




















 biology
biologySimilar presentations:










Организация хроматина. Лекция 4
1. Организация хроматина
2.
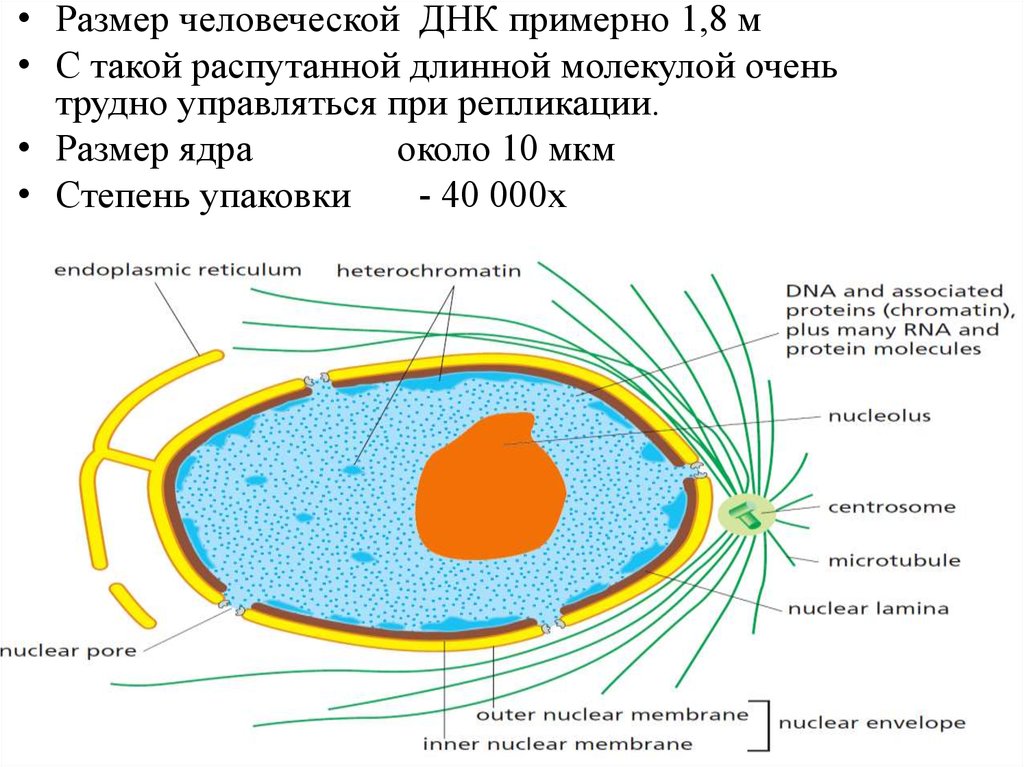
• Размер человеческой ДНК примерно 1,8 м• С такой распутанной длинной молекулой очень
трудно управляться при репликации.
• Размер ядра
около 10 мкм
• Степень упаковки
- 40 000х
3.
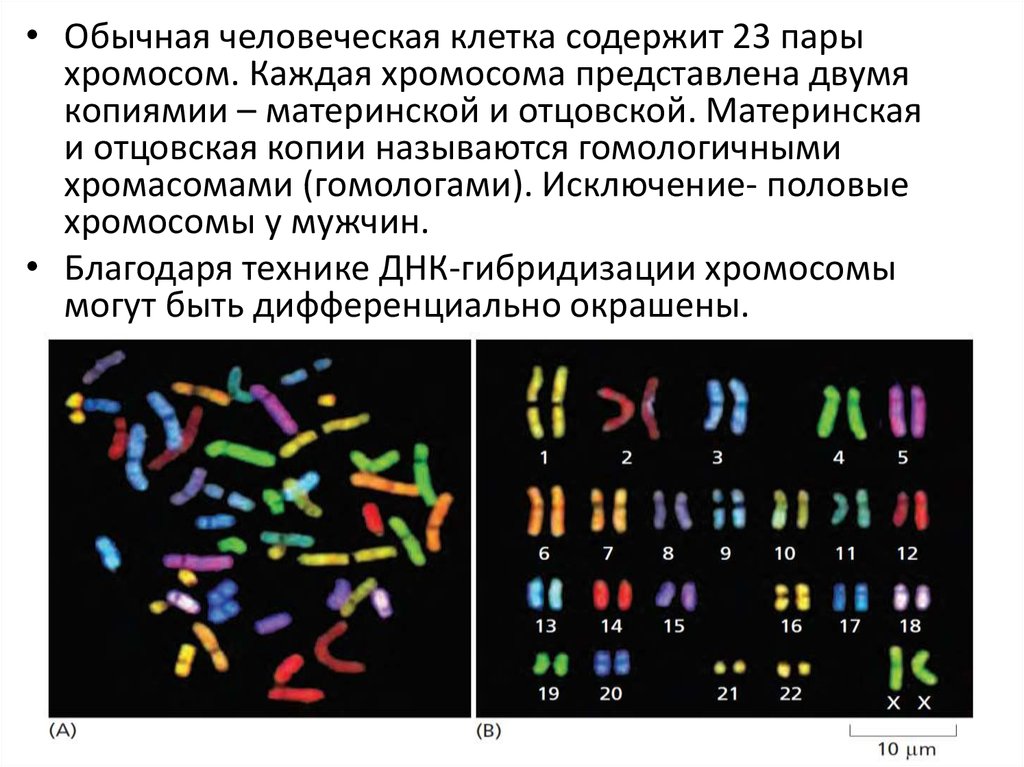
• Обычная человеческая клетка содержит 23 парыхромосом. Каждая хромосома представлена двумя
копиямии – материнской и отцовской. Материнская
и отцовская копии называются гомологичными
хромасомами (гомологами). Исключение- половые
хромосомы у мужчин.
• Благодаря технике ДНК-гибридизации хромосомы
могут быть дифференциально окрашены.
4. Распределение генов в S. cerevisiae по сравнению с человеком
5. Нуклеотдная последовательность человеческого генома может рассказать об его устройстве
6. Свойства генома человека
• Малая фракция кодирует структурнуюинформацию;
• Примерно половина генома состоит из
мобильных (повторяющихся) элементов;
• Большая средняя длиня гена (ок. 27 кB), при
том что для кодирования
среднестатсистического белка (ок 430 аа)
нужно всего ок 1300 B. В малых геномах в
генах практически нет интронов и размеры
их генов почти в 20 раз меньше
человеческого.
7.
• Помимо экзонов и интронов, в состав геноввходят регуляторные последовательности ДНК,
отвечающие за включение/выключение гена,
экспрессию его на должном уровне и только в
определенных типах клеток.
• В человеческом геноме кроме генов,
кодирующих белки (ок 21 тыс), имеется порядка
9 тыс генов, кодирующих различные РНК.
8.
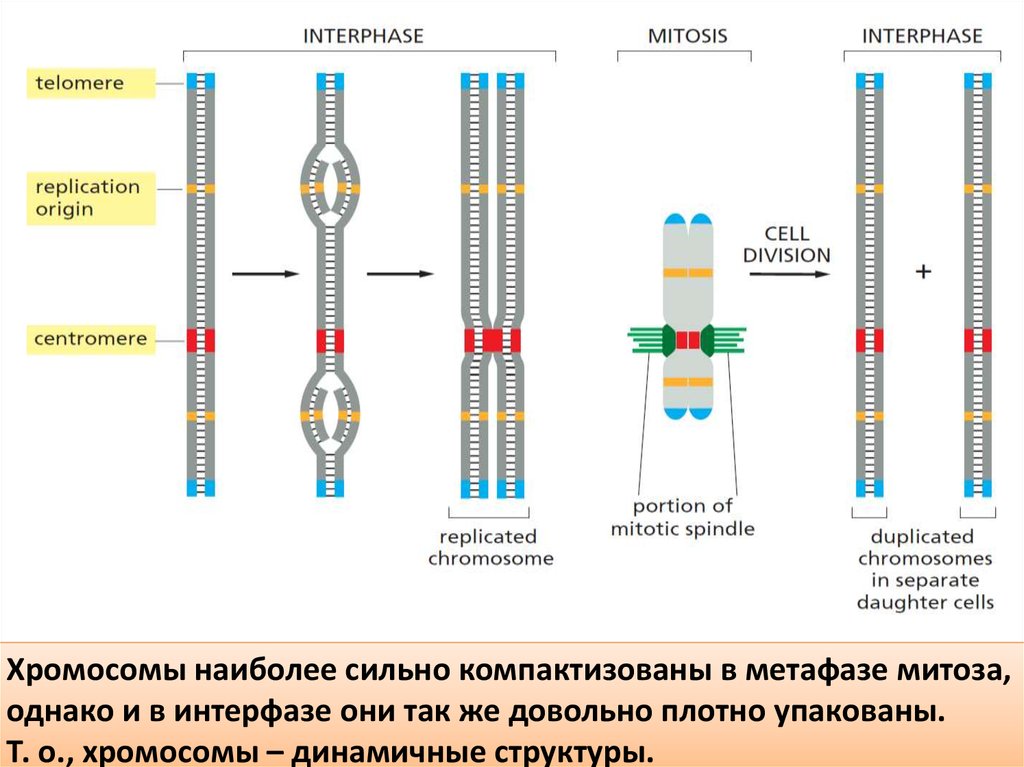
Хромосомы наиболее сильно компактизованы в метафазе митоза,однако и в интерфазе они так же довольно плотно упакованы.
Т. о., хромосомы – динамичные структуры.
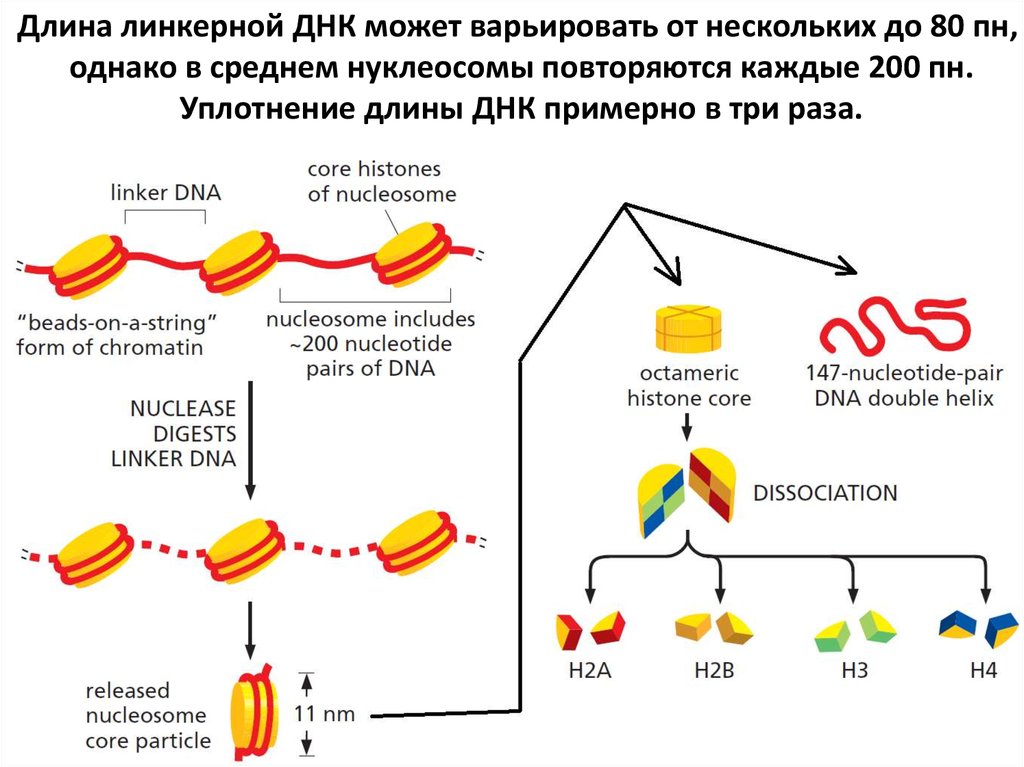
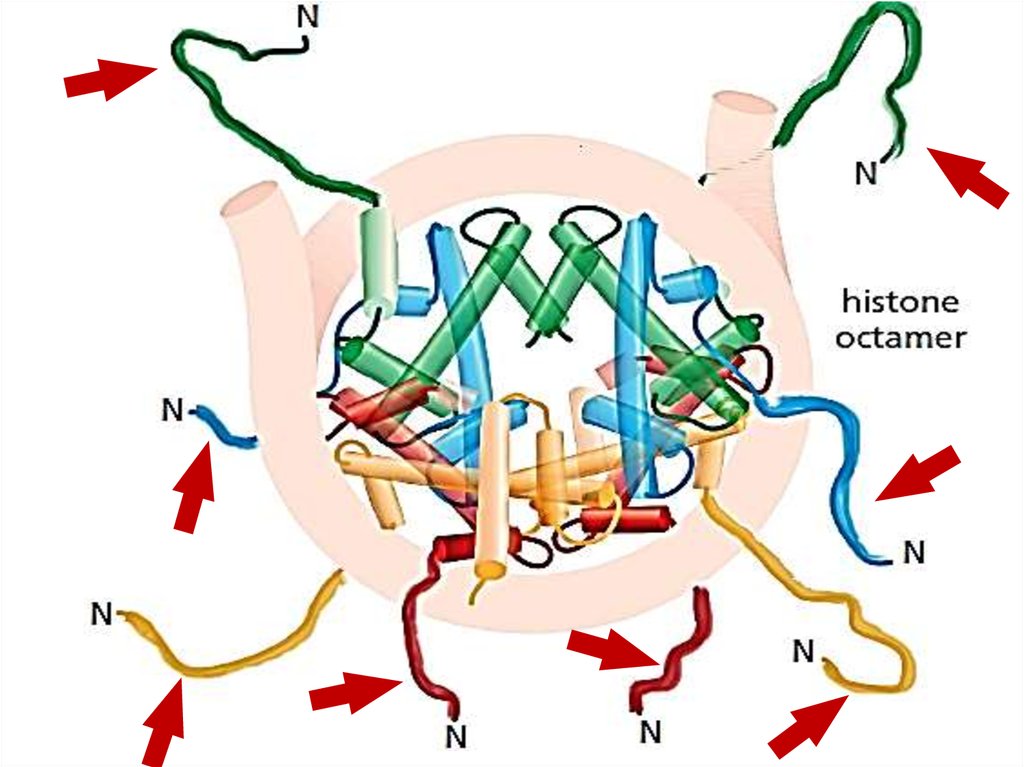
9. Нуклеосома – базовая структура эукариотической хромосомы
10.
11.
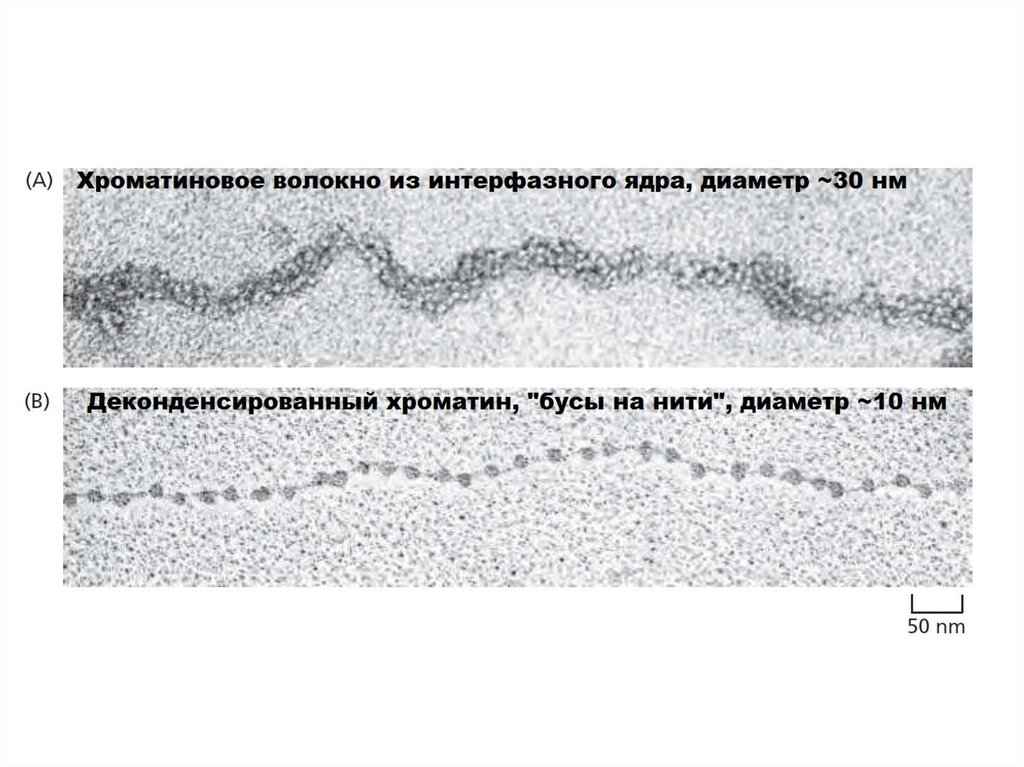
Длина линкерной ДНК может варьировать от нескольких до 80 пн,однако в среднем нуклеосомы повторяются каждые 200 пн.
Уплотнение длины ДНК примерно в три раза.
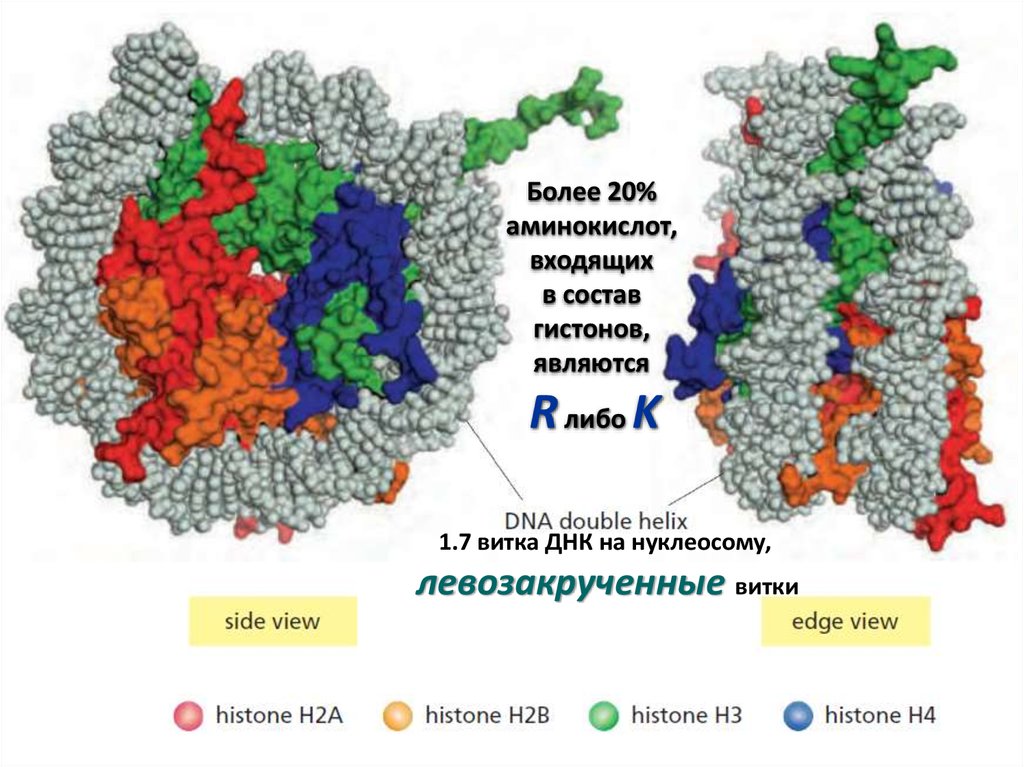
12.
Более 20%аминокислот,
входящих
в состав
гистонов,
являются
R либо K
1.7 витка ДНК на нуклеосому,
левозакрученные витки
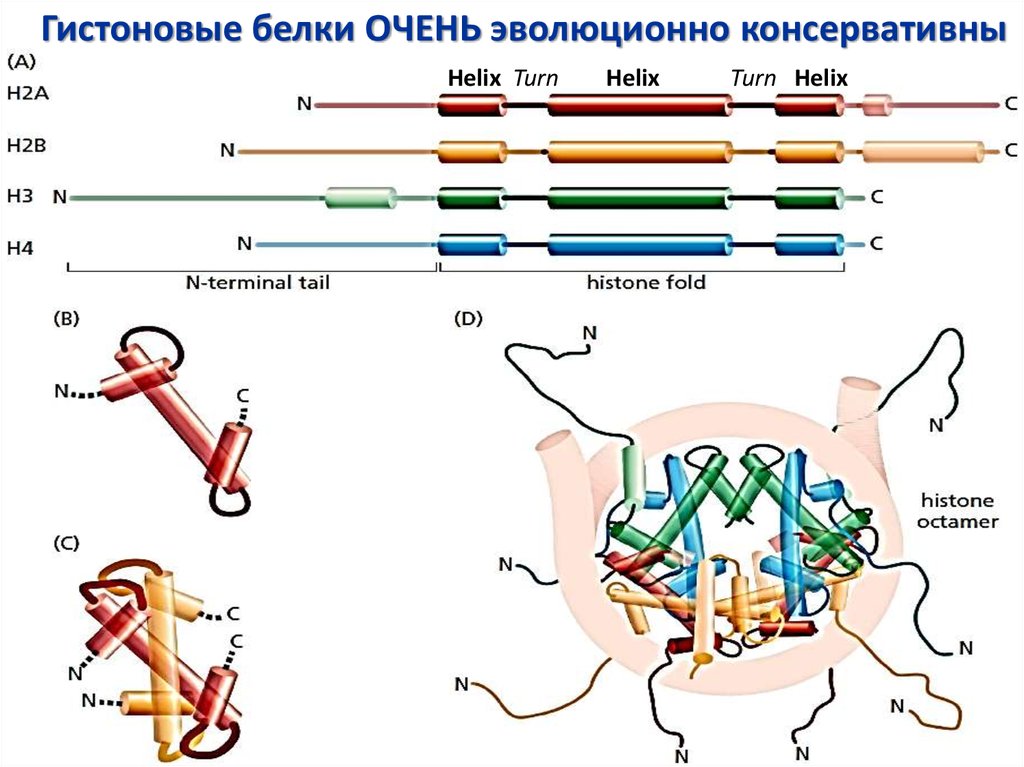
13.
Гистоновые белки ОЧЕНЬ эволюционно консервативныHelix Turn
Helix
Turn Helix
14.
• Взаимодействие между ДНК и гистоновым коромдовольно интенсивно: 142 водородные связи
образуются между ДНК и гистоновым кором в
каждой нуклеосоме.
• Около половины этих водородных связей
образовывается между аминокислотами
гистонового кора и сахарофосфатным остовом
ДНК.
• Около 1/5 аминокислот гистонового кора
представлено АК с положительно заряженной
боковой цепью – аргинином и лизином, которые
нейтрализуют отрицательно заряженную ДНК.
• Такие многочисленные взаимодействия между ДНК
и гистоновым кором частично объясняют, почему
ДНК практически любой последовательности
может быть связана с октамерным гистоновым
кором.
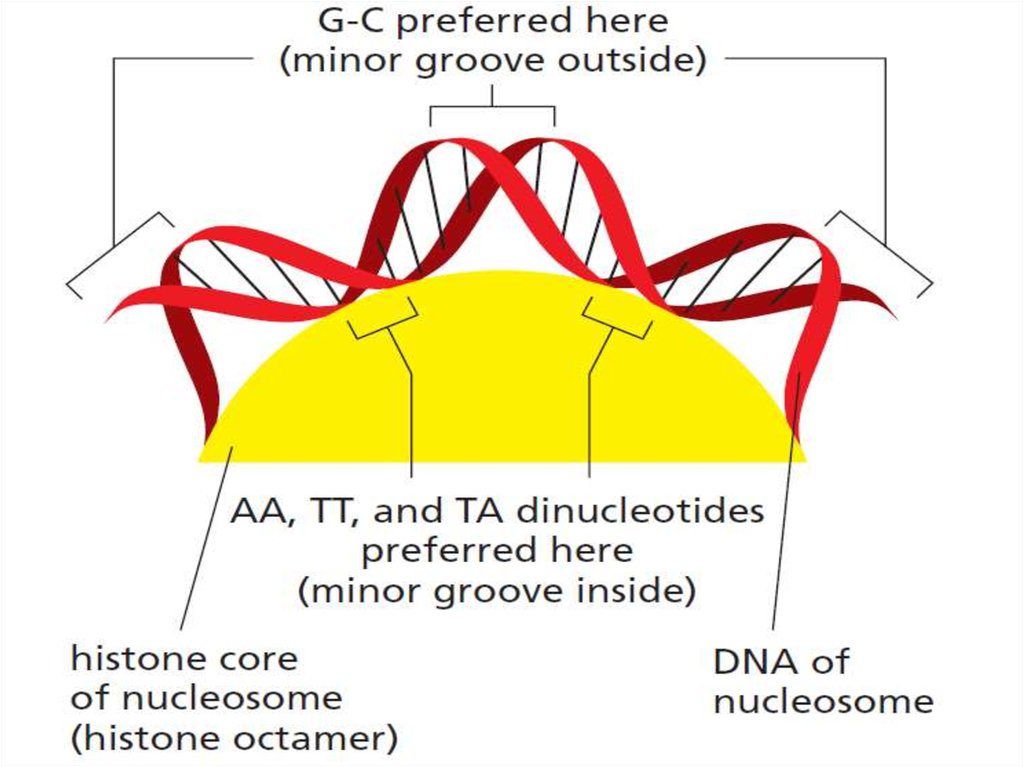
15.
• Накручивание ДНК на гистоновый кор приводитк частичнму сжатию (сдавливанию) малой
бороздки.
• Некоторые динуклеотиды в малой бороздке
более легко переносят такое сжатие, а
некоторые – связываются с нуклеосомой более
сильно, чем другие, что приводит к
предпочтительному относительному
расположению некоторых
последовательностей относительно
гистонового кора. Однако такое влияние
последовательности на положение
относительно кора является довольно слабым.
16.
17.
18.
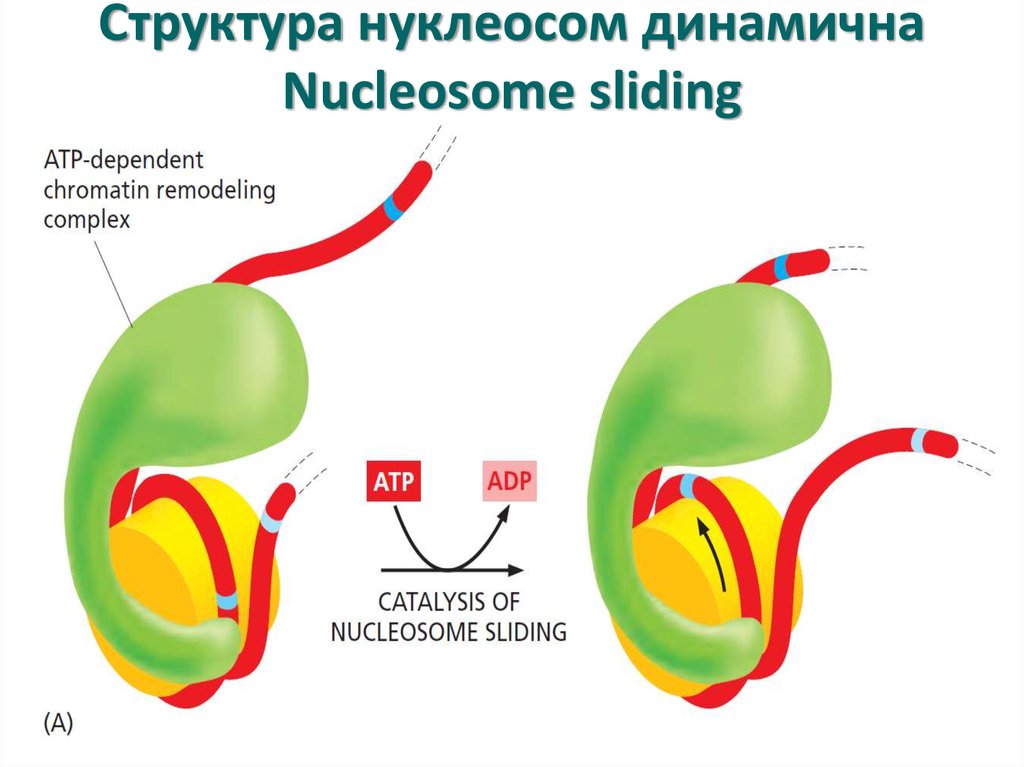
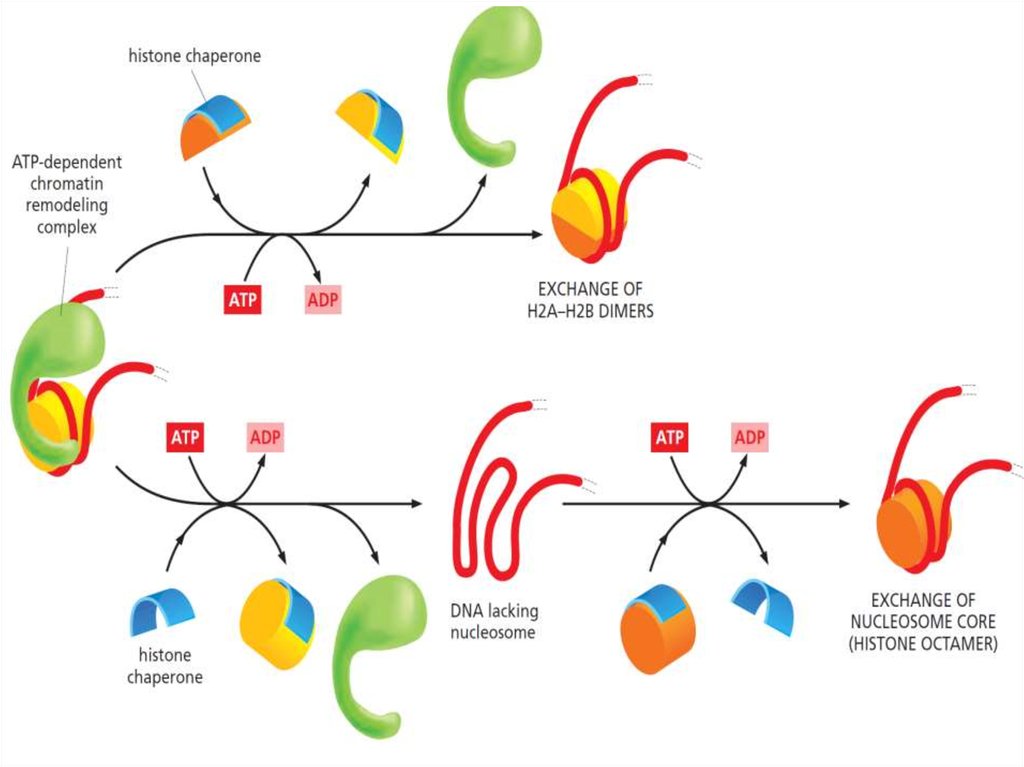
Структура нуклеосом динамичнаNucleosome sliding
19.
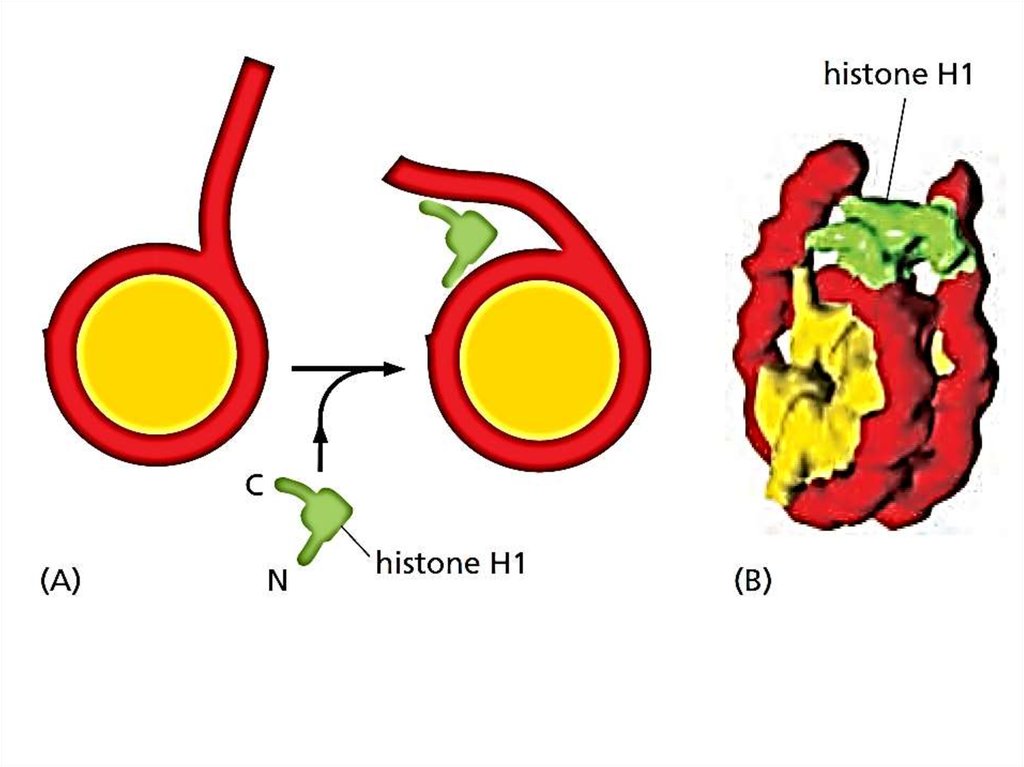
20.
Нуклеосомы упаковываютсяв компактное хроматиновое волокно
21.
22.
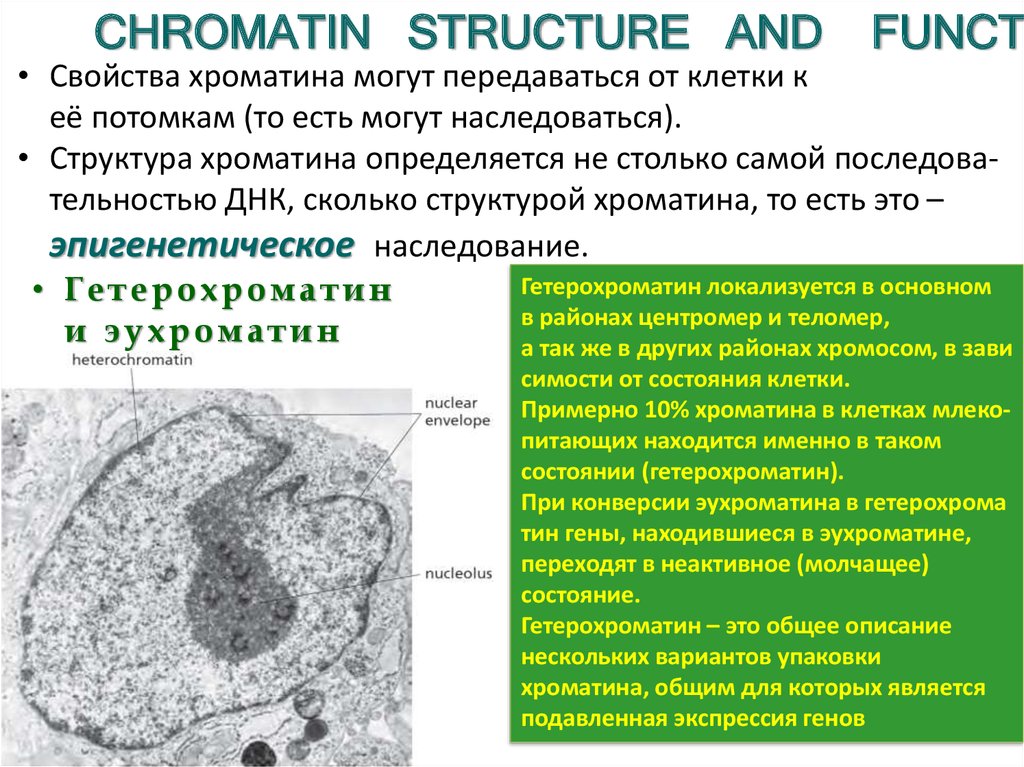
CHROMATIN STRUCTURE ANDFUNCT
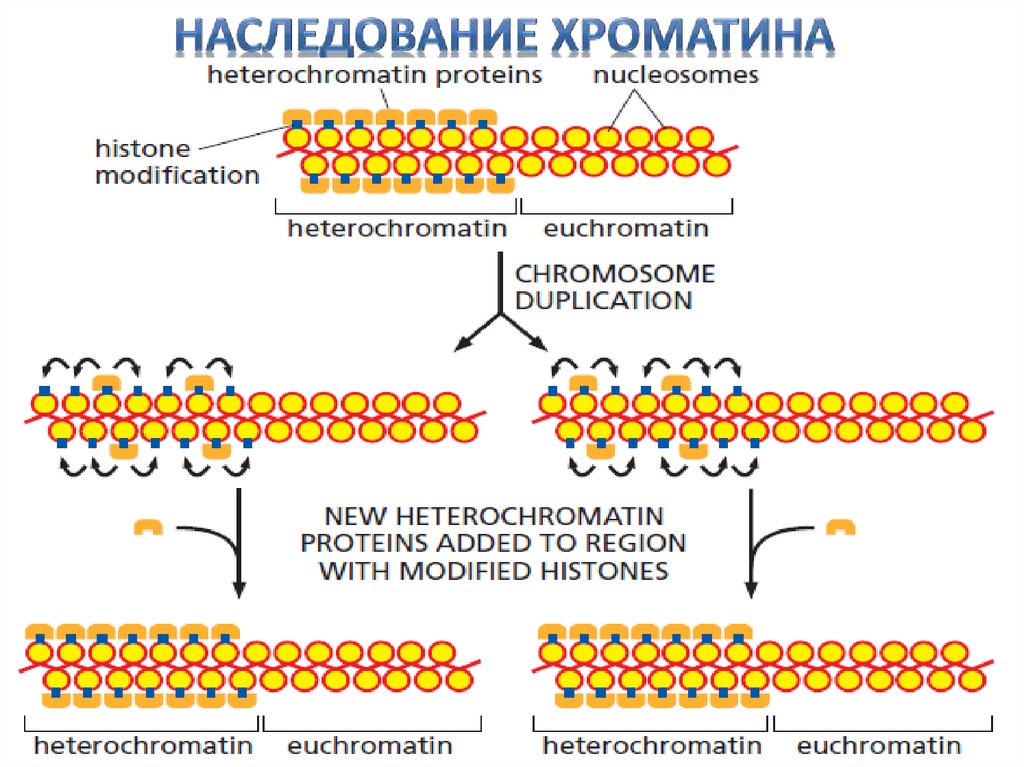
• Свойства хроматина могут передаваться от клетки к
её потомкам (то есть могут наследоваться).
• Структура хроматина определяется не столько самой последовательностью ДНК, сколько структурой хроматина, то есть это –
эпигенетическое наследование.
Гетерохроматин локализуется в основном
• Гетерохроматин
в районах центромер и теломер,
и эухроматин
а так же в других районах хромосом, в зави
симости от состояния клетки.
Примерно 10% хроматина в клетках млекопитающих находится именно в таком
состоянии (гетерохроматин).
При конверсии эухроматина в гетерохрома
тин гены, находившиеся в эухроматине,
переходят в неактивное (молчащее)
состояние.
Гетерохроматин – это общее описание
нескольких вариантов упаковки
хроматина, общим для которых является
подавленная экспрессия генов
23.
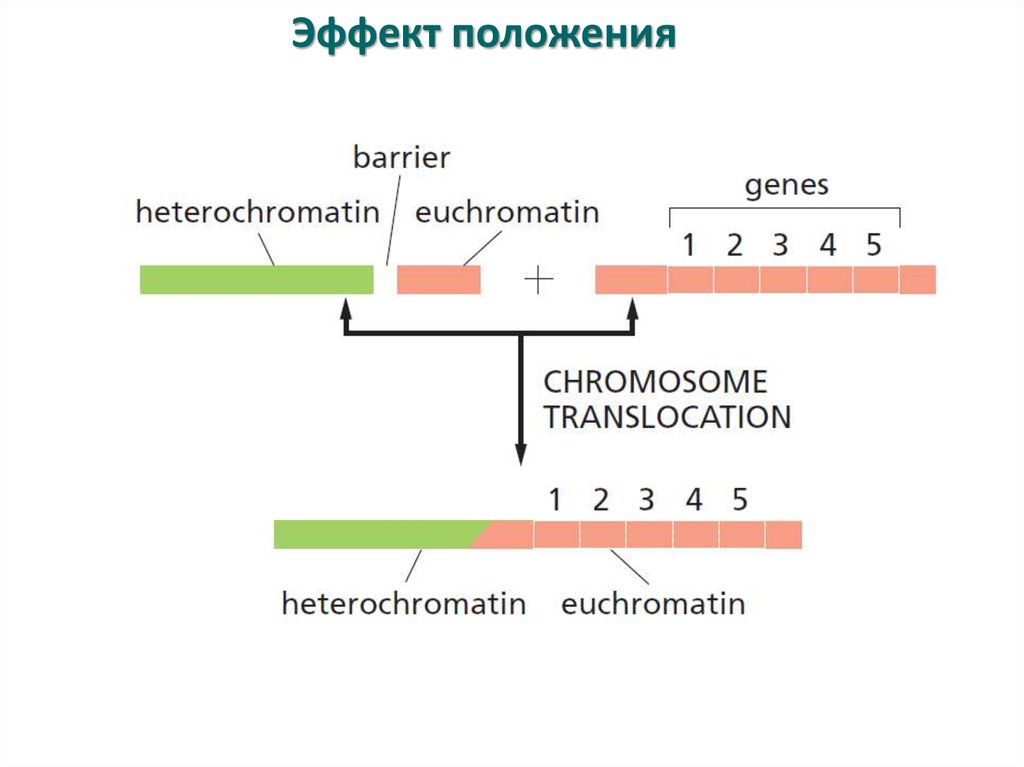
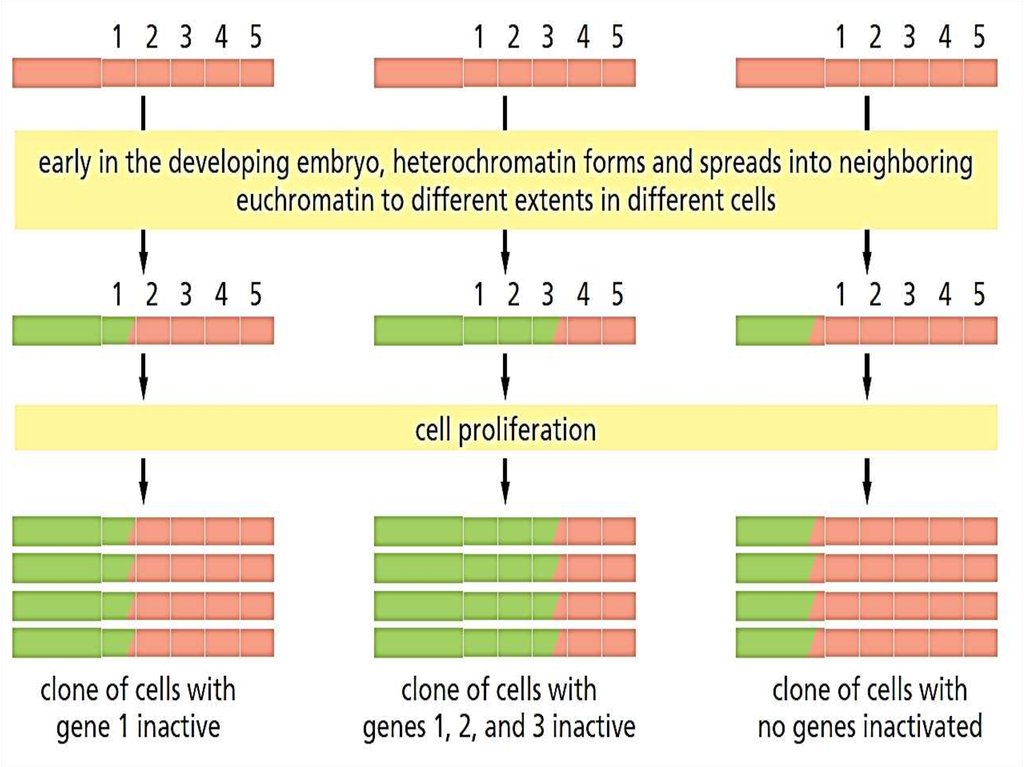
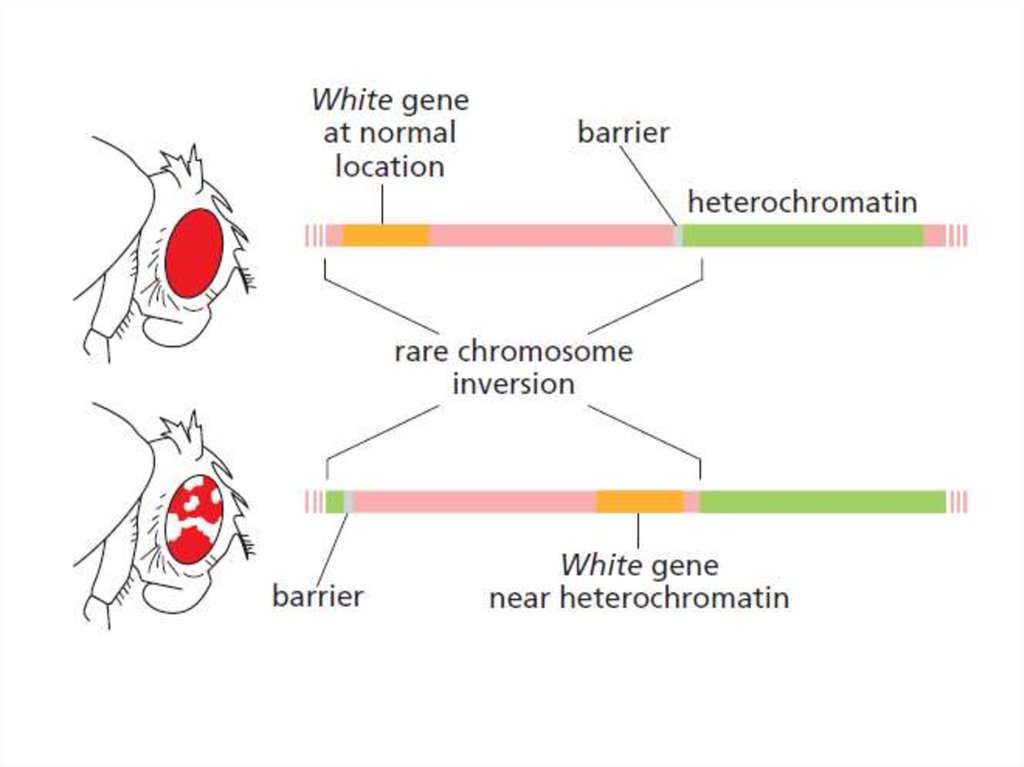
Эффект положения24.
25.
26.
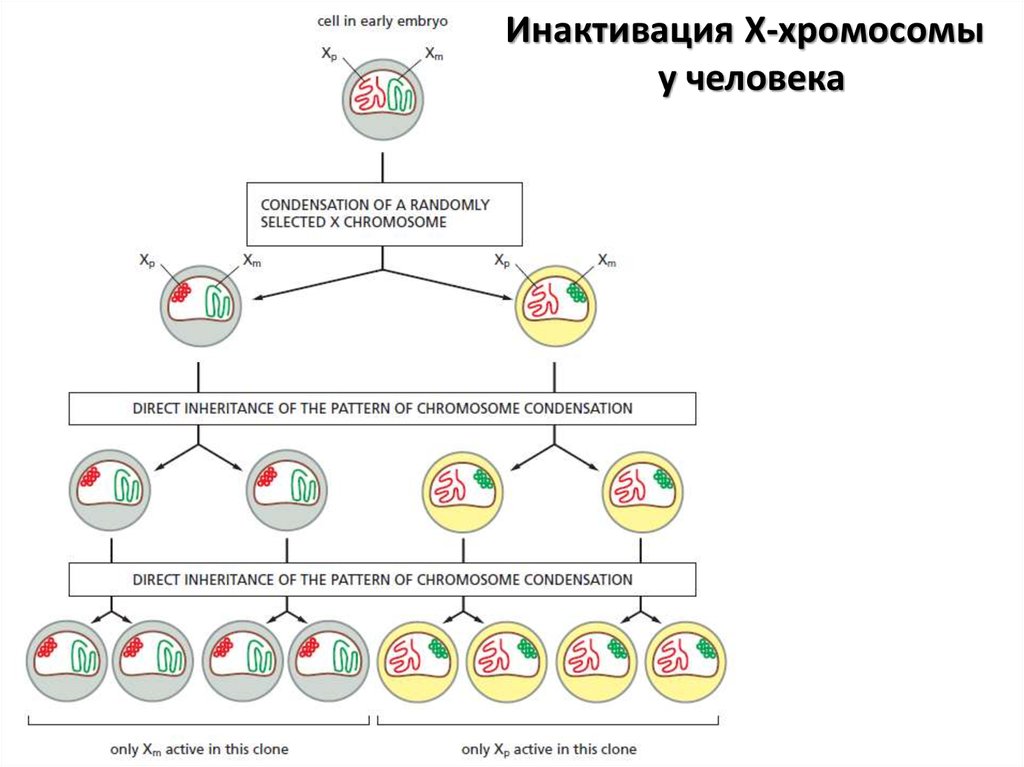
Инактивация Х-хромосомыу человека
27.
Путем скрининга многочисленных мутантовбыло выявлено более 100 генов,
влияющих на распространение гетерохроматина.
Многие из этих генов кодируют
негистоновые хромосомальные белки,
которые взаимодействуют с гистонами
и модифицируют или поддерживают их
в определенном состоянии
28.
29.
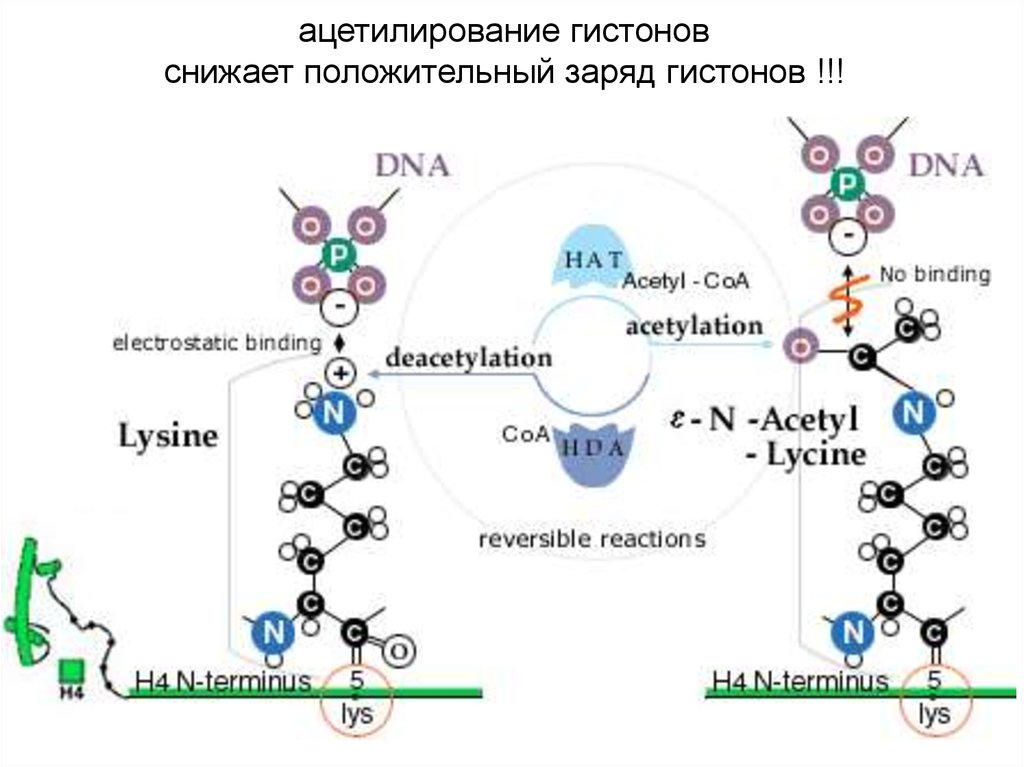
ацетилирование гистоновснижает положительный заряд гистонов !!!
30.
31.
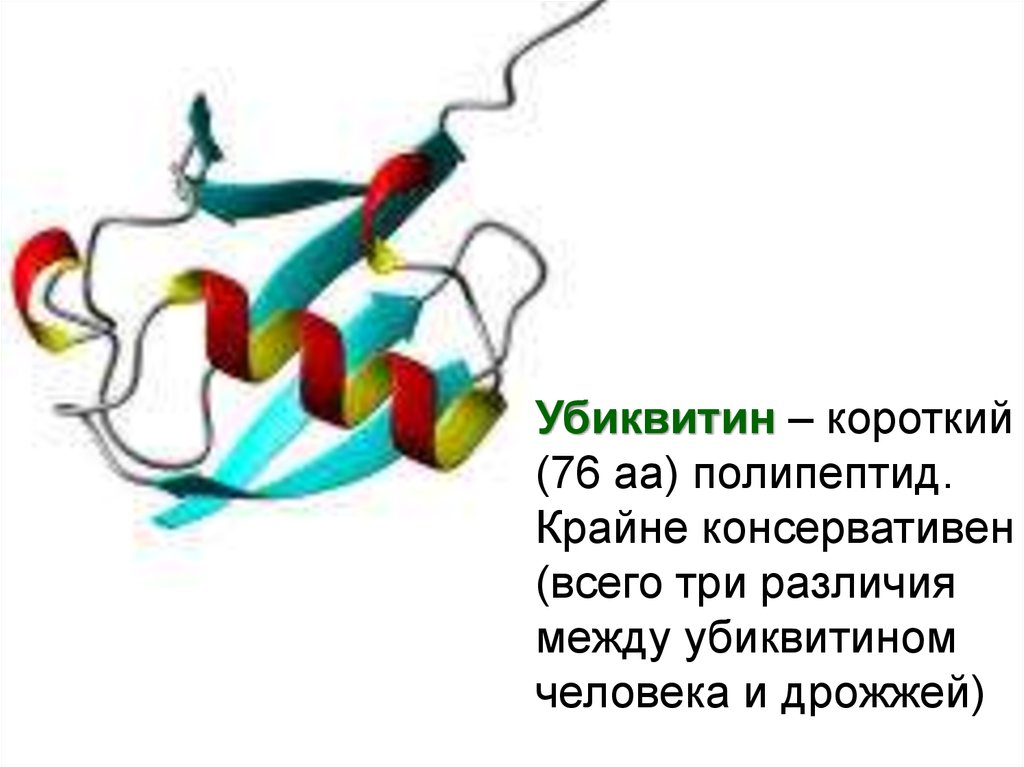
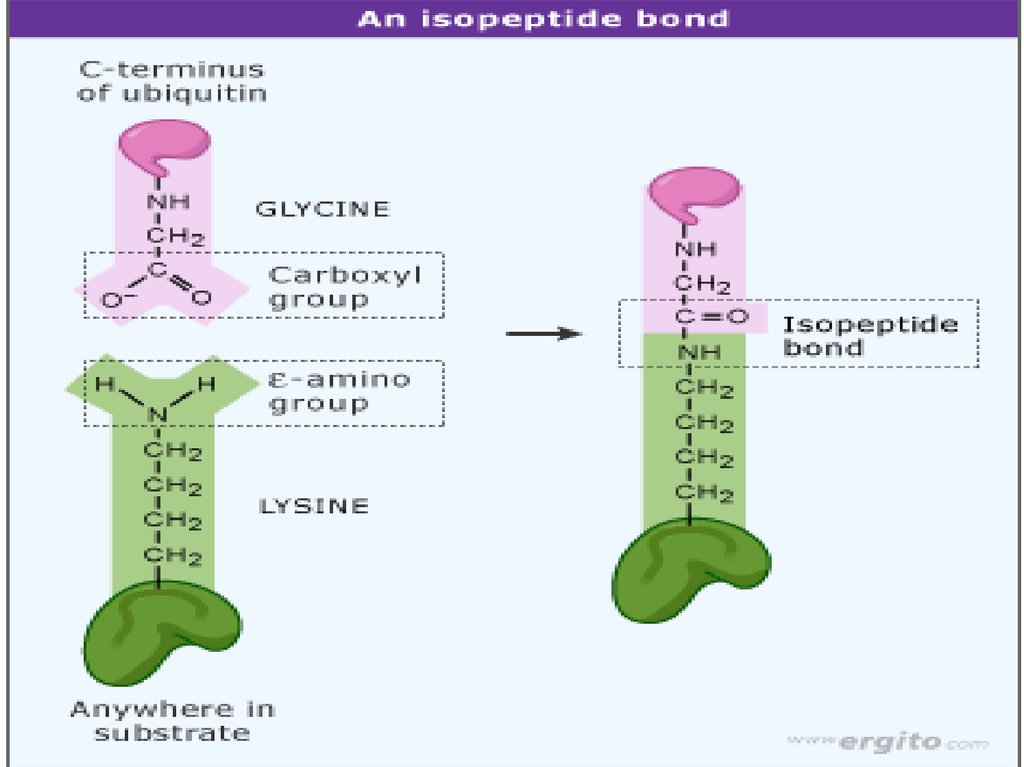
Убиквитин – короткий(76 аа) полипептид.
Крайне консервативен
(всего три различия
между убиквитином
человека и дрожжей)
32.
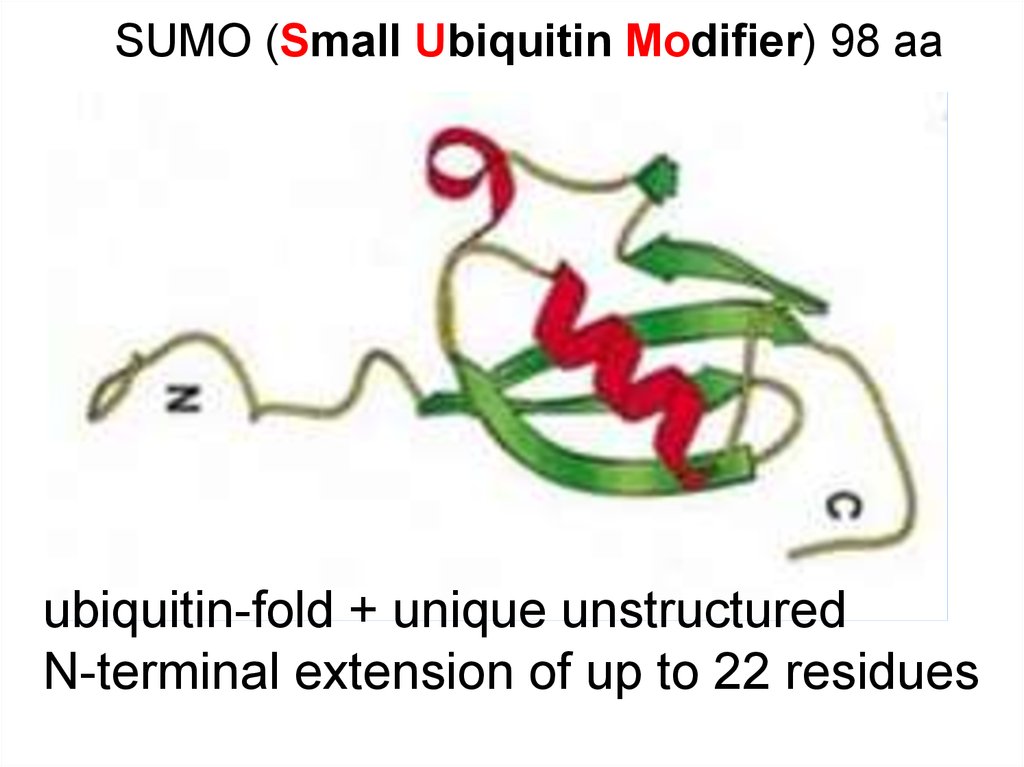
SUMO (Small Ubiquitin Modifier) 98 ааubiquitin-fold + unique unstructured
N-terminal extension of up to 22 residues
33.
34.
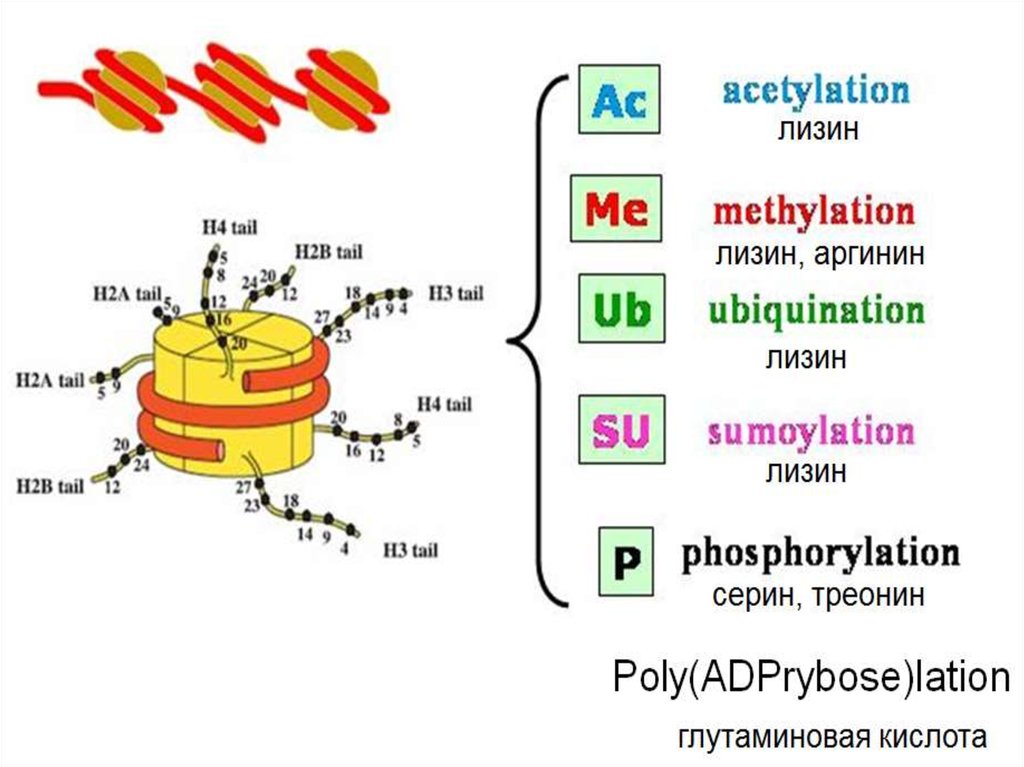
Модификации гистонов могут:1. прямо влиять на свойство октамера гистонов
(например посредством снижения заряда)
2. Выполнять сигнальную функцию,
способствую привлечения различных белков
(ГИСТОНОВЫЙ КОД)
Важен не только тип модификации и позиция,
по которой осуществляется модификация,
но и количественные характеристики модификаций.
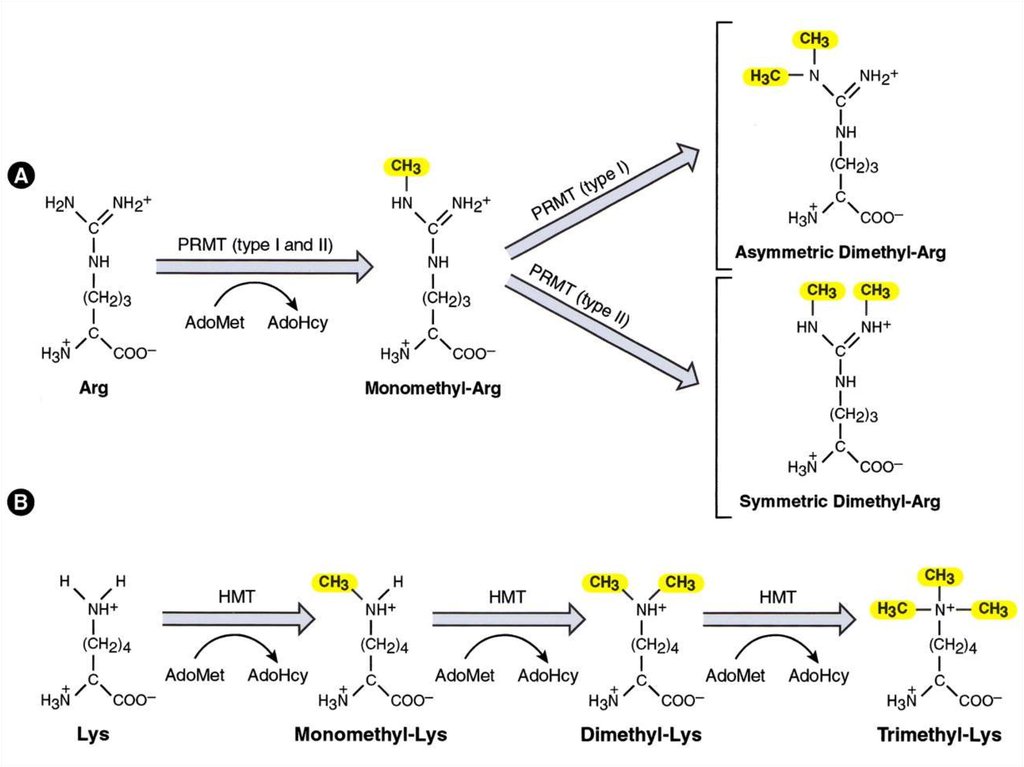
Например, монометилирование и триметилирование
лизина могут иметь прямо противоположные последствия
35.
3. Модифицироваться могут не только N-концы, но ипозиции в глобулах гистонов (около 20 различных
модификаций)
4. Модификации гистонов обратимы. Одни ферменты
вносят модификацию, другие её снимают:
histone acetyl transferases (HATs) ацетилируют лизин,
а histone deacetylase complexes (HDACs) снимают
ацетильную группу.
Каждый из этих белков в определенное время
рекрутируется на определенный сайт ДНК.
Рекрутирование инициируется, в частности, активностью
транскрипционных факторов, узнающих определенные
участки ДНК.
Таким образом, нуклеотидная последовательность ДНК
полностью определяет паттерн модификации гистонов.
36.
Тем неменее, некоторые модификации гистонов могутоставаться и после транскрипции, представляя собой
клеточную память.
Различные варианты гистонов и их функции
37.
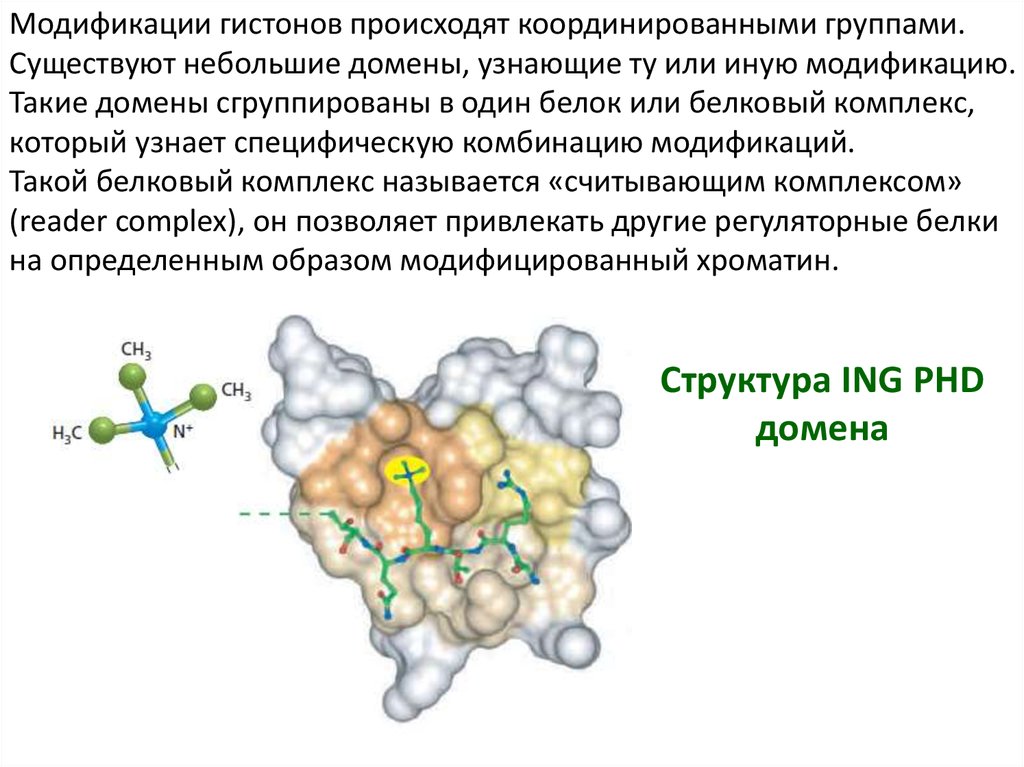
Модификации гистонов происходят координированными группами.Существуют небольшие домены, узнающие ту или иную модификацию.
Такие домены сгруппированы в один белок или белковый комплекс,
который узнает специфическую комбинацию модификаций.
Такой белковый комплекс называется «считывающим комплексом»
(reader complex), он позволяет привлекать другие регуляторные белки
на определенным образом модифицированный хроматин.
Структура ING PHD
домена
38.
39.
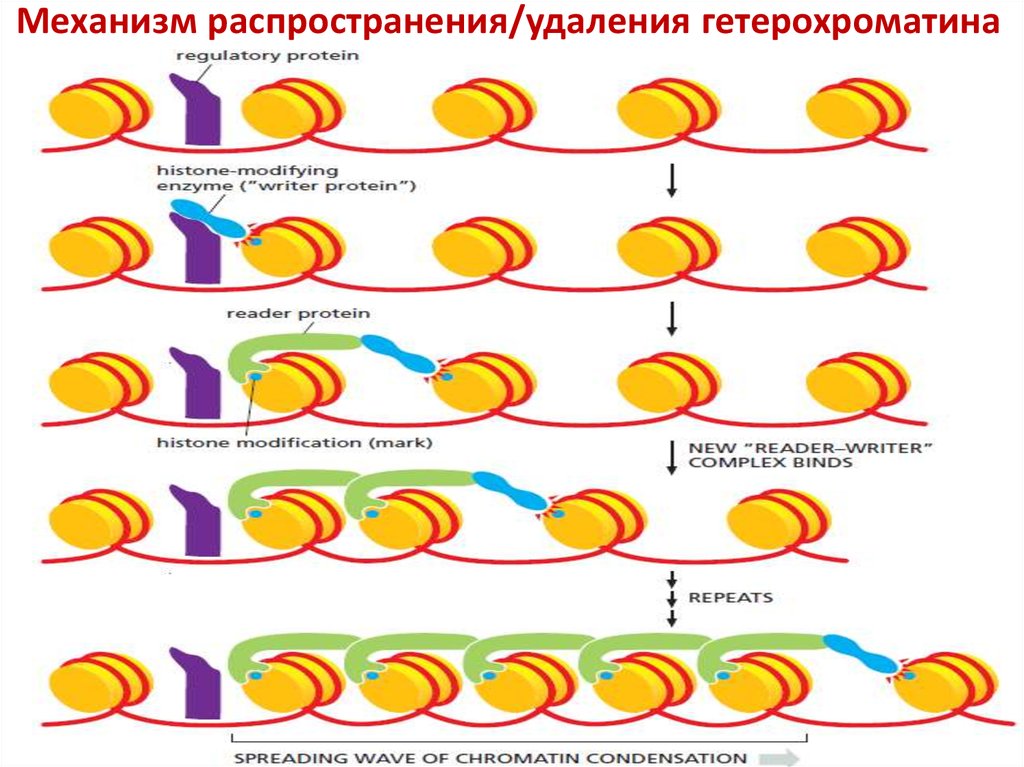
Механизм распространения/удаления гетерохроматина40.
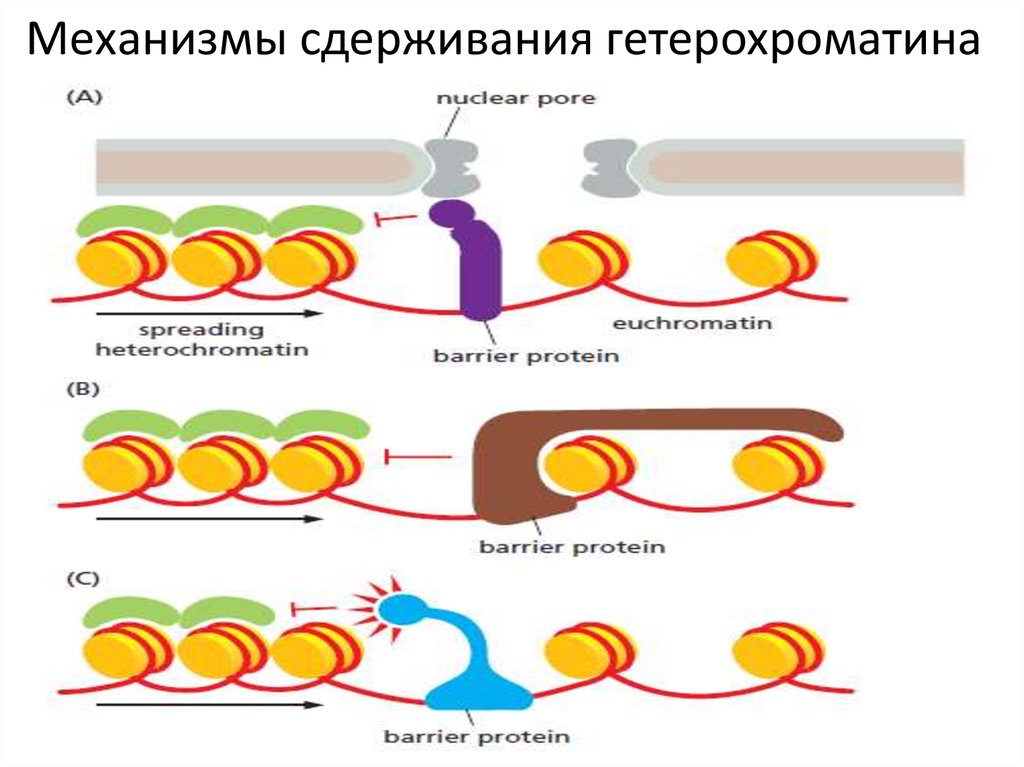
Как преодолевается противоречивое поведениесоседних участков хроматина с разной функцией?
Существуют специфические последовательности
ДНК, служащие барьером между соседними
участками хроматина.
Пример: в предшественниках эритроцитов генетический
локус HS4 разделяет участок активного хроматина, несущего ген
β-глобина, от соседнего гетерохроматинового региона. При делеции
HS4 ген β-глобина оказывается под влиянием распространившегося
соседнего гетерохроматина, вследствие чего глобиновый ген слабо
экспрессируется, что ведет к тяжелой форме анемии.
HS4 сайт несет в себе многочисленные участки связывания с гистоновой ацетилазой. А поскольку ацетилирование лизина несовместимо с метилированием
(метилир-е необходимо для гетерохроматизации), то гистоновые ацетилазы –
кандидаты в барьерные белки.
41.
Механизмы сдерживания гетерохроматина42.
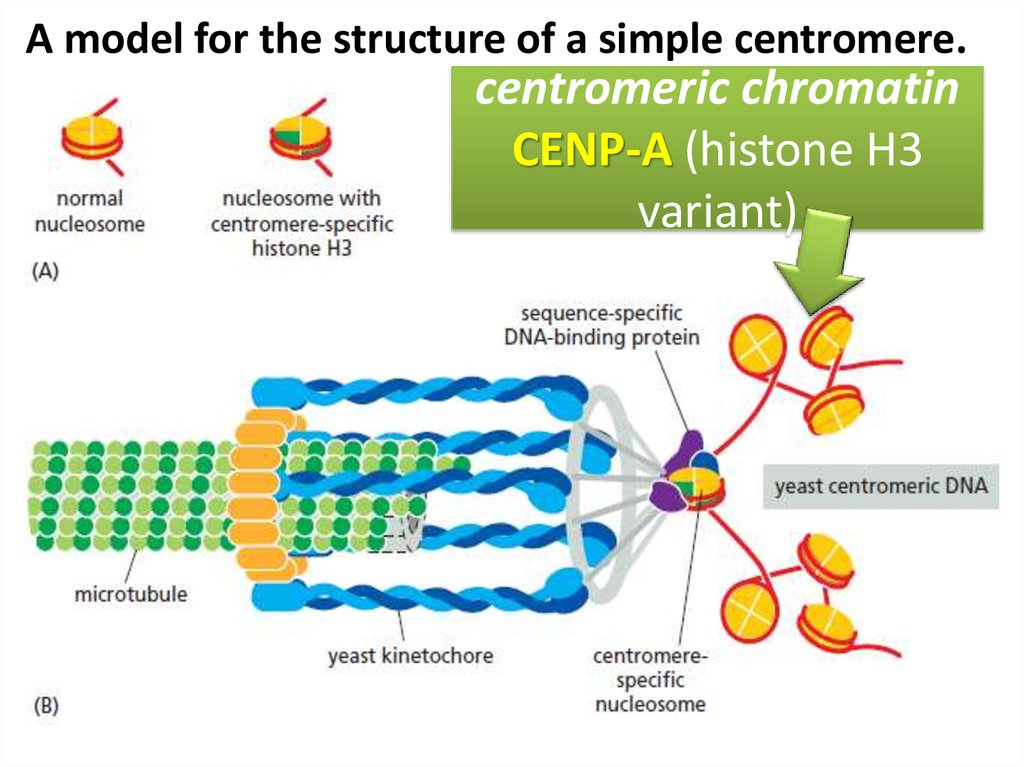
A model for the structure of a simple centromere.centromeric chromatin
CENP-A (histone H3
variant)
43.
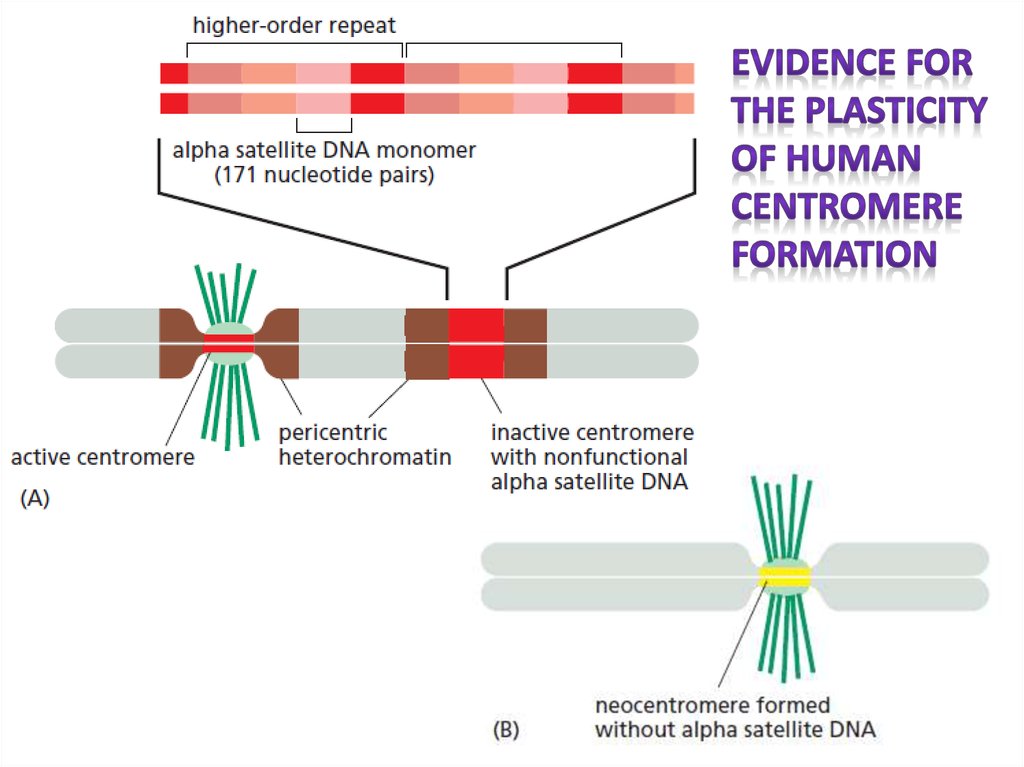
У человека центромерная ДНК состоитиз коротких повторяющихся последовательностей,
известных как альфа-сателлитная ДНК.
Однако такие последовательности находят и в других частях хромосом. Т. о, сборка центромеры зависит скорее от специфических белков, чем от
последовательности ДНК, что подтверждают редкие случаи образования неоцентромер (иногда
в районах эухроматина и без альфа-сателлитных
повторов)
44.
45.
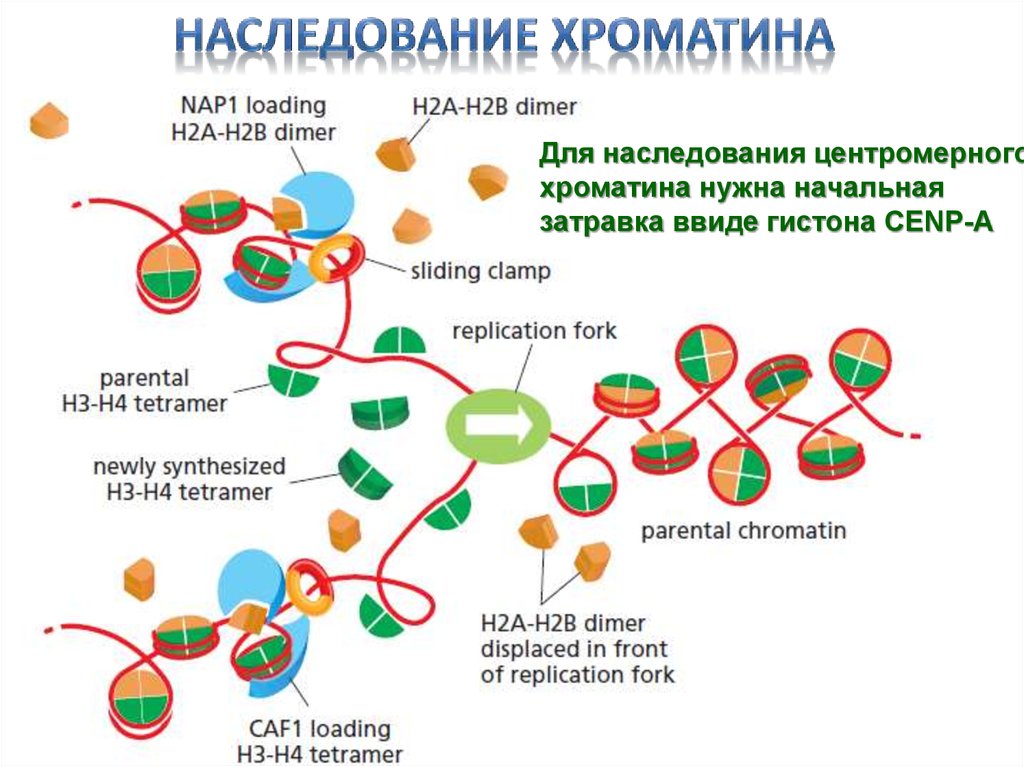
Для наследования центромерногохроматина нужна начальная
затравка ввиде гистона CENP-A
46.
47.
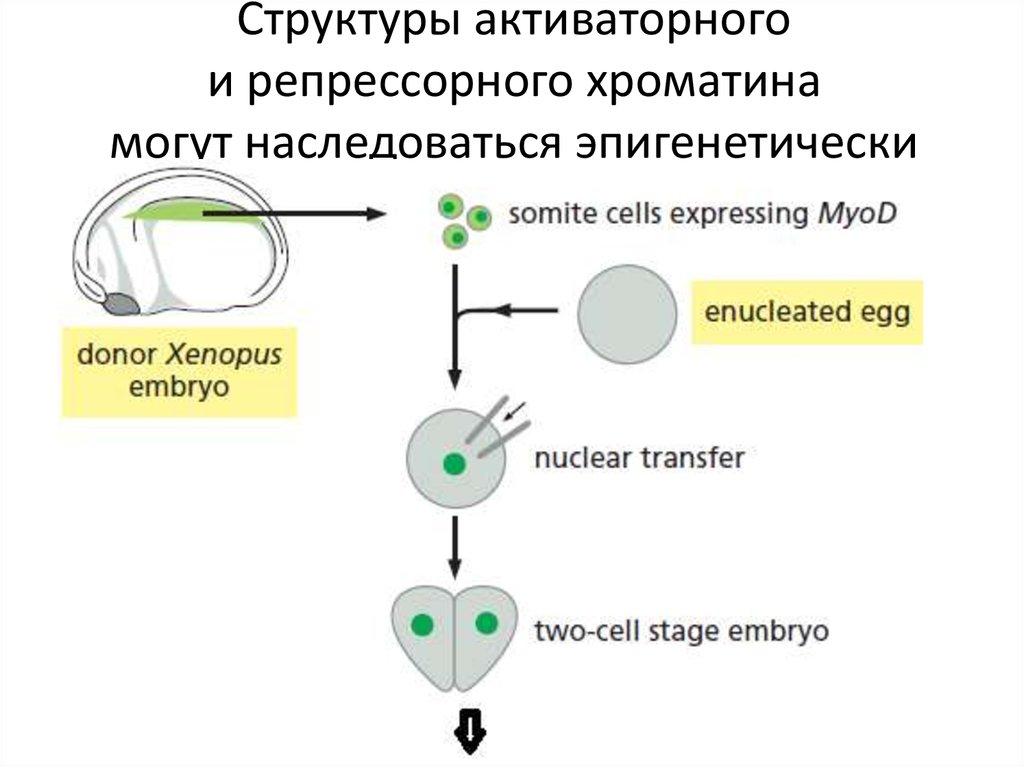
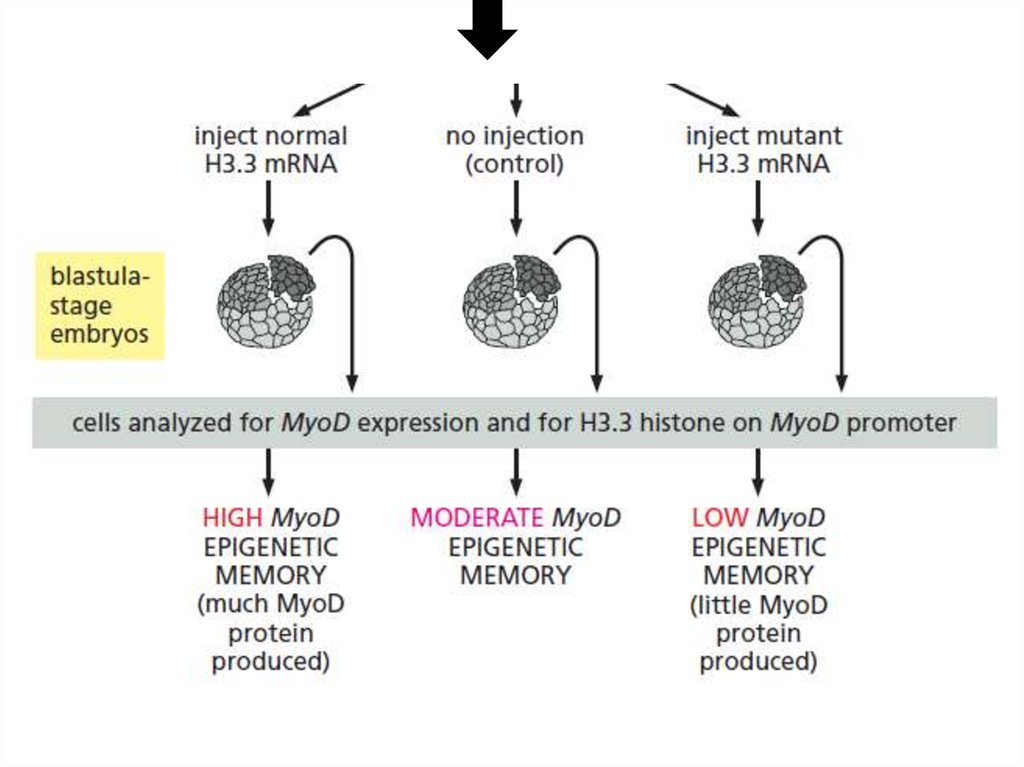
Структуры активаторногои репрессорного хроматина
могут наследоваться эпигенетически
48.
49.
50.
Даже свернутая в 30-нм волокно ДНК все равно будетиметь размер окло 1 мм, что в 100 раз больше, чем
размер ядра. Значит, она должна подвергнуться
дополнительной укладке. Такая укладка состоит из серии
петель и спиралей. Эта укладка довольно мобильна и
может изменяться в зависимости от нужд клетки.
51.
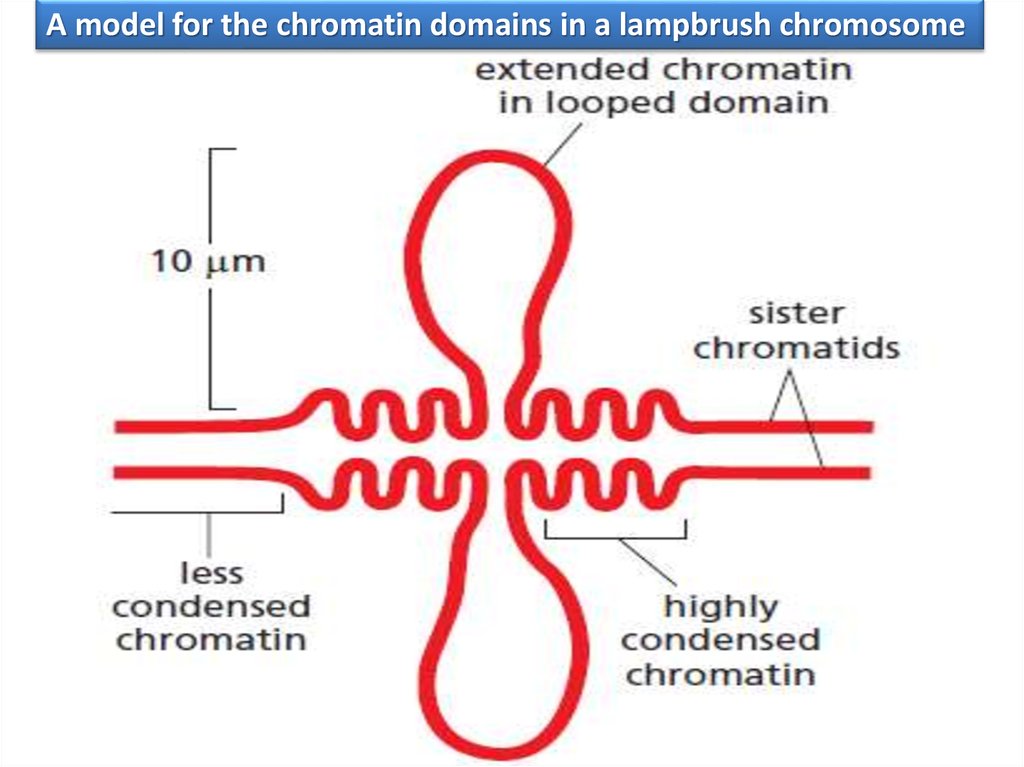
A model for the chromatin domains in a lampbrush chromosome52.
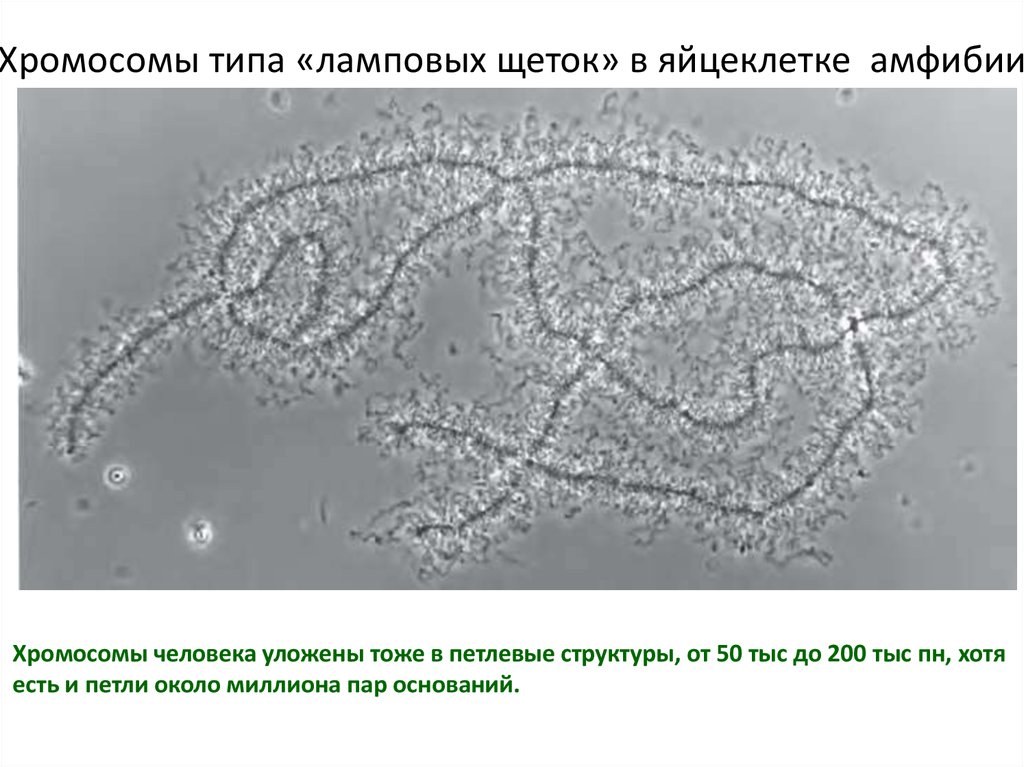
Хромосомы типа «ламповых щеток» в яйцеклетке амфибииХромосомы человека уложены тоже в петлевые структуры, от 50 тыс до 200 тыс пн, хотя
есть и петли около миллиона пар оснований.
53.
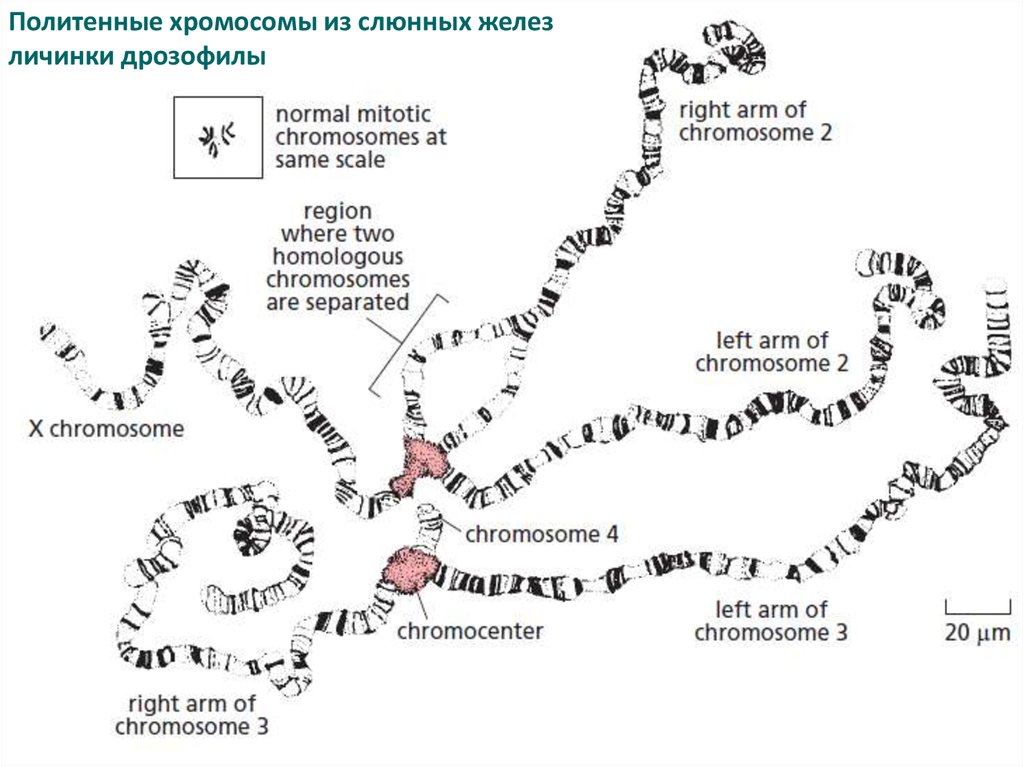
Политенные хромосомы из слюнных железличинки дрозофилы
54.
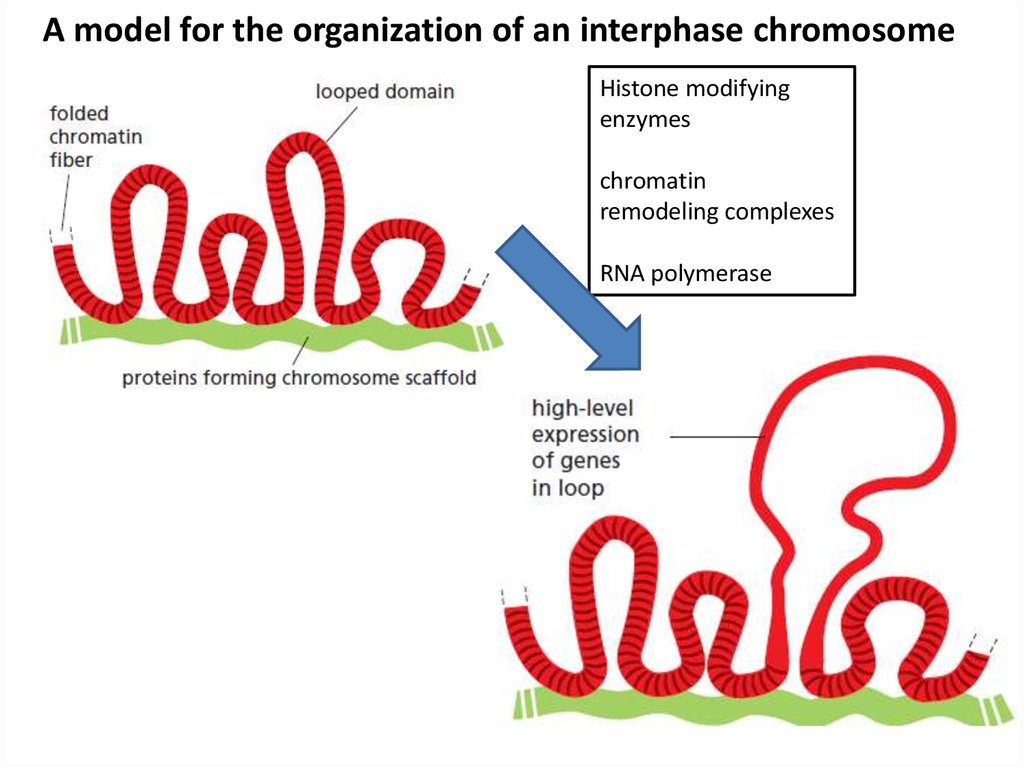
A model for the organization of an interphase chromosomeHistone modifying
enzymes
chromatin
remodeling complexes
RNA polymerase
55.
У дрозофилы в политенных хромосомах выявлено три основных типарепрессирующего хроматина (один содержит классический HP1-белок среди ещё
пяти; другой – белки семейства Polycomb (PcG белки и другие).
В процессе развития насекомого новые пуфы включаются, старые - выключаются
56.
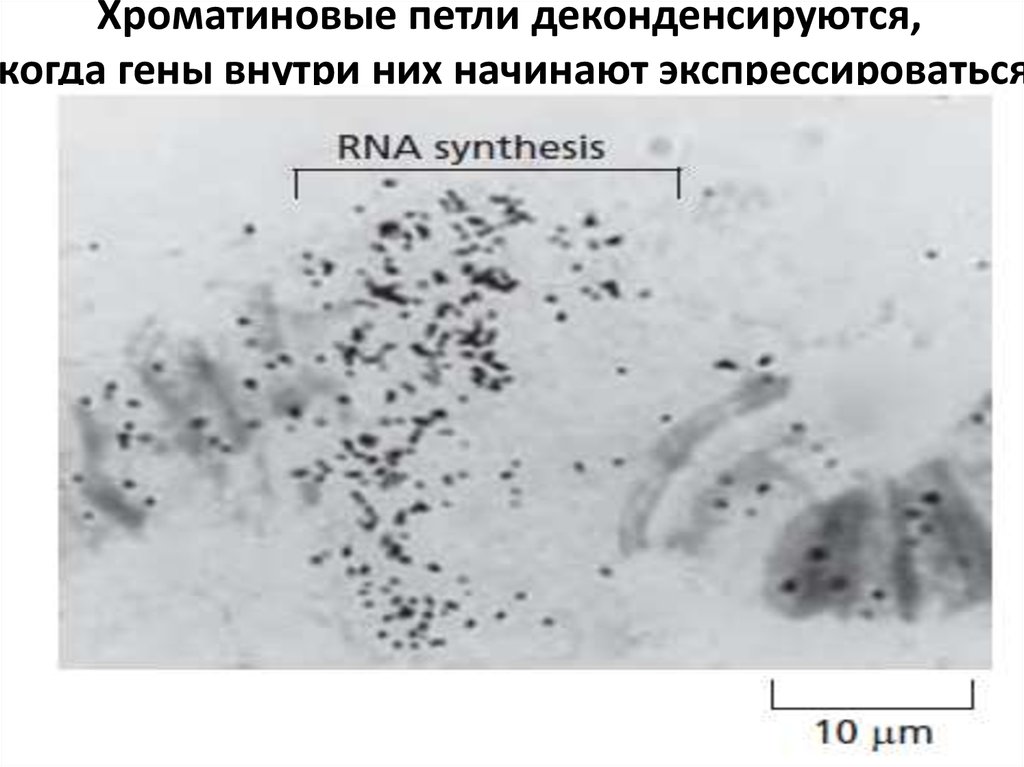
Хроматиновые петли деконденсируются,когда гены внутри них начинают экспрессироваться
57.
Хромосомы занимают в ядре определенные территории58.
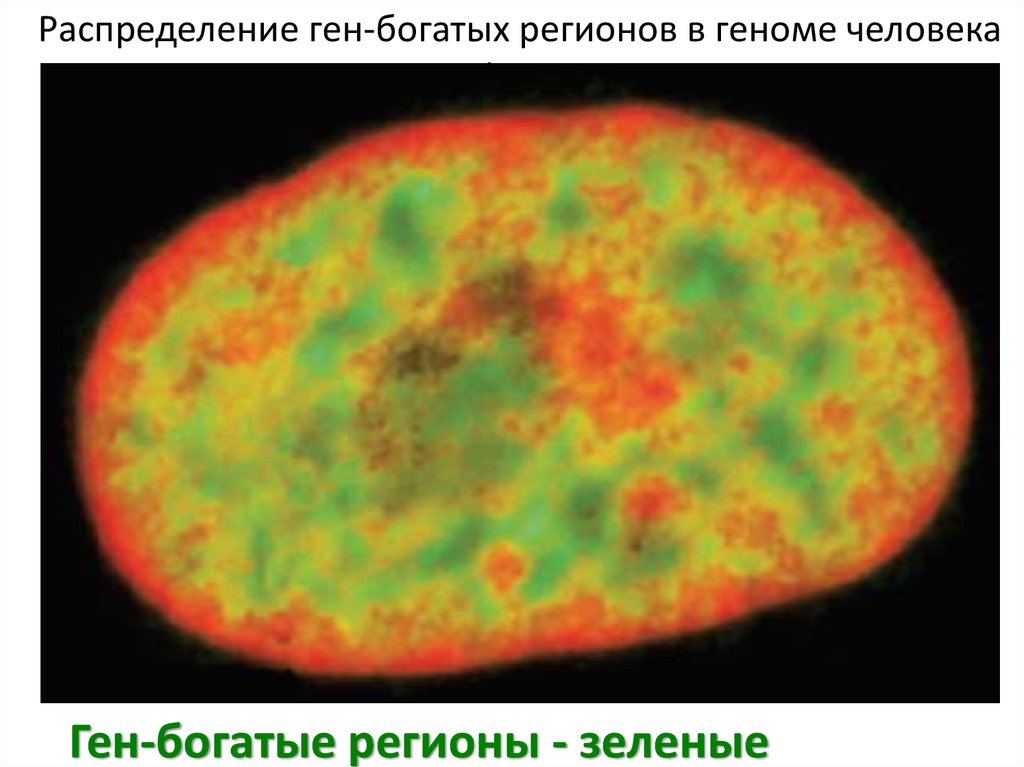
Распределение ген-богатых регионов в геноме человекав интерфазном ядре
Ген-богатые регионы - зеленые
59.
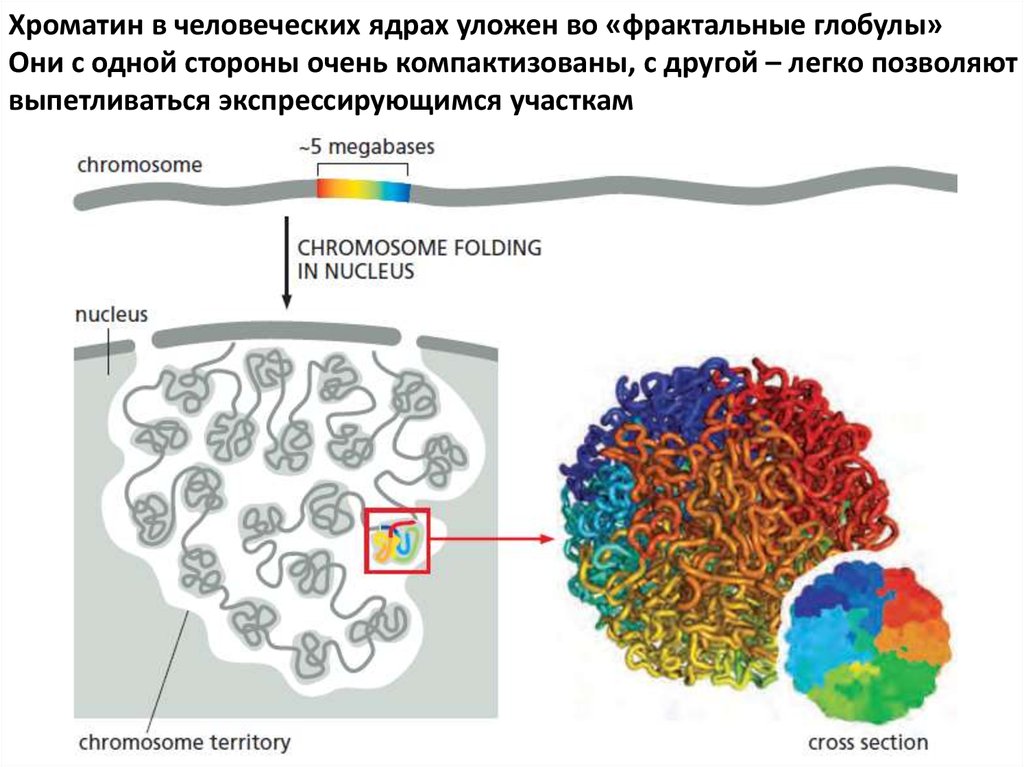
Хроматин в человеческих ядрах уложен во «фрактальные глобулы»Они с одной стороны очень компактизованы, с другой – легко позволяют
выпетливаться экспрессирующимся участкам
60.
Экспрессирующийся ген меняет свою территорию61.
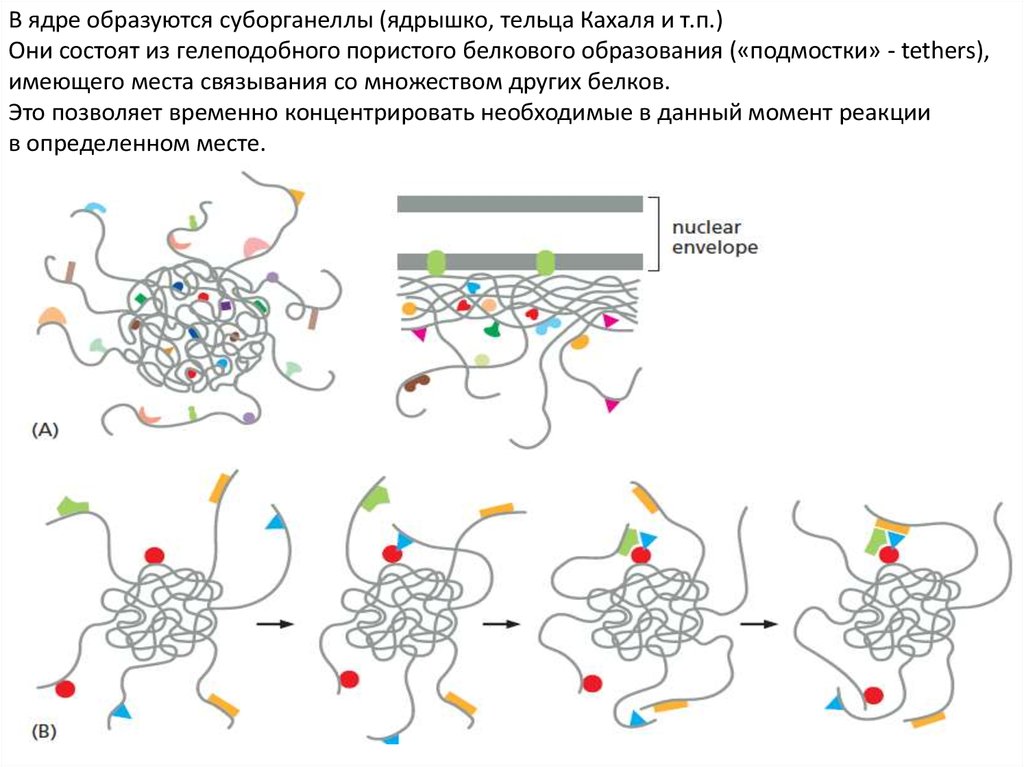
В ядре образуются суборганеллы (ядрышко, тельца Кахаля и т.п.)Они состоят из гелеподобного пористого белкового образования («подмостки» - tethers),
имеющего места связывания со множеством других белков.
Это позволяет временно концентрировать необходимые в данный момент реакции
в определенном месте.