









")
")

")





")




")
 medicine
medicineSimilar presentations:










Мышечные ткани. Виды мышечных тканей
1. Мышечные ткани
Виды мышечных тканейI. Поперечнополосатые
(исчерченные)
мышечные ткани
II. Гладкие
(неисчерченные)
мышечные ткани
Происхождени
е
1. Скелетная
мышечная ткань
Из миотомов
2. Сердечная
мышечная ткань
Из
миоэпикардиальной
пластинки
(находящейся в
составе
висцерального
листка
спланхнотома).
1. Гладкая мышечная
ткань сосудов и
внутренних органов
2. Мышечная ткань
нейрального
происхождения
Из мезенхимы
Из клеток
нейрального зачатка
2. Типы мышечных тканей
3. Гладкая мышечная ткань
1 – продольныйсрез гладких
мышечных клеток,
1
2
3
2 – поперечный
срез пучков
мышечных клеток,
3 – рыхлая
соединительная
ткань (эндомизий)
4. Сократительный аппарат гладкой мышечной ткани
• в покое в клетках нет миофибрилл (отчегоклетки не имеют поперечной исчерченности).
• Тонкие (актиновые) миофиламенты состоят
только из актина (т.е. не содержат тропонина
и тропомиозина) и прикрепляются к т.н.
плотным тельцам
• Толстые же (миозиновые) миофиламенты в
состоянии покоя диссоциированы на
отдельные молекулы миозина и поэтому не
имеют фиксированного положения
5. Процесс сокращения гладких миоцитов
• при возбуждении ионы Ca входят в клетку черезCa2+-каналы (а не из СПР!!!)
• ионы Ca2+, связавшись с белком кальмодулином,
активируют миозинкиназу, которая
фосфорилирует молекулы миозина.
• Это придает молекулам миозина способность объединяться в толстые миофиламенты и
взаимодействовать с тонкими миофиламентами.
• Толстые миофиламенты внедряются между тонкими
– образуются временные миофибриллы.
• миофиламенты перемещаются навстречу друг другу
(за счёт образования и разрыва мостиков и
гидролиза АТФ).
6. Выход из сокращения
• инициируется удалением ионов Ca2+ изгладкого миоцита Ca2+-насосами.
• Миозинфосфатаза
дефосфорилирилирует миозин
• толстые миофиламенты распадаются на
фрагменты или молекулы миозина и клетка
возвращается в расслабленное состояние
• C другой стороны, гладкие миоциты могут
пребывать в сокращённом состоянии
достаточно долго без заметного
утомления. Это объясняется тем, что часть
миозиновых мостиков сохраняется и после
дефосфорилирования миозина.
7. Гладкая мышечная ткань
8. Поперечнополосатая мышечная ткань
1a
b
c
2
d
e
3
4
1 – продольный срез
поперечнополосатых
мышечных волокон:
а – А диски
(анизотропные темные
диски),
b – I диски (изотропные
светлые диски),
c – ядро,
2 – поперечный срез
поперечнополосатых
мышечных волокон: d –
миофибриллы, e – ядро,
3 – эндомизий
(прослойки рыхлой
соединительной ткани),
4 – кровеносные сосуды
9. Поперечнополосатая мышечная ткань
10. Васкуляризация поперечнополосатой мышечной ткани
11. Диафрагма крысы (СЭМ)
Скол мышечноговолокна диафрагмы
крысы. Видны
саркомеры и цистерны
саркоплазматического
ретикулума (указано
стрелкой).
Увеличение 12 500.
12. Тонкая структура саркомера (ЭМФ и схема)
Z - телофрагма1 - актиновые миофиламенты
2 - миозиновые миофиламенты
Н - Н-зона м/у концами актиновых миофиламентов
М - мезофрагма
миофибрилла
Поперечные срезы миофибриллы
вокруг каждого толстого миофиламента находятся 6 тонких,
а вокруг каждого тонкого - 3 толстые.
13. Тонкие и толстые миофиламенты
• Тонкие, или актиновые, миофиламентыобразованы глобулярным белком
актином, 350 молекул которого
объединяются в двойную спираль,
глобулярным белком тропонином и
фибриллярным белком тропомиозином
• в состоянии покоя тропонином и
тропомиозин блокируют активные
центры актина, что исключает
взаимодействие миофиламентов.
• В толстом миофиламенте – примерно 300
молекул миозина. Причём их стержни
плотно упакованы в толстом филаменте, а
головки выступают наружу и при
сокращении участвуют во взаимодействии
с тонкими филаментами.
14. Мембраны мышечного волокна (схема)
• 1-миофибрилы,• 2-СПР-гладкая
ЭПС,
• 3-терминальные
цистерны,
• 4-Т-трубочкивпячивания
плазмолеммы,
• 5-митохондрия
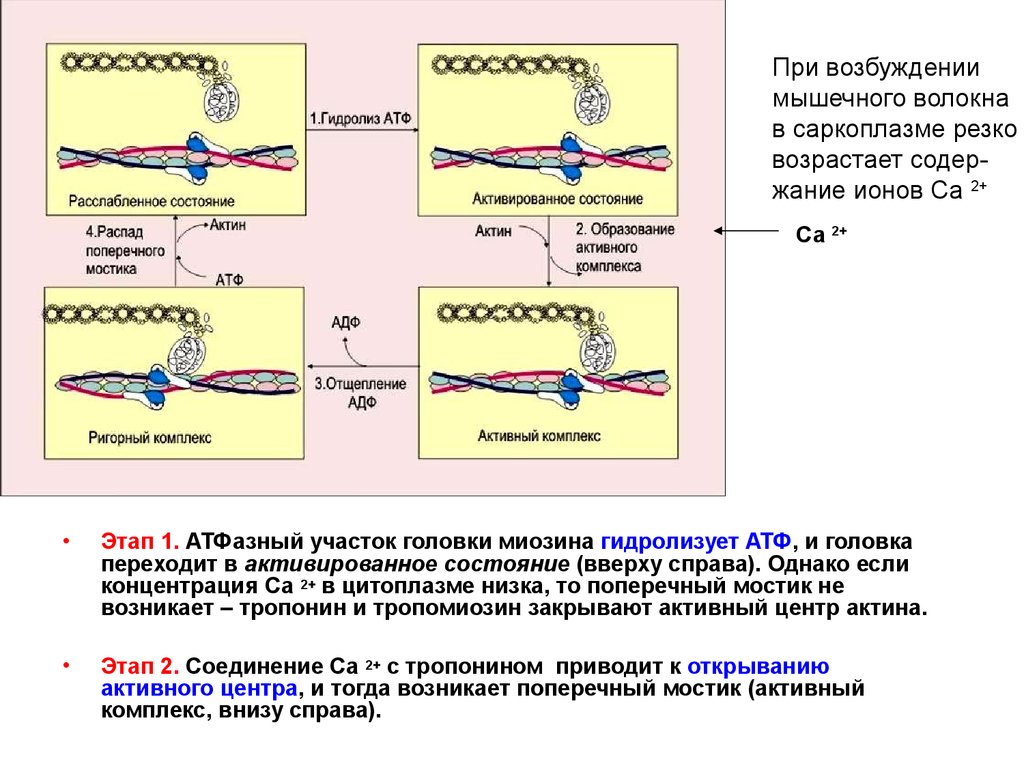
15.
При возбуждениимышечного волокна
в саркоплазме резко
возрастает содержание ионов Са 2+
Са 2+
Этап 1. АТФазный участок головки миозина гидролизует АТФ, и головка
переходит в активированное состояние (вверху справа). Однако если
концентрация Са 2+ в цитоплазме низка, то поперечный мостик не
возникает – тропонин и тропомиозин закрывают активный центр актина.
Этап 2. Соединение Са 2+ с тропонином приводит к открыванию
активного центра, и тогда возникает поперечный мостик (активный
комплекс, внизу справа).
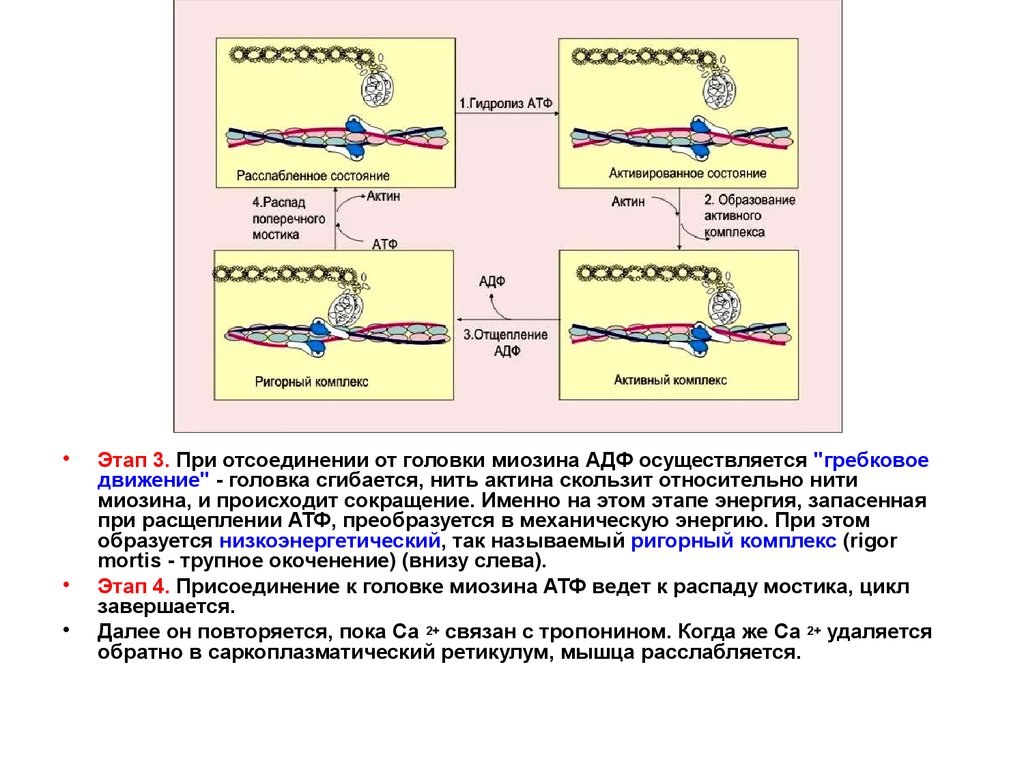
16.
Этап 3. При отсоединении от головки миозина АДФ осуществляется "гребковое
движение" - головка сгибается, нить актина скользит относительно нити
миозина, и происходит сокращение. Именно на этом этапе энергия, запасенная
при расщеплении АТФ, преобразуется в механическую энергию. При этом
образуется низкоэнергетический, так называемый ригорный комплекс (rigor
mortis - трупное окоченение) (внизу слева).
Этап 4. Присоединение к головке миозина АТФ ведет к распаду мостика, цикл
завершается.
Далее он повторяется, пока Са 2+ связан с тропонином. Когда же Са 2+ удаляется
обратно в саркоплазматический ретикулум, мышца расслабляется.
17. Схема, демонстрирующая молекулярные механизмы мышечного сокращения с точки зрения теории "скользящих нитей": (Источник: San Diego State University College
Схема, демонстрирующая молекулярные механизмы мышечногосокращения с точки зрения теории "скользящих нитей":
(Источник: San Diego State University College of Sciences [www.sci.sdsu.edu]
каталитический центр расщепления АТФ - АТФаза –
располагается непосредственно на миозиновой головке, однако
активируется он актином в присутствии ионов Mg 2+
18. Феномен трупного окоченения
• а) После смерти, в связи с угасанием метаболическихпроцессов, в мышцах быстро снижается
концентрация АТФ. Поэтому
• перестаёт функционировать Са2+ -насос,
и в саркоплазме повышается концентрация ионов
Са2+.
• б) Под их влиянием в мышцах замыкаются
мостики между тонкими и толстыми
миофиламентами.
• в) А разомкнуться они не в состоянии, т.к. для
этого требуется АТФ.
• г) Таким образом, существо трупного окоченения –
это постепенное замыкание всё большего числа
мостиков между миофиламентами.
19. Связь мышцы с сухожилием
32
1
1 – мышечные волокна, 2 – коллагенновые волокна, 3 – область контакта
мышечных и коллагенновых волокон. Здесь коллагеновые волокна проникают в
узкие впячивания сарколеммы и прикрепляются к базальной мембране,
окружающей мышечные волокна
20. Типы мышечных волокон
Реакция на сукцинатдегидрогеназу Реакция на АТФазную активность21. Двигательные нервные окончания
22. Двигательные нервные окончания (СЭМ)
23. Сердечная поперечнополосатая мышечная ткань
12
3
4
5
1 – кардиомиоциты,
формирующие
сетевую структуру
миокарда,
2 – вставочные диски
(места контактов
кардиомиоцитов),
3 – ядро,
4 – саркоплазма,
5 – кровеносный
сосуд
24. Сердечная поперечнополосатая мышечная ткань
25. Стенка желудочка сердца
Вставочные диски• В области вставочных дисков
(9) между кардиомиоцитами
существуют контакты трёх
видов:
• интердигитации –
пальцевидные впячивания
клеток друг в друга;
• десмосомы (10) – контакты,
обеспечивающие более прочное
сцепление клеток;
• нексусы (11) – контакты,
пронизанные гидрофильными
каналами и потому
обеспечивающие электрическую
и метаболическую связь между
кардиомиоцитами.
26. Вставочные диски
(нексусы и десмосомы)27. Вставочные диски (нексусы и десмосомы)
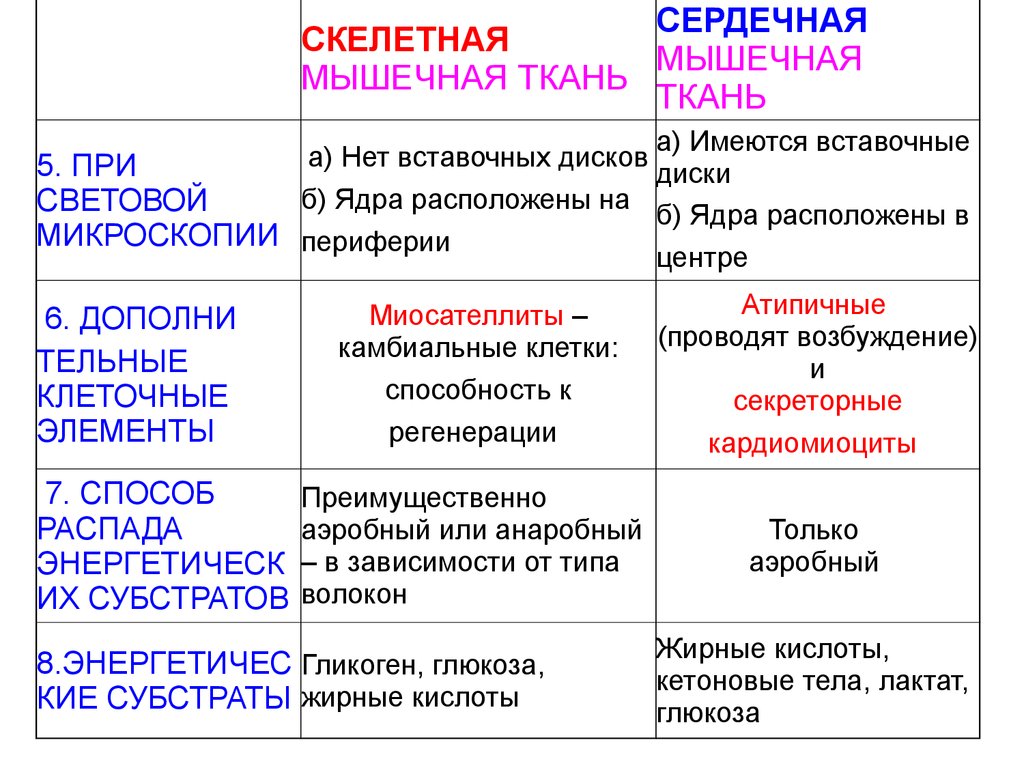
СКЕЛЕТНАЯМЫШЕЧНАЯ
ТКАНЬ
СЕРДЕЧНАЯ
МЫШЕЧНАЯ ТКАНЬ
1.ПРОИСХОЖДЕНИЕ
Из миотомов
Из миоэпикардиальной
пластинки
2. ОСНОВНЫЕ
СТРУКТУРНЫЕ
ЭЛЕМЕНТЫ
ТКАНИ
Миосимпласты –
основа мышечных
волокон
Типичные
кардиомиоциты,
объединяющиеся в
функциональные волокна
3. ПРИМЕРНЫЙ
ДИАМЕТР
ВОЛОКОН
50-70 мкм
20 мкм
70% объёма
волокна
40% объёма
кардиомиоцита
4. СОДЕРЖАНИЕ
МИОФИБРИЛЛ
28.
СЕРДЕЧНАЯСКЕЛЕТНАЯ
МЫШЕЧНАЯ
МЫШЕЧНАЯ ТКАНЬ
ТКАНЬ
а) Имеются вставочные
а)
Нет
вставочных
дисков
5. ПРИ
диски
б) Ядра расположены на
СВЕТОВОЙ
б) Ядра расположены в
МИКРОСКОПИИ периферии
центре
6. ДОПОЛНИ
ТЕЛЬНЫЕ
КЛЕТОЧНЫЕ
ЭЛЕМЕНТЫ
7. СПОСОБ
РАСПАДА
ЭНЕРГЕТИЧЕСК
ИХ СУБСТРАТОВ
Миосателлиты –
камбиальные клетки:
способность к
регенерации
Атипичные
(проводят возбуждение)
и
секреторные
кардиомиоциты
Преимущественно
аэробный или анаробный
– в зависимости от типа
волокон
Только
аэробный
8.ЭНЕРГЕТИЧЕС Гликоген, глюкоза,
КИЕ СУБСТРАТЫ жирные кислоты
Жирные кислоты,
кетоновые тела, лактат,
глюкоза