





 перенос глюкозы возможен и против градиента концентрации.")

















")

")
















 biology
biology chemistry
chemistrySimilar presentations:










Обмен углеводов: переваривание углеводов. Гликолиз. Глюконеогенез
1. ОБМЕН УГЛЕВОДОВ:
Переваривание углеводов.Гликолиз. Глюконеогенез
1
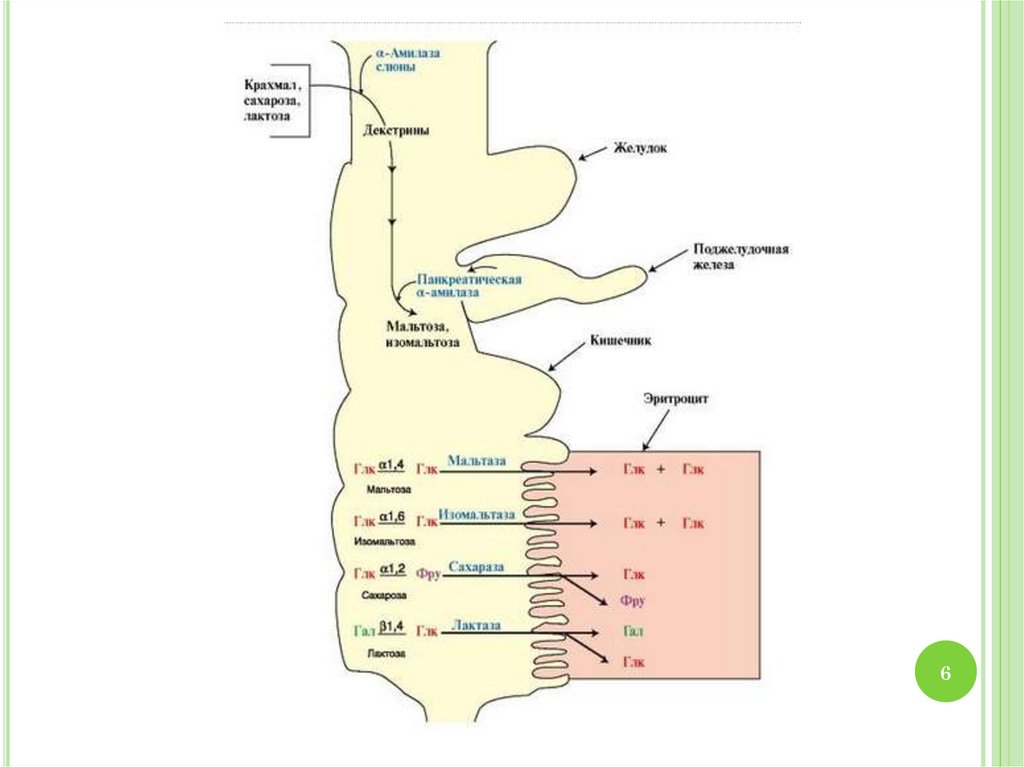
2. Переваривание углеводов: ротовая полость
Амилазаслюны расщепляет α1,4гликозидные связи в крахмале и
гликогене.
В ротовой полости происходит лишь
частичное переваривание крахмала, так
как действие фермента на эти углеводы
кратковременно.
Основными продуктами переваривания
крахмала в ротовой области являются
декстрины.
2
3. Желудок
Желудочныйсок не содержит ферментов,
расщепляющих пищевые углеводы.
Амилаза слюны инактивируется в желудке, так
как оптимальное значение рН для ее
активности составляет 6,7, а рН желудочного
сока равно ~ 2.
Лишь внутри пищевого комка этот фермент
продолжает действовать некоторое время.
3
4. 12-перстная кишка
Переваривание нерасщепленного или частичнорасщепленного крахмала происходит в кишечнике.
В 12-перстной кишке pH желудочного содержимого
нейтрализуется бикарбонатами, содержащимися в
секрете поджелудочной железы, и создается
оптимальное значение pH 7,5-8 для действия
панкреатической α-амилазы.
α-Амилаза поджелудочной железы гидролизует
декстрины и оставшиеся нерасщепленными
молекулы крахмала и гликогена, расщепляя α1,4гликозидные связи
α-1,6-гликозидные связи расщепляют ферменты
амило- и олиго-1,6 гликозидазы из поджелудочной
4
железы
5. Тонкий кишечник
Мальтоза и изомальтоза вместе с другими пищевымидисахаридами - сахарозой и лактозой - гидролизуются
специфическими гликозидазами на поверхности
клеток тонкого кишечника (возможно и внутри клеток) до
соответствующих мономеров.
Сахараза расщепляет сахарозу
Мальтаза расщепляет мальтозу
Изомальтаза расщепляет изомальтозу
Лактаза расщепляет лактозу
Трегалаза расщепляет трегалозу (грибной сахар,
дисахарид, содержащий 2 остатка глюкозы, соединенных
α-1,1 гликозидной связью)
5
6.
67. Всасываются в кишечнике только моносахара
Их перенос в клетки слизистой оболочкикишечника (энтероциты) может происходить :
1) способом пассивной диффузии
по градиенту концентрации
из просвета кишечника (где концентрация
сахара после еды выше)
в клетки кишечника (где она ниже).
7
8. 2) перенос глюкозы возможен и против градиента концентрации.
Это активный транспорт: идёт с затратой энергии,используются специальные белки-переносчики
(GLUT).
Глюкоза
Белок–переносчик + АТФ
8
9.
ДАЛЬНЕЙШИЕ ПРЕВРАЩЕНИЯ САХАРОВ ВОРГАНИЗМЕ
В
результате переваривания углеводов
пищи и инверсии других сахаров
основным моносахаридом оказывается
глюкоза.
Из кишечника она поступает в кровь
воротой вены и далее с током крови в
печень.
9
10. Пищевые волокна
Растительнаяпища богата пищевыми волокнами
(целлюлоза, гемицеллюлоза, пектины, лигнины),
которые не перевариваются ферментами желудочнокишечного тракта.
Некоторые из них (целлюлоза) перевариваются
ферментами бактерий толстого кишечника:
целлюлаза расщепляет целлюлозу до целлобиозы;
целлобиаза расщепляет целлобиозу на две молекулы
β-D-глюкозы.
β-D-Глюкоза затем превращается в органические
кислоты: молочную, пировиноградную и др.
10
11. Роль пищевых волокон
Пищевые волокна выполняют ряд важных функций ворганизме:
1) необходимы для нормальной перистальтики
кишечника;
2) адсорбируют вещества, продуцируемые
кишечными бактериями (например, токсины);
3) предупреждают развитие опухолевых процессов в
кишечнике;
4) снижают всасывание холестерола из кишечника и
способствуют выведению желчных кислот из
организма.
11
12. Основные источники глюкозы
1)2)
3)
пища;
распад гликогена;
синтез глюкозы из неуглеводных
предшественников (глюконеогенез).
12
13. Основные пути использования глюкозы
1)2)
3)
4)
распад глюкозы с целью получения
энергии (аэробный и анаэробный
гликолиз);
синтез гликогена;
пентозофосфатный путь распада для
получения других моносахаридов и
восстановленного НАДФН;
синтез других соединений (жирные
кислоты, аминокислоты,
гетерополисахариды и др.).
13
14. Источники и пути расходования глюкозы
1415. ГЛИКОЛИЗ
Гликолиз (Greek glucose – сахар, lysis –разрушение) – последовательность реакций
превращения глюкозы до пирувата.
В процессе гликолиза часть свободной энергии
распада глюкозы превращается в АТФ и
НАДН.
Суммарная реакция гликолиза:
Глюкоза + 2 Рн + 2 АДФ + 2 НАД+→
2 пируват + 2 АТФ + 2 НАДН + 2Н+ + 2 Н2О
15
16. ОСНОВНЫЕ ПРИЗНАКИ ГЛИКОЛИЗА
1. Протекает во всех клетках организма. Ферментылокализованы в цитозоле клеток.
2. В анаэробных условиях (в отсутствии
кислорода) образующийся пируват превращается
в лактат.
3. В аэробных условиях (присутствие кислорода)
пируват окисляется до ацетил-КоА, который
затем вступает в ЦТК, где его ацетильная группа
окисляется до СО2 и Н2О.
4. В клетках, лишенных митохондрий
(эритроциты, роговая оболочка глаза, хрусталик
глаза) гликолиз является главным источником
образования АТФ.
5. В клетках, имеющих митохондрии, гликолиз
является этапом аэробного окисления
углеводов.
16
17.
4. Наиболее активно протекает в мозге,поскольку окисление глюкозы
является основным источником
энергии для нервной ткани.
5. В анаэробных условиях гликолиз –
единственный процесс в организме
животных, растений и многих
микроорганизмов, приводящий к
образованию АТФ;
6. В организме человека и животных
гликолиз позволяет поддерживать
интенсивную работу скелетной мышцы в
условиях недостатка кислорода.
7. Промежуточные продукты гликолиза
17
18. АЭРОБНЫЙ И АНАЭРОБНЫЙ РАСПАД ГЛЮКОЗЫ
1819. АЭРОБНЫЙ И АНАЭРОБНЫЙ РАСПАД ГЛЮКОЗЫ
Анаэробный гликолиз включает в себя реакцииспецифического пути распада глюкозы до
пирувата и восстановление пирувата в
лактат. АТФ при анаэробном гликолизе образуется
только путем субстратного фосфорилирования.
Аэробный распад глюкозы до конечных
продуктов (СО2 и Н2О) включает в себя реакции
аэробного гликолиза и последующее окисление
пирувата в общем пути катаболизма.
Таким образом, аэробный распад глюкозы - это
процесс полного ее окисления до СО2 и Н2О, а
аэробный гликолиз - часть аэробного распада
глюкозы.
19
20. МЕХАНИЗМ АЭРОБНОГО ГЛИКОЛИЗА
2021.
2122.
2223.
2324.
2425.
2526.
2627.
2728.
2829.
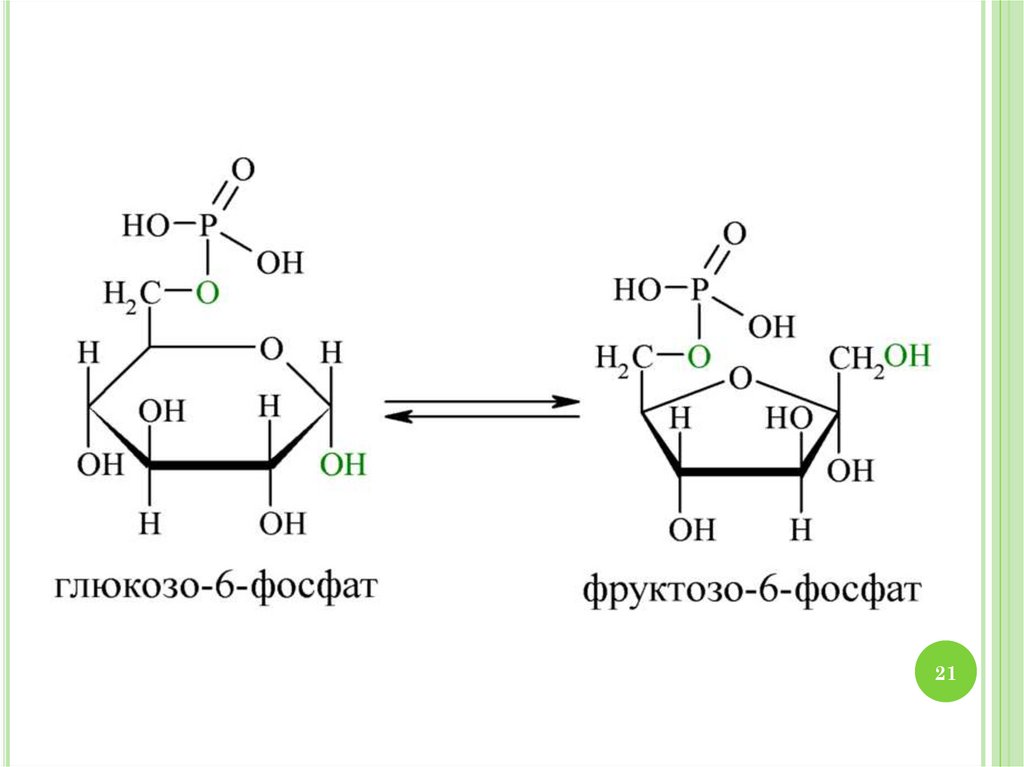
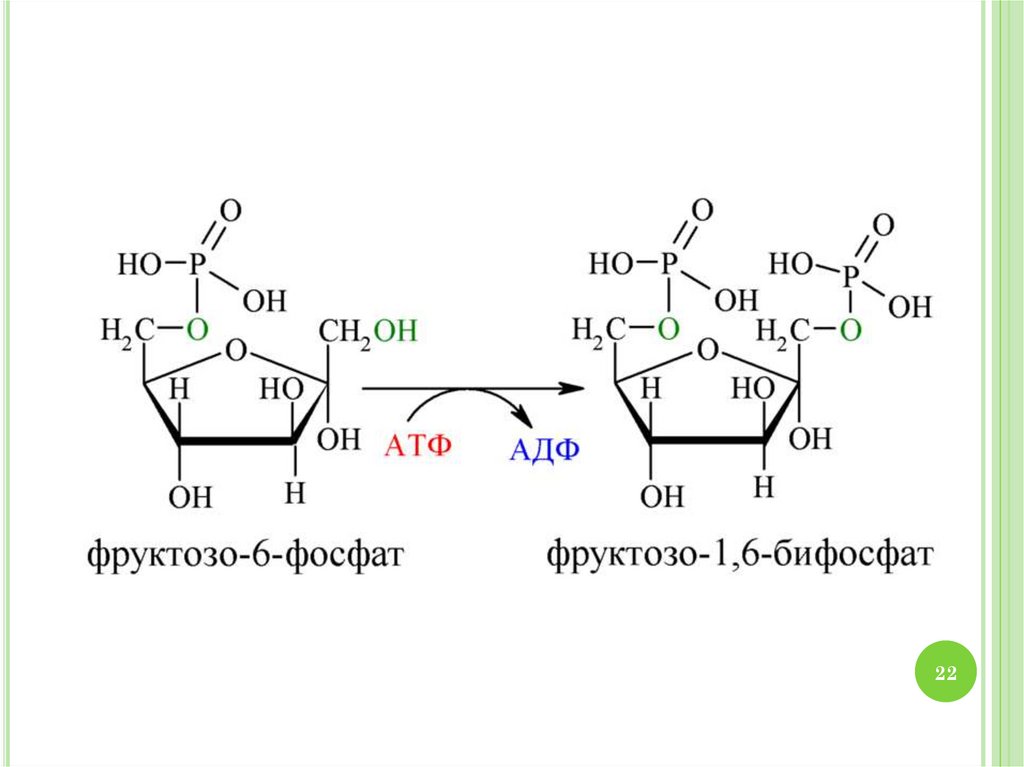
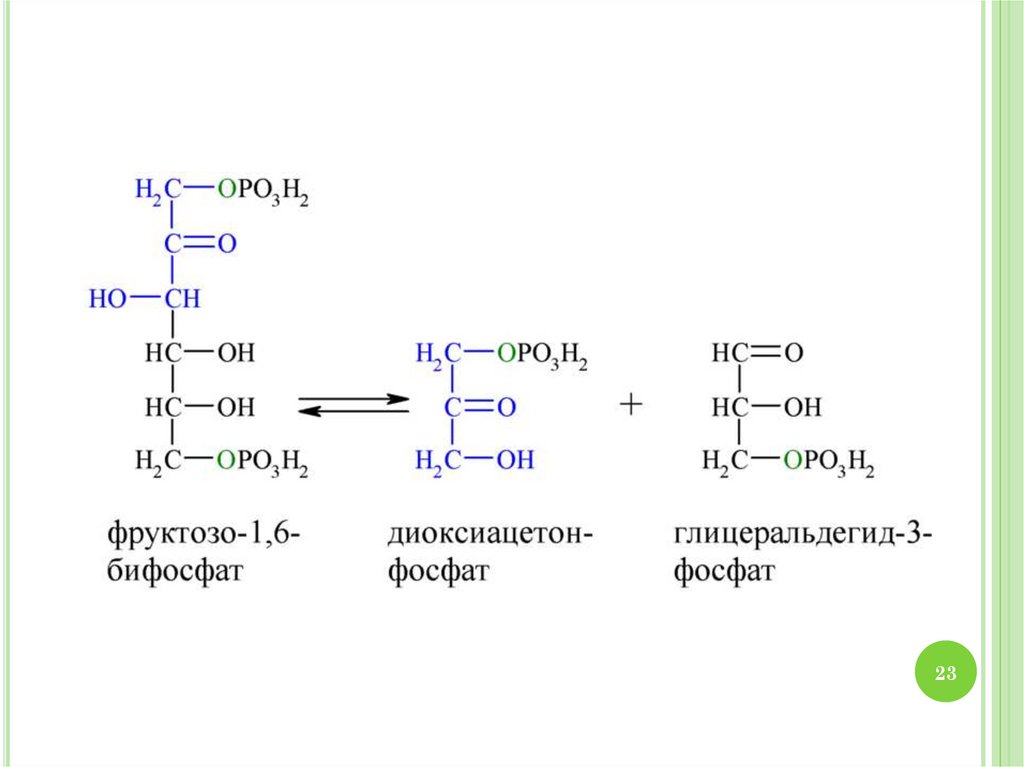
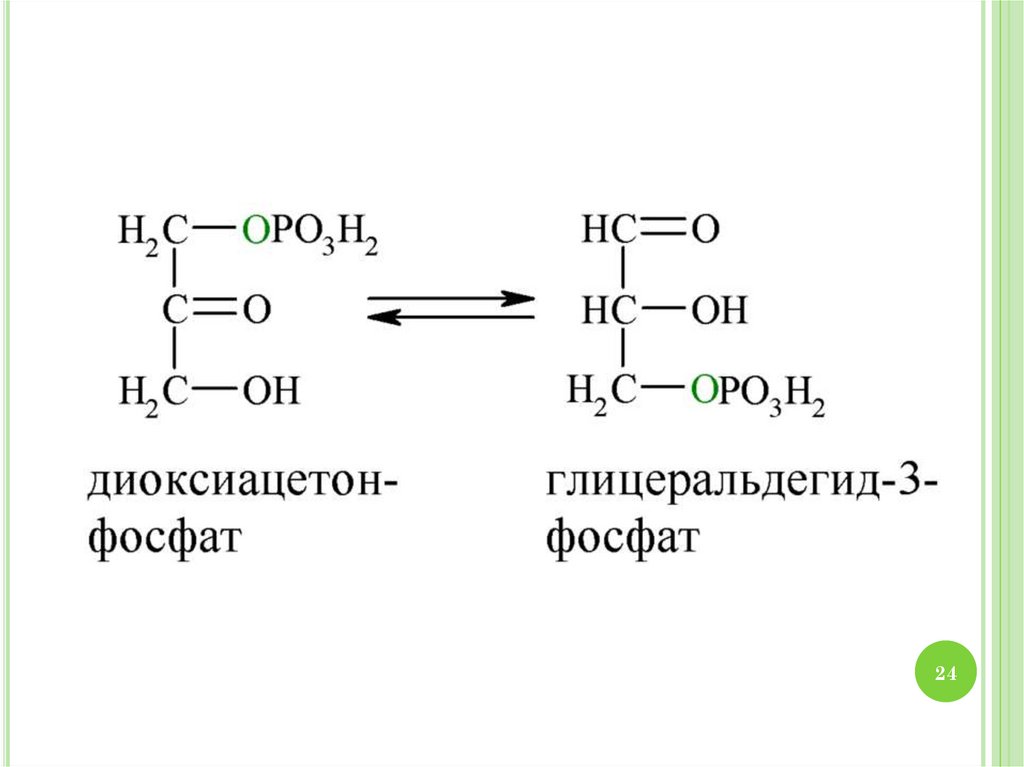
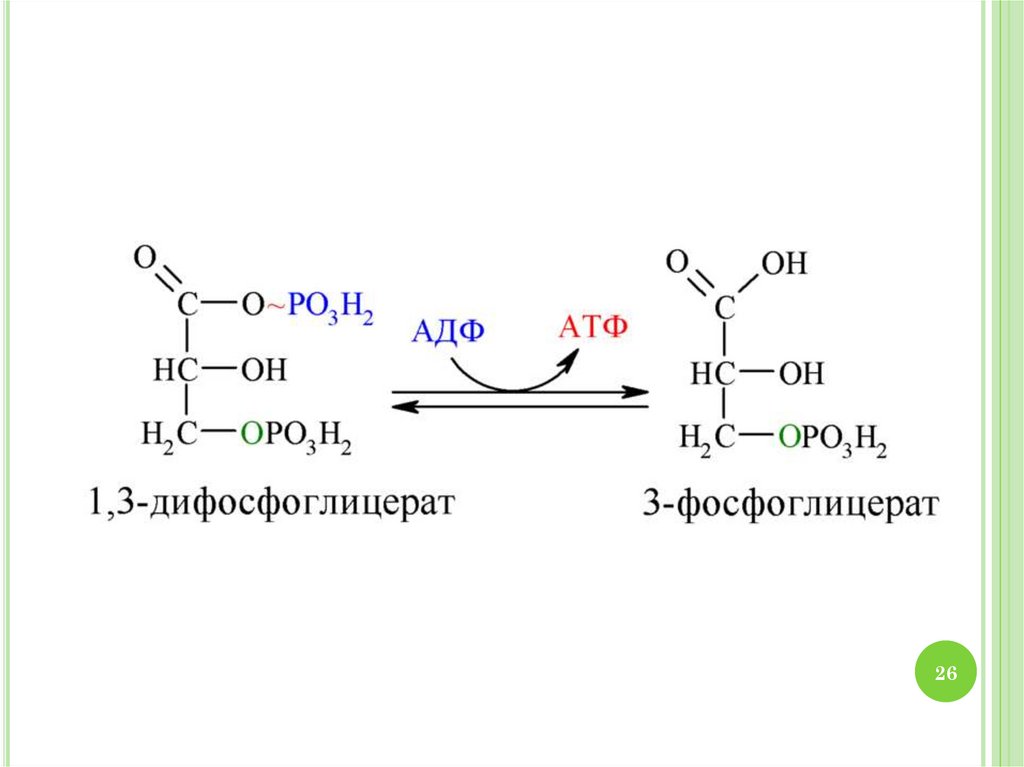
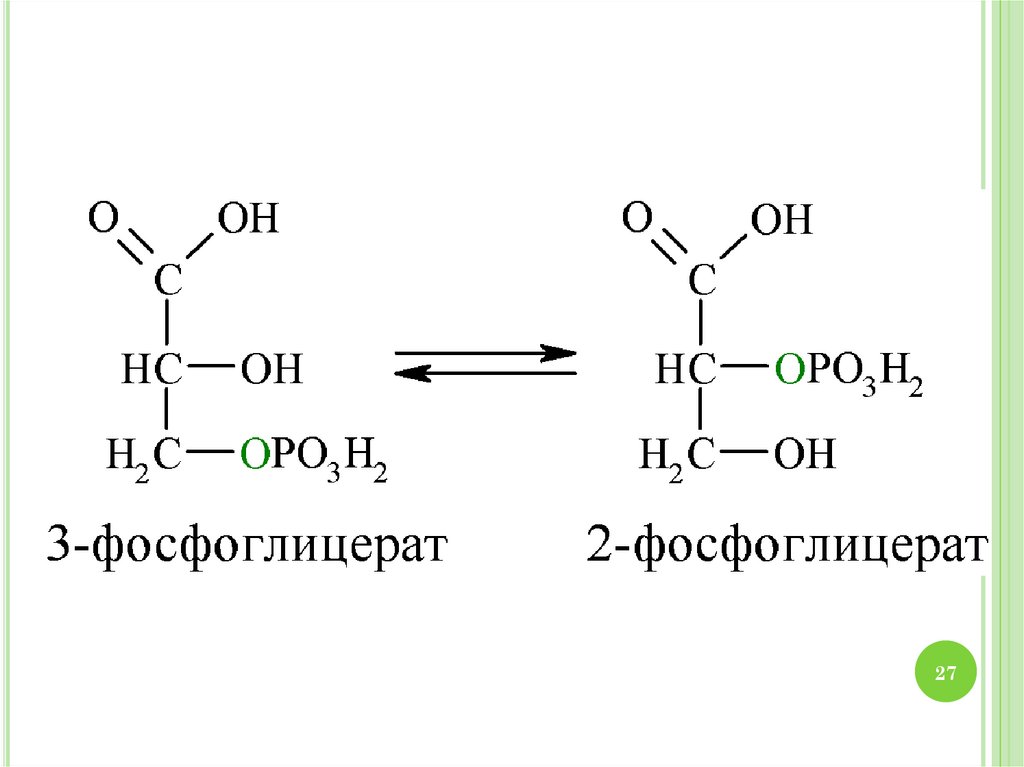
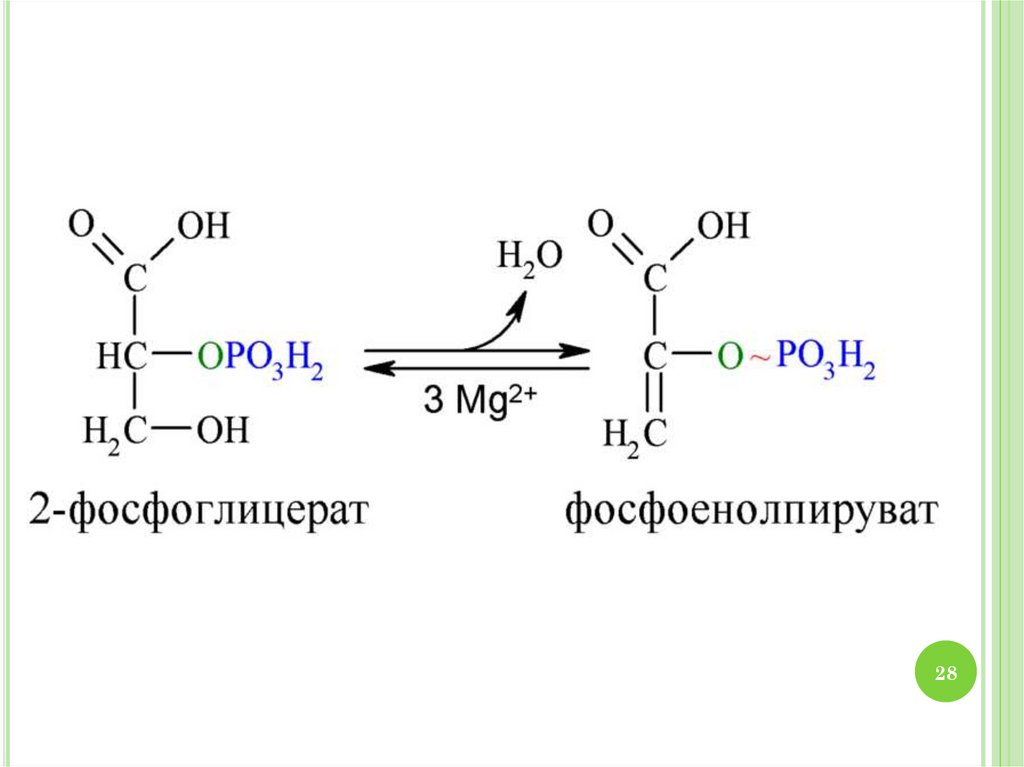
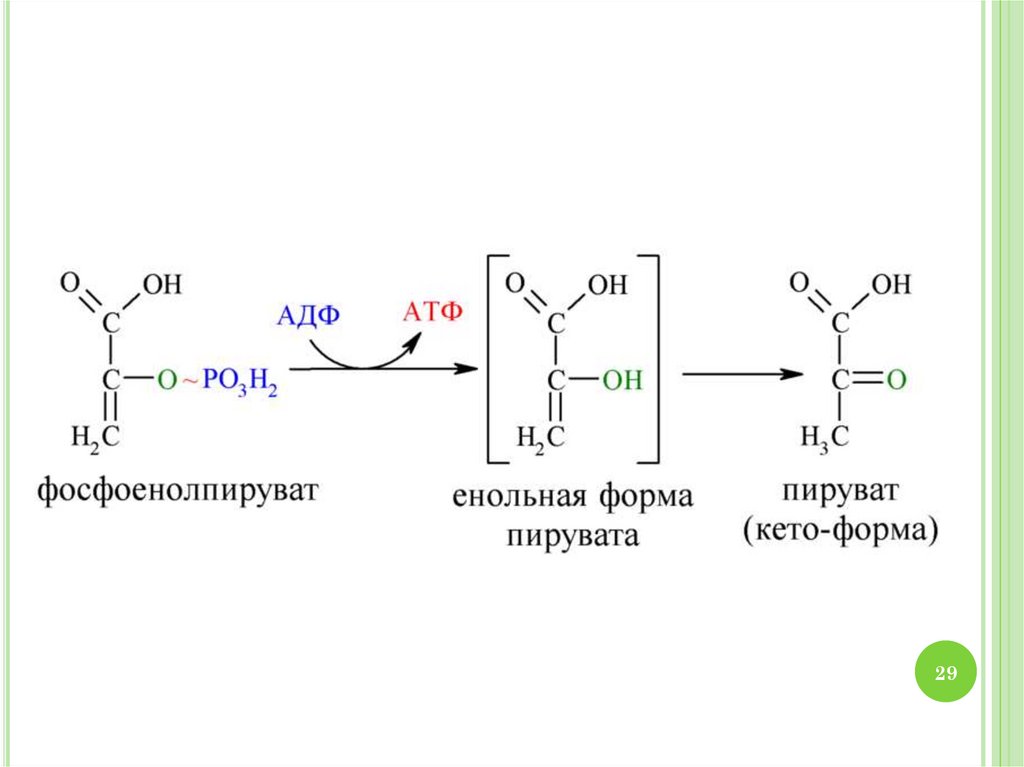
2930. МЕХАНИЗМ ГЛИКОЛИЗА
1.2.
3.
4.
30
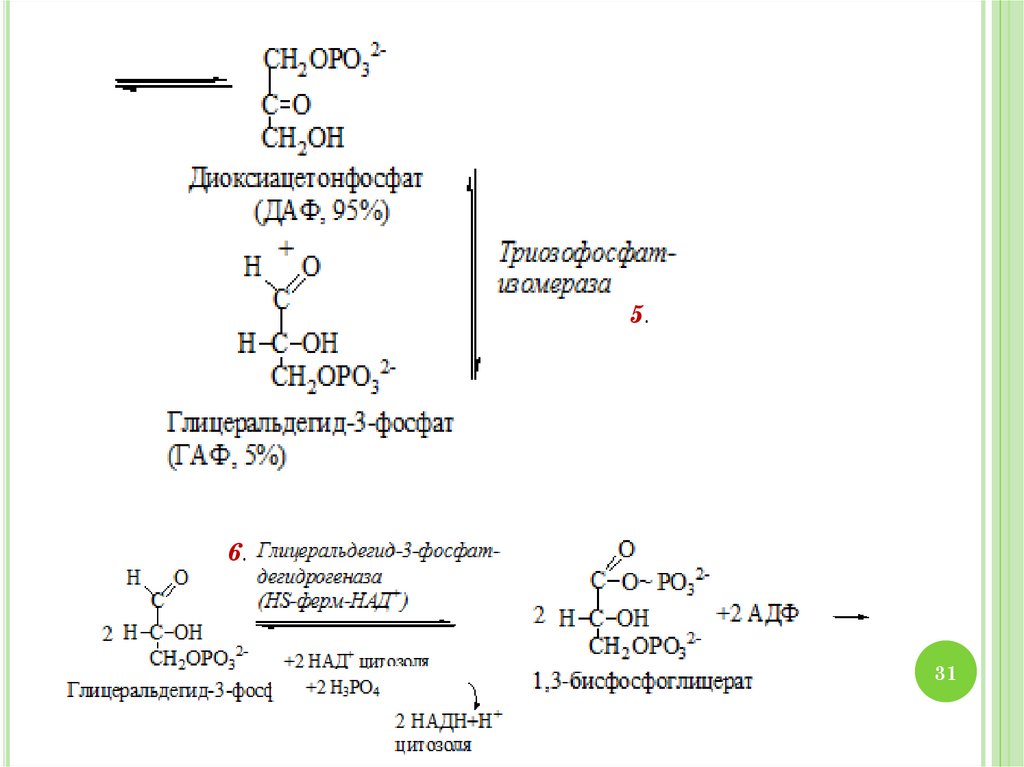
31.
5.6.
31
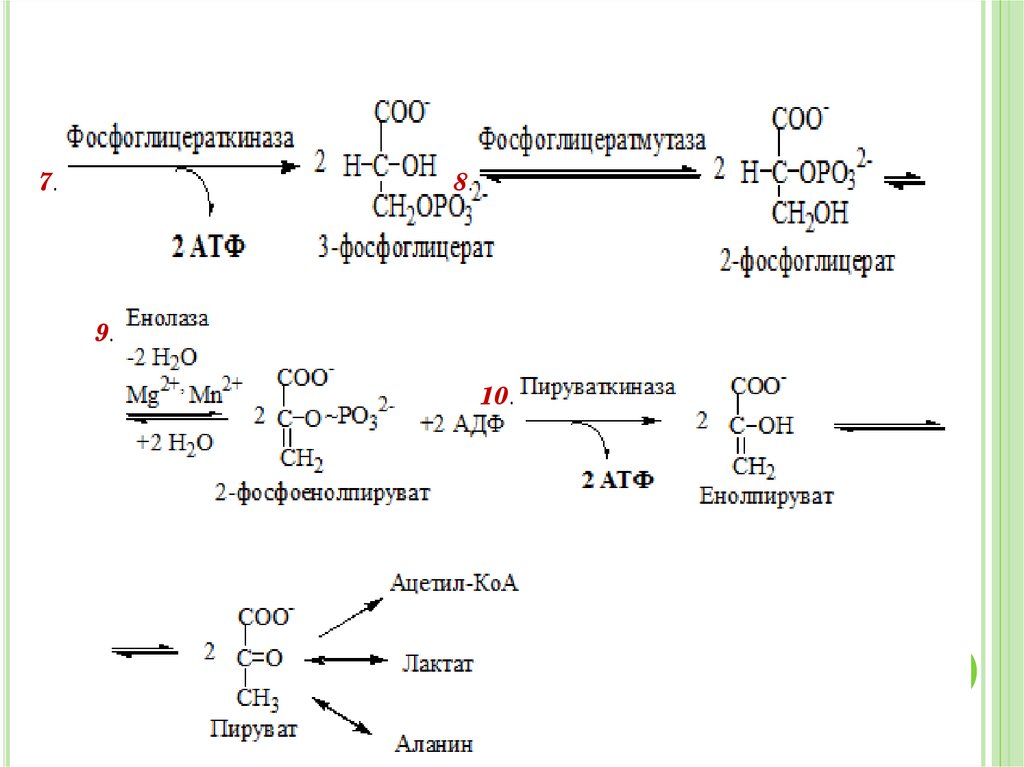
32.
7.8.
9.
10.
32
33. РЕГУЛЯЦИЯ ГЛИКОЛИЗА
1.2.
3.
4.
Первый фермент гликолиза – гексокиназа
ингибируется продуктом реакции (глюкозо-6фосфат). Высокая внутриклеточная концентрация
глюкозо-6-фосфата сигнализирует о том, что не
требуется поступление глюкозы в клетку для
получения энергии или запасания в виде
гликогена.
Высокий уровень АТФ и цитрата
аллостерически ингибирует активность
фосфофруктокиназы в печени
АМФ отменяет ингибиторный эффект АТФ и
активность фосфофруктокиназы повышается, если
отношение АТФ/АМФ уменьшается.
Иными словами, гликолиз активируется при
уменьшении энергетического заряда клетки.
33
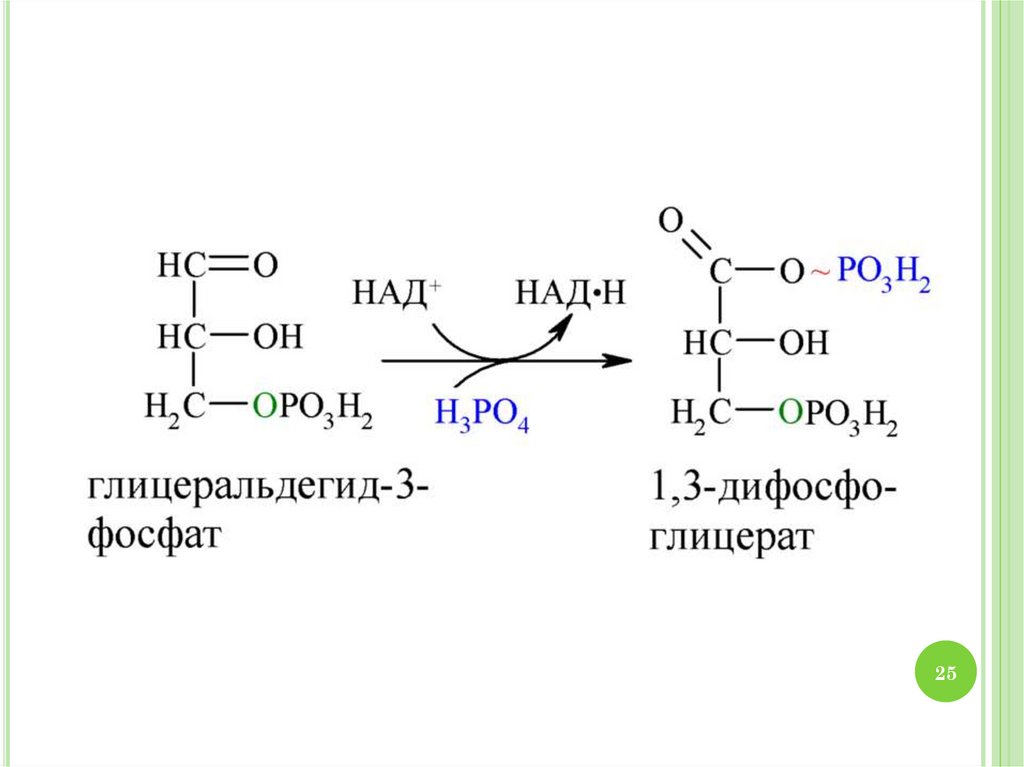
34. МАЛАТНЫЙ ЧЕЛНОЧНЫЙ МЕХАНИЗМ
На стадии окисления глицеральдегид-3-фосфата в1,3-бисфосфоглицериновую кислоту образуется
восстановленный цитозольный НАДН+Н+.
Перенос восстановительных эквивалентов от НАДН+Н+
в митохондрии происходит с помощью малатного
челночного механизма
34
35. ЭНЕРГЕТИЧЕСКИЙ БАЛАНС АЭРОБНОГО ОКИСЛЕНИЯ ГЛЮКОЗЫ
1. В специфическом пути распада глюкозы образуется 2молекулы пирувата, 2 АТФ (субстратное
фосфорилирование) и 2 молекулы НАДН+Н+.
2. Окислительное декарбоксилирование каждой
молекулы пирувата - 2,5 АТФ;
декарбоксилирование 2-х молекул пирувата дает 5
молекул АТФ.
3. В результате окисления ацетильной группы
ацетил-КоА в ЦТК и сопряженных ЦПЭ – 10 АТФ;
2 молекулы ацетил-КоА образуют 20 АТФ.
4. Малатный челночный механизм переносит
НАДН+Н+ в митохондрии – 2,5 АТФ; 2 НАДН+Н+
образуют 5 АТФ.
Итого: при распаде 1 молекулы глюкозы в аэробных
условиях образуется 32 молекулы АТФ.
35
36. ХАРАКТЕРИСТИКА ГЛЮКОНЕОГЕНЕЗА
Глюконеогенез - это процесс синтеза глюкозы извеществ неуглеводной природы - лактата,
глицерола, аминокислот, пропионовой кислоты.
Важнейшей функцией глюконеогенеза является
поддержание уровня глюкозы в крови в период
длительного голодания и интенсивных
физических нагрузок.
Постоянное поступление глюкозы в качестве
источника энергии особенно необходимо для
нервной ткани и эритроцитов.
При интенсивной физической работе синтез
глюкозы происходит из лактата и глицерола
При голодании – из глицерола, образующегося при
распаде жиров и аминокислот, освобождающихся в
процессе катаболизма белков мышц.
36
36
37. ХАРАКТЕРИСТИКА ГЛЮКОНЕОГЕНЕЗА (ПРОДОЛЖЕНИЕ)
Процесс протекает главным образом в печени именее интенсивно - в почках, а также в слизистой
оболочке кишечника.
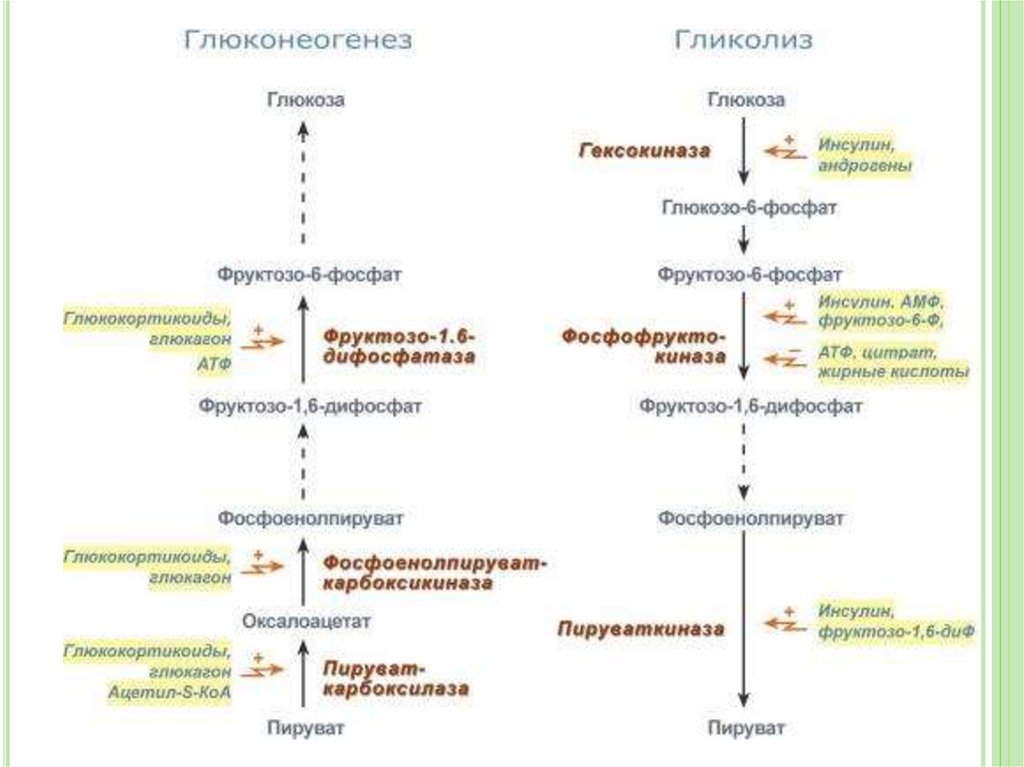
Большинство реакций гликолиза и глюконеогенеза
являются обратимыми и катализируются одними и
теми же ферментами.
Существуют три необратимые реакции гликолиза –
реакции, катализируемые гексокиназой (1-я реакция),
фосфофруктокиназой (3-я реакция) и пируваткиназой
(10-я реакция)
В процессе глюконеогенеза для обращения этих трех
необратимых этапов используются другие ферменты
37
37
38.
3838
39. ГЛЮКОНЕОГЕНЕЗ ИЗ ЛАКТАТА
1. При интенсивной мышечной работе образуетсялактат, который током крови поступает в печень.
В цитозоле под действием лактатдегидрогеназы
лактат окисляется в пируват, который поступает в
митохондрии.
39
39
40. СИНТЕЗ ФОСФОЕНОЛПИРУВАТА (1-Я НЕОБРАТИМАЯ РЕАКЦИЯ ГЛИКОЛИЗА)
Прямой синтез фосфоенолпирувата из пируватаневозможен из-за необратимости пируваткиназной
реакции, поэтому используется обходной путь из
нескольких ферментов.
Вначале пируват поступает в митохондрии и при
участии биотинзависимой пируваткарбоксилазы,
СО2 и АТФ карбоксилируется с образованием
оксалоацетата. Это ключевой момент для
энергетического преодоления пируваткиназной реакции
в обходных реакциях.
40
40
41.
Для транспорта из митохондрий в цитозольоксалоацетат восстанавливается в малат с
участием фермента НАД-зависимой
малатдегидрогеназы.
41
41
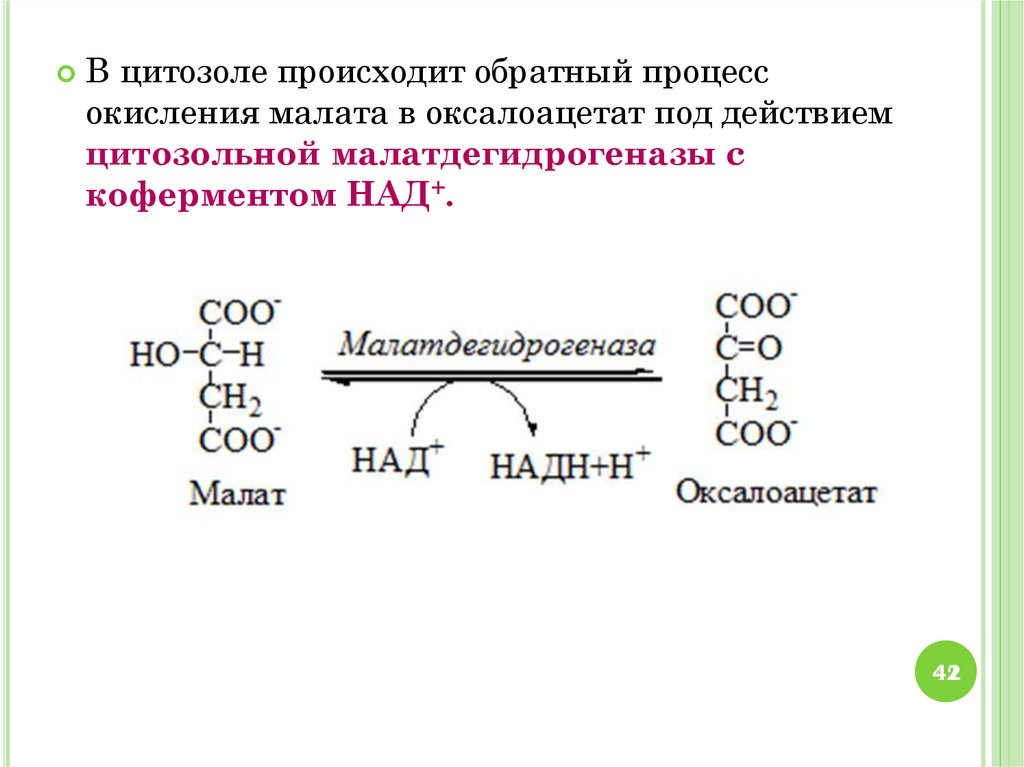
42.
В цитозоле происходит обратный процессокисления малата в оксалоацетат под действием
цитозольной малатдегидрогеназы с
коферментом НАД+.
42
42
43.
Оксалоацетат в результатедекарбоксилирования и фосфорилирования
под влиянием фермента
фосфоенолпируваткарбоксикиназы
превращается в фосфоенолпируват. Донором
фосфатного остатка служит ГТФ. На этом
завершается путь обхода необратимой
пируваткиназной реакции
43
43
44. 2-Я И 3-Я НЕОБРАТИМЫЕ РЕАКЦИИ ГЛИКОЛИЗА
Фосфоенолпируват в результате рядаобратимых реакций гликолиза превращается
во фруктозо-1,6-бисфосфат.
Превращение фруктозо-1,6-бисфосфата во
фруктозо-6-фосфат катализируется
специфической фосфатазой – фруктозо-1,6бисфосфатазой (обход необратимой
фосфофруктокиназной реакции).
В последующей обратимой стадии биосинтеза
глюкозы фруктозо-6-фосфат превращается в
глюкозо-6-фосфат, который дефосфорилируется
под влиянием фермента глюкозо-6-фосфатазы
(обход необратимой гексокиназной реакции).
44
44
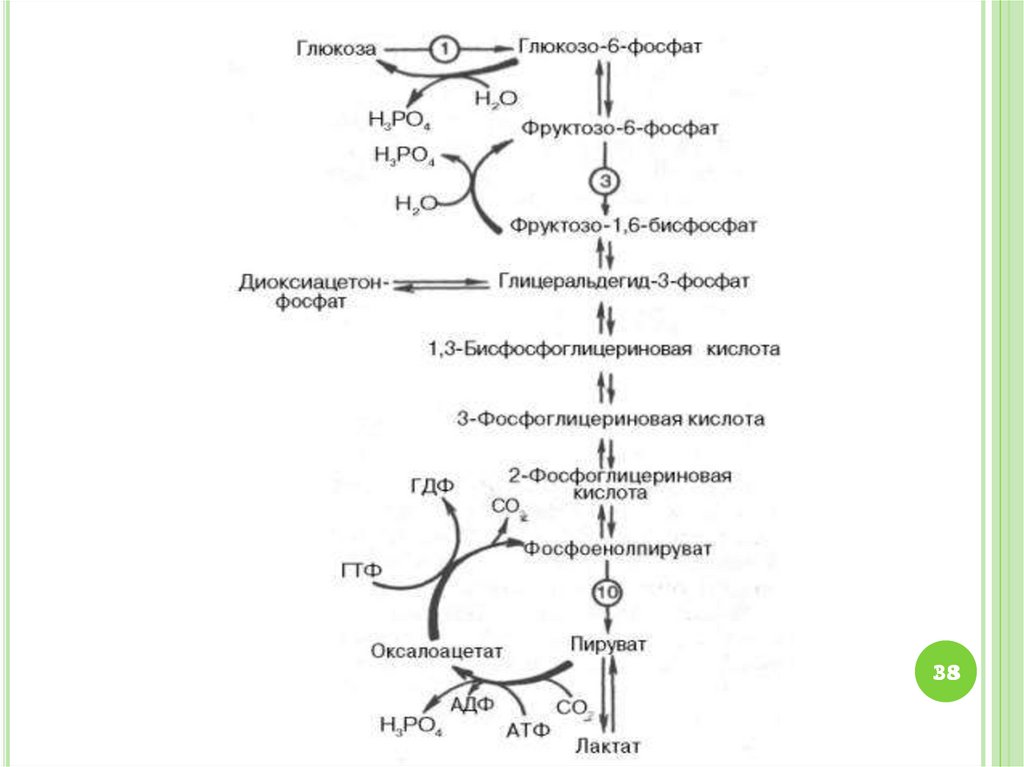
45. СХЕМА ГЛИКОЛИЗА И ГЛЮКОНЕОГЕНЕЗА
4545
46.
4646
47. ВЗАИМОСВЯЗЬ ГЛИКОЛИЗА И ГЛЮКОНЕОГЕНЕЗА
1. Основным субстратом для глюконеогензаявляется лактат, образованный активной
скелетной мышцей. Плазматическая мембрана
обладает высокой проницаемостью для лактата.
2. Поступив в кровь, лактат переносится в печень,
где в цитозоле окисляется в пируват.
3. Пируват затем превращается в глюкозу по
пути глюконеогенеза.
4. Глюкоза поступает далее в кровь и поглощается
скелетными мышцами. Эти превращения
составляют цикл Кори.
47
47
48. ЦИКЛ КОРИ
4848
49. ХАРАКТЕРИСТИКА ПЕНТОЗОФОСФАТНОГО ПУТИ
Пентозофосфатный путь распада глюкозы (ПФП) называетсятакже гексозомонофосфатным шунтом или фосфоглюконатным
путем.
Этот альтернативный гликолизу и ЦТК путь окисления
глюкозы был описан в 50-х годах ХХ века Ф.Дикенсом,
Б.Хорекером, Ф.Липманном и Е.Рэкером.
Ферменты пентозофосфатного пути локализованы в цитозоле.
Наиболее активно ПФП протекает в почках, печени,
жировой ткани, коре надпочечников, эритроцитах,
лактирующей молочной железе. В большинстве из этих
тканей протекает процесс биосинтеза жирных кислот и
стероидов, что требует НАДФН.
Выделяют две фазы ПФП: окислительную и
неокислительную
49
49
50. СХЕМА ПФП
5050
51. ОКИСЛИТЕЛЬНЫЙ ЭТАП ПФП
5151
52. НЕОКИСЛИТЕЛЬНЫЙ ЭТАП ПФП
5252
53.
I-я стадия окислительная:1) генерируется НАДФН + Н + –
восстановительные эквиваленты, которые
используются как донор Н + и ẽ при
восстановительных биосинтезах.
2) синтезируется рибоза –
нужна для синтеза КоА, НАД, ФАД, АТФ,
нуклеотидов ДНК и РНК.
53
53
54. 2-я стадия НЕОКИСЛИТЕЛЬНАЯ :
1) идёт перенос (класс - трансферазы)2-углеродного компонента
-СО-СН2ОН
транскетолазой
3-углеродного компонента -НОСН-СО-СН2ОН
трансальдолазой
2) работает эпимераза (класс - изомеразы)
Образуются разные сахара
с количеством углеродных атомов от 3 до 7.
Следовательно, этот этап ПФП обеспечивает
взаимопревращение 3-, 4-, 5-, 6-, 7-углеродных
сахаров (система структурной перестройки).
54
54
55.
Для синтезов клеткам нужно гораздо больше НАДФН,чем требуется рибозы для включения
в нуклеотиды и нуклеиновые кислоты.
Поэтому роль II-й неокислительной
ветви ПФП:
1) Излишек рибозо-5-ф переключён на ресинтез
более нужного
глюкозо-6-Ф.
2) Через отдельные промежуточные продукты
(ГА-3-Ф, Ф-6-Ф) возможна связь с гликолизом.
55
55
56.
Из 6 молекул Г-6-Ф, поступающих в ПФП,1 молекула глюкозы сгорает (в ходе синтеза 12
НАДФН и отщепления 6 СО2) остальные 5 молекул
Г-6-Ф образуются снова.
Все процессы ПФП протекают в цитозоле клетки и
тесно связаны с синтезом жирных кислот,
холестерола, стероидных гормонов и т.д. (жировая
ткань, половые железы, надпочечники).
56
56
57. ФУНКЦИИ ПЕНТОЗОФОСФАТНОГО ПУТИ
1. Образование НАДФН+Н+ (50% потребности организма),необходимого 1) для биосинтеза жирных кислот,
холестерола; 2) для реакции детоксикации
(восстановление и окисление глутатиона,
функционирование цитохром Р-450 зависимых
монооксигеназ – микросомальное окисление); 3) для
восстановления железа (III) в метгемоглобине в железо
(II) нормального гемоглобина (эритроциты).
2. Синтез рибозо-5-фосфата, используемого для
образования 5-фосфорибозил-1-пирофосфата, который
необходим для синтеза пуриновых нуклеотидов и
присоединения оротовой кислоты в процессе биосинтеза
пиримидиновых нуклеотидов.
3. Синтез углеводов с различным числом атомов углерода
(С3-С7).
4. У растений образование рибулозо-1,5-бисфосфата,
который используется как акцептор СО2 в темновой
стадии фотосинтеза.
57
57