
























 biology
biologySimilar presentations:

")








Реакции глюконеогенеза
1. Реакции глюконеогенеза
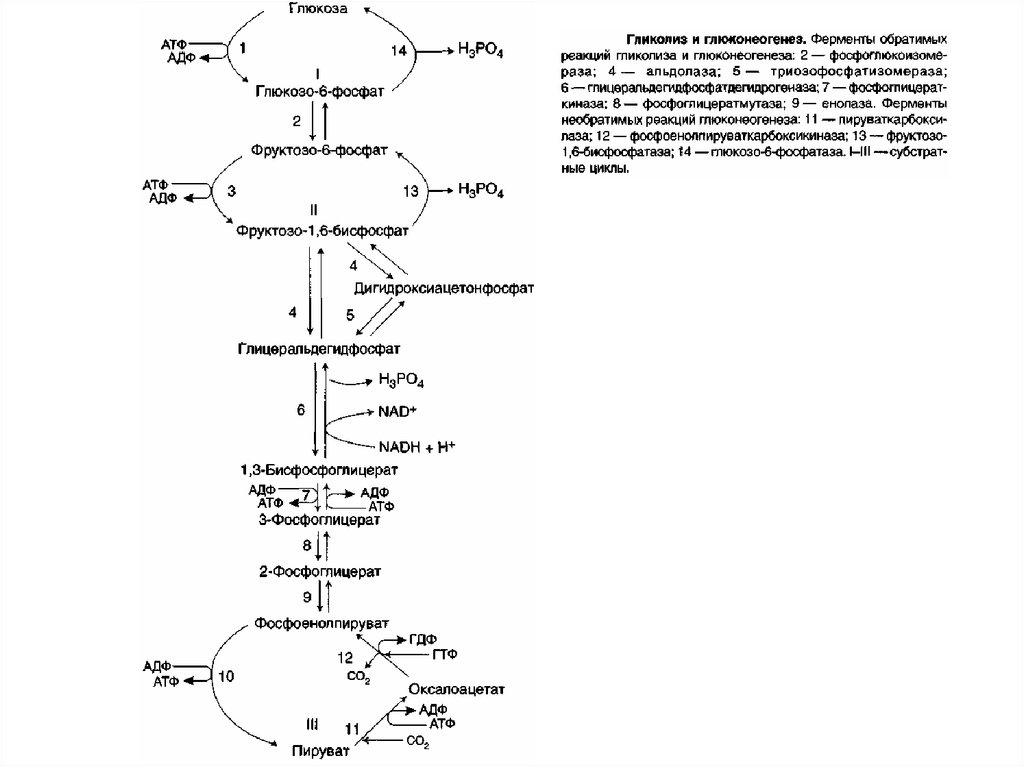
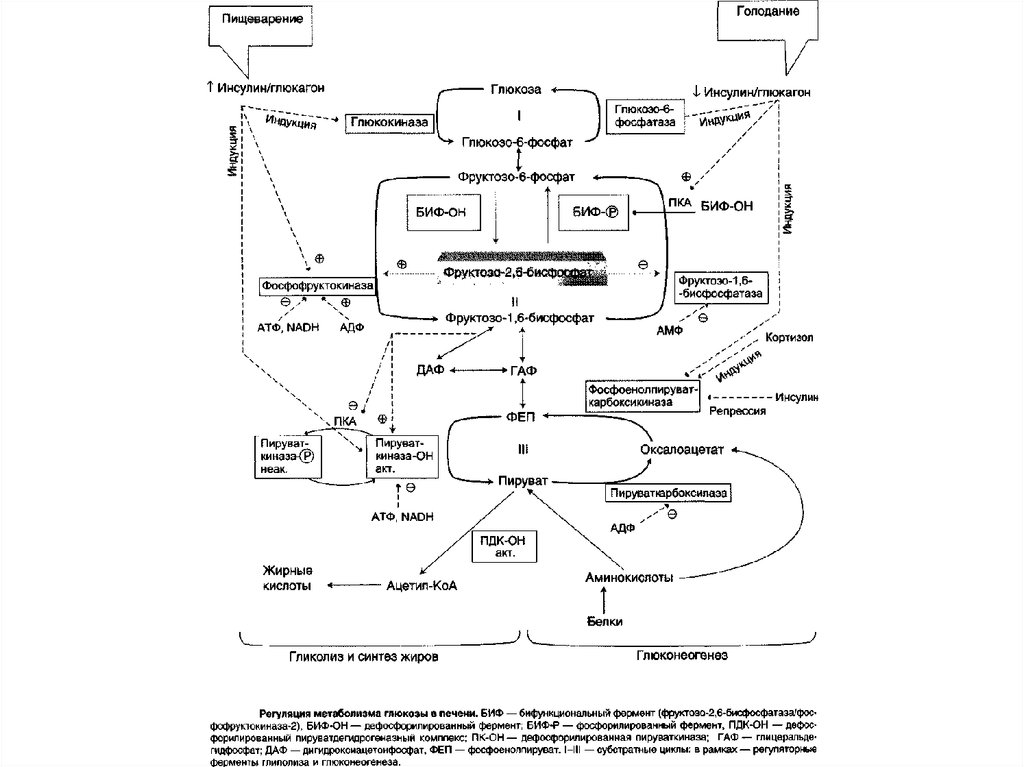
Большинство реакций глюконеогенеза протекает засчёт обратимых реакций гликолиза и катализируется
теми же ферментами. Однако 3 реакции гликолиза
термодинамически необратимы. На этих стадиях
реакции глюконеогенеза протекают другими путями.
Необходимо отметить, что гликолиз протекает в
цитозоле, а часть реакций глюконеогенеза происходит
в митохондриях.
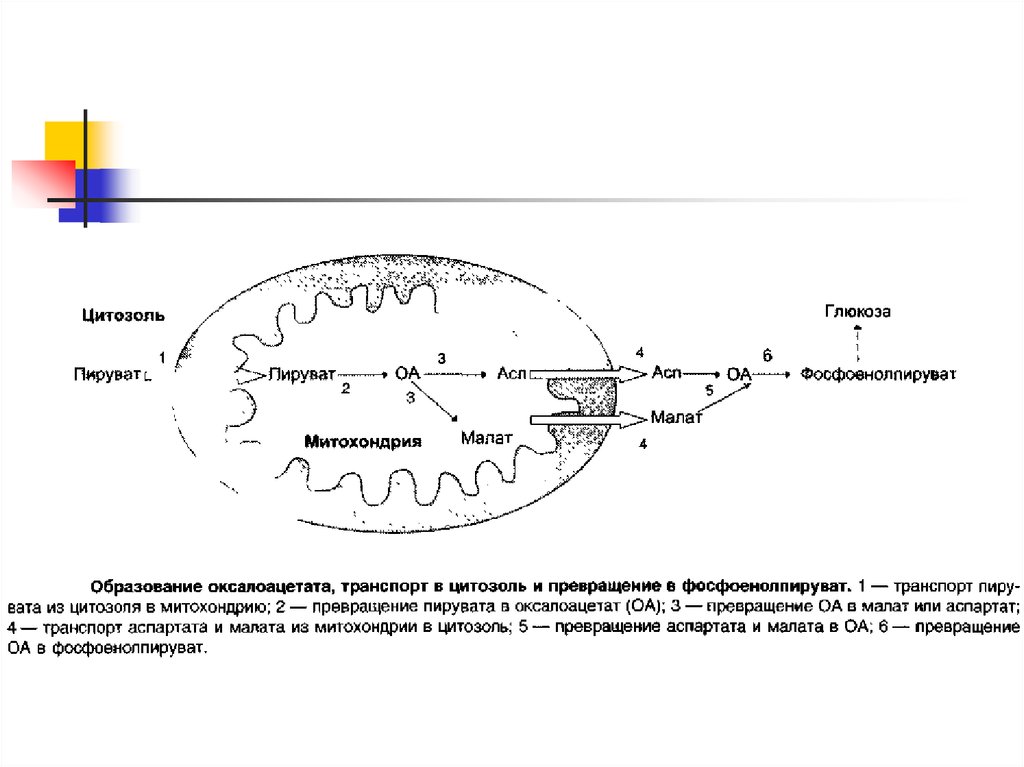
2. Образование фосфоенолпирувата из пирувата — первая из необратимых стадий глюконеогенеза
Образованиефосфоенолпирувата
из
пирувата
происходит в ходе двух реакций, первая из которых
протекает в митохондриях. Пируват, образующийся из
лактата
или
из
некоторых
аминокислот,
транспортируется в матрикс митохондрий и там
карбоксилируется с образованием оксалоацетата.
Пируваткарбоксилаза,
катализирующая
данную
реакцию, — митохондриальный фермент, коферментом
которого является биотин. Реакция протекает с
использованием АТФ.
3. Включение субстратов в глюконеогенез
4.
5. Образование оксалоацетата из пирувата
6. Превращение оксалоацетата в малат
7. Превращение оксалоацетата в фосфоенолпируват
8.
9. Гидролиз фруктоза-1,6-бисфосфата и глюкоза-6-фосфата
Отщепление фосфатной группы из фруктозо-1,6бисфосфатаи
глюкозо-6-фосфата
—
также
необратимые реакции глюконеогенеза. В ходе
гликолиза эти реакции катализируют специфические
киназы с использованием энергии АТФ. В
глюконеогенезе они протекают без участия АТФ и
АДФ и ускоряются не киназами, а фосфатазами —
ферментами, принадлежащими к классу гидролаз.
Ферменты фруктозо-1,6-бисфосфатаза и глюкозо-6фосфатаза катализируют отщепление фосфатной
группы от фруктозо-1,6-бисфосфата и глюкозо-6фосфата. После чего свободная глюкоза выходит из
клетки в кровь.
10. Энергетический баланс глюконеогенеза из пирувата
Суммарный результат глюконеогенеза из пируватавыражается следующим уравнением:
2 Пируват + 4 АТФ + 2 ГТФ + 2 (NADH+Н+) + 4 Н2О →
Глюкоза + 4 АДФ + 2 ГДФ +6 Н3РО4 + 2 NAD+.
11. Синтез глюкозы из лактата
Лактат, образованный в анаэробном гликолизе, неявляется
конечным
продуктом
метаболизма.
Использование лактата связано с его превращением в
печени в пируват. Лактат как источник пирувата важен
не столько при голодании, сколько при нормальной
жизнедеятельности организма. Его превращение в
пируват и дальнейшее использование последнего
являются способом утилизации лактата.
12.
Лактат, образовавшийся в интенсивно работающихмышцах или в клетках с преобладающим анаэробным
способом катаболизма глюкозы, поступает в кровь, а
затем в печень. В печени отношение NADH/NAD+
ниже, чем в сокращающейся мышце, поэтому лактатдегидрогеназная реакция протекает в обратном
направлении, т.е. в сторону образования пирувата из
лактата. Далее пируват включается в глюконеогенез, а
образовавшаяся глюкоза поступает в кровь и
поглощается скелетными мышцами.
13.
Цикл Кори выполняет 2 важнейшие функции:1 — обеспечивает утилизацию лактата;
2
— предотвращает накопление лактата и, как
следствие этого, опасное снижение рН (лактоацидоз).
Часть пирувата, образованного из лактата, окисляется
печенью до СО2 и Н2О. Энергия окисления может
использоваться для синтеза АТФ, необходимого для
реакций глюконеогенеза.
14. Синтез глюкозы из аминокислот
В условиях голодания часть белков мышечной тканираспадается
до
аминокислот,
которые
далее
включаются в процесс катаболизма. Аминокислоты,
которые при катаболизме превращаются в пируват или
метаболиты цитратного цикла, могут рассматриваться
как потенциальные предшественники глюкозы и
гликогена и носят название гликогенных. Например,
оксалоацетат, образующийся из аспарагиновой
кислоты, является промежуточным продуктом как
цитратного цикла, так и глюконеогенеза.
15.
Из всех аминокислот, поступающих в печень,примерно 30% приходится на долю аланина. Это
существует следующая последовательность событий
(глюкозо-аланиновый цикл): глюкоза в мышцах →
пируват в мышцах → аланин в мышцах → аланин в
печени → глюкоза в печени → глюкоза в мышцах. Весь
цикл не приводит к увеличению количества глюкозы в
мышцах, но он решает проблемы транспорта аминного
азота из мышц в печень и предотвращает лактоацидоз.
16. Синтез глюкозы из глицерола
Глицерол образуется при гидролизе триацил-глицеролов,главным образом в жировой ткани. Использовать его могут
только те ткани, в которых имеется фермент глицерол
киназа, например печень, почки. Этот АТФ-зависимый
фермент катализирует превращение глицерола в αглицерофосфат (глицерол-3-фосфат). При включении
глицерол-3-фосфата в глюконеогенез происходит его
дегидрирование
NAD-зависимой
дегидрогеназой
с
образованием дигидроксиацетонфосфата, который далее
превращается в глюкозу.
17. Глюкозо-аланиновый цикл.
18. Превращение глицерола в дигидроксиацетон-фосфат.
19.
20. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ПРЕВРАЩЕНИЯ ГЛЮКОЗЫ
Пентозофосфатныйпуть,
называемый
также
гексомонофосфатным
шунтом,
служит
альтернативным путём окисления глюкозо-6-фосфата.
Пентозофосфатный путь состоит из 2 фаз —
окислительной и неокислительной.
В окислительной фазе глюкозо-6-фосфат необратимо
окисляется в пентозу — рибулозо-5-фосфат, и
образуется восстановленный NADPH.
В неокислительной фазе рибулозо-5-фосфат обратимо
превращается в рибозо-5-фосфат и метаболиты
гликолиза.
21.
Пентозофосфатный путь обеспечивает клетки рибозойдля синтеза пуриновых и пиримидиновых нуклеотидов
и гидрированным коферментом NADPH, который
используется в восстановительных процессах.
Суммарное уравнение пентозофосфатного пути
выражается следующим образом:
3 Глюкозо-6-фосфат + 6 NADP+ → 3 СО2 +6 (NADPH +
Н+) + 2 Фруктозо-6-фосфат + Глицеральдегид-3фосфат.
Ферменты пентозофосфатного пути, так же, как и
ферменты гликолиза, локализованы в цитозоле.
22. Окислительный этап
В окислительной части пентозофосфатного путиглюкозо-6-фосфат
подвергается
окислительному
декарбоксилированию,
в
результате
которого
образуются пентозы. Этот этап включает 2 реакции
дегидрирования.
23.
• Суммарноеуравнение
окислительного
этапа
пентозофосфатного пути можно представить в виде:
• Реакции окислительного этапа служат основным
источником
NADPH
в
клетках.
Гидрированные
коферменты снабжают водородом биосинтетические
процессы, окислительно-восстановительные реакции,
включающие защиту клеток от активных форм кислорода.
NADPH как донор водорода участвует в анаболических
процессах, например в синтезе холестерина. Это источник
восстановительных эквивалентов для цитохрома Р450,
катализирующего образование гидроксильных групп при
синтезе стероидных гормонов, жёлчных кислот, при
катаболизме лекарственных веществ и других чужеродных
соединений
24. Окислительный этап пентозофосфатного пути
25. Неокислительный этап
Неокислительный этап пентозофосфатного путивключает серию обратимых реакций, в результате
которых рибулозо-5-фосфат превращается в рибозо-5фосфат и ксилулозо-5-фосфат, и далее за счёт переноса
углеродных фрагментов в метаболиты гликолиза —
фруктозо-6-фосфат и глицеральдегид-3-фосфат. В этих
превращениях
принимают
участие
ферменты:
эпимераза, изомераза, транскетолаза и трансальдолаза.
26.
Суммарный результат метаболизма 3 молекулрибулозо-5-фосфата
в
неокислительной
фазе
пентозофосфатного пути — образование 2 молекул
фруктозо-6-фосфата и 1 молекулы глицеральдегид-3фосфата. Далее фруктозо-6-фосфат и глицеральдегид3-фосфат могут превратиться в глюкозу. С учётом
стехиометрического коэффициента, равного 2, для
образования 5 молекул глюкозы (содержащих 30
атомов углерода) потребуются 4 молекулы фруктозо-6фосфата и 2 молекулы глицеральдегид-3-фосфата (в
сумме содержащие также 30 атомов углерода) или,
соответственно, 6 молекул рибулозо-5-фосфата. Таким
образом, неокислительный путь можно представить
как процесс возвращения пентоз в фонд гексоз.
27. Пентозофосфатный цикл
Окислительныйэтап
образования
пентоз
и
неокислительный этап (путь возвращения пентоз в
гексозы) составляют вместе циклический процесс.
Такой процесс можно описать общим уравнением:
6 Глюкозо-6-фосфат + 12 NADP+ + 2 Н2О → Глюкозо-6фосфат + 12 NADPH +12 Н+ + СО2.
Это означает, что из 6 молекул глюкозы образуются 6
молекул рибулозо-5-фосфат (пентозы) и 6 молекул
СО2.