





























 medicine
medicineSimilar presentations:
")









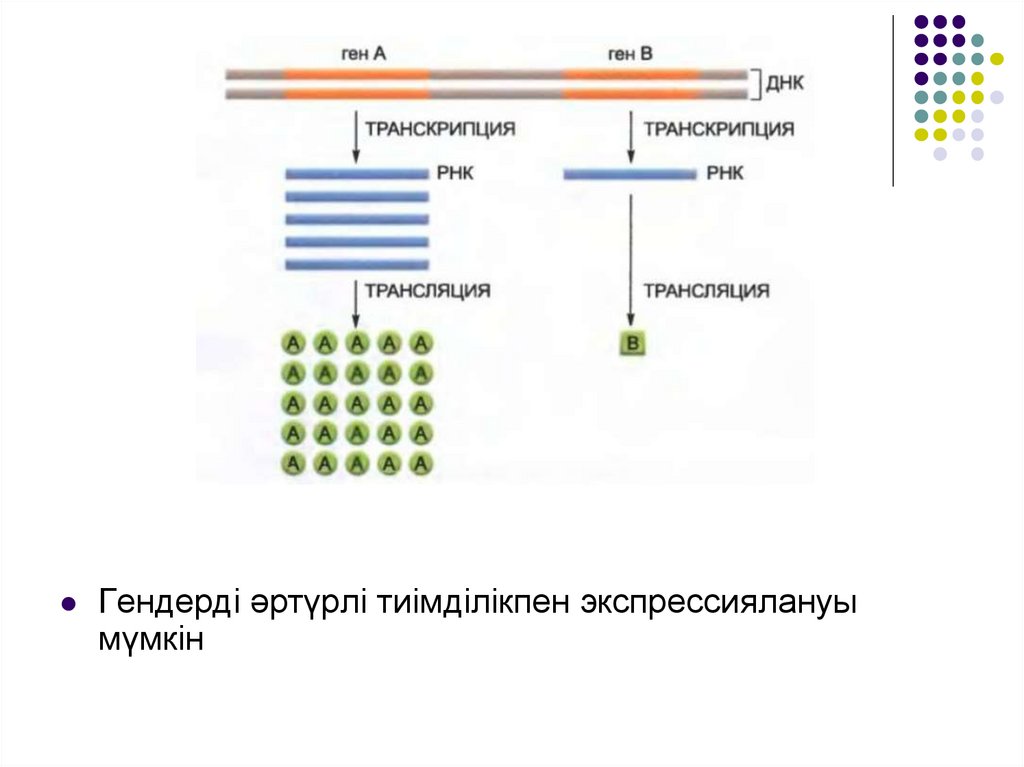
Транскрипция. Гендерді әртүрлі тиімділікпен экспрессиялануы мүмкін
1. Транскрипция
2.
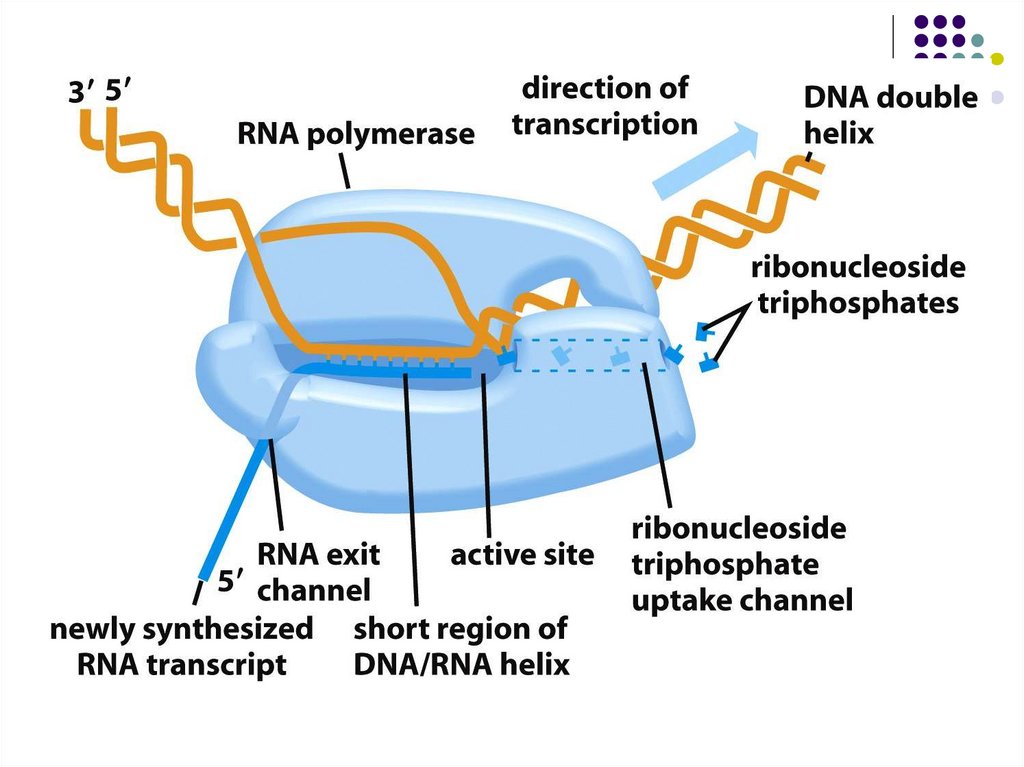
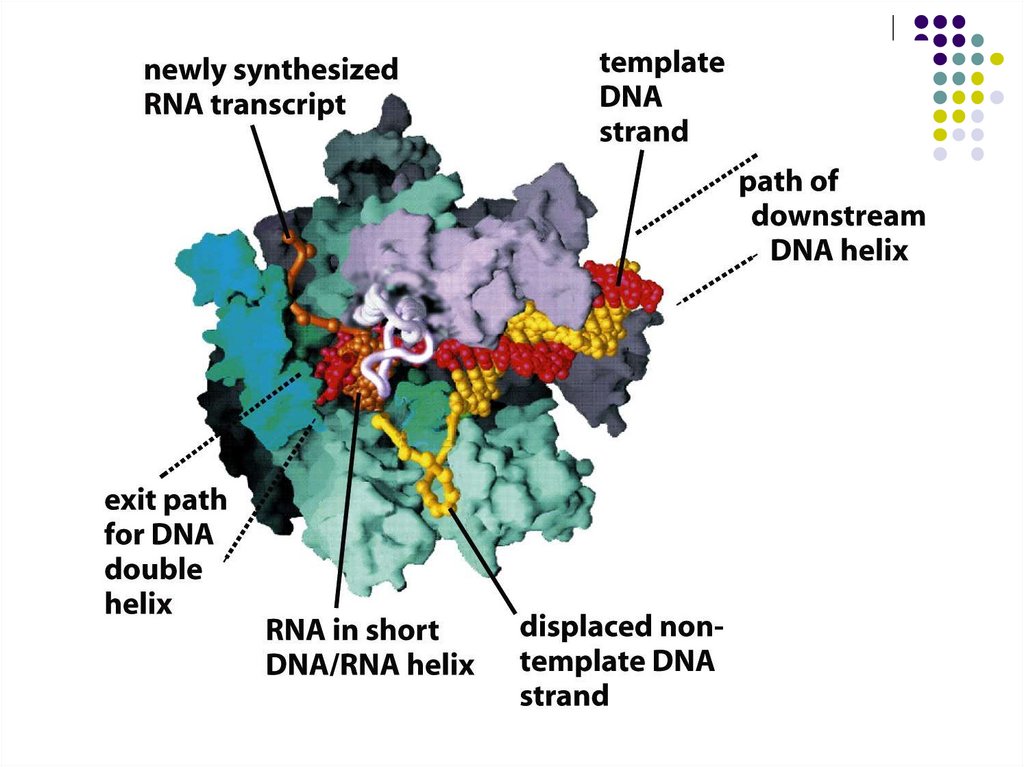
Ақпараттың екі тізбекті ДНҚ -дан бір тізбекті РНҚ- ға РНК-транскриптазанемесе РНК-полимераза деп аталатын ферменттер көмегімен
тасымалдануы транскрипция деп аталады.
Ширатылған хромосомалық ДНҚ-ның белгілі бір бөлігінің РНҚ- молекуласы
транскрипциялық бірлік деп аталады.
РНҚ биосинтезі үшін субстрат ретінде рибонуклеозидтрифосфаттар
пайдаланылады.
РНК-транскрипт синтезі 5‘тен - 3‘ ұшына бағытталып жүреді.
РНҚ синтезі аймағында ДНҚ спиралінің екі жіпшесінің (16-18 қос негіздер)
тарқатылуы жүреді,осылай 3‘тен - 5‘ке бағытталған матрицалық - ДНҚ тізбегі
пайда болады.
ДНҚ -дағы транскрипциялық бірлік бір жағынан транскрипциялық инициация
бөлімі - промотормен, ал келесі жағынан транскрипция кідіру бөлімі –
терминатормен шектеледі.
3.
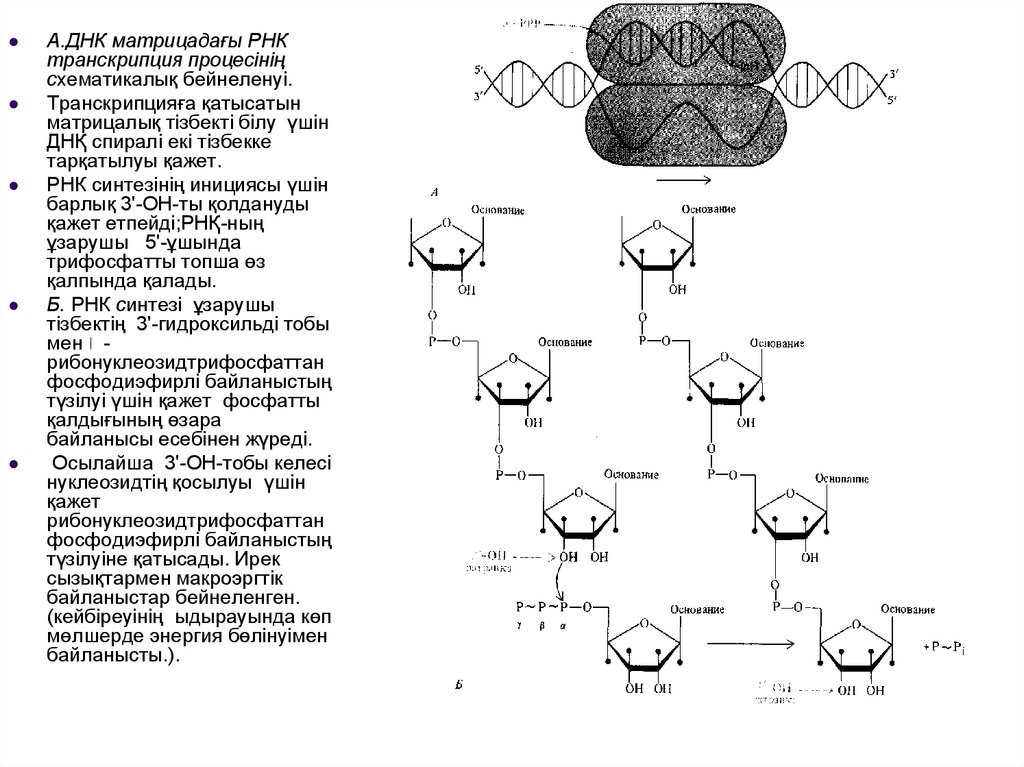
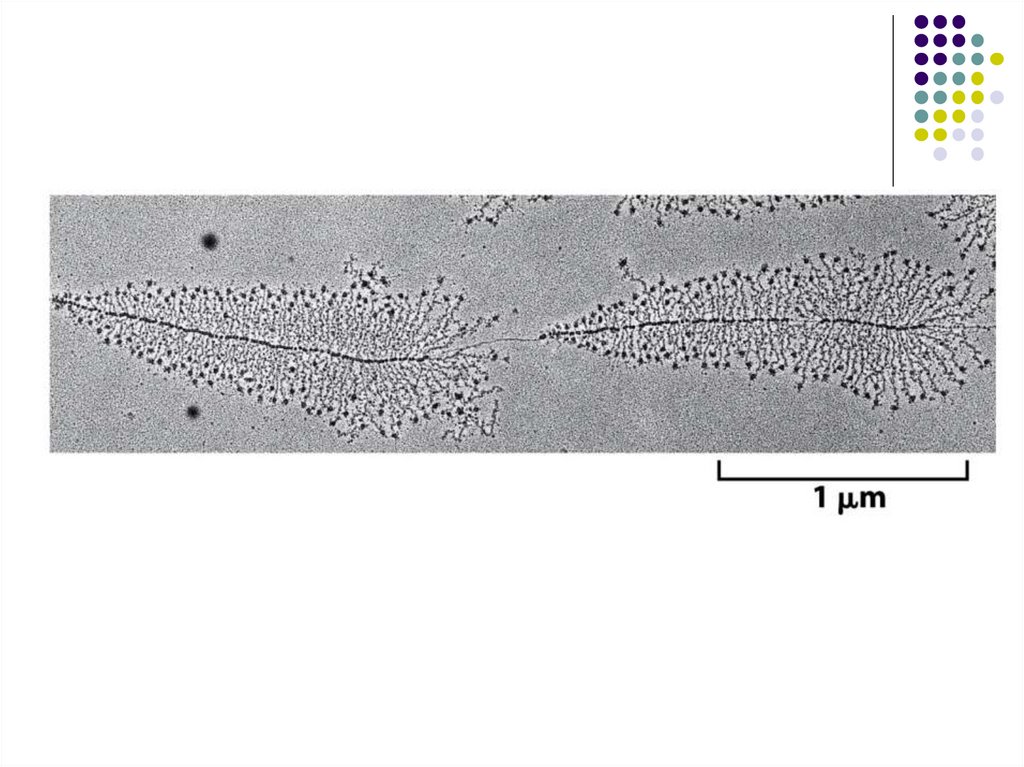
А.ДНК матрицадағы РНКтранскрипция процесінің
схематикалық бейнеленуі.
Транскрипцияға қатысатын
матрицалық тізбекті білу үшін
ДНҚ спиралі екі тізбекке
тарқатылуы қажет.
РНК синтезінің инициясы үшін
барлық 3'-ОН-ты қолдануды
қажет етпейді;РНҚ-ның
ұзарушы 5'-ұшында
трифосфатты топша өз
қалпында қалады.
Б. РНК синтезі ұзарушы
тізбектің 3'-гидроксильді тобы
мен рибонуклеозидтрифосфаттан
фосфодиэфирлі байланыстың
түзілуі үшін қажет фосфатты
қалдығының өзара
байланысы есебінен жүреді.
Осылайша 3'-ОН-тобы келесі
нуклеозидтің қосылуы үшін
қажет
рибонуклеозидтрифосфаттан
фосфодиэфирлі байланыстың
түзілуіне қатысады. Ирек
сызықтармен макроэргтік
байланыстар бейнеленген.
(кейбіреуінің ыдырауында көп
мөлшерде энергия бөлінуімен
байланысты.).
4.
Гендерді әртүрлі тиімділікпен экспрессиялануымүмкін
5.
6.
7.
Урацил аденинмен комплементарлы тізбек түзеді8.
9.
10.
11.
12.
Е.СоІі-дің ДНҚ полимеразасыРНҚның барлық 3 классының:рибосомалық рРНҚ, транспорттық тРНҚ, ақпараттық немесе
матрицалық мРНҚ молекулаларының реті жайында ақпарат беретін
бөлімдерінің транскрипциясын қамтамасыз етеді.
Эукариоттар жасушасысында, керісінше, 3 түрлі РНҚ полимераза
болады. Олардың әрқайсысы промоторды арнайы танып, РНҚның 3
класссының да транскрипциясын бақылап отырады.
Эукариоттарда РНҚ полимераза І ядрошықта орналасып, негізгі рРНҚ
синтезіне жауап береді.
РНҚ полимераза ІІІ транспорттық РНҚ және рибосоманың 5SРНҚ
компоненттерінің транскрипциясын қамтамасыз етеді.
Ақуыз құрамы жайлы ақпарат беретін мРНҚмен бірге, қалған РНҚ
молекулаларының транскрипциясына РНҚ полимераза ІІІ қатысады.
13.
Эукариоттардың РНК-полимеразасы жәнеоның транскрипттері
14.
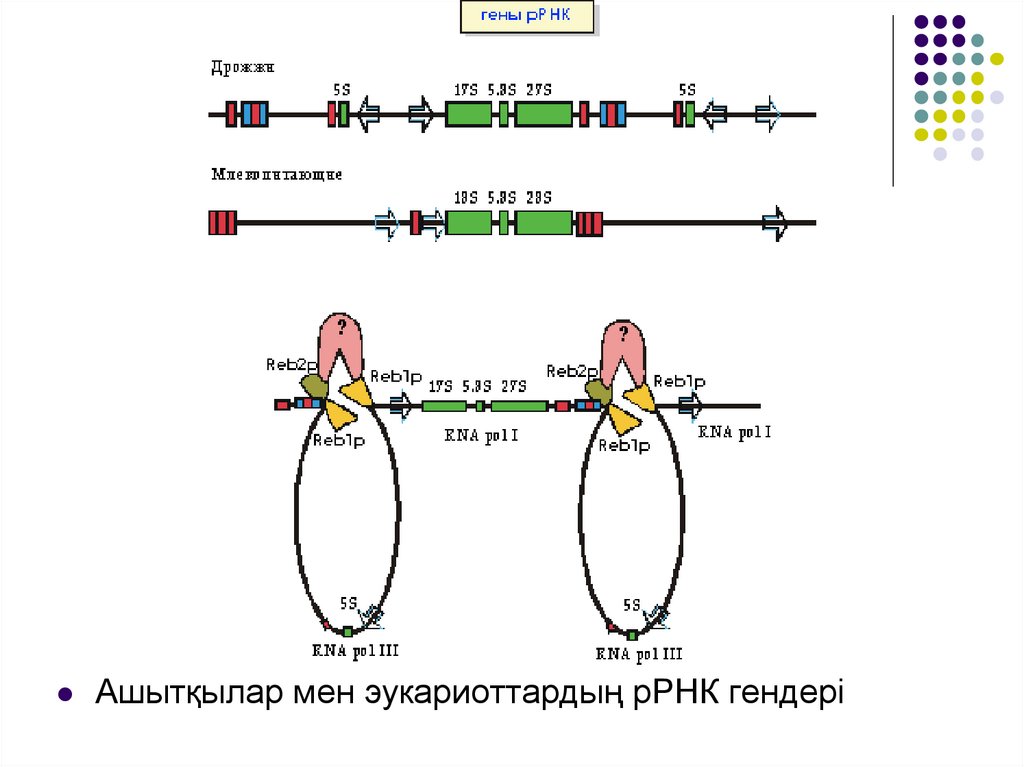
РНҚ полимераза І ядрошықта орналасып, 18S, 5,8S, 28Sгендерін транскрипциялайды.
Бұл РНҚ тізбегі тұтас ізашар ретінде синтезделіп, көп
сатылы бөліну арқылы “жетілген” рРНҚға айналады.
РНҚ полимераза І транскрипцияламайтын жалғыз рРНҚ – 5S
рРНҚ, оның транскрипциясы РНҚ полимераза ІІІ -пен
қамтамасыз етіледі.
рРНҚ ізашарларын кодтайтын гендер жүздеген
транскрипционды бірліктерге дейін жететін тандемді
қайталанулар ретінде орналасады.
рРНҚ гендерінің транскрипциясы мен рибосомалардың
жинақталуы ядрошықта жүреді.
15.
Ашытқыларда 5S рРНҚ гені рРНҚның қайталама ізашаргендерінің арасында қарама – қарсы бағытта орналасады.
Эукариоттарда 5S рРНҚ гені басқа рРНҚ гендерінің
қайталамаларынан тыс орналасады.
рРНҚның транскрипционды бірліктерінің басы мен аяғы
транскрипция терминациясынан кейін бірден реинициациясы
үшін байланысады деп есептеледі.
Ашытқыларда оған Reb1 және Reb2 ақуыздары қатысуы
мүмкін. Олардың байланысы терминатордан кейін, рРНҚ
ізашар генінің промоторының алдында орналасады.
16.
Ашытқылар мен эукариоттардың рРНК гендері17.
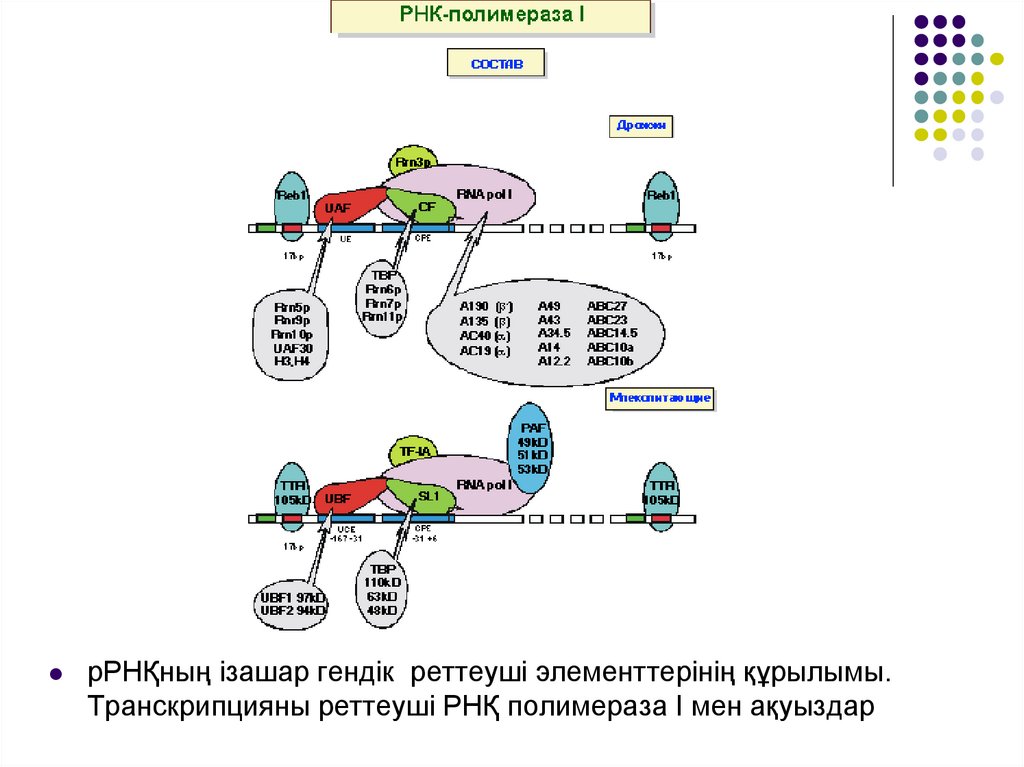
РНҚ полимеразаның суббірліктері І,ІІ,ІІІ түрінеқатысына байланысты А,В,С әріптерімен
белгіленеді.
Кейбір ақуыздар бірнеше РНҚ полимеразаның
құрамына кіріп, екі немесе үш әріппен
белгіленуі мүмкін.
Әріптен кейін осы ақуыз тізбегінің
молекулалық салмағы жазылады.
Мысалы, АС40, бұл РНҚ полимеразада ІІ не ІІІ
суббірлік бар және молекулалық массасы 40
kD тең екендігін көрсетеді.
18.
19.
РНҚ полимераза І –ді тек бір транскрипт синтездейді.РНҚ полимераза І –дің транскрипциондық белсенділігі жасушалық циклдың
фазасына/митоз кезінде тежеледі/ және жасушаның пролиферативтік
белсендігіне/интенсивті бөліну кезінде рРНҚны көп мөлшерде қажет етеді/
байланысты.
РНҚ полимеразаның барлық транскрипциондық аппаратының компоненттері
реттелінеді.
Rm3p (TiF-IA)мен тиімді әсерлесуі үшін РНҚ полимераза фосфорилденуі қажет.
Болжам бойынша, инициацияға қабілетті РНҚ полимераза І мөлшері осылайша
реттеледі.
Негізгі инициация факторларының (SL1 и UBF) белсенділігі жасуша циклының
сатысына сәйкес реттеледі.
SL1 митоз кезінде фосфорилдеу арқылы инактивацияланады, ал UBF керісінше,
G1фазасында фосфорлану арқылы белсенеді.
Белсенді UBF концентрациясы сонымен қатарретинобластомамен
сәйкестендірілген Rb ақуызбен де реттеледі. Rb транскрипцияның күшті ко –
репрессоры болып табылады. Оның тежеуші қабілеті S – фазасында жүретін
фосфорилдену кезінде бәсеңдейді. Осыған ұқсас p53 SL – 1 құрамына кіретін
TBPді инактивациялайды.
РНҚ полимераза І –мен эффективті транскрипцияны қамтамасыз ету үшін, TTFI
ақуызы нуклеосомалардың ДНҚ бойымен қозғлуына жағдай жасайды.
Басқа РНҚ полимеразалар транскрипцияны реттеу үшін гистондар модификациясы
мен хроматиннің конформациялық озгерістерін пайдаланады. РНҚ полимераза І –
дің хроматин құрылымымен байланысы әлі де зерттеуді қажет етеді.
20.
рРНҚның ізашар гендік реттеуші элементтерінің құрылымы.Транскрипцияны реттеуші РНҚ полимераза І мен ақуыздар
21.
РНҚ полимераза ІІІ рРНҚның транспорттық РНҚның және бірнеше кішіядролы РНҚның 5S генін транскрипциялайды. Гендердің бұл үш классы
өздерінің промоторларының қызметін пайдаланады.
рРНҚның 5Sгені ішінде өзіндік промоторы бар, яғни
транскрипцияныңинициациясына қажетті және жеткілікті болатын барлық
аймақтар ДНҚның транскрипцияланушы бөлімінде орналасады.
Бұл фактордың байланысы TFIIIС мен ақуыз комплексінің шөгуіне алып
келеді. TFIIIС ДНҚның А тізбегін байланыстырады, алайда, TFIIIA –сыз бұл
ДНҚ – ақуыздық әркеттестік 5S геннің транскрипциясы үшін жеткіліксіз
болады.
TFIIIС пен рРНҚның 5S гені байланысқаннан кейін оған TFIIIВ қосылады.
TFIIIВ құрамына ТВР мен 2 қосымша ТАFІІІ ақуызы кіреді -В және Brf.
TFIIIВ фактор РНК полимераза ІІ –нің қосылуы мен транскрипция
басталуына жағдай жасайды.
22.
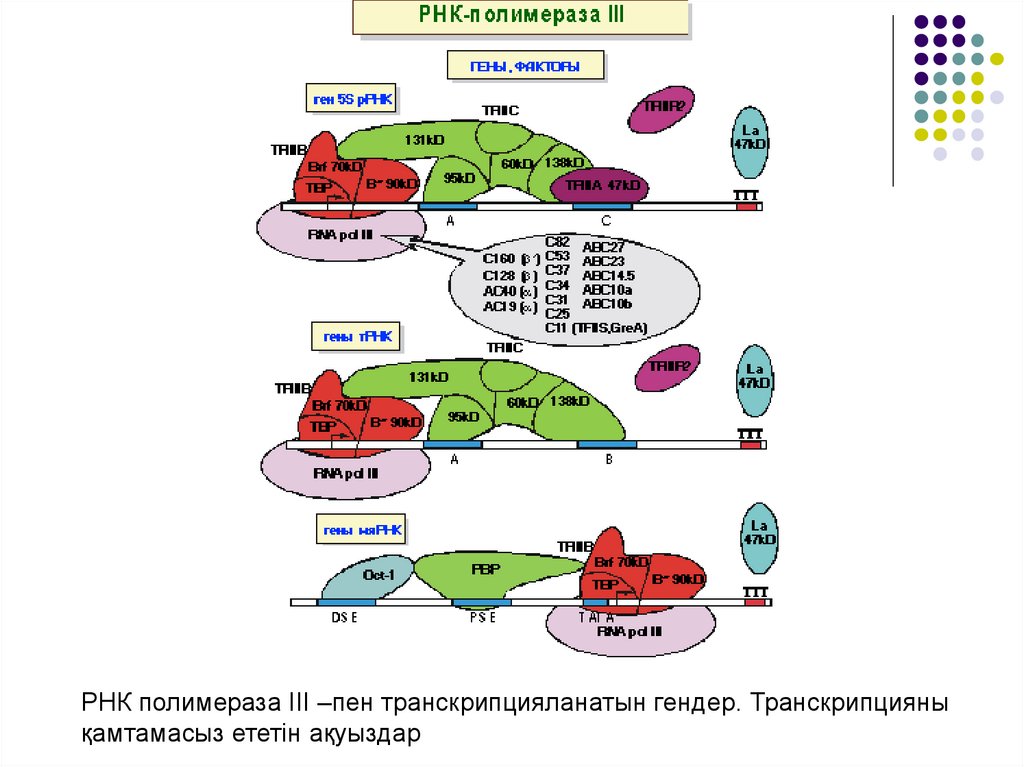
РНК полимераза III –пен транскрипцияланатын гендер. Транскрипциянықамтамасыз ететін ақуыздар
23.
Кіші ядролы РНҚ генінің промоторы мүлдем өзгеше құрылған (РНҚполимераза ІІІ –пен транскрипцияланады). Бұл гендердің барлық
реттеуші элементтері транскрипцияланушы аймақтан тыс орналасқан.
ДНҚның PSE және DSE тізбектері промотордың сәйкес жақын және алыс
орналасқан элементтері болып табылады.
PSE –ге РВВ, ал DSE ақуызына Ost-1 транскрипциялаушы факторы
қосылады. Ost-1 РНҚ полимераза ІІ –мен байланысып, кейбір гендердің
транскрипциясын белсендіреді.
Ost-1 мен DSР кооперативті байланысады және оған ТҒІІІВ келіп
қосылады. РНҚ полимераза промоторларының соңғы классында ғана
ТАТА тізбегі бар.
РНҚ полимераза ІІІ –пен транскрипцияланатын кяРНҚ промоторлары
РНК полимераза ІІ промоторына ұқсас келеді. Сондықтан кяРНҚның
кейбір гендері жағдайға байланысты 2 полимеразамен де синтезделіне
алады.
24.
Барлық 3 типтегі промоторларға ортақмеханизм – басқа ДНҚ байланыстырушы
белсендіруші ақуыздар көмегімен ТҒІІІВ –ның
промоторға келіп шөгуі.
РНҚ полимераза ІІІ –тің де қызметі РНҚ
полимераза ІІ ұқсас клеткалық цикл фазасына
тәуелді реттеледі
25.
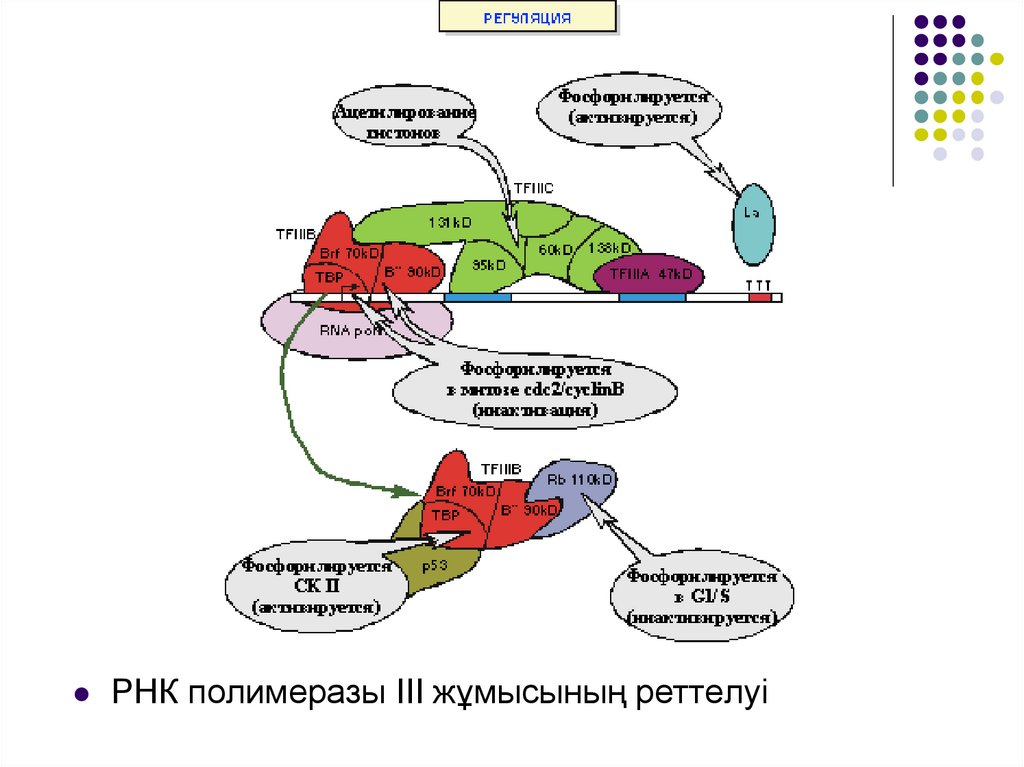
РНК полимеразы III жұмысының реттелуі26.
•ТВІІІВ компоненттері тікелей фосфорилдену көмегіменде, репрессор ақуыз Rb және p53 арқылы да реттеледі. La
ақуызы да фофорилдену кезінде белсенеді. Реттеуші
фосфорилдеу РНҚ полимераза І жағдайындағы сияқты
митоз кезіндетранскрипцияны тежеуге бағытталған.
РНҚ полимераза ІІІ транскрипциясы хроматин
құрылымына сезімтал.
131 kD факторының суббірлігі ТҒІІІС гистондарды
ацетилдеу қабілетіне ие, сол арқылы ол хроматин
құрылымын транскрипцияға қолайлы етіп өзгерте алады.
27.
РНҚ полимераза ІІ РНҚның мРНҚ және кіші РНҚның түрлікласстарын транскрипцилайды. Бұл фермент полимеразаның 3
классының ішіндегі реттелуге ең бейім түріне жатады.
РНҚ полимераза ІІ суббірлігінің ерекшелігі болып оның YSPTSPS
гаптомерлерінің қайталауларынан тұратын – С соңды домені
саналады.
Бұл қайталаулардың саны әр түрлі ағзаларда әр түрлі болып келеді
және ондап саналады. С –соңды доменнің қысқаруы жасуша өліміне
алып келеді. SP қайталануларындағы қалдықты сериндер
фосфорилденуі мүмкін.
28.
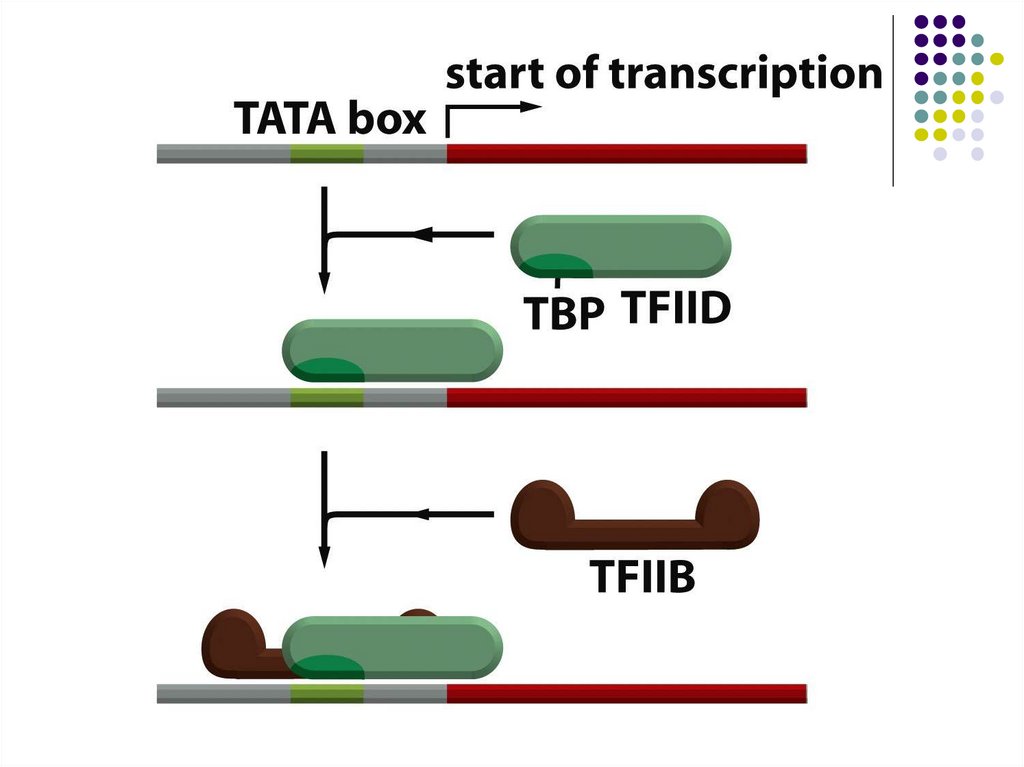
РНК полимераза I және ІІІ сияқты соңғы транскрипция инициация стадиясы
РНК полимераза ІІ промоторға тігілуі,атап айтқанда ТАТА тізбегіне ТВР және
(ТАF II) факторымен ас оцирленген саны жағынан ең көбі.
ТВР және ТАF II ақуыздар комплексі ТFIID деп аталады.
ТFII А және ТFII В ақуыздары осы комплекске тігілуге мүмкіндік туғызады
Одан басқа РНК полимераза ІІ комплексіне және ТЕIF, ТFІІS ақуыздарына
ТFІІВ тігілуге мүмкіндік туғызады.
ТFІІ F ақуыздарының үлкен суббірлігі ТАF 250 фосфорлайды. Бұл үлкен
суббірлік АТФ тәуелді ДНК-хеликаза болады
РНК С-соңды синтезі бастамасынан бұрын,РНК полимераза ІІ домені
дефосфорленген.
РНК полимеразасы қосылған соң,комплекс ТFІІЕ инициаторымен
байланысады.
Өз кезегінде ТFІІН жалғастырады
Бұл бифункционалды ақуызды комплекс,репарацияда және транскрипцияда
роль ойнайды. Оның құрамына : хеликазалар(ДНК-ны екі бағытта (5 және 3)
жергілікті тарқатушы ) және РНК полимераза С-сонды доменді киназасы.
С-сонды домен фосфорланған сон РНК полимераза инициаторлы
комплекске ұқсастығын жоғалтады және РНК синтезі басталады.
СТD басқа ТFІІН циклин-тәуелді киназаларды фосфорлайды: сdk2,4 және
сонымен қатар ТFІІЕ(56 kD)және ТFІІF(74 kD)
29.
30.
31.
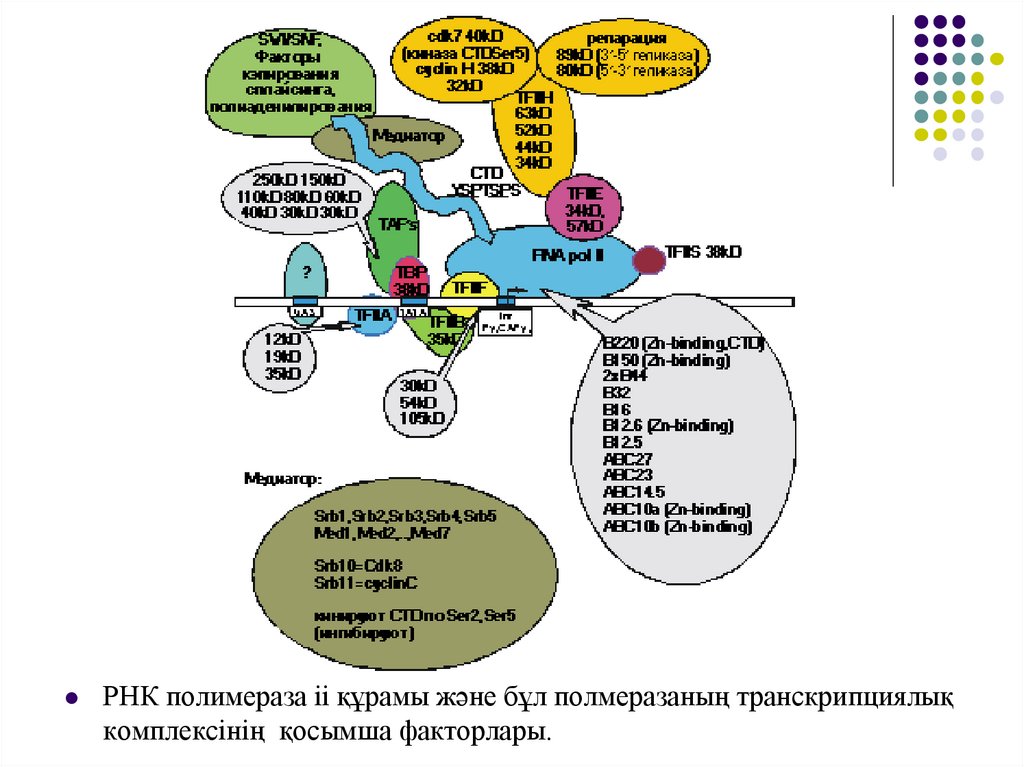
РНК полимераза іі құрамы және бұл полмеразаның транскрипциялықкомплексінің қосымша факторлары.
32.
Жазылған факторлардан басқа РНК полимераза ІІтранскрипция инициациясында басқа да көптегендері
қатысады. Олар әртүрлі гендердің транскрипциасының
реттелуін қамтамасыз етеді.
РНК полимераза ІІ барлық промоторларды бірегей және
реттеуші факторлары өзінің арнаулы комбинациясын қажет
етеді.
Реттеуші фактор промотор аймағында ДНК-мен байланысады
және транскрипцияны активтейді немесе ингиберлейді.
Прокариоттарға қарағанда реттеуші факторлардың
байланысу орны транскрипция инициация нүктесінен
бірталай алшақ орналасуы мүмкін.
33.
Фосфорленбеген (инициация стадиясында) мен РНКполимераза С-сонды домені ақуызды комплекспен
әрекеттеседі, ол медиатор деп аталады.
Кейбір оның ақуызды суббірліктері жеке гендердің
активациясы /ингибирлеуге қажет және тікелей немесе тікелей
емес ДНК байланыстырушы транскрипционды факторлармен
өзара әрекеттеседі.
Жеке ақуыздардын қосылмай қалуы,медиатор
компоненттері,белгілі ген топтарының транскрипциасын
алдын-ала болдырмайды немесе басқа топтың
транскрипциасын күшейтеді.
34.
РНК полимераза ІІ барлық белсенділігін реттеуфосфорленген оның С-соңды домені Ser2 және Ser5
көмегімен іске асуы мүмкін.
Бұл фосфорлану полимеразаны транскрипция
инициациясына қабілетсіз етеді.
Медиатор CDK8 компоненттері және циклин С белгілі
гендер жиынтығына арналған сияқты ингибирлеу шақырады.
Басқаша қарағанда, сол сияқты фосфорлену MPF киназамен
жүзеге асады және митоздың басында транскрипцияның
тоқталуын қамтамасыз етеді.
РНК полимераза ІІ СТD-дан басқа, әр түрлі TFIID
компоненттері реттелуге тап болады. Активаторлар мен
ингибиторлар (тікелей ДНК-байланыстырушы сияқты, ДНК
арқылы байланысқан басқа да ақуыздар) әр түрлі ТАҒІІ
өзара әрекеттеседі. Сонымен қатар жасушалық циклдің әр
түрлі фазасында ТАҒІІ экспрессиясы өзгереді.
35.
Медиатор – бұл ақуызды комплекс, РНКполимераза ІІ С-соңды доменмен
ассоцирленген.
Алғаш ол ашытқыдан табылған және
оның жеке суббірліктері әр түрлі гендер
жиынтығының экспрессиясына қажет
болған немесе РНК полимеразаның Ссоңды қысқаруын супрестеуге
көмектесті.
36.
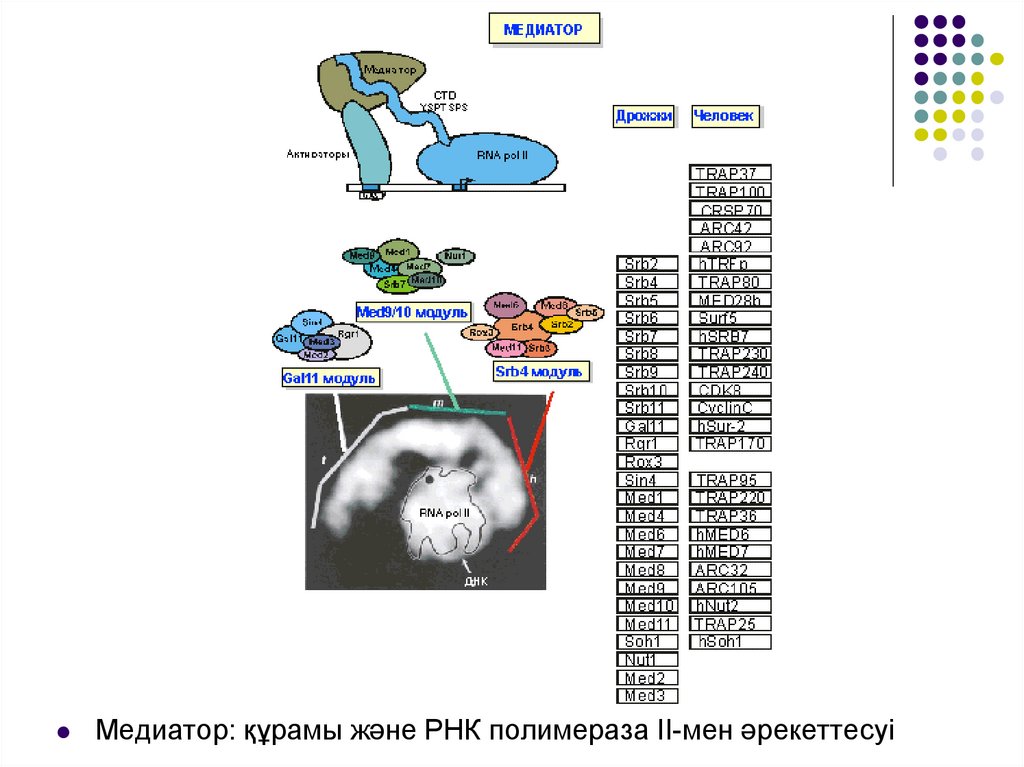
Медиатор: құрамы және РНК полимераза ІІ-мен әрекеттесуі37.
Медиаторда 3 модулді айыру мүмкін болды.Gal11 модулі ДНК байланыстырушы активаторымен немесе
транскрипция ингибиторымен жеке бірлесіп әрекет жасайтын
ақуыздан тұрады.
Med9\10 модулі медиатор бөліктерін және РНК полимераза ІІ-нің Ссоңды доменін байланыстырады.
Srb4 негізінде жасалған модуль РНК полимеразамен өзара әрекет
жасайды және белсенділігін модульдейді.
Қазіргі уақытта медиаторды ақуызды комплекс түрінде
қарастырады, реттеуші сигналдарды интегрирлеуші және РНК
полимеразаға белсенді әсер етеді. Осы уақытқа дейін Srb8-11
ақуыздарынан тұратын 4-ші модульдің табылғаны анық емес.
Олардың арасында циклинге және циклин-тәуелді киназаға сәйкес
CTD-РНК полимеразаға әрекет етуші және белгілі жағдайда оны
инициацияға дейін инактивтейтін Srb10\11 жұбы да бар.
38.
Жоғарғы эукариоттарда медиатордың гомологтарытабылған болатын. Күтуге болатындай, ұқсас
комплекстердің бірнешеуі табылды, сонымен қатар
олардың құрамы бөлшектеп жабылады.
Осы комплекстердің жеке суббірліктердің
мутациялары ашытқы медиаторында сияқты жеке
гендер жиынтығы транскрипциясының бұзылуына
әкеледі, айырмашылығы жоғарғы эукариот
жағдайында бұл өзгерістер, әр түрлі тіндерде және
әр түрлі даму этаптарында әр түрлі болады.
39.
РНК полимераза ІІ-нің элонгационды күйінің басты маркері фосфорленген СTD.Элонгация процесінде фосфорленген күйінің ұсталуына Сdk9
және циклин Т (ашытқыдағы Ctk1, Ctk2 және Ctk3)-дан
тұратын рТЕҒb факторы жауап береді. Бұл фактордың
инактивациясы белгілі бір жинақталған гендердің
транскрипциясына әсер етеді.
рТЕҒb-дан басқа, РНК полимераза ІІ элонгациясының
факторы гетеротример элонгин болып табылады. Бұл
фактордың теріс реттеушісі – VHL, элонгин В және С
комплекстерін байланыстырып, олардың элонгин А-мен
ассоциациясын болдырмайды.
40.
РНК полимераза ІІ транскрипциясының элонгациясы41.
Элонгация процессінде РНК полимеразасы прокариоттың РНКполимеразасы сияқты мәселесімен кездесуі мүмкін: ДНК
зақымдалуы, пауза туғызатын және “тұтқындалған” күйге
түсіретін бөлімдер.
Егер жұмысшы РНК полимераза репарация қажет ететін ДНК
зақымдануын кездестірсе, онда сондай тоқталған комплекспен
СSА және CSB ақуыздары байланысады. Бұл ақуыздар
зақымдалған репарация факторының тігілуін хабарлайды және
репарациядан кейін транскрипцияның қалпына келуінде басты
роль ойнайды. СSА және CSB мутациялары кокейн синдромын
шақырады.
ДНК полимераза кідірісінің супрессиясына ТҒІІҒ, ал
“тұтқындалған” күйден шығуға ТҒІІS жауап береді.
Транскрипция элонгациясын қамтамасыз етуші факторлардан
басқа, фосфорланған РНК полимераза ІІ формасы пре-мРНК
процессингінде қатысатын ақуыздармен байланысады.
Олардың арасында терминация транскрипциясына жауап
беретін кесу-полиаденилдеу аппараты да бар.
42.
43.
44.
45.
Эукариоттыф клеткалардын инициациясы басталуы үшінДНҚ-ның транскрипциялық факторы керек.
Эукариоттарда транскрипцияның in vivo басталуы үшін
белокты комплектерді қажет етеді, оны медиатор деп
атайды. Ол активаторлы белоктар мен полимераза ІІ жалпы
факторлармен байланыстырады
Эукариотты клеткалардың транскрипцияның инициациясына
рекрутирлеуші модификациялаушы хроматин ферменттер
қажет, сонымен қоса хроматинді қайта тузү комплектері
және гистонды-модификациялаушы ферменттерді қажет
етеді.