




















































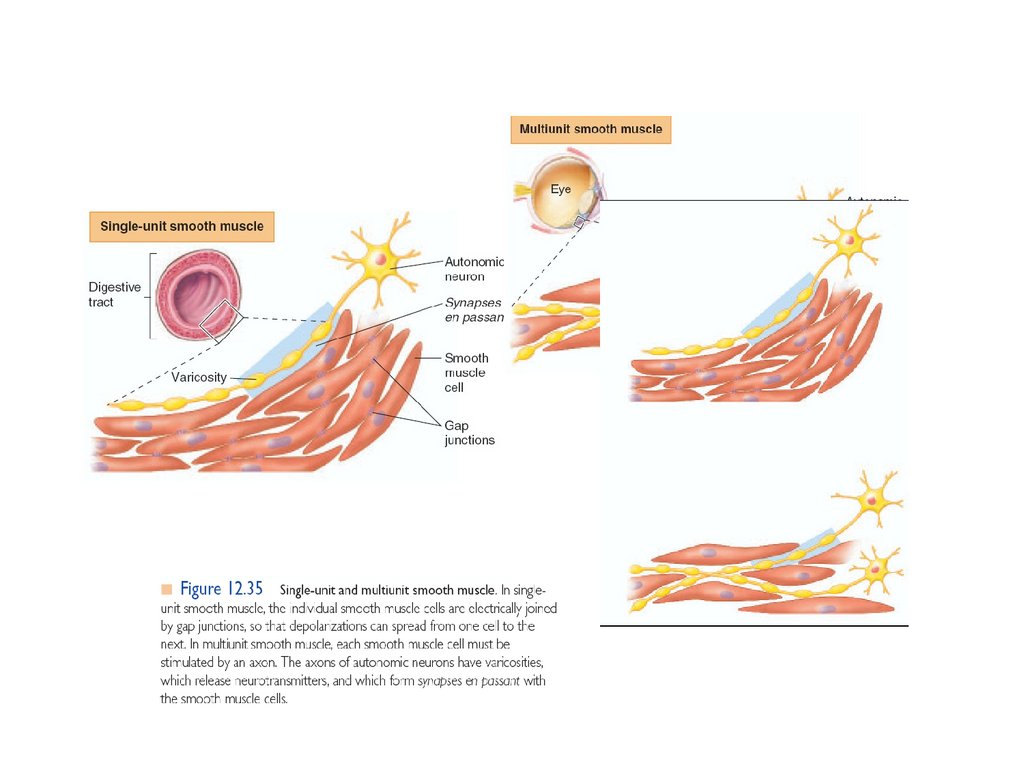



 medicine
medicine biology
biologySimilar presentations:










Физиология мышц
1.
ГОУ ВПО ОрГМА Росздрава кафедранормальной физиологии
Физиология
мышц
2.
Активное движение• Одно из характерных свойств всех живых
систем начиная от простейших и кончая
самыми сложными
3.
Биологическое движениеСокращение различных мышц,
Движение листьев,
Биение ресничек,
Движение жгутиков,
Деление клеток,
Движение протоплазмы.
4.
• Все разнообразные формы проявлениядвигательной активности имеют общую
черту – превращение химической энергии в
механическую.
5.
Формы движения• Амебойдное
• Мерцательное
• Мышечное
(для тел большой массы)
6.
Мышечное движение наиболееэффективный способ перемещения
•10,2 метра в
секунду
•километры
•0,2 миллиметра в
секунду
•сантиметры
7.
• Мышечными называют все типы клеток,функция которых состоит в сокращении.
8.
• У млекопитающих имеются три главныхтипа клеток, специально приспособленных
для сокращения: волокна скелетных мышц ,
клетки сердечной мышцы ,
гладкомышечные клетки.
9.
Классификации мышцГистологическая
АНАТОМИЧЕСКАЯ
СКЕЛЕТНЫЕ
Функциональная
произвольные
Поперечнополосатые
СЕРДЕЧНАЯ
непроизвольные
Гладкие
ВИСЦЕРАЛЬНЫЕ
10.
Скелетнаямышца
11.
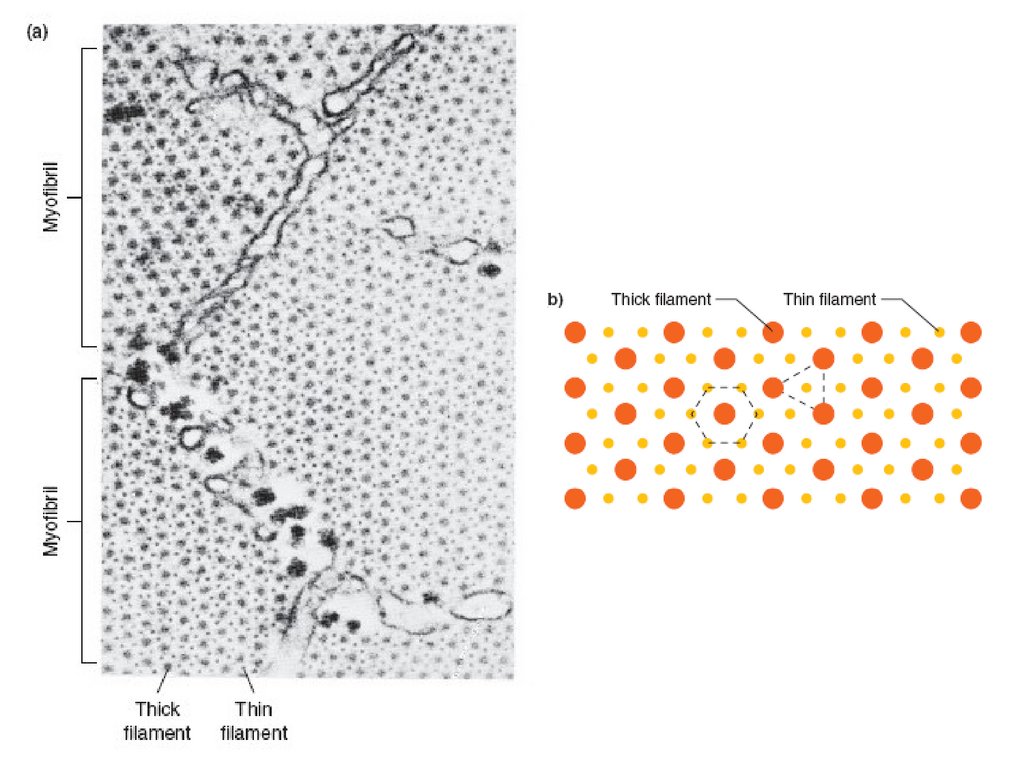
Уровни структурнойорганизации скелетных
мышц
Мышца
х1
Пучок мышечных
волокон
х5
Мышечное волокно
Х 500
Миофибрилла
Х 10000
Саркомер
Х 50000
Миофиламенты
Х 1000000
12.
миофибрилласаркомер
Z - линия
Z - линия
Тонкие нити
М - линия
Титиновые нити
Толстые нити
13.
14.
15.
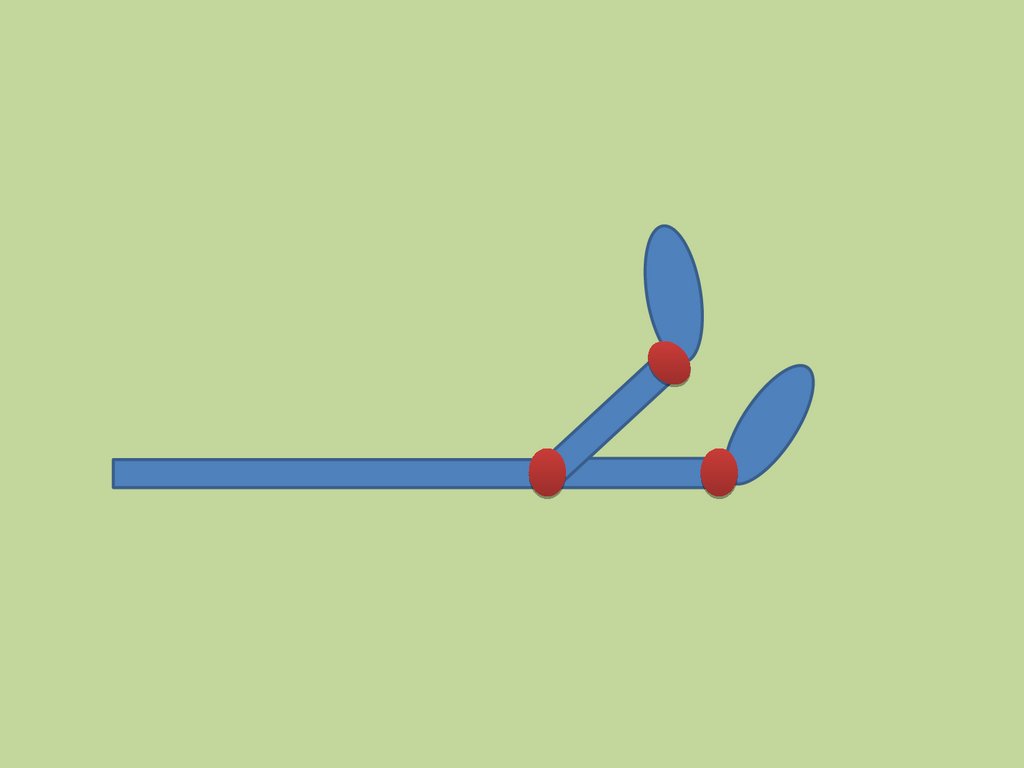
Миозиновые нити(толстые нити)
Каждая миозиновая нить состоит из
300–400 молекул миозина.
16.
Строение молекулы миозинаМиозин — гексaмер (две тяжёлые и четыре лёгкие цепи).
Области присоединяющиеся к
актиновой нити.
Области присоединения АТФ.
Тяжёлые цепи — две
спирально закрученные
полипептидные нити,
несущие на своих концах
глобулярные головки.
В области головок с
тяжёлыми цепями
ассоциированы лёгкие
цепи.
17.
шарнирные участкисубфрагмент S2
субфрагмент S1
ЛЁГКИЙ МЕРОМИОЗИН
ТЯЖЕЛЫЙ МЕРОМИОЗИН
18.
19.
20.
21.
22.
23.
24.
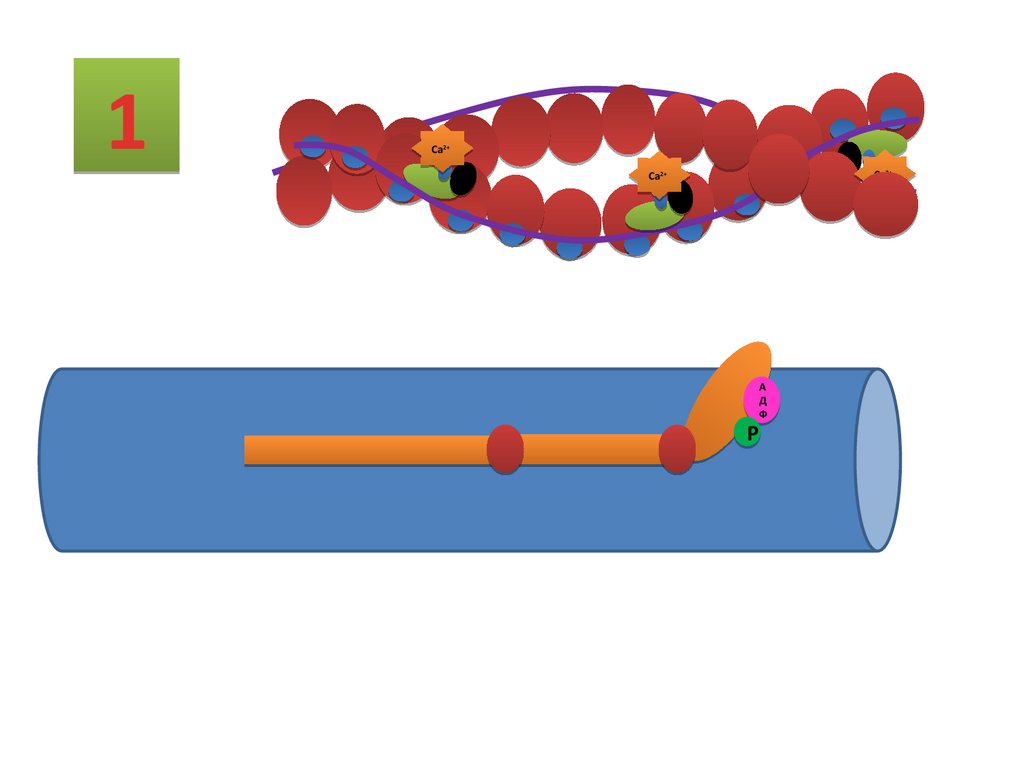
Строение актиновых нитейтропомиозин
тропонин
G – актин мономер
Комплекс регуляторных белков
F – актин полимер
Функциональная актиновая нить
25.
тропонинтропомиозин
G - актин
26.
Последовательность процессов примышечном сокрашении
Деполяризация постсинаптической мембраны и генерация ПД.
Распространение ПД по плазмолемме МВ .
Передача сигнала в триадах на саркоплазматический ретикулум.
Выброс Ca2+ из саркоплазматического ретикулума.
Связывание Ca2+ тропонином С тонких нитей.
Взаимодействие тонких и толстых нитей (формирование мостиков), появление тянущего усилия и
скольжение нитей относительно друг друга.
Цикл взаимодействия нитей.
Укорочение саркомеров и сокращение МВ.
Расслабление.
27.
28.
12
Т-трубочка
ПД
ПД
саркоплазматический ретикулюм
ПД
3
2+
Са2+
2+
Са2+
рецепторы
дигидропиридина
Рецепторы рианодина
Потенциалозависимый
кальциевый канал
АТФ
4
АДФ
2+
Са2+
Концентрация кальция в цитоплазме
29.
тропомиозинзин
мио
Область
связывания
миозина
Область
связывания
актина
Са 2+< 10-9 M
тропонин
Са 2+ не связан с
тропонином.
G - актин
G - актин
Взаимодействие
актина с миозином
заблокировано.
н
миози
2+
Са
Са2+
Са 2+< 10-5 M
Са 2+ связан с
тропонином.
з ин
о
и
м
5
G - актин
Взаимодействие
актина с миозином
разблокировано.
G - актин
н
миози
6
2+
Са
Са2+
30.
Модель мостика Хаксли-Симмонса1
2
3
4
1
2
3
4
31.
Циклвзаимодействия
нитей
32.
АА
Д
Д
Ф
Ф
Р
33.
12+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
А
А
Д
Д
Ф
Ф
Р
34.
12+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
А
А
Д
Д
Ф
Ф
Р
А
А
Д
Д
Ф
Ф
Р
35.
22+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
А
А
Д
Д
Ф
Ф
Р
36.
32+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
А
А
Д
Д
Ф
Ф
2+
Са
Са2+
37.
42+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
А
А
Д
Д
Ф
Ф
АТФ
АТФ
38.
52+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
АТФ
АТФ
АТФ
АТФ
АТФ
АТФ
39.
62+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
А
А
Д
АТФ
Д
АТФ
Ф
Ф
Р
40.
17
2+
Са
Са2+
2+
Са
Са2+
2+
Са
Са2+
А
А
Д
Д
Ф
Ф
Р
А
А
Д
Д
Ф
Ф
Р
41.
Цикл взаимодействия нитей42.
43.
Укорочение саркомеров и сокращение МВ.44.
45.
46.
47.
Общеенапряжение
5
сил
а
4
3
2
1
0
0
длина
время
7
8
длина
9
пассивное
ативное
9
8
7
ативное
6
0
1
2
3
время
4
5
пассивное
Общее
напряжение