
 белгілі нүклеотидтер бірізділігі.")











 хромосома 1- хроматида, (S-фазадан кейінгі пайда болған екі бірдей хроматидтер); 2 – центромера,")
 және q (ұзын). Метафазалық хромосоманың морфологиялық 4 түрі бар: метацентрикалық,")



")


")




































 medicine
medicineSimilar presentations:









")
Жасушаның генетикалық аппараты. Ген туралы түсінік
1.
Тақырыбы:. Жасушаның генетикалықаппараты. Ген туралы түсінік
Мақсаты: Эукариот клеткасы
хромосомасының химиялық ұйымын, гендердің
негізгі қасиеттерін оқып білу.
Дәрістің жоспары:
1. Гендердің негізгі қасиеттері.
2. Гендердің жіктелуі.
3. Хромосоманың химиялық құрамы.
4. Гистондардың сипаттамасы.
5. Гистонды емес ақуыздардың сипаттамасы.
6. Гистондардың қызметі.
7. Гистонды емес ақуыздардың сипаттамасы.
2. Ген – тұқымқуалаушылықтың элементарлы бірілігі (нақты белгіні анықтайтын ДНҚ бөлігі) белгілі нүклеотидтер бірізділігі.
3.
1. Тұрақты - ген құрылымының сақталуы,мутацияға сирек ұшырау ерекшелігі.
2. Спецификалық (өзгешелігі) - әрбір
гендегі нуклеотидтердің орналасуы нақты
және белгілі полипептид синтезін
анықтайды.
3. Лабильділігі мутацияға немесе
өзгеруге қабілетті.
4. Экспрессивтік - белгінің көріну дәрежесі
немесе берілген геннің фенотипті көріну
дәрежесі.
4.
5. Пенетранттық - геннің немесе белгініңкөріну жиілігі.
6. Тұтас ген - полипептид синтезін
бағдарлау кезінде ген бөлінбейтін бірлік
болып табылады.
7. Дискреттік - гендегі суббірлікті анықтайды:
минимальді құрылымды бірлігі–
нуклеотидтердің комплементарлы жұптары,
минимальді функциональді бірлігі-кодон.
8. Плейотроптық - бір ген бірнеше белгінің
түзілуіне жауап береді.
5.
9. Аллельділік - диплоидтық ағзагенотипінде гендердің тек екі түрі
болады.
10. Амплификация - геномда кейбір
гендердің көптеген көшірмелері
болады.
11. Мөлшерлік - ген анықталған,
шектелген мөлшерге дейін белгілердің
дамуын қамтамасыз етеді.
6. Эукариот генінің жіктелуі
I. Құрылымдық гендер- Ақуыздарды кодтайтын гендер,
тРНҚ, рРНҚ (300-1600 рет қайталанады).
II- Реттеуші гендер - Құрылымдық гендердің
белсенділігін реттейді.
III. Миграцияланатын генетикалық элементтер –
транспозондар – геном бойынша жылжиды,
ферменттердің көмегімен көрші гендердің
белсенділігіне әсер етеді.
IV. Псевдогендер- интрондары жоқ, құрылымдық
гендердің көшірмесі, транскрипцияға
қатыспайды.
Адам геномында ақуызды кодтайтын 20-25 мың ген
бар. Алтернативті сплайсингтің
нәтижесінде бір геннен бірнеше әртүрлі
ақуыздарды құрастырады.
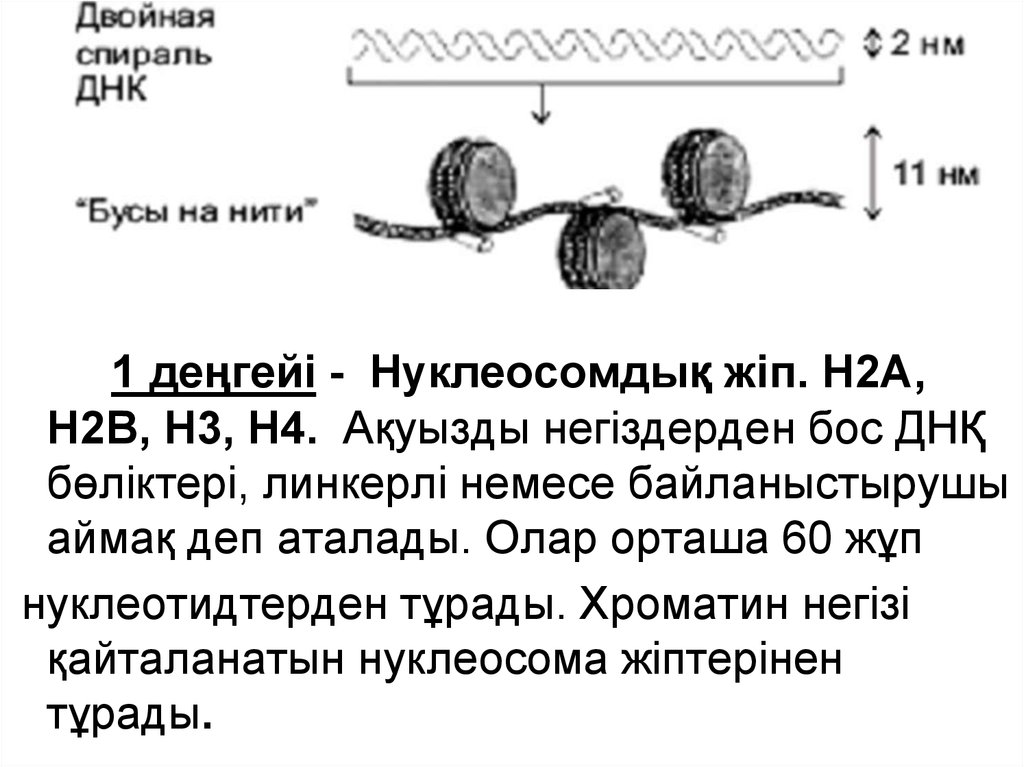
7. Хромосомадағы ДНҚ ұйымы
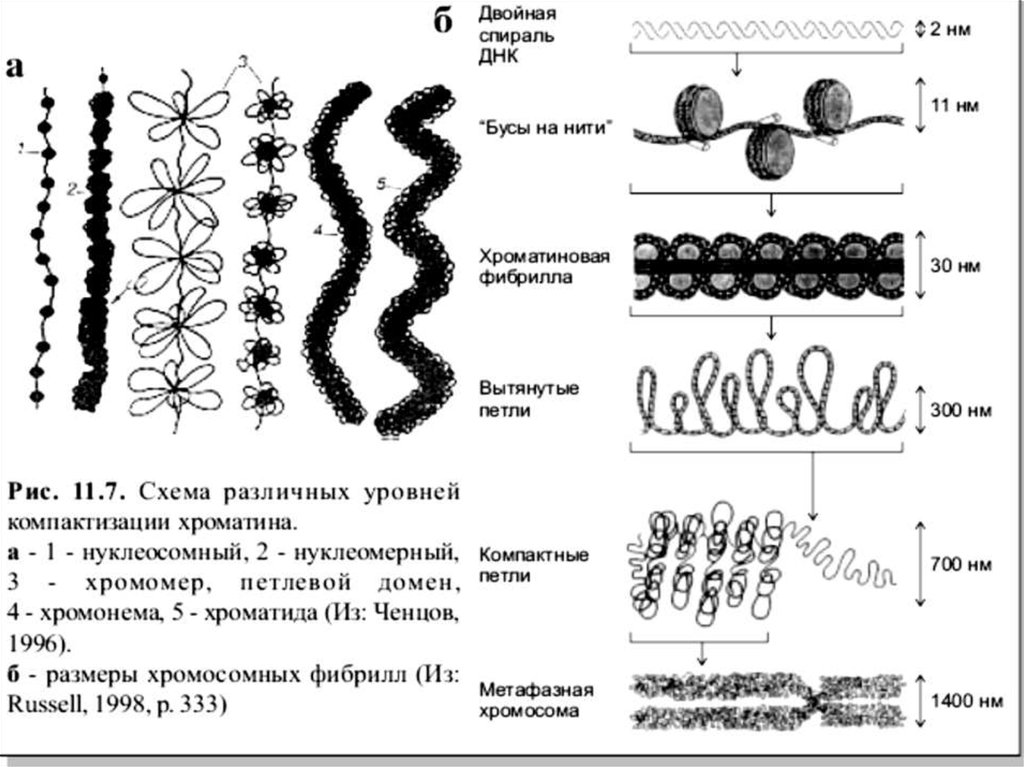
1 деңгейі - Нуклеосомдық жіп – d -10нм.Хроматиннің бұл құрылымдық деңгейіне
келесі гистондар қатысыды: Н2А, Н2В,
Н3, Н4.
Олар ақуызды дене немесе негіздерді
құрайды, 8 молекуладан тұрады (әрбір гистоннан
2 молекуладан). ДНҚ молекуласы ақуыздық
негіздерде спиральді оралады.
Ақуызды негіздерде 146 жұп нуклеотидтерден
тұратын ДНҚ бөлігі болады.
8.
1 деңгейі - Нуклеосомдық жіп. Н2А,Н2В, Н3, Н4. Ақуызды негіздерден бос ДНҚ
бөліктері, линкерлі немесе байланыстырушы
аймақ деп аталады. Олар орташа 60 жұп
нуклеотидтерден тұрады. Хроматин негізі
қайталанатын нуклеосома жіптерінен
тұрады.
9.
Нуклеосомдық - электрондық микроскоппенкөргенде «жіптегі моншақтарға» ұқсайды.
10.
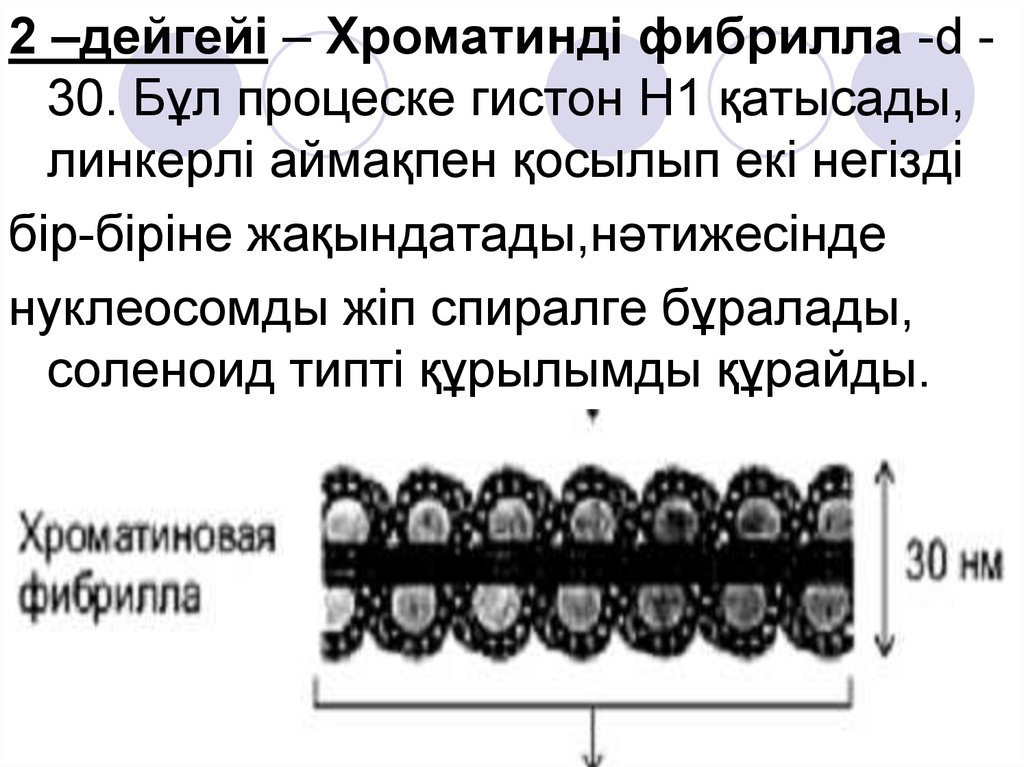
2 –дейгейі – Хроматинді фибрилла -d 30. Бұл процеске гистон Н1 қатысады,линкерлі аймақпен қосылып екі негізді
бір-біріне жақындатады,нәтижесінде
нуклеосомды жіп спиралге бұралады,
соленоид типті құрылымды құрайды.
11.
3 – деңгейі– Интерфазалық хромонема.Бұл этап ілмекті, күрделі. Соленоидты
фибрилла әртүрлі ұзындықтағы ілмекті ( 50 000
– 200 000 мың жұп нуклеотидтерге дейін)
құрастырады. Хроматинді фибрилла
нәтижесінде 300 нм диаметрге дейінгі құрылым
түзіледі, ол интерфазалы хромонема деп
Аталады.
12.
Хромонемалар қосымша жинақталады,диаметрі 600-700 нм хроматидтерді
құрайды. Хроматидтер метафазалы
хромосома құрамына кіреді, диаметрі 1400
нм немесе 1,4 мкм.
13.
14. Хроматиннің жинақталу этаптары
15.
Кариотип дегеніміз – хромосоманың өлшемімен,түрлерімен және санымен сипатталатын жасушаның
хромосомалық жинағы. Кариограммада гомологиялық
хромосомалардың ұзындығы мен центромераларының
орналасуы бойынша орналасқаны көрсетілген (Денверлік
жіктелу, 1964 ж.).
16.
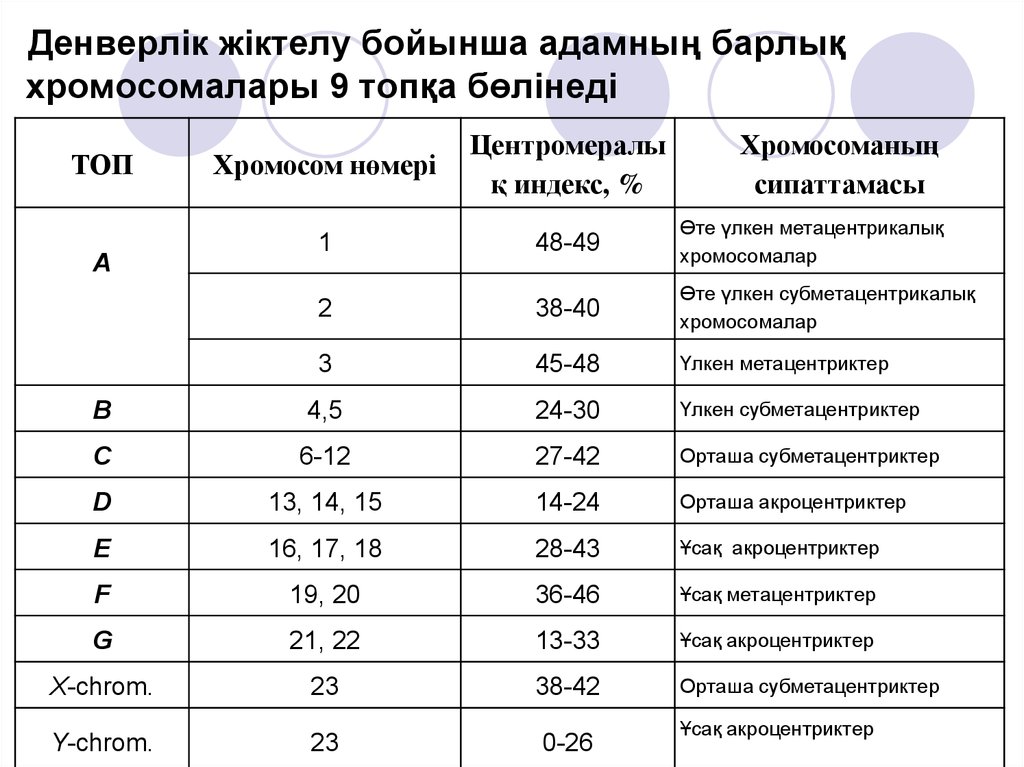
Денверлік жіктелу бойынша адамның барлықхромосомалары 9 топқа бөлінеді
Хромосом нөмері
Центромералы
қ индекс, %
1
48-49
Өте үлкен метацентрикалық
хромосомалар
2
38-40
Өте үлкен субметацентрикалық
хромосомалар
3
45-48
Үлкен метацентриктер
В
4,5
24-30
Үлкен субметацентриктер
С
6-12
27-42
Орташа субметацентриктер
D
13, 14, 15
14-24
Орташа акроцентриктер
Е
16, 17, 18
28-43
Ұсақ акроцентриктер
F
19, 20
36-46
Ұсақ метацентриктер
G
21, 22
13-33
Ұсақ акроцентриктер
X-chrom.
23
38-42
Орташа субметацентриктер
Y-chrom.
23
0-26
ТОП
А
Хромосоманың
сипаттамасы
Ұсақ акроцентриктер
17.
Хромоссомалардың париждік
жіктелуі (1971
ж.).
Хромосоманы
дифференциалді
бояумен бояған
кезде олардың
көлденең
жолақтары жақсы
көрініп тұр
18. Конденсирленген (метафазалы) хромосома 1- хроматида, (S-фазадан кейінгі пайда болған екі бірдей хроматидтер); 2 – центромера,
Конденсирленген(метафазалы)
хромосома
1- хроматида, (Sфазадан кейінгі пайда
болған екі бірдей
хроматидтер);
2 – центромера,
хроматидтердің
қосылатын орны және
микротүтікшелердің
бекінуі;
3- қысқа иықтар;
4-ұзын иықтар
19. Әрбір хромасома екі иықтан тұрады: р (қысқа) және q (ұзын). Метафазалық хромосоманың морфологиялық 4 түрі бар: метацентрикалық,
субметацентрикалық, акроцентрикалық жәнетелоцентрикалық.
20. Эукариот ядросындағы хромосомалар деконденсирлен-ген жағдайда болады. ДНҚ гистонды ақуыздармен қоршалып, хроматинді түзеді. Екі
Эукариотядросындағы
хромосомалар
деконденсирленген жағдайда
болады. ДНҚ
гистонды
ақуыздармен
қоршалып,
хроматинді түзеді.
Екі түрге бөлінеді:
эухроматин және
гетерохроматин.
21. Политенді хромосомалар гомологтардың конъюгациясы, олардың репликация кезінде хромосомалардың ары қарай ажырамау нәтижесінде
Политендігомологтардың
репликация
ары
қарай
хромосомалар
конъюгациясы,
олардың
кезінде
хромосомалардың
ажырамау
нәтижесінде
пайда
болады.
22. Гендердің 23% экспрессияға, репликацияға және геномның қызметін ұстап тұруға жауап береді; 20% - жасуша аралық сигналды беруге,
17% - жасуша қалыпты жұмыс істеуі үшін, ал басқа белсендік үшінжауап беретін гендердің жұмысы белгісіз.
23. Ауру гендері (2005ж)
2001 жылы адам геномында 1112 “ауругендері” анықталынды (яғни, мұндай
гендердің мутациясы ауруларға алып
келеді);
Бұл гендерден 44,500 мутация
табылды (орташа 1 генге 26),
аурулармен байланысты;
Қалғандарында-шамамен 10 млн.
мутациясы бар, бірақ олар аурулармен
байланысы жоқ.
24. Гендердің экспрессиялы реттелуі-
бұл транскрипция (сайт) басталатынаймақта орналасқан ақуыздардың, әртүрлі
ДНҚ бөліктерімен өзара спрецификалы
байланысуы.
- Гендердің экспрессиясы (белсенділігі)
уақытқа байланысты жүреді, яғни гендер
қосылып және ажырап отырады немесе
гендер белсенді немесе белсенсіз халде
болады.
- Реттеуші ретінде жиі ақуыздар, кейде
металдар және басқа төменгі
молекулярлық қосылыстар жатады.
25. Эукариот гендері жұмысының реттелуі келесі кезеңдерден тұрады:
1.2.
3.
4.
Ген жұмысының реттелуі келесі этаптардан
тұрады:
транскрипциялық
посттранскрипциялық (сплайсинг,
полиаденирлену)
м-РНҚ-ның ядродан цитоплазмаға шығу
деңгейі
Трансляциялық
26. Прокариот генінің экспрессиялы реттелуі Оперон моделі (1961 Жакоб және Моно)
Прокариоттарда реттелудің 2-і типібар:негативті және позитивті.
Лак-оперон – позитивті реттелу
мысалы ( ақуыз-репрессор
әрдайым жұмыс халінде болады).
Триптофанды-оперон – негативті
реттелу мысалы (репрессор
белсенсіз халде болады)
27.
28. ЛАК-опероны
29. Триптофан опероны
30.
Эукариотгенінің
экспрессиялық
реттелуінің
этаптары
РЕТТЕЛУДІҢ
ҚОСЫМША
ДЕҢГЕЙІ
(ЭНХАНСЕРЛЕР,
САЙЛЕНСЕРЛЕР,
Ada-зона)
РЕТТЕЛУДІҢ
БАЗАЛЫҚ
ДЕҢГЕЙІ
(Предпромотор
+
промотор)
ҚҰРЫЛЫМДЫҚ
ГЕН
31. Эукариот генінің экспрессиялы реттелуі
Эукариоттарда гендердің белсенділігі 2-іреттеуші аймақтан тұрады: базалы және
қосымша
Базалы элементтің реттелу деңгейі:
промотор алдындағы бөліктен, промотордан
(ТАТА-бокс және ЦААТ-бокс) тұрады.
Қосымша элементтердің реттелу деңгейі:
энхансерлер, сайленсерлер, адаптивті
жауап.
32. Эукариоттағы экспрессияның ерекшеліктері:
Эукариот жасушасында гистондарақуыз-репрессор ролін атқарады.
Экспрессия нәтижесінде, ядрода
көптеген ақуыздар туралы ақпараты бар
м-РНҚ молекуласы синтезделінеді.
Содан кейін, м-РНҚ-ның бір бөлігі
ыдырайды, ал қалған бөлігі
альтернативті сплайсингке ұшырайды.
33.
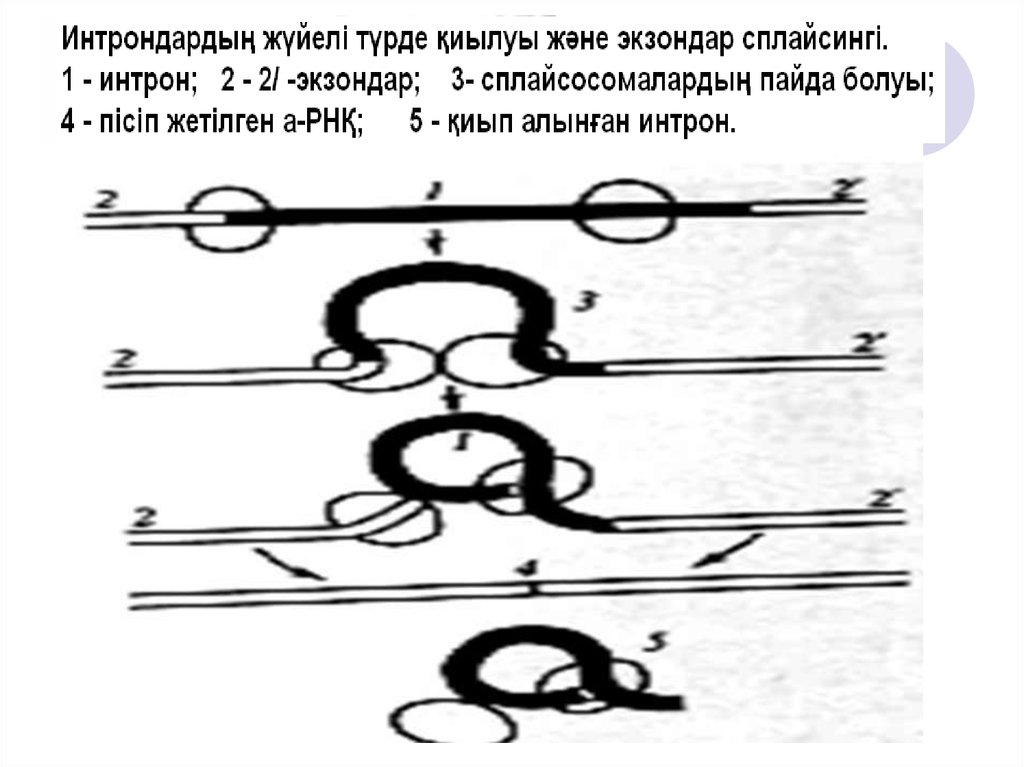
34. м-РНҚ АЛЬТЕРНАТИВТІ СПЛАЙСИНГІ
35. Ақуыз синтезінің этаптары:
Транскрипция (инициация, элонгация,терминация, РНҚ-транскриптат синтезі)
Процессинг (кэпирлену, м-РНҚ-ның
полиаденилденуі)
Сплайсинг (интрондарды қиып тастау)
Трансляция (рибосомада, полипептидті
тізбекке аминқышқылын жинақтау)
36. Процессинг және сплайсингтен кейінгі РНҚ-транскриптаттың құрылысы:
Процессинг және сплайсингтен кейінгітранскриптаттың құрылысы:
Кэп
АУГ
(старткодон)
Экзон
Экзон
Экзон
UAA
Стопкодон
РНҚ-
А-А-А-Ақұйрық
37.
38.
ҚММУМолекулярлық биология және
медициналық генетика кафедрасы
Дәріс
Тақырыбы: «Нуклеин қышқылдарының
биосинтезі. ДНҚ репликациясы».
Мамандығы: 5В130100 «Жалпы
медицина»
Курс: 1
Уақыты: 50 мин.
Дәріскер: б.ғ.к. Есілбаева Б.Т.
39.
Тақырыбы: «Нуклеин қышқылдарыныңбиосинтезі. ДНҚ репликациясы».
Мақсаты: ДНҚ синтезін, оның ерекшеліктерін
және маңызын оқып білу.
Дәрістің жоспары:
1. Репликация механизмі.
2. Репликон, репликациялық вилка (аша) туралы
түсінік.
3. Ферментті комплекстің компоненттері.
4. Жетекші тізбектің және Оказаки
фрагменттерінің синтезі.
5. Теломерлер және теломердің қызметі.
6. Репарацияға түсінік.
40.
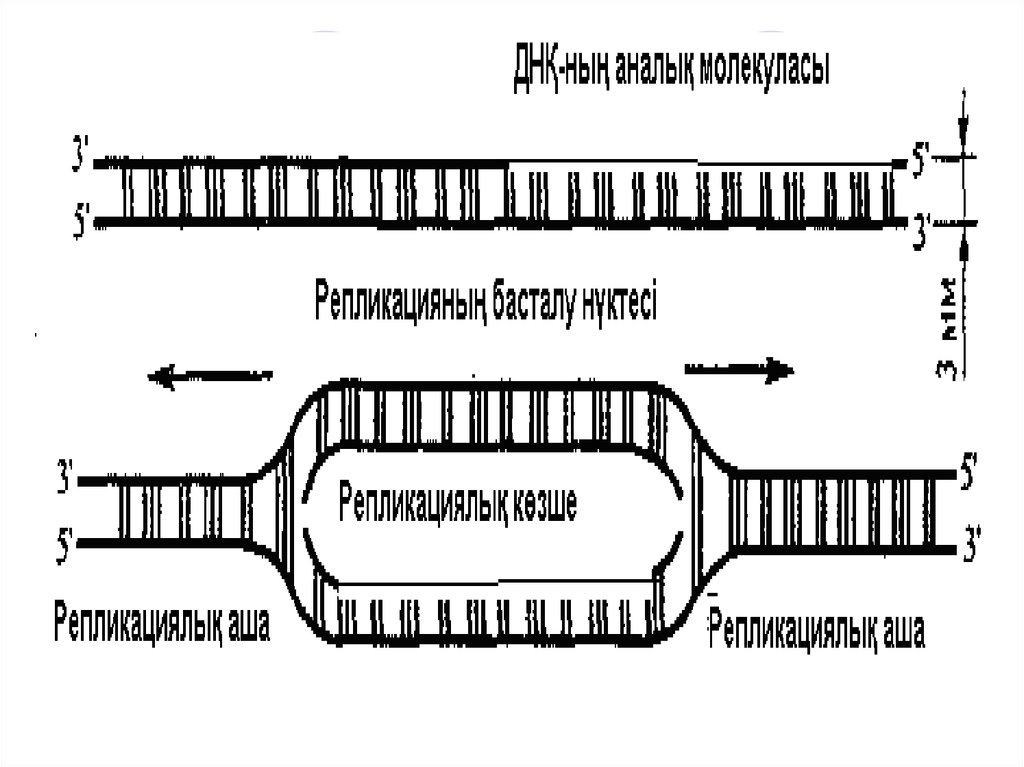
ДНҚ репликациясы.Репликация – дегеніміз ядроішілік маңызды
процесс. Бұл өзін-өзі көшіруге қабылетті,
ДНҚ санының екі еселенуіне алып келеді.
Репликация екі комплементарлық тізбектен
тұратын, ДНҚ молекуласының химиялық
ұйымы ерекшелігінің нәтижесінде болады.
Репликация процессінде аналық ДНҚ
молекуласының әрбір полинуклеотидті
тізбегінде оған комплементарлы тізбек
синтезделінеді.
Нәтижесінде, екі спиральді ДНҚ-ң
біреуінен, екі ұқсас қос спираль пайда
41.
Репликация – дегеніміз ядроішілік маңыздыпроцесс. Бұл өзін-өзі көшіруге қабылетті,
ДНҚ санының екі еселенуіне алып келеді.
Репликация екі комплементарлық тізбектен
тұратын, ДНҚ молекуласының химиялық
ұйымы ерекшелігінің нәтижесінде болады.
Репликация процессінде аналық ДНҚ
молекуласының әрбір полинуклеотидті
тізбегінде оған комплементарлы тізбек
синтезделінеді.
Нәтижесінде, екі спиральді ДНҚ-ң
біреуінен, екі ұқсас қос спираль пайда
болады.
42.
43.
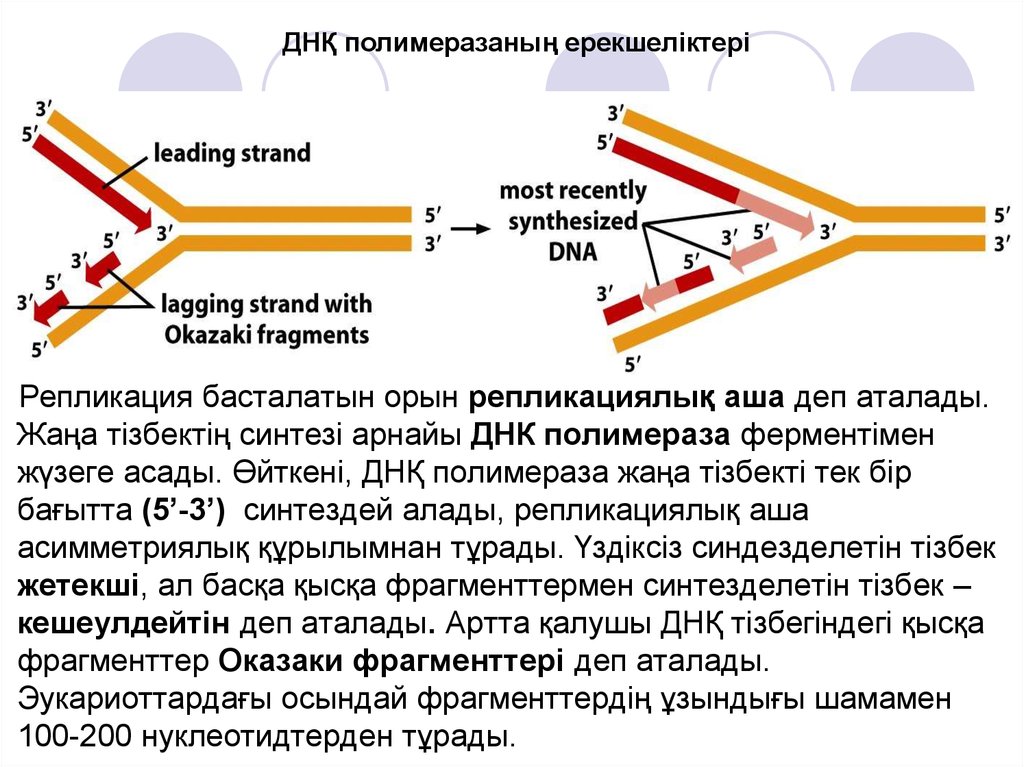
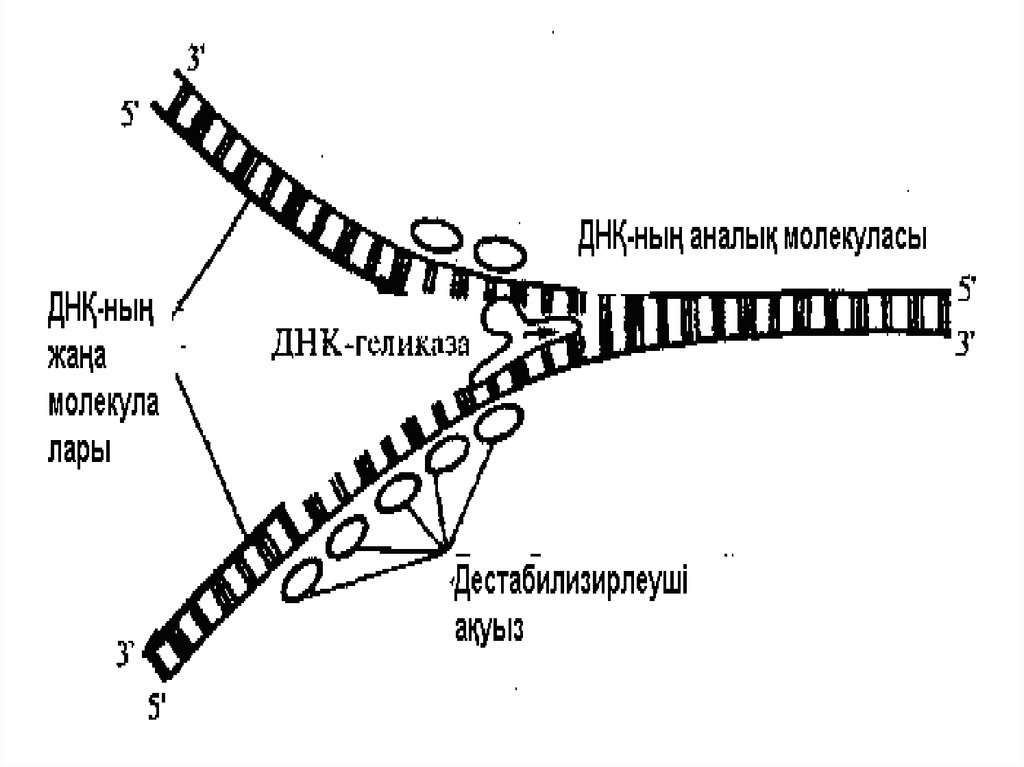
ДНҚ полимеразаның ерекшеліктеріРепликация басталатын орын репликациялық аша деп аталады.
Жаңа тізбектің синтезі арнайы ДНК полимераза ферментімен
жүзеге асады. Өйткені, ДНҚ полимераза жаңа тізбекті тек бір
бағытта (5’-3’) синтездей алады, репликациялық аша
асимметриялық құрылымнан тұрады. Үздіксіз синдезделетін тізбек
жетекші, ал басқа қысқа фрагменттермен синтезделетін тізбек –
кешеулдейтін деп аталады. Артта қалушы ДНҚ тізбегіндегі қысқа
фрагменттер Оказаки фрагменттері деп аталады.
Эукариоттардағы осындай фрагменттердің ұзындығы шамамен
100-200 нуклеотидтерден тұрады.
44.
45.
46.
Прокариот және әукариоттардарепликацияның бірізділігінің болуы
ұқсас, бірақ ДНҚ-ң синтезделу
жылдамдылығы прокариоттарда жоғары
(1000 нуклеотидтер /сек), ал
эукариоттарда төмен (100
нуклеотидтер /сек). Неге бұлай болады?
Репликацияның жоғарғы жылдамдылығының
болуы ферменттер жүйесінің қатысуымен жүреді:
геликазалар, топоизомеразалар,
дестабилизирлеуші ақуыздар, ДНҚполимеразалар және т.с.с.
47.
48.
Кешеулдейтін тізбек қысқа фрагмент түріндеболады (1500 мың нуклеотидтерге дейін). Бұл
Оказаки фрагменттері.
Прокариот жасушасында Оказаки фрагменттері
1000-2000 нуклеотидтерден, ал эукариотта 100200 нуклеотидтерден тұрады.
Жетекші тізбек ұзын үздіксіз фрагмент түрінде
болады, (сперматогоний үшін – бұл 1млн. 600
мың нуклеотидтер).
Әрбір осындай фрагменттің синтезделуі кезінде
ұзындығы шамамен 10 нуклеотидтен тұратын
ұйтқы РНҚ -пайда болады.
49.
50.
51.
Репарация механизміГенетикалық материалдың тұрақтылығын
қамтамасыз ету үшін репликацияның өте
дәл механизмі жеткіліксіз. Өйткені, күнделікті
ДНҚ молекуласында әртүрлі факторлармен
шақырылатын (жылудың, радиацияның,
әртүрлі метоболиттердің және т.б. әсерінен)
мыңдаған әртүрлі өзгерістер болып тұрады.
Адамның әрбір жасушасының ДНҚ
спонтанды депуринизация салдарынан
шамамен 5000 пуриндік негіздерді
молекуласындағы екі комплементарлық
тізбектердің болуына негізделген.
52.
Нуклеотидтердің бірізділігінің бұзылуыферменттермен анықталынып, бұл бөлік
қиылып алынады, ДНҚ-ң екінші
комплементарлы тізбегіндегі жаңа
синтезделген фрагментпен ауыстырылады.
Жасушада күніне цитозиннің урацилге
спонтанды дезаминирлеу жылдамдығы
шамамен 100 негіздер болады. Бірақ,
репарация жүйесін құрайтын ферменттердің
жұмысының арқасында1000 –ның тек 1 –
еуінде, кездейсоқ өзгерістер мутацияға
айналады
53.
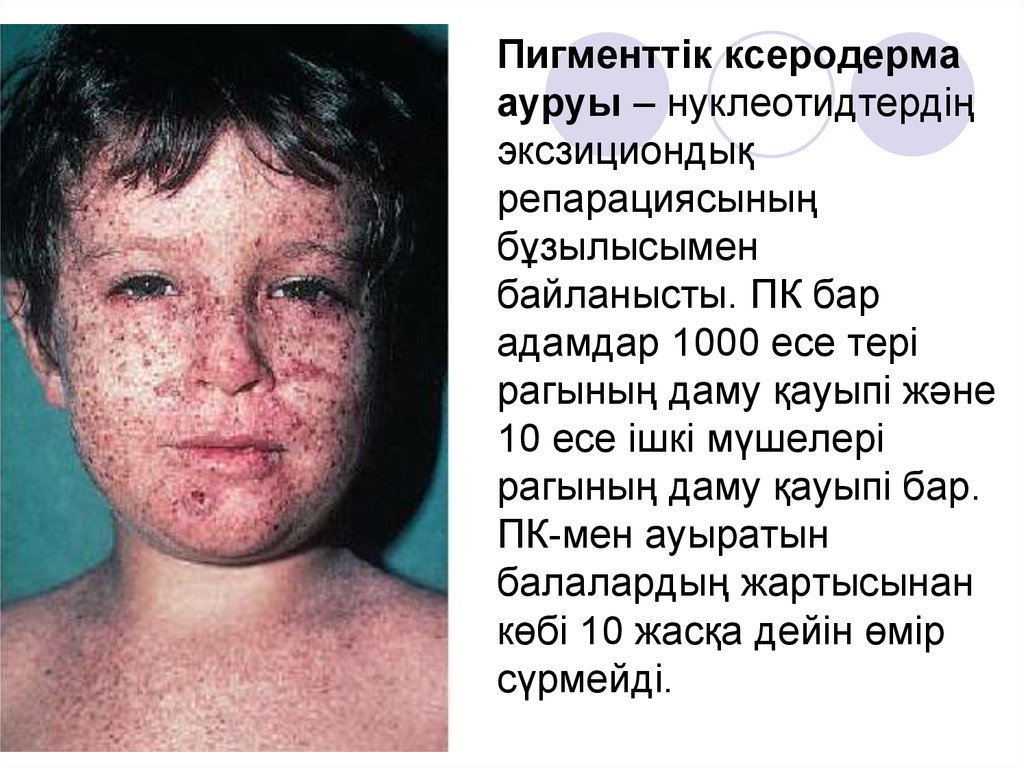
Пигменттік ксеродермаауруы – нуклеотидтердің
эксзициондық
репарациясының
бұзылысымен
байланысты. ПК бар
адамдар 1000 есе тері
рагының даму қауыпі және
10 есе ішкі мүшелері
рагының даму қауыпі бар.
ПК-мен ауыратын
балалардың жартысынан
көбі 10 жасқа дейін өмір
сүрмейді.
54.
Тақырып: Гомеостаз бұзылысы.Мақсаты: гомеостаз механизмдерін және
гемеостаз ауруларын оқып білу
Дәріс жоспары:
1. Гомеостаздың механизмдері және
биологиялық маңызы.
2. Гомеостаз аурулары.
55.
Ағза өзінің ішкі тепе-теңдігін сыртқы ортажағдайының өзрегісіне сақтай алатын
қабылеттілігін – гомеостаз деп атаймыз.
Жылы қанды жануарларда көптеген көрсеткіштер
сақталады. Мысалы, O2, CO2, плазманың pH,
глюкозаның және K+ концентрациясы және т.б.
Көп жасушалы ағзалардың гомеостазы –
тұрақтылықты сақтау, өмур сүру, ортаның
өзгерісіне бейімделу және даму болып табылады.
56.
Гомеостаздың механизмдері:Барлық гомеостаздық механизмдер бір-біріне
тәуелді бірнеше механизмдерден тұрады.
Рецептор – өте сезімтал кешене, сыртқы ортаның
өзгерісін қадағалайды және оларға жауап береді.
Рецептордан алған Сигнал ары қарай бақылау
орталығына беріледі (көптеген гомеозтаздық
механизмдер көбінесе миға беріледі.
Бақылау орталығы ары қарай сигналды
эффекторға (бұлшық-еттер, мүшелер немесе басқа
құрылымдар) жібереді. Сигналды қабылдағаннан
кейін ауытқуларды коррекциялайтын гомеостаздық
жағдайдан позитивті немесе негативті кері
байланыс, өзгерістер жүреді.
57.
Оң кері байланыс механизмі сыртқы ортажағдайының өзгерісіне жауап беру
реакциясын жоғарылатады,
дестабилизирлеуші эффект көрсетеді.
Теріс кері байланыс механизмі жиі жеке
мүшелердің белсенділігін төмендетеді. Бұл
механизм ағзаның ішкі ортасының тұрақты
сақталуына бағытталады.
58. Қандай механизмдер дененің температурасын төмендетеді?
1. Тер шығаруБіздің денеміз қызған кезде бездерден
тер бөлінеді.
Тер газға айналады (буланады)
Булану кезінде дененің қызуы
қолданылады.
Дененің беткейі суыйды.
59. Тер бөліну
The skin60.
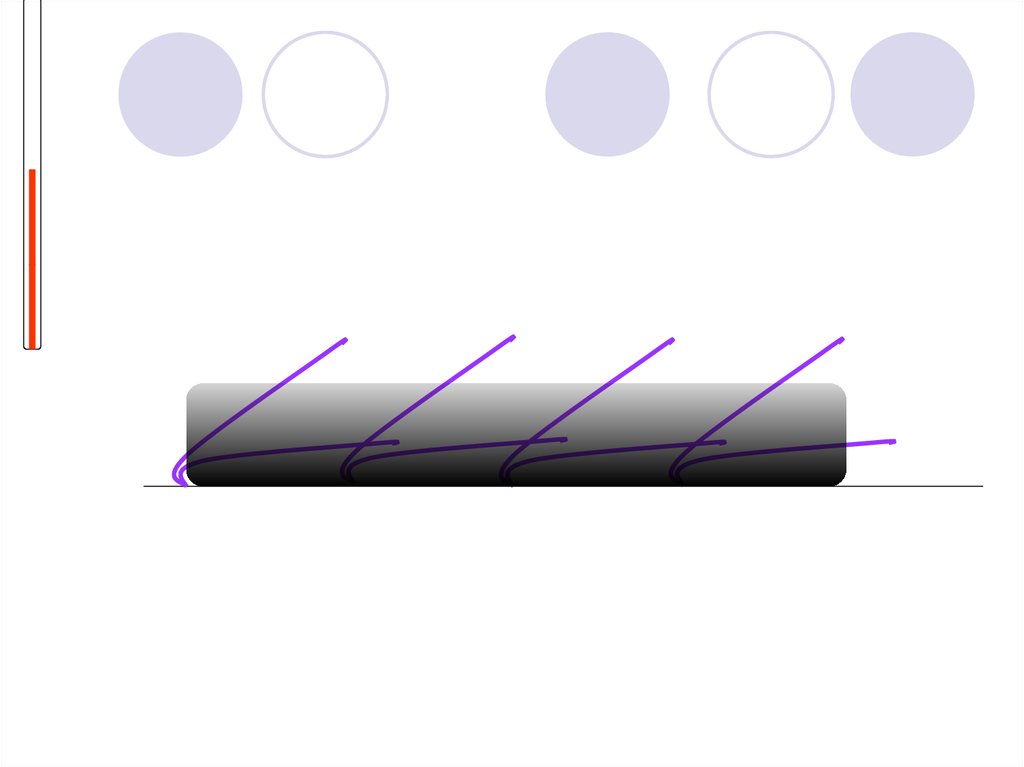
2. Тамырлардың кеңеюі.Біздің қанымыз жылу энергиясының көп бөлігін денеге
тасымалдайды.
Тері астында капилларлар бар, олар дене жылыған
кезде кеңейеді.
Осылай, ағза сыртқы ортаға жылуды көптеп шығарады.
Сондықтан да, ыстықтаған кезде қызарып кетеміз.
61.
Демек, тері беткейінен сыртқы ортаға көп жылубөлінеді
Егер
температура
жоғарыласа,
веналар
кеңейеді
62. Қандай механизмдер дененің температурасын көтереді?
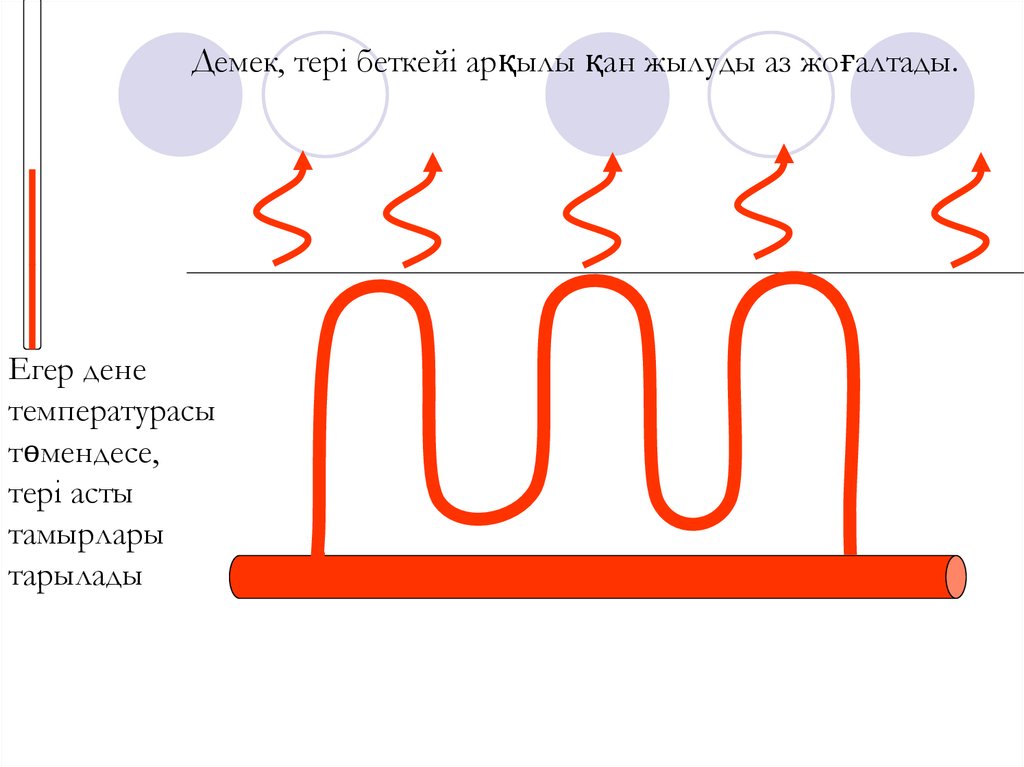
1. Тамыр саңлауларының төмендеуі.Венаның кеңеюіне қарама-қарсы механизм.
Тері астындағы капиллярлар жиырылады.
Сондықтан да, қан жылуды тері беткейі арқылы сыртқы
ортаға аз бөледі.
63.
Демек, тері беткейі арқылы қан жылуды аз жоғалтады.Егер дене
температурасы
төмендесе,
тері асты
тамырлары
тарылады
64.
2. Пилоэрекция – бұл тері бетіндегіқылшықтардың тітіркенуі, яғни «құс терісі»
деп аталады, нәтижесінде денеде жылу
сақталынады.
65.
66.
Ағзадағы глюкозадеңгейі екі ұйқы
безінің гормондары
инсулин,
глюкагонмен
реттеледі.
Егер
қанда
глюкоза көп болса,
инсулин
оны
гликогенге
айналдырады
Гликоген
Қандағы глюкоза
67.
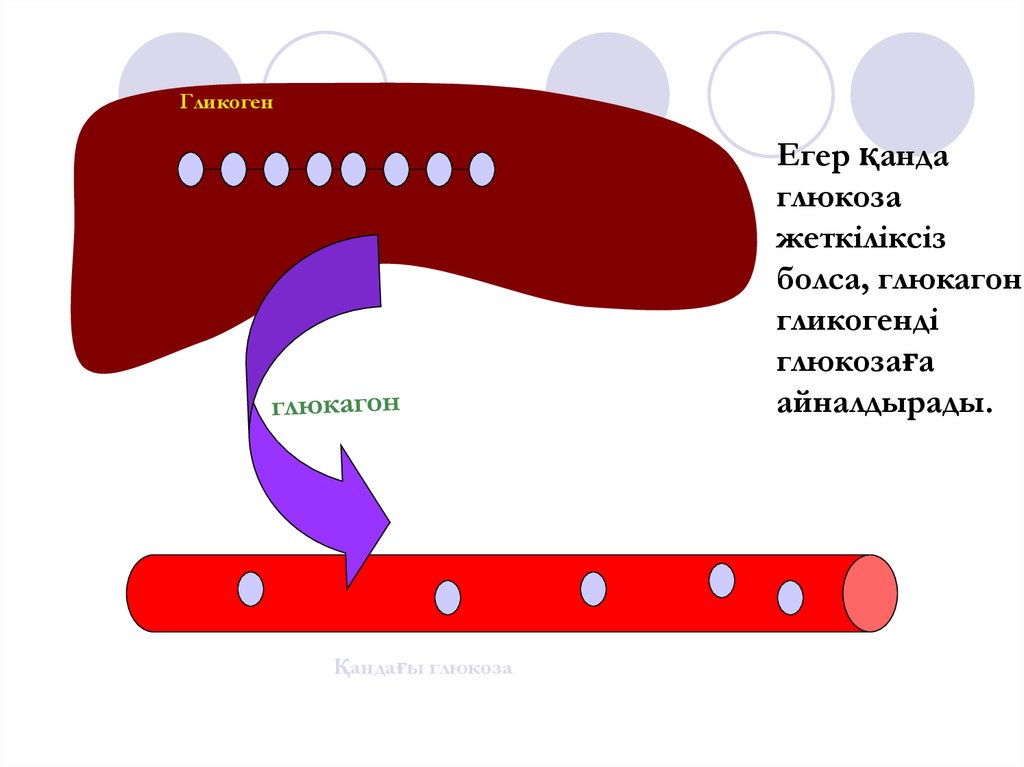
ГликогенЕгер қанда
глюкоза
жеткіліксіз
болса, глюкагон
гликогенді
глюкозаға
айналдырады.
Қандағы глюкоза
68.
Глюкозаныгликогенге
айналдыру үшін
инсулин жоқ.
Гликоген
Глюкозаның
концентрациясы
өте көп.
Қандағы глюкоза
69.
Гомеостаздың бұзылысы - дисгомеостаздеп аталады. Нәтижесінде аурулар жетіледі:
- диабет, подагра кан ағымында әртүрлі
токсиндердің болу нәтижесінде пайда
болады. Гомеостазды бақылайтын
механизмдер ағзаның биологиялық
сұйықтарында токсиндердің жинақталуын
болдырмау керек, бірақ кейбір даралардың
гомеостаз механизмдері жеткілікті
эффективті жұмыс істемейді.
70.
Қант диабеті – жалпы әлемдік мәселе.Әртүрлі елдерде ауыратындардың саны
тұрғындардың 1-2 %- құрайды
Гипергликемия көптеген тамырлардың
бұзылысын, миокард инфаркті, аяқтардың
жаралық зақымданулары, бүйректік
жетіспеушіліктер, соқырлық асқынуларын
шақырады.
Адам қант диабетінің екі типімен ауырады: 1
типтегі диабет (инсулинге тәуелді түрі) және
диабеттің 2 типі (инсулинге тәуелсіз түрі).
71.
Диабеттің 1-ші типінде ұйқы безіндегі βжасушалар аутоиммундік процесстіңсалдарынан бұзылады және инсулин
түзілмейді.
Диабеттің II типінде перифериялық
ұлпалардың жасушалары инсулинге
резистенттілік көрсетеді, нәтижесінде
гипергликемия (қанттың жоғарылауы)
дамиды.