы хранения наследственной
информации,
б. Изучение проблемы передачи наследственной
информации,
в. Изучение проблемы реализации наследственной
информации,
г. Изучение проблемы изменения наследственной
информации.
Практические (для медицины):
а. Управление развитием наследственных признаков,
б. Защита наследственности человека от вредных
воздействий окружающей среды.
4
5.
Методы генетики:а. Гибридологический,
б. Цитогенетические,
в. Биохимические,
г. Генеалогический,
д. Близнецовый,
е. Популяционно-статистический,
ж. Иммунологические.
5
6.
Этапы развитиягенетики.
Г. Мендель опубликовал
свои работы в 1865 году.
Однако, они не нашли
понимания и не получили
должного отклика научной
общественности. Поэтому
генетика как наука берет
свое начало в 1900 году,
когда законы Г.Менделя
были вторично открыты
другими учеными.
6
7.
Грегор Иоганн Мендель (GregorJohann Mendel; 1822-1884) —
австрийский биолог и ботаник,
сыгравший огромную роль в
развитии представления о
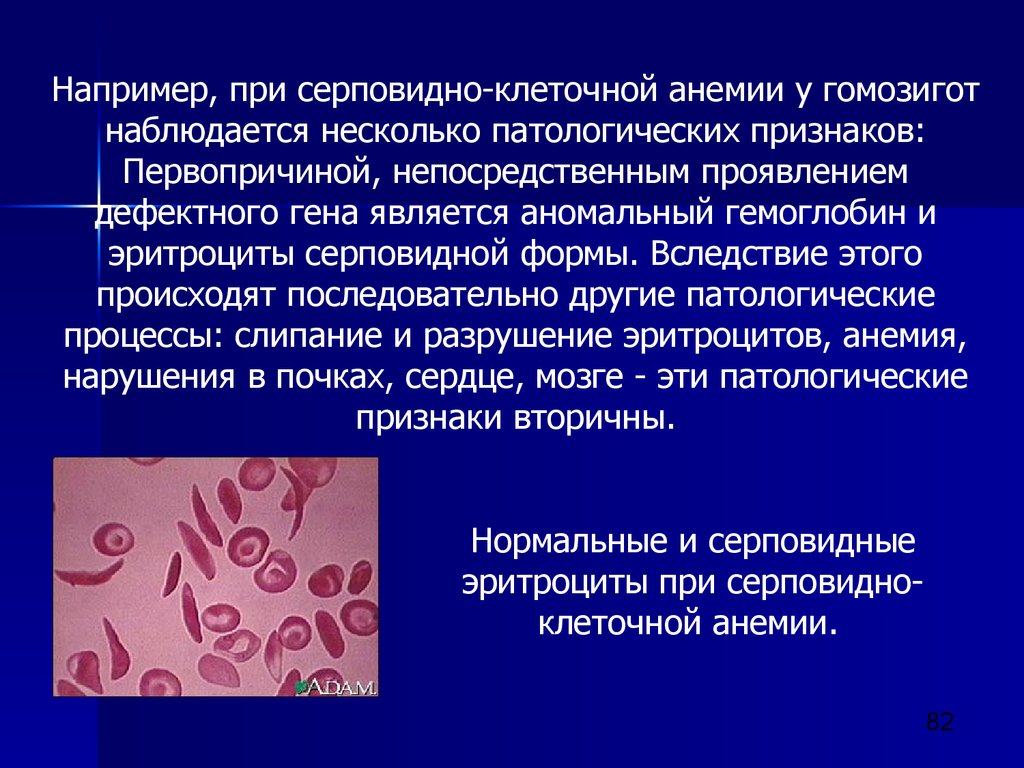
наследственности. Открытие им
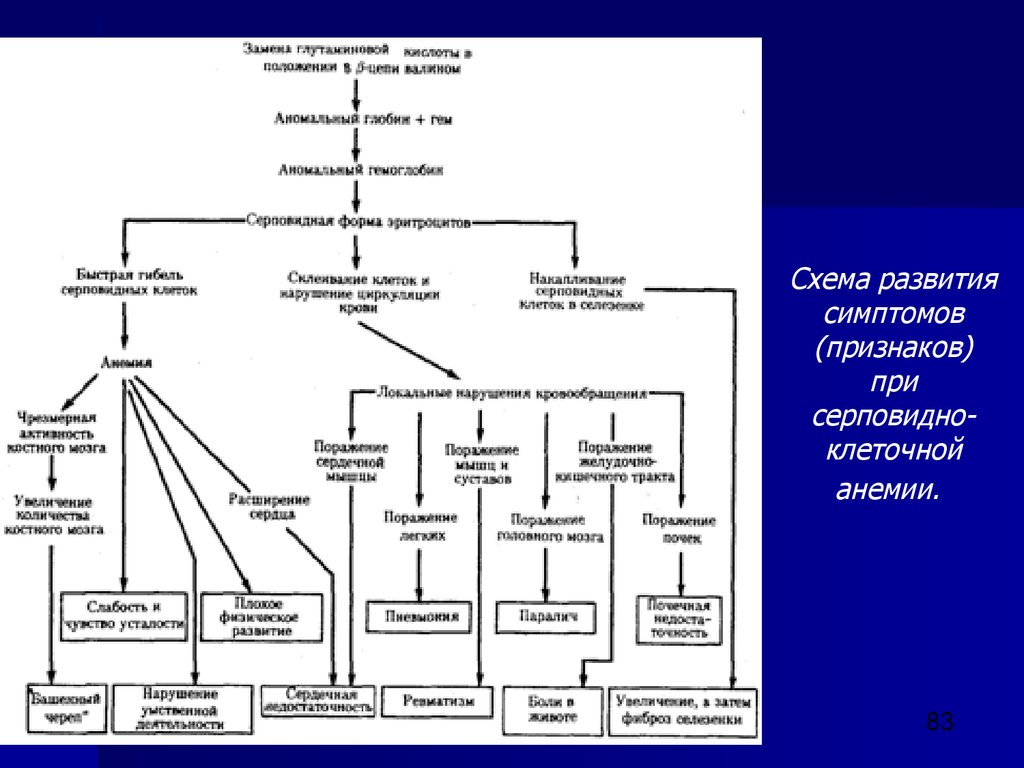
закономерностей наследования
моногенных признаков (эти
закономерности известны теперь
как Законы Менделя) стало
первым шагом на пути к
современной генетике.
Грегор Иоганн Мендель.
Портрет 1884 года.
7
8.
Выделяют пять этапов развития генетики:1 - 1900-1912;
2 - 1912-1925;
3 - 1925-1940;
4 - 1940-1955;
5 - 1955 - настоящее время.
Первый этап (1900-1912) - триумф менделизма. В
1900 году независимо друг от друга Х. Де Фриз, Э.
Чермак и К. Э. Корренс повторно открыли законы
Г.Менделя.
8
9.
Де Фриз Хуго (1848-1935)- нидерландский ботаник,
один из основателей
учения об изменчивости и
эволюции, Провел первые
систематические
исследования
мутационного процесса.
Разработал концепцию
эволюции посредством
мутаций (мутационная
теория Де Фриза).
Одновременно с К.
Э. Корренсом и Э.
Чермаком-Зейзенеггом
вторично открыл законы
Менделя (1900).
9
10.
Эрих Чермак-Зейзенегг Э.(1871-1962) - австрийский
ботаник. Проводил
исследования по
гибридизации рас садового
гороха. Независимо и
одновременно с Х. де
Фризом и К. Корренсом в
1900 г. ученый вторично
описал закономерности
наследственности,
сформулированные за 35 лет
до этого Г. Менделем. Э.
Чермак сосредоточил свое
внимание на практическом
применении генетических
закономерностей в селекции
10
культурных растений.
11.
Карл Эрих Корренс(1864-1933) –
германский ученый.
Независимо от Х. де
Фриза и Э. Чермака
вторично открыл
законы Г. Менделя
(1900 г.). К. Корренсу
принадлежат труды по
генетике пола, им была
впервые описана
цитоплазматическая
наследственность на
примере передачи
признака мозаичности
окраски листьев через
гены, заключенные в
11
хлоропластах.
12.
В 1907 году У. Бетсонпредложил
термин
"Генетика".
Бэтсон, Бейтсон (Bateson) Уильям (1861-1926), английский биолог,
один из основателей генетики, само название которой было
предложено им в 1907. Первые работы посвящены филогении
хордовых. Отстаивал ненаследование приобретённых признаков,
прерывистый характер изменчивости, учение о чистоте гамет. В 1910
12
основал английский генетической журнал «Journal of Genetics».
13.
Второй этап (1912-1925) - создания хромосомнойтеории наследственности.
Эта теория разработана школой Т.Моргана (США).
Томас Морган и его ученики (Г. Дж. Меллер, А. Г.
Стертевант и др.) обосновали хромосомную теорию
наследственности.
Установленные закономерности расположения генов
в хромосомах, способствовали выяснению
цитологических механизмов законов Грегора Менделя и
разработке генетических основ теории естественного
отбора.
13
14.
Томас Хант Морган(Thomas Hunt Morgan, 1866
—1945) — американский
биолог, один из
основоположников
генетики, Лауреат
Нобелевской премии по
физиологии и медицине
1933 года «За открытия,
связанные с ролью
хромосом в
наследственности».
14
15.
Третий этап (1925-1940) - этап открытиявозможности искусственного получения
мутаций.
В 1925 году этот феномен описали Г.А.
Надсон и Г.С. Филиппов.
В 1927 году Мюллер показал, что
мутагенным действием обладает
ионизирующее излучение.
15
16.
Георгий АдамовичНадсон (1867-1940),
русский микробиолог,
доказавший возможность
искусственной мутации.
Первая
экспериментальная работа
Г.А.Надсона, выполненная
совместно с
Г.С.Филипповым,
посвящена получению
мутаций под влиянием
рентгеновских лучей у
низших грибов из
семейства Mucoraceae
(1925 г.).
16
17.
Герман ДжозефМёллер (Hermann Joseph
"H. J." Muller 1890-1967),
американский генетик,
ученик Томаса Ханта
Моргана, лауреат
Нобелевской премии по
физиологии и медицине
(1946).
Наиболее известен
своими работами в области
мутагенного действия
рентгеновских лучей на мух
дрозофил.
17
18.
В этот период складываются иработают школы Н.И.Вавилова,
Н.К.Кольцова.
В 1926 году Николай
Иванович Вавилов открыл закон
гомологических рядов в
наследственной изменчивости.
Николай Иванович Вавилов российский генетик,
растениевод, географ. Автор закона гомологических рядов
в наследственной изменчивости организмов, учения о
биологических основах селекции и центрах происхождения
18
и разнообразия культурных растений.
19.
В 1928 году Н. К. Кольцовпервым
разработал
гипотезу
молекулярного
строения
и
матричной
репродукции
хромосом
(«наследственные
молекулы»),
предвосхитившую
принципиальные положения
современной молекулярной
биологии и генетики.
Кольцов Николай Константинович (1872-1940), российский
биолог, основоположник российской экспериментальной
биологии. Организатор и первый директор (1917-1939)
19
Института экспериментальной биологии.
20.
В тридцатые годы прошлого века в России развиваетсягенетика человека и медицинская генетика.
В Москве в начале 30-х годов прошлого века
существовали
медико-евгенические
консультации,
организованные
С.Н.
Давиденковым
и
Медикогенетический институт во главе с С.Г.Левитом.
20
21.
Сергей НиколаевичДавиденков (1880—1961),
невропатолог, академик АМН
СССР (1945). Основоположник
отечественной клинической
нейрогенетики.
21
22.
Соломон ГригорьевичЛевит (1894-1943), советский
генетик, один из
основоположников медицинской
генетики в СССР. В 1930-1937 –
директор Медико-генетического
института им. М. Горького. Был
организатором клиникогенетических исследований
близнецов.
22
23.
Четвертый этап (1940-1955) - этап исследованиймолекулярных основ генетики.
Д.Бидл (Beadle G.W.) и Э.Тетум (Tatum E.L.) (США)
показали, что один ген отвечает за синтез одного
фермента.
23
24.
Бидл Джордж Уэлс (BeadleGeorge Wells 1903-1989),
американский генетик.
Основополагающие труды по
биохимической генетике. Один
из авторов фундаментальной
концепции: "один ген - один
фермент" (каждый ген отвечает
за синтез только одного
фермента).
24
25.
Эдвард Лаури Тетум (EdwardLawrie Tatum 1909–1975),
американский генетик.
Нобелевская премия по
медицине (1958) совместно с
Д.У.Бидлом и Дж. Ледербергом
(Joshua Lederberg) за исследования
по генетике микроорганизмов.
25
26.
В 1944 году О. Эвери, С. Маклеод и М. Маккартнидоказали генетическую роль ДНК. Они показали, что
ДНК является носителем наследственных задатков.
С 1950 года Фрэнсис Крик и Джеймс Уотсон изучают
строение ДНК и в 1953 году открывают структуру ДНК.
26
27.
Крик (Crick) Фрэнсис Харри Комптон (1916-2004),английский биофизик и генетик. В 1953 совместно с Дж.
Уотсоном создал модель структуры ДНК (двойную
спираль), что позволило объяснить многие ее свойства и
биологические функции и положило начало молекулярной
генетике. Нобелевская премия (1962, совместно с Дж.27
Уотсоном и М. Уилкинсом).
28.
Джеймс Дьююи Уою тсон(James Dewey Watson, род. 6
апреля 1928), американский
биолог. Лауреат Нобелевской
премии по физиологии и
медицине (1962). — совместно
с Фрэнсисом Криком и Морисом
Х. Ф. Уилкинсом за открытие
структуры молекулы ДНК.
С 1955 года генетика вступает в современный этап
развития.
28
29.
Основные понятия генетики.Фенотип - совокупность всех признаков и свойств
организма, проявление которых обусловлено
взаимодействием генотипа с факторами внутренней и
внешней среды.
Генотип - совокупность всех генов организма.
Ген - участок молекулы ДНК, несущий информацию об
одном белке.
29
30.
Аллели (аллеломорфы) - различные формы одногои того же гена, расположенные в одинаковых участках
(локусах) гомологичных хромосом и определяющие
альтернативные варианты развития одного и того же
признака.
В диплоидном организме может быть два
одинаковых аллеля одного гена, в этом случае организм
называется гомозиготным, или два разных, что приводит
к гетерозиготному организму.
30
31.
Аллельные гены – гены, отвечающие за развитиеальтернативных признаков.
Неаллельные гены – гены, отвечающие за развитие
разных (не альтернативных) признаков.
Гомозигота – организм, дающий один тип гамет.
Гетерозигота – организм, дающий несколько типов
гамет.
Гемизигота – организм, имеющий по одной аллели
какого-либо гена (генов).
31
32.
Доминантность (доминирование) — формавзаимоотношений между аллелями одного гена, при
которой один из них (доминантный) подавляет
(маскирует) проявление другого (рецессивного) и таким
образом определяет проявление признака как у
доминантных гомозигот, так и у гетерозигот.
Рецессивный признак — признак, не
проявляющийся у гетерозиготных особей вследствие
подавления проявления рецессивного аллеля.
32
33.
Моногибридное скрещивание – скрещиваниеорганизмов, различающихся по проявлениям одного
изучаемого признака, за которые отвечают аллели
одного гена.
Дигибридное скрещивание – скрещивание
организмов, различающихся по проявлениям двух
изучаемых признаков, за которые отвечают аллели двух
неаллельных генов.
Полигибридное скрещивание – скрещивание
организмов, различающихся по проявлениям нескольких
(многих) изучаемых признаков, за которые отвечают
аллели нескольких (многих) неаллельных генов.
33
34.
2. Принцип гибридологического анализа. ЗаконыМенделя, их цитологические основы. Закон
«чистоты гамет». Анализирующее скрещивание.
Летальные гены.
Гибридологический
анализ
–
способ
изучения
наследственных свойств организма путём скрещивания
(гибридизации)
его
с
родственной
формой
и
последующим анализом признаков потомства.
Гибридологический анализ впервые применил Г. Мендель
(1865) для изучения механизма передачи наследственных
задатков от родителей потомкам и для изучения
взаимодействия генов у одного и того же организма.
34
35.
Принципы гибридологического анализа.Из всех признаков организма выбирают только
альтернативные признаки — такие, которые имеют только
два четко различающихся варианта.
Производится большое количество скрещиваний в
не менее чем в двух поколениях.
Производится точный количественный учет и
анализ результатов большого числа скрещиваний для
устранения роли случайных отклонений.
35
36.
Законы Менделя.Первый закон Менделя (Закон единообразия гибридов
первого поколения или закон доминирования признаков).
При скрещивании гомозиготных организмов,
различающихся по одной паре альтернативных
(контрастных) признаков, получается первое поколение
гибридов единообразное по фенотипу и генотипу.
По фенотипу все гибриды первого поколения
характеризуются доминантным признаком, по генотипу
всё первое поколение гибридов гетерозиготное.
36
37.
Схема первого закона МенделяА-
доминант
ный
аллель
гена
P
♀ AA
а-
рецессив
ный
аллель
гена
G
A
x
♂ aa
a
F1
Aa
генотип
100% гетерозиготы
фенотип
100% имеют
доминантный признак
37
38.
Цитологические основы первого закона Менделя.1. При формировании гамет (гаметогенезе)
родительский гомозиготный организм образует только
один сорт гамет.
2. При оплодотворении гаметы родителей
сливаются только в одном варианте, формируя только
один сорт зигот.
38
39.
Условия выполнения законов Менделя.В соответствии с законами Менделя наследуются
только моногенные признаки.
Если за фенотипический признак отвечает более
одного гена (а таких признаков абсолютное
большинство), он имеет более сложный характер
наследования.
39
40.
Второй закон Менделя (Закон расщепления признаков).При скрещивании гибридов (гетерозигот),
анализируемых по одной паре альтернативных признаков
(при моногибридном скрещивании), в потомстве
наблюдается расщепление по фенотипу в соотношении
3:1 (3/4 гибридов поколения имеют доминантный признак,
около 1/4 - рецессивный), а по генотипу 1:2:1 (1/4
потомков – рецессивные гомозиготы, ½ - гетерозиготы,
1/4 – доминантные гомозиготы).
Расщепление – явление, при котором скрещивание
гетерозиготных особей приводит к образованию
потомства, часть которого несет доминантный, а часть –
рецессивный признак.
40
41.
Схема первого закона МенделяА-
доминантный
аллель гена
P1
♀ AA
а-
рецессивный
аллель гена
G
A
x
♂ aa
a
F1
Aa
генотип
100% гетерозиготы
фенотип
100% имеют доминантный признак
Схема второго закона Менделя
P2
G
♀ Aa
А
x
а
♂ Aa
А
а
F2
1АА
2Аа
1аа
генотип
25% (1/4)
доминантные
гомозиготы
50% (2/4)
гетерозиготы
25% (1/4)
рецессивные
гомозиготы
фенотип
75% (3/4) имеют доминантный
признак
25% (1/4)
имеют
рецессивный
признак
41
42.
В основе выполнения законов Менделя лежитгипотеза «чистоты гамет».
Гипотеза «чистоты гамет». Мендель предположил,
что при образовании половых клеток в каждую гамету
попадает только один наследственный фактор из пары
данных наследственных факторов. При образовании
гибридов наследственные факторы не смешиваются, а
сохраняются в неизменном виде.
42
43.
Во времена Менделя эта гипотеза не имеладостаточных подтверждений. Термин «ген» не
использовали.
В дальнейшем гипотеза была подтверждена
цитологическими наблюдениями и стала законом.
Закон чистоты гамет. В каждую гамету
попадает только одна аллель из пары аллелей
данного гена родительской особи.
43
44.
Цитологические основы второго закона Менделя.1. При формировании гамет (гаметогенезе)
родительский гетерозиготный организм образует
несколько сортов гамет. Например, при моногибридном
скрещивании у каждого родителя равновероятно
образуется два сорта гамет: А, а.
2. При оплодотворении гаметы родителей сливаются в
разных сочетаниях, формируя разные сорта зигот.
Например, при моногибридном скрещивании в потомстве
образуются зиготы с вероятностями 1АА:2Аа:1аа.
44
45.
Условия выполнения закона расщепления примоногибридном скрещивании.
1. Изучается большое число скрещиваний (большое
число потомков).
2. Гаметы, содержащие аллели А и а, образуются в
равном числе (обладают равной жизнеспособностью).
3. Нет избирательного оплодотворения: гаметы,
содержащие любой аллель, сливаются друг с другом с
равной вероятностью.
4. Зиготы (зародыши) с разными генотипами
одинаково жизнеспособны.
45
46.
Третий закон Менделя (Закон независимогонаследования признаков).
При дигибридном (полигибридном) скрещивании
гетерозигот каждая пара признаков наследуется
независимо от других пар и дает расщепление 3:1 по
каждой паре.
Например, скрещивались растения, отличающиеся по двум парам
признаков, таким как белые и пурпурные цветы и желтые или
зелёные горошины. Первое поколение после скрещивания обладало
доминантным фенотипом по всем признакам. Во втором поколении
наблюдалось расщепление фенотипов по формуле 9:3:3:1, то есть
9:16 были с пурпурными цветами и желтыми горошинами, 3:16 с
белыми цветами и желтыми горошинами, 3:16 с пурпурными цветами
и зелёными горошинами, 1:16 с белыми цветами и зелёными
горошинами.
46
47.
Схема третьего закона МенделяА
-
доминантный
аллель 1-го гена
P1
♀ ААВВ
♂ аавв
а-
рецессивный
аллель 1-го гена
G
АВ
ав
В
-
доминантный
аллель 2-го гена
F1
в-
рецессивный
аллель 2-го гена
генотип
100% гетерозиготы
фенотип
100% имеют два
доминантных признака
АаВв
47
48.
P2♀ АаВв
х
G АВ
Ав
аВ
ав
F2
♀\♂
АВ
Ав
♂ АаВв
АВ
аВ
Ав аВ ав
ав
АВ ААВВ ААВв АаВВ АаВв
Ав
ААВв ААвв АаВв Аавв
аВ
АаВВ АаВв ааВВ ааВв
ав
АаВв
Аавв
ааВв
аавв
Расщепление по фенотипу
9 (А_В_):3(А_вв):3(ааВ_):1(аавв)
48
49.
Цитологические основы третьего закона Менделя.1.При формировании гамет (гаметогенезе)
родительский гетерозиготный организм образует
несколько сортов гамет. При этом доминантные аллели
каждой пары аллельных генов представлены в 50%
гамет и рецессивные аллели каждой пары аллельных
генов представлены в 50% гамет.
2. При оплодотворении гаметы родителей сливаются
в разных сочетаниях, формируя разные сорта зигот в
определенных соотношениях.
49
50.
Условия выполнения закона независимогонаследования.
1.Все условия, необходимые для выполнения закона
расщепления.
2. Расположение генов, отвечающих за изучаемые
признаки, в разных парах хромосом (несцепленность).
50
51.
Анализирующее скрещивание.Анализирующее скрещивание - скрещивание особи
с доминантным признаком (анализируемой особи) с
особью, гомозиготной по рецессивным аллелям
(анализатором).
Анализирующее скрещивание позволяет определить
генотип и соотношение гамет разного типа,
образуемых анализируемой особью.
51
52.
Если в потомстве от анализирующего скрещиваниярасщепления нет, то анализируемая особь гомозиготна.
P
♀ AA
G
A
F1
фенот
ип
x
♂ aa
a
Aa
100% имеют доминантный признак. Расщепления
нет. Анализируемая особь гомозиготна
52
53.
Если расщепление есть, то анализируемая особьгетерозиготна.
P
G
F1
♀ Aа
A
x
а
Aa
фенот
50% имеют
ип
доминантный признак
♂ aa
a
аа
50% имеют
рецессивный признак.
Расщепление есть. Анализируемая особь
гетерозиготна
53
54.
Летальные гены.Летальные гены (летальные аллели) - гены, которые
вызывают гибель организма.
Эти гены, как правило, рецессивные. Гибель, как
правило, наступает у гомозигот. Гибель организма
наступает при определенных условиях, или на
определенных этапах развития (чаще всего на
эмбриональных стадиях развития). Летальные аллели
возникают в результате летальных мутаций генов,
ответственных за какую-либо жизненно важную
функцию.
54
55.
В случае наличия летальных генов возникает отклонениеот менделирующего (возникающего по законам Менделя)
расщепления.
Схема скрещивания при наличии летального гена. Расщепление 2:1.
А доминантный
аллель гена
P2
а
G
летальный
рецессивный
аллель гена
♀ Aa
А
x
а
♂ Aa
А
а
F2
1АА
2Аа
1аа
генотип
25% (1/4)
доминантные
гомозиготы
50% (2/4)
гетерозиготы
25% (1/4)
рецессивные
гомозиготы
фенотип
Расщепление
среди
выживших 2:1.
75% выживают
1/3 выживших имеет
одно проявление
признака
25% (1/4)
гибнут
2/3 выживших имеет
другое проявление
признака
55
56.
3. Доминантный и рецессивный типынаследования признаков человека.
Промежуточный характер наследования.
Человек, как объект генетических исследований,
имеет ряд особенностей. Над человеком нельзя
проводить эксперименты, в том числе,
экспериментальное скрещивание. У человека
небольшое количество потомков. Продолжительность
жизни человека, за проявлениями генетических
закономерностей которого наблюдают, приблизительно
равна продолжительности жизни наблюдающих ученых.
Однако, генотип и фенотип человека является хорошо
изученными, что облегчает исследования генетики
человека.
56
57.
Наследование признаков человека (доминантных ирецессивных), которое подчиняется законам Менделя,
называется менделирующим.
По доминантному типу наследуются некоторые нормальные
менделирующие признаки человека, например: белый локон
надо лбом; волосы жесткие, прямые; шерстистые волосы короткие, легко секущиеся, курчавые, пышные; толстая
кожа; способность свертывать язык трубочкой.
По доминантному типу наследуются некоторые
патологические менделирующие признаки человека,
например: полидактилия (многопалость); синдактилия
(сращение мягких или костных тканей фаланг двух или
более пальцев); брахидактилия (короткопалость,
недоразвитие дистальных фаланг пальцев).
57
58.
По рецессивному типу наследуются некоторыенормальные признаки человека, например:
мягкие, прямые волосы; тонкая кожа;
отрицательный резус-фактор; неощущение вкуса
фенилкарбамида; неумение складывать язык
трубочкой.
По рецессивному типу наследуются некоторые
патологические признаки человека, например:
фенилкетонурия; галактоземия; альбинизм.
58
59.
Промежуточный характер наследования.Промежуточный характер наследования наблюдается
при неполном доминировании, когда один аллельный ген
не полностью подавляет фенотипическое проявление
другого аллельного гена. У гетероозигот формируется
промежуточный признак.
59
60.
Схема промежуточного наследования признака.АА
- Генотип, при котором
формируется
доминантный признак
P2
♀ Aa
Аа
- Генотип, при котором
формируется
промежуточный признак
G
аа
- Генотип, при котором
формируется
рецессивный признак
F2
1АА
2Аа
1аа
ге
но
ти
п
25% (1/4)
доминантные
гомозиготы
50% (2/4)
гетерозиготы
25% (1/4)
рецессивные
гомозиготы
А
x
а
♂ Aa
А
а
Расщепление по генотипу 1:2:1
фе
но
ти
п
25% (1/4) имеют
доминантный
признак
50% (2/4) имеют
промежуточный
признак
25% (1/4) имеют
рецессивный
признак
Расщепление по фенотипу 1:2:1
60
61.
Например. По промежуточному типу наследуетсяталассемия человека.
Талассемия (анемия гемолитическая мишеневидноклеточная) – наследственная гемолитическая анемия,
характеризующаяся нарушением синтеза глобина
(белка, входящего в состав гемоглобина).
Талассемия большая – гомозиготная форма
талассемии (генотип аа), характеризующаяся
выраженной гипохромной анемией, наличием
мишеневидных эритроцитов, увеличением селезенки,
утолщением длинных трубчатых костей и костей
черепа.
61
62.
Мишеневидные эритроциты.Талассемия малая – гетерозиготная форма талассемии
(генотип Аа), характеризующаяся легкой гипохромной
анемией, небольшим количеством мишеневидных
эритроцитов, некоторым увеличением селезенки.
62
63.
4. Взаимодействие аллельных инеаллельных генов. Полигенное наследование
признаков.
Взаимодействие генов - участие двух или более
генов в генетическом контроле формирования
признака.
Виды взаимодействия аллельных генов.
Полное доминирование – взаимодействие двух
аллелей одного гена, при котором доминантная
аллель полностью исключает проявление действия
второй аллели. В фенотипе присутствует только
признак, задаваемый доминантной аллелью.
Например, прямые волосы у человека.
63
64.
Неполное доминирование – доминантная аллель вгетерозиготном состоянии не полностью подавляет
действие рецессивной аллели. Гетерозиготы имеют
промежуточный характер признака. Например,
микрофталмия, таласемия у человека.
Сверхдоминирование – более сильное проявление
признака у гетерозиготной особи, чем у гомозиготной.
Например, у человека гетерозиготы по
серповидноклеточной анемии обладают большей
резистентностью к тропической малярии, вызываемой
протозойным паразитом Plasmodium falciparum, чем
индивиды, гомозиготные по нормальной доминантной
аллели гена, определяющего форму эритроцитов.
У дрозофилы у гетерозигот по отдельным мутациям
наблюдаются более высокие показатели плодовитости,
чем у гетерозигот.
64
65.
Кодоминирование – проявление у гибридов новогопризнака, обусловленного взаимодействием двух
разных аллелей одного гена. При этом оба аллеля гена
проявляются в фенотипе. Например, наследование
четвертой группы крови у человека.
Аллельное исключение – вид взаимодействия
аллельных генов, при котором в части клеток
организма фенотипически проявляется одна аллель, а
в другой части клеток организма – другая аллель.
Например, в данном В лимфоците экспрессируется
только одна аллель гена иммуноглобулина.
65
66.
Виды взаимодействия неаллельных генов.Комплементарность – форма взаимодействия
неаллельных генов, при котором одновременное
действие нескольких доминантных генов дает новый
признак.
Например, развитие нормального слуха – это
результат совместного действия нескольких неаллельных
генов.
66
67.
Например, синтез какого-либо гормона определяетсяодним геном, а проникновение этого гормона в клеткимишени зависит от белков рецепторов, которые
кодируются другим неаллельным геном.
Стандартное расщепление (расщепление по фенотипу в
потомстве от скрещивания двух гетерозиготных
родительских организмов) при комплеметарном
взаимодействии наблюдается в отношении 9:3:3:1 (но
здесь нельзя найти независимого расщепления по
каждой паре аллелей в отношении 3:1), 9:7 и 9:6:1.
67
68.
Примерскрещивания
при
комплементарном
взаимодействии генов. Форма гребня у кур может быть
листовидной гороховидной, розовидной и ореховидной.
При скрещивании кур, имеющих ореховидные гребни,
потомство получилось со всеми четырьмя формами
гребней в отношении: девять ореховидных, три
гороховидных, три розовидных, один листовидный.
Определите генотипы родителей.
68
69.
Пример скрещивания при комплементарностиА_В_
- ореховидная форма гребня
А_вв
- гороховидная форма гребня
ааВ_
- розовидная форма гребня
аавв
- листовидная форма гребня
P
G
F
♀ АаВв
АВ
Ав
аВ
х
ав
♂ АаВв
АВ
Ав
аВ
ав
♀\
♂
АВ
Ав
аВ
ав
АВ
АВАВ
ААВв
АаВВ
АаВв
Ав
ААВв
ААвв
АаВв
Аавв
аВ
АаВВ
АаВв
ааВВ
ааВв
ав
АаВв
Аавв
ааВв
аавв
Расщепление по фенотипу 9 ореховидных (А_В_) : 3 гороховидных (А_вв) : 3 розовидных
(ааВ_): 1 листовидный (аавв).
Ответ: генотипы родителей ♀ АаВв, ♂ АаВв
69
70.
Эпистаз – взаимодействие генов, при которомактивность одного гена подавляется влиянием других
неаллельных ему генов.
Ген, подавляющий фенотипические проявления
другого, называется эпистатичным; ген, чья активность
изменена или подавлена, называется гипостатичным.
Если эпистатический ген является доминантным, то
имеет место доминантный эпистаз, если рецессивным, то
рецессивный эпистаз.
70
71.
Стандартное расщепление (расщепление по фенотипу впотомстве от скрещивания двух гетерозиготных
родительских организмов) при доминантном эпистазе
оказывается 13:3 или 12:3:1. Стандартное расщепление
при рецессивном эпистазе наблюдается в отношении
9:3:4.
Примером рецессивного эпистаза у человека является
бомбейский феномен. У женщины с первой и у мужчины
со второй группой крови родился ребенок с четвертой
группой крови. При исследовании данного случая у
матери был найден эпистатический ген с, который
подавлял ген IВ.
71
72.
Пример скрещивания при рецессивном эпистазе.«Бомбейский феномен».
IA,IB,I 0
- аллели гена группы крови
С
- ген, который не подавляет ген IВ
с
- ген, который подавляет ген IВ
P
♀ IBI Bсс
G
IBс
F
х
♂ IAIAСС
IA С
IAIBСс
IV (AB) группа крови. Подавления нет.
72
73.
Пример скрещивания при доминантном эпистазе.Белое оперение кур определяется двумя парами не
сцепленных неаллельных генов. В одной паре
доминантный определяет окрашенное оперение
рецессивный — белое. В другой паре доминантный ген
подавляет окраску, рецессивный — не подавляет
окраску. При скрещивании белых кур получено
потомство из 1680 цыплят. 315 цыплят было
окрашенных, остальные белые. Определите генотипы
родителей.
73
74.
Пример скрещивания при доминантном эпистазеА
— окрашенное оперение
а
— белое оперение
В
— подавляет окраску
в
— не подавляет окраску
P
G
F
♀ АаВв
АВ
Ав
аВ
х
♂ АаВв
ав
АВ
Ав
аВ
ав
♀\♂
АВ
Ав
аВ
ав
АВ
АВАВ
ААВв
АаВВ
АаВв
Ав
ААВв
ААвв
АаВв
Аавв
аВ
АаВВ
АаВв
ааВВ
ааВв
ав
АаВв
Аавв
ааВв
аавв
Расщепление по фенотипу: 9 белые (А_В_) : 3 окрашенные (А_вв) : 3 белые (ааВ_)
: 1 белый (аавв).
Ответ: генотипы родителей ♀ АаВв, ♂ АаВв
74
75.
Полимерия (полигения, полигенное наследованиепризнаков) – обусловленность развития одного
признака многими неаллельными генами, действие
которых суммируется в признаке.
Стандартное расщепление (расщепление по
фенотипу в потомстве от скрещивания двух
гетерозиготных родительских организмов) при
полимерии оказывается 1:4:6:4:1.
Бывают случаи, когда полимерные гены не
усиливают друг друга. Тогда расщепление будет 15:1.
75
76.
Примером полимерии является наследование цветакожи у человека. Этот признак определяется четырьмя
генами, ответственными за выработку пигмента
меланина. Чем больше доминантных неаллельных
генов, тем темнее кожа.
Широко распространенные болезни человека,
например, артериальная гипертензия, ишемическая
болезнь сердца, бронхиальная астма, язвенная болезнь
желудка, наследуются полигенно. При этом тяжесть
заболевания зависит не только от совокупного действия
множества генов, но и от провоцирующих средовых
факторов.
76
77.
5. Первичная и вторичная плейотропия.Плейотропия множественное действие гена,
способность одного гена воздействовать на проявление
нескольких разных признаков организма.
Плейотропное действие гена может быть первичным и
вторичным.
77
78.
При первичной плейотропии ген проявляет своймножественный эффект в формировании разных
признаков одновременно. Причинно-следственная
связь в развитии этих признаков отсутствует.
Это обусловлено тем, что продукт гена участвует в
нескольких, а иногда и в очень многих процессах.
Например, ген, обуславливающий рыжие волосы,
обуславливает более светлую окраску кожи и
появление веснушек.
78
79.
При синдроме Марфана арахнодактилия, вызываемаядоминантной мутацией, проявляется одновременно в
изменениях пальцев рук и ног, вывихах хрусталика глаза
и врождённых пороках сердца.
79
80.
При болезни Хартнупа мутация гена приводит кнарушению всасывания аминокислоты триптофана в
кишечнике и его реабсорбции в почечных канальцах.
При этом поражаются одновременно мембраны
эпителиальных клеток кишечника и почечных канальцев
с расстройствами пищеварительной и выделительной
систем.
80
81.
При вторичной плейотропии есть один первичныйэффект проявление гена, вслед за которым развивается
ступенчатый процесс вторичных изменений, приводящих
к множественным эффектам. При этом в развитии
признаков присутствует причинно-следственная связь.
81
82.
Например, при серповидно-клеточной анемии у гомозиготнаблюдается несколько патологических признаков:
Первопричиной, непосредственным проявлением
дефектного гена является аномальный гемоглобин и
эритроциты серповидной формы. Вследствие этого
происходят последовательно другие патологические
процессы: слипание и разрушение эритроцитов, анемия,
нарушения в почках, сердце, мозге - эти патологические
признаки вторичны.
Нормальные и серповидные
эритроциты при серповидноклеточной анемии.
82
83.
Схема развитиясимптомов
(признаков)
при
серповидноклеточной
анемии.
83
84.
6. Множественные аллели.Множественный аллелизм - это существование в
популяции более двух аллелей данного гена.
Они возникают в результате разных мутаций одного
гена.
Например, для у глобиновых генов человека известно
несколько сотен аллелей.
У дрозофилы была обнаружена серия множественных
аллелей по гену white, состоящая из нескольких
десятков мутантных и одной нормальной аллели.
84
85.
7. Наследование групп крови человека посистемам АВ0 и MN. Наследование резусфактора. Резус-конфликт.
Группа крови - описание индивидуальных антигенных
характеристик эритроцитов, определяемое с помощью методов
идентификации специфических групп углеводов и белков,
включённых в мембраны эритроцитов (антигенов).
У человека открыто несколько систем антигенов,
основные из них: система AB0, система Rh (резуссистема).
Известно также 46 классов других антигенов, из которых
большинство встречается гораздо реже, чем AB0 и резус-фактор.
Это такие системы антигенов как система Келл (Kell), система
Кидд (Kidd), система Даффи (Duffy), система MNSs, система Lewis,
система P, система Xg и другие.
85
86.
Наследование групп крови человека по системе АВ0(ABO histo-blood group glycosyltransferases).
В 1924 г. Bernstein установил, что система групп
крови АВ0 контролируется серией множественных
аллелей.
В системе АВ0 синтез агглютиногенов и
агглютининов определяется аллелями гена I: I0, IA, IB
(пример множественных аллелей у человека).
86
87.
Ген I контролирует и образование антигенов, иобразование антител. При этом наблюдается полное
доминирование аллелей IA и IB над аллелем I0, но
совместное доминирование (кодоминирование)
аллелей IA и IB. Генный локус для этих аллелей
находится на длинном плече хромосомы 9.
В плазме крови человека могут содержаться
агглютинины α и β, в эритроцитах - агглютиногены A и
B, причём из белков A и α содержится только один, то
же самое - для белков B и β.
87
88.
Таблица соответствия генотипов, агглютиногенов,агглютининов и групп крови (фенотипов).
Антигены
(агглютиногены)
Антитела
(агглютинины)
Группы
крови
(фенотипы)
I 0 I0
нет
,
I (0)
IAIA, IAI 0
А
II (A)
IBI B, IBI0
В
III (B)
IA IB
А, В
нет
Генотипы
IV (AB)
88
89.
Если по каким-либо причинам агглютиноген Австречается с агглютинином a или агглютиноген В
встречается с агглютинином b, то происходит реакция
агглютинации – склеивания эритроцитов. В дальнейшем
агглютинированные эритроциты подвергаются гемолизу
(разрушению), продукты которого ядовиты. Такое
возможно при переливании несовместимой по группе
крови.
89
90.
Наследование групп крови человека по системе MN(MNSs).
Групповая система MNSs является сложной
системой; она состоит из 9 групп крови: MNSs, MNs, Ns,
Mss, Ms, MS, NSs, MNS и Ns.
Гены антигенов системы групп крови MNSs
локалазованы в 4-ой хромосоме.
Имеется ген системы групп крови MN с двумя
кодоминантными аллелями M и N. Близко к нему
располагается другой ген, с доминантным аллелем S и
рецессивным s, определяющий систему групп крови Ss.
90
91.
Наследование групп крови системы Rhesus.Резус-система определяется тремя сцепленными
генами (CDE); все эти гены локализованы в 1-й
хромосоме.
Наиболее сильным антигеном резус-системы является
антиген RhD, который контролируется соответствующим
геном D.
При этом резус-положительная группа крови
доминирует над резус-отрицательной.
91
92.
Наследование резус-фактора происходит сложнымобразом, но,
учитывая ведущую роль гена D, его можно представить
как моногенное наследование с полным доминированием:
при генотипе DD или Dd резус положительный (Rh+),
а при генотипе dd – отрицательный (Rh–).
92
93.
Таблица соответствия генотипов, антигенов, антители групп крови резус-системы.
Генотипы
Антигены
Группы крови
(фенотипы)
DD, Dd
есть
Rh+
dd
нет
Rh–
93
94.
Резус-конфликт.Если резус-отрицательная женщина вынашивает
резус-положительный плод, то при попадании
эритроцитов плода в кровоток матери происходит
иммунизация организма матери резус-антигеном.
Вырабатываются резус-антитела.
Резус-антитела через плаценту попадают в
кровоток плода. Резус-антитела матери разрушают
резус-положительные эритроциты плода. Это
приводит или к гибели плода, или к рождению
неполноценного ребенка.
94
95.
Развитие резус конфликта.I. Если у плода кровь резус-положительная (красные круги), а у матери
резус-отрицательная (синие круги), развивается резус-конфликт. В ответ на
резус-положительный белок, содержащийся в эритроцитах плода, организм
матери вырабатывает антитела (черные полукружия).
II. Через плаценту антитела проникают в кровь плода и разрушают его
95
эритроциты. Образующийся при этом продукты повреждает структуры плода.
96.
8. Иммуногенетика: предмет и задачи.Тканевая и видовая специфичность белков.
Антигенные свойства белков.
Иммуногенетика - раздел генетики, исследующий
закономерности наследования антигенов, других
факторов иммунитета и тканевой несовместимости.
Начало иммуногенетики положили работы немецких
учёных П. Эрлиха и Ю. Моргенрота, обнаруживших в
начале двадцатого века. Группы крови у коз, и
открытие К. Ландштейнером групп крови у человека.
Термин «иммуногенетика» предложен американским
учёным М. Ирвином в 1930.
96
97.
Основные задачи иммуногенетики:1) изучение проблем генетики гистосовместимости;
2) изучение проблем генетического контроля
структуры иммуноглобулинов и других
иммунологически значимых молекул;
3) изучение проблем генетического контроля силы
иммунного реагирования;
4) изучение генетики антигенов.
97
98.
Изучение проблем генетики гистосовместимостисвязана с развитием несовместимости тканей при
внутривидовых пересадках.
Описан комплекс генов, контролирующих
поверхностные клеточные структуры, которые
вызывают иммунную реакцию отторжения чужеродной
ткани.
Эти гены кодируют молекулы (антигены)
гистосовместимости (главный комплекс
гистосовместимости - MHC или HLA).
98
99.
Антигены HLA подразделяются на антигены класса I иантигены класса II.
Антигены HLA класса I необходимы для распознавания
трансформированных клеток цитотоксическими Тлимфоцитами.
Функция антигено