










































 biology
biology chemistry
chemistrySimilar presentations:


")
")






Окисление и синтез глюкозы: гликолиз, глюконеогенез, пентозофосфатный путь
1.
ЛекцияОкисление и синтез глюкозы:
гликолиз, глюконеогенез,
пентозофосфатный путь
• Дисциплина: Б1.Б.8 –Биологическая химия, биохимия полости рта
• Специальность: 31.05.03 –Стоматология
НГМУ, КАФЕДРА МЕДИЦИНСКОЙ ХИМИИ
К.М.Н., ДОЦЕНТ ЛЕБЕРФАРБ ЕЛЕНА ЮРЬЕВНА
2.
План лекции• 1. Катаболизм глюкозы
• 2. Глюконеогенез
• 3. Регуляция гликолиза и глюконеогенез
• 4. Пентозофосфатный путь превращения
глюкозы
• 5. Биохимические основы сахарного
диабета
3.
ГЛИКОЛИЗОкисление глюкозы происходит по двум
направлениям:
1. Окисление с получением энергии.
Этот путь, т.е. тот по которому глюкоза
окисляется для получения энергии,
называется гликолиз. Конечным продуктом
гликолиза является пировиноградная
кислота (пируват).
В зависимости от дальнейшей судьбы пирувата
различают аэробное и анаэробное окисление
глюкозы. Целью обоих типов окисления
является получение АТФ.
2. Окисление с образованием пентоз: рибозы,
рибулозы, ксилулозы. Этот путь
называется пентозофосфатный шунт и не
связан с получением энергии
4.
В аэробном и анаэробном гликолизе можно выделить два этапа.1. Превращение глюкозы в две молекулы глицеральдегид-3-фосфата. Эта серия
реакции протекает с потреблением АТФ.
5.
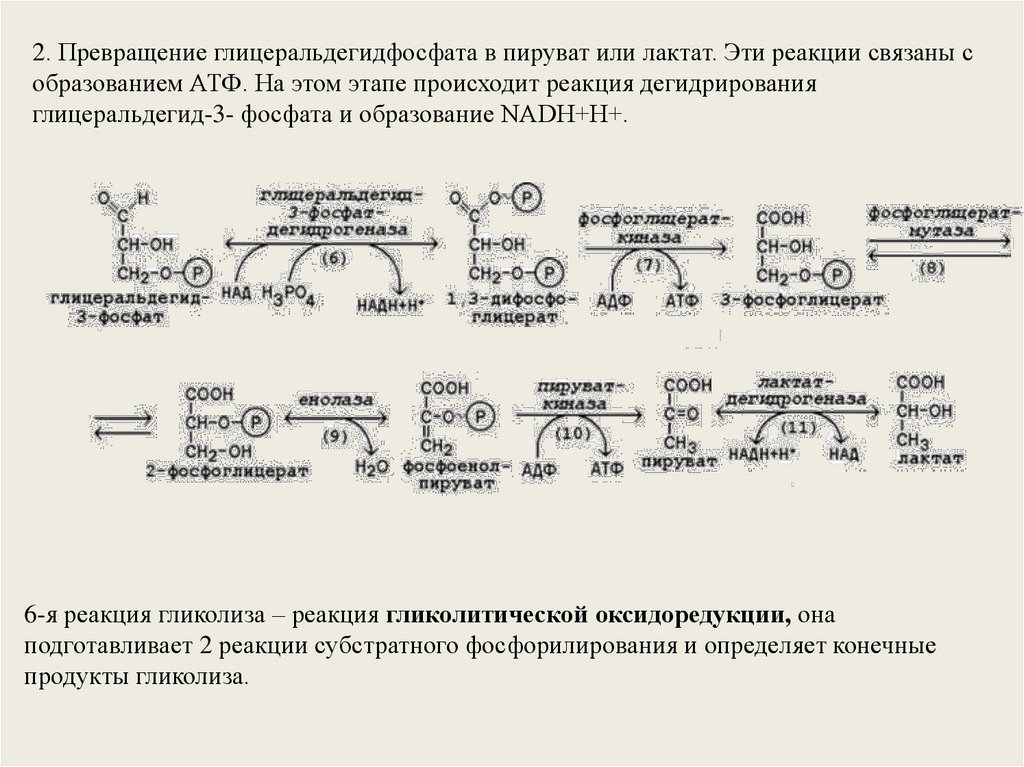
2. Превращение глицеральдегидфосфата в пируват или лактат. Эти реакции связаны собразованием АТФ. На этом этапе происходит реакция дегидрирования
глицеральдегид-3- фосфата и образование NADH+H+.
6-я реакция гликолиза – реакция гликолитической оксидоредукции, она
подготавливает 2 реакции субстратного фосфорилирования и определяет конечные
продукты гликолиза.
6.
Аэробным гликолизом называют процесс окисления глюкозы до пировинограднойкислоты, протекающий в присутствии кислорода. Этот процесс составляет
специфический для глюкозы путь катаболизма.
Суммарное уравнение
C6H12O6 + 6 O2 + 38 АДФ + 38 Фнеорг → 6 CO2 + 44 H2О + 38 АТФ
Аэробный распад глюкозы включает несколько стадий:
• аэробный гликолиз — процесс окисления глюкозы с образованием двух молекул
пирувата;
• общий путь катаболизма, включающий превращение пирувата в ацетил-СоА и его
дальнейшее окисление в цитратном цикле;
• цепь переноса электронов на кислород, сопряженная с реакциями дегидрирования,
происходящими в процессе распада глюкозы
Таким образом, аэробный распад глюкозы до CO2 и H2O включает реакции
аэробного гликолиза и последующее окисление пирувата в общих путях катаболизма.
7.
Характеристика аэробного гликолиза:Большинство реакций обратимо, за исключением трех (реакции 1,3,10);
Все метаболиты находятся в фосфорилированной форме
Источниками фосфатной группы в реакциях фосфорилирования являются АТФ
(реакции 1 и 3) или неорганический фосфат (реакция 6);
Регенерация NAD+ (реакции 6,11), являющаяся необходимым условием
протекания гликолиза, происходит при аэробном гликолизе посредством
дыхательной цепи. В этом случае водород транспортируется в митохондрии с
помощью челночного механизма при участии переносчиков. Это происходит
потому, что мембрана митохондрий непроницаема для протонов;
Образование АТФ при гликолизе может идти двумя путями: либо субстратным
фосфорилированием, когда для фосфорилирования АДФ используется энергия
макроэргической связи субстрата (реакции 7,10), либо путем окислительного
фосфорилирования АДФ, сопряженного с дыхательной цепью (реакции 6, 11).
8.
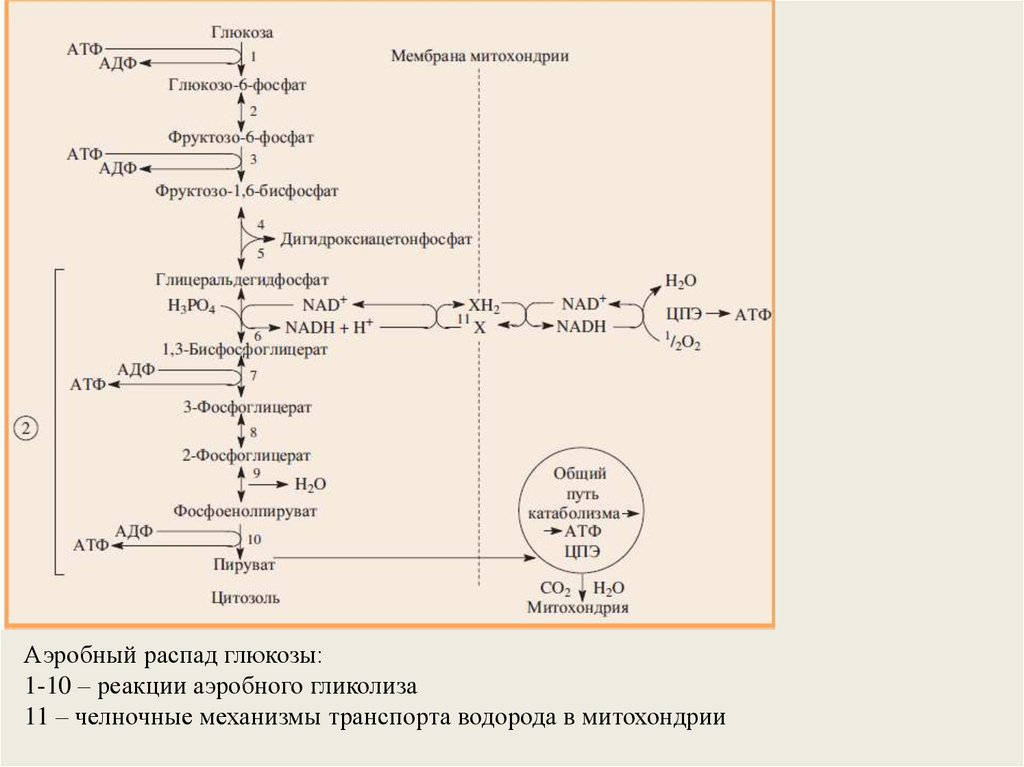
Аэробный распад глюкозы:1-10 – реакции аэробного гликолиза
11 – челночные механизмы транспорта водорода в митохондрии
9.
Анаэробный гликолиз включает те же реакции специфического пути распада глюкозы до пирувата, но с последующим превращением пирувата влактат, т.е. термины «анаэробный распад» и «анаэробный гликолиз»
совпадают.
Анаэробный гликолиз протекает без использования кислорода, так как в
определенных ситуациях обеспечение кислородом тканей может не
соответствовать их потребностям.
Суммарное уравнение анаэробного гликолиза:
C6H12O6 + 2 АДФ + 2 Фнеорг → 2 Лактат + 2 H2O + 2 АТФ
Все ферменты, катализирующие реакции этого процесса, локализованы в
цитозоле клетки.
10.
Последняя реакция бескислородного окисления глюкозы, одиннадцатая –образование молочной кислоты из пирувата под
действием лактатдегидрогеназы. Важно то, что эта реакция
осуществляется только в анаэробных условиях. Эта реакция необходима
клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие
кислорода не может окисляться в митохондриях.
11.
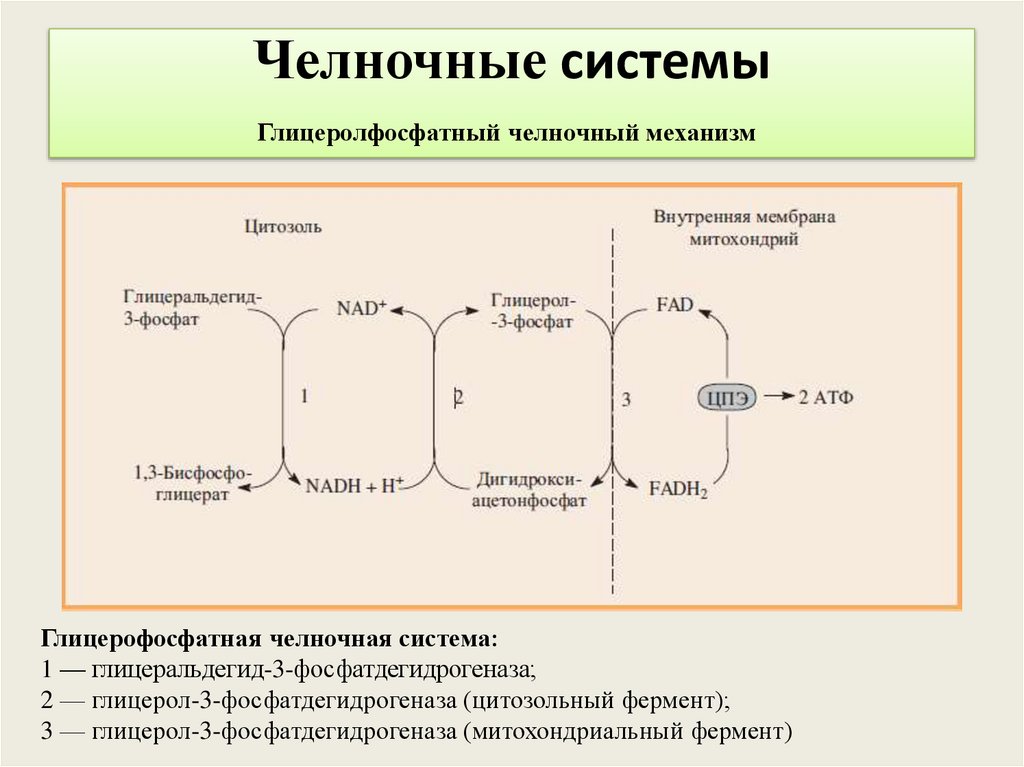
Челночные системыГлицеролфосфатный челночный механизм
Глицерофосфатная челночная система:
1 — глицеральдегид-3-фосфатдегидрогеназа;
2 — глицерол-3-фосфатдегидрогеназа (цитозольный фермент);
3 — глицерол-3-фосфатдегидрогеназа (митохондриальный фермент)
12.
Глицерофосфатный челночный циклНАДН, образующаяся в 6-й реакции не может транспортировать
Н+ из цитозоля.
ДАФ восстанавливается до глицерол-3-ф,используя
глицероальдегид-3-фосфат- дегидрогеназу с образованием
глицерол-3ф, который легко проникает через мембрану. В
митохондриях этот фермент ФАД-зависимый, окисляет
глицерол-3-фосфат с образованием ДАФ и ФАДН2,который
окисляясь в дыхательной цепи дает 4 АТФ
13.
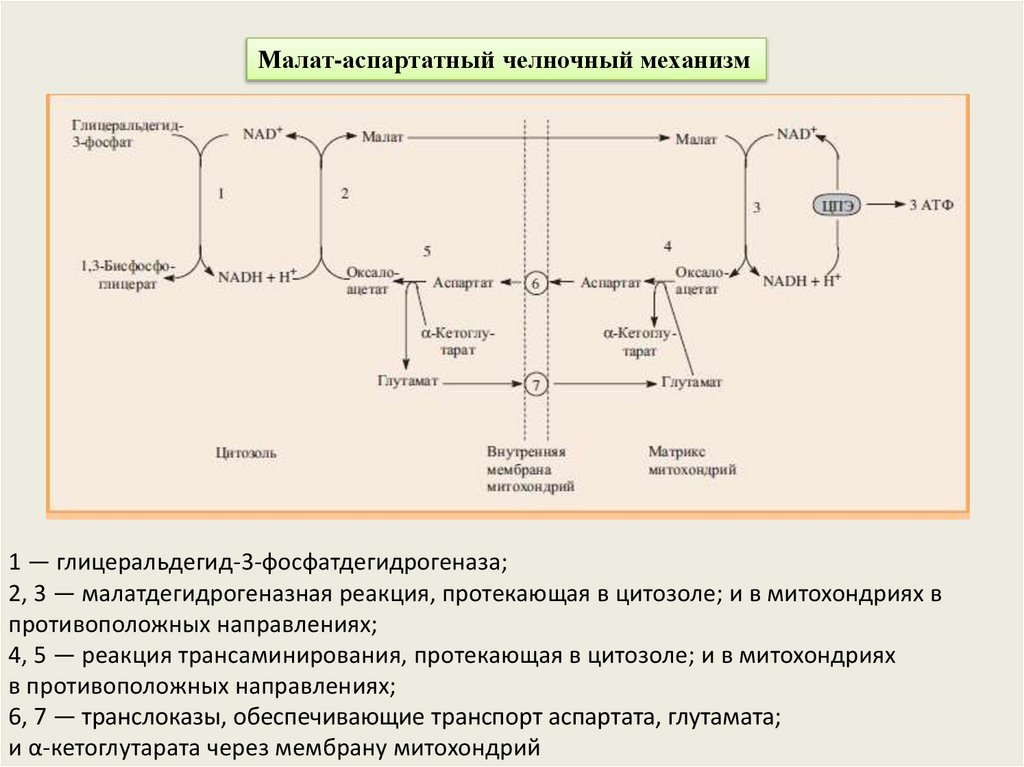
Малат-аспартатный челночный механизм1 — глицеральдегид-3-фосфатдегидрогеназа;
2, 3 — малатдегидрогеназная реакция, протекающая в цитозоле; и в митохондриях в
противоположных направлениях;
4, 5 — реакция трансаминирования, протекающая в цитозоле; и в митохондриях
в противоположных направлениях;
6, 7 — транслоказы, обеспечивающие транспорт аспартата, глутамата;
и α-кетоглутарата через мембрану митохондрий
14.
Малат-аспартатный челночный циклКлючевыми ферментами этого челнока являются изоферменты
малатдегидрогеназы – цитоплазматический и митохондриальный.
Оксалоацетат, который под действием цитозольного пула
малатдегидрогеназы и за счет "гликолитического" НАДН восстанавливается
до яблочной кислоты (малата).
Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь
метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как
мембрана митохондрий непроницаема для оксалоацетата, то он при помощи
аспартатаминотрансферазы трансаминируется до аспарагиновой кислоты,
которая в обмен на глутамат выходит в цитозоль.
Таким образом, атомы водорода от цитозольного НАДН перемещаются в
состав митохондриального НАДН, который окисляясь в дыхательной цепи дает
6 АТФ
15.
Выход АТР при аэробном распаде глюкозы до конечных продуктов16.
Кроме энергетической функции процесс катаболизма глюкозы можетвыполнять и анаболические функции.
Метаболиты гликолиза используются для синтеза новых соединений.
Так, фруктозо-6-фосфат и глицеральдегид-3-фосфат участвуют в образовании
рибозо-5-фосфата — структурного компонента нуклеотидов;
3-фосфоглицерат может включаться в синтез аминокислот, таких, как серин,
глицин, цистеин.
В печени и жировой ткани ацетил-КоА, образующийся из пирувата,
используется как субстрат при биосинтезе жирных кислот, холестерина, а
дигидроксиацетонфосфат как субстрат для синтеза глицерол-3-фосфата.
17.
Регуляция гликолизаГормональная регуляция
В печени гликолиз стимулируется инсулином, повышающим количество
ключевых ферментов гликолиза
(гексокиназы, фосфофруктокиназы, пируваткиназы).
В печени активность глюкокиназы, кроме инсулина, регулируется другими
гормонами:
активацию вызывает андрогены,
подавляют ее активность глюкокортикоиды и эстрогены.
В других тканях активность гексокиназ повышается
тиреоидными гормонами, снижается – глюкокортикоидами и соматотропином.
18.
Регуляция гликолизаМетаболическая регуляция
Гексокиназа непеченочных клеток ингибируется продуктом собственной
реакции – глюкозо-6-фосфатом.
Фосфофруктокиназа активируется АМФ и собственным субстратом
(фруктозо-6-фосфат),
ингибируется – АТФ, лимонной кислотой, жирными кислотами.
Пируваткиназа активируется фруктозо-1,6-дифосфатом (прямая
положительная регуляция).
19.
Регуляция катаболизма глюкозы вскелетных мышцах
20.
Синтез глюкозы (глюконеогенез)Глюконеогенез — это процесс синтеза глюкозы из веществ неуглеводной
природы. Его основной функцией является поддержание уровня глюкозы в крови в
период длительного голодания и интенсивных физических нагрузок.
Процесс протекает главным образом в печени и менее интенсивно - в
корковом веществе почек, а также в слизистой оболочке кишечника.
Первичными субстратами глюконеогенеза являются лактат, аминокислоты и
глицерол. Включение этих субстратов в глюконеогенез зависит от физиологического состояния организма.
• Лактат является продуктом анаэробного гликолиза. Он образуется при любых
состояниях организма (эритроциты, работающие мышцы). Таким образом, лактат
используется в глюконеогенезе постоянно.
• Глицерол высвобождается при гидролизе жиров в жировой ткани в период
голодания или при длительной физической нагрузке.
• Аминокислоты образуются в результате распада мышечных белков и
включаются в глюконеогенез при длительном голодании или продолжительной
мышечной работе.
21.
Реакции глюконеогенезаБольшинство реакций
глюконеогенеза
протекают
за счет обратимых
реакций гликолиза и
катализируются теми
же ферментами.
Однако три реакции
гликолиза термодинамически
необратимы. На этих
стадиях реакции
глюконеогенеза
протекают
другими путями.
22.
Образование фосфоенолпирувата из пирувата происходит в ходе двухреакций, первая из которых протекает в митохондриях. Пируват, образующийся из
лактата или из некоторых аминокислот, транпортируется в матрикс митохондрий и там
карбоксилируется с образованием оксалоацетата. Пируваткарбоксилаза,
катализирующая данную реакцию, — митохондриальный фермент, коферментом
которого является биотин. Реакция протекает с использованием АТФ. Оксалоацетат
транспортируется в цитозоль и включается в глюконеогенез, превращаясь в
фосфоенолпируват в ходе реакции, катализируемой
фосфоеполпируваткарбоксикиназой — ГТФ-зависимым ферментом. Все остальные
реакции глюконеогенеза протекают в цитозоле. В ходе этого процесса на синтез 1 моль
глюкозы из 2 моль пирувата расходуется 4 моль АТФ и 2 моль ГТФ.
23.
Отщепление фосфатной группы из фруктозо-1,6-бисфосфата и глюкозо-6-фосфатакатализируют ферменты фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза.
Свободная глюкоза выходит из клетки в кровь.
24.
Итак, в печени существует четыре фермента, которые принимают участиетолько в глюконеогенезе и катализируют обходные реакции необратимых
стадий гликолиза.
Это пируваткарбоксилаза, фосфоенолпируваткарбоксикиназа,
фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза.
Энергетический баланс глюконеогенеза из пирувата.
В ходе этого процесса расходуется 6 моль АТФ на синтез 1 моль глюкозы из
2 моль пирувата.
4 АТФ расходуется на стадии синтеза фосфоенолпирувата из оксалоацетата
и еще 2 моль АТФ на стадиях образования 1,3-бисфосфоглицерата из
3-фосфоглицерата.
25.
Синтез глюкозы из лактатаЛактат - конечный продукт анаэробного гликолиза транспортируется в другие
ткани, например в печень, сердечную мышцу, где превращается в пируват вступает
на путь глюконеогенеза
Значение цикла Кори:
Обеспечивает утилизацию
лактата
Предотвращает накопление
лактата и лактатный ацидоз
На глюконеогенез
использется энергия окисления
лактата
Снижение использования лактата в качестве субстрата в синтезе глюкозы, вызванное
дефектом ферментов глюконеогенеза, может приводить к повышению концентрации
молочной кислоты в крови, понижению рН и, следовательно, к лактоацидозу.
26.
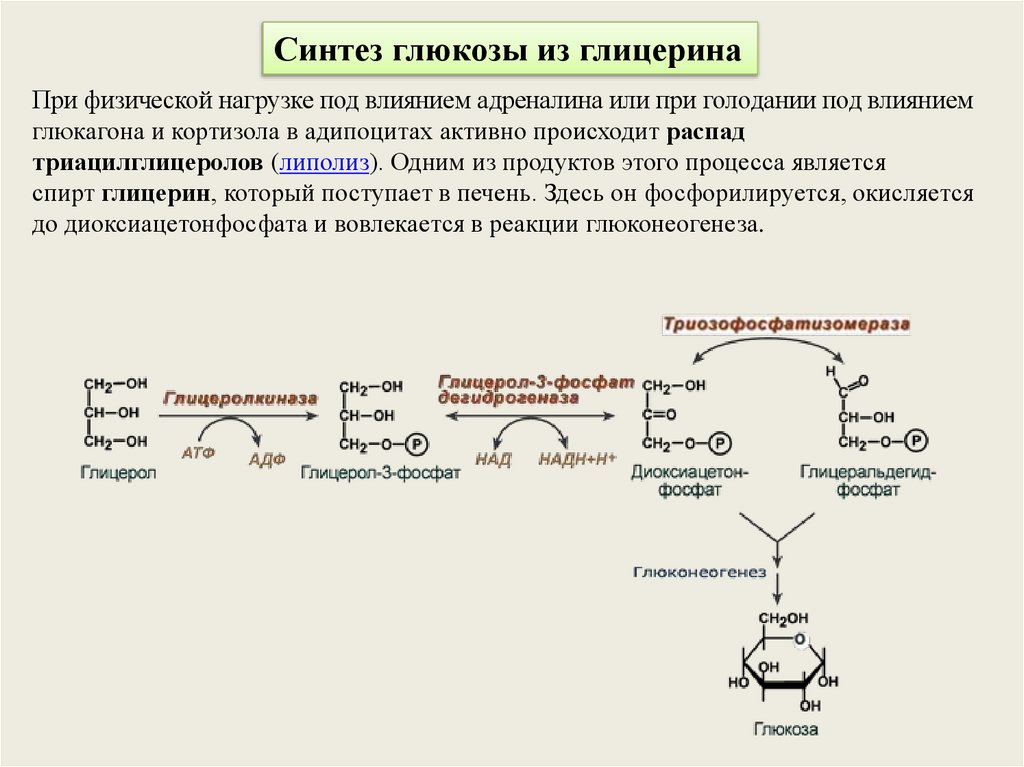
Синтез глюкозы из глицеринаПри физической нагрузке под влиянием адреналина или при голодании под влиянием
глюкагона и кортизола в адипоцитах активно происходит распад
триацилглицеролов (липолиз). Одним из продуктов этого процесса является
спирт глицерин, который поступает в печень. Здесь он фосфорилируется, окисляется
до диоксиацетонфосфата и вовлекается в реакции глюконеогенеза.
27.
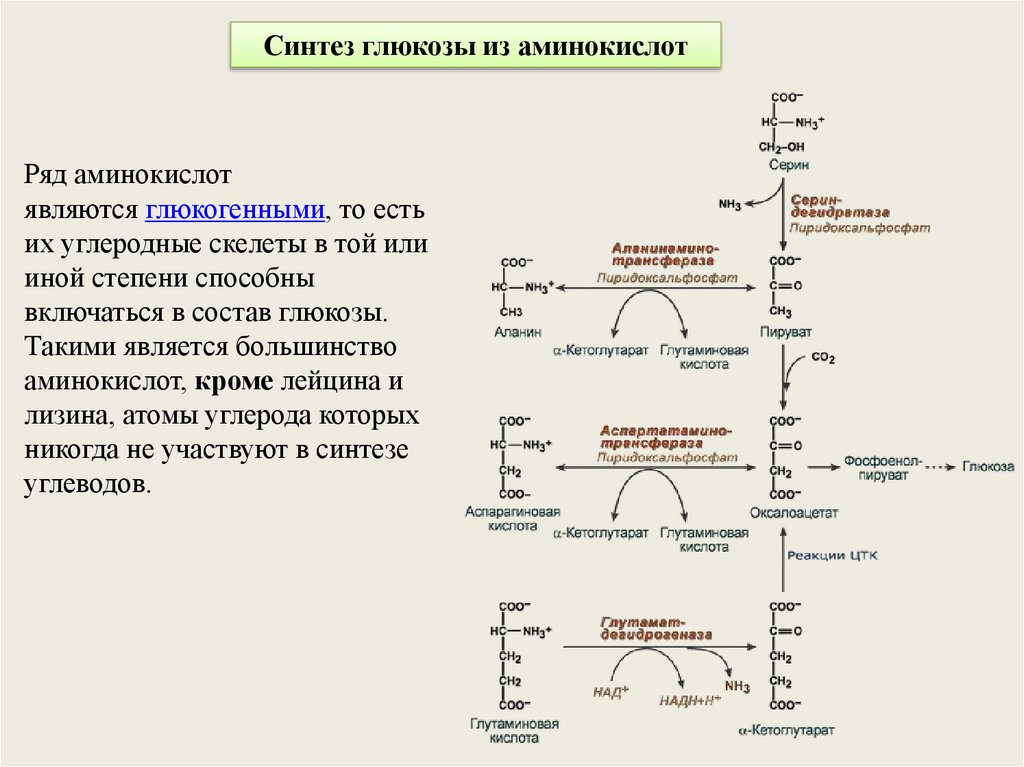
Синтез глюкозы из аминокислотРяд аминокислот
являются глюкогенными, то есть
их углеродные скелеты в той или
иной степени способны
включаться в состав глюкозы.
Такими является большинство
аминокислот, кроме лейцина и
лизина, атомы углерода которых
никогда не участвуют в синтезе
углеводов.
28.
Регуляция глюконеогенезаКлючевыми ферментами глюконеогенеза являются
пируваткарбоксилаза и
фосфоэнолпируваткарбоксиназа.
Гормональная активация
осуществляется глюкокортикоидами, которые увеличивают синтез
пируваткарбоксилазы, фосфоенолпируват-карбоксикиназы, фруктозо-1,6дифосфатазы.
Глюкагон стимулирует те же самые ферменты через аденилатциклазный механизм
путем фосфорилирования.
Энергия для глюконеогенеза поступает от β-окисления жирных кислот. Конечный
продукт этого окисления, ацетил-SКоА аллостерически стимулирует активность
первого фермента глюконеогенеза – пируваткарбоксилазы. Кроме этого, фруктозо-1,6дифосфатаза стимулируется при участии АТФ.
29.
Пентозофосфатный путь - альтернативный путь окисления глюкозо6-фосфатаНаиболее активно реакции пентозофосфатного пути идут в цитозоле клеток
печени, жировой ткани, эритроцитах, коре надпочечников, молочной железе
при лактации. Этот путь окисления глюкозы не связан с образованием
энергии, а обеспечивает
анаболизм клеток.
Метаболизм глюкозо-6-фосфата в пентозофосфатном пути может
идти по различным вариантам. Судьба метаболитов
пентозофосфатного пути зависит от ситуации, потребностей клетки и
ее типа.
30.
2 этапа:Первый этап. Окислительный.
Глюкозо-6-фосфат в трех реакциях превращается в рибулозо-5-фосфат,
реакции сопровождаются восстановлением двух молекул НАДФ до НАДФН.
На этом этапе происходит регуляция процесса:
инсулин повышает активность глюкозо-6-фосфат-дегидрогеназы и
фосфоглюконат-дегидрогеназы.
Второй этап. Структурных перестроек (неокислительный).
В этих реакциях молекулы рибулозо-5-фосфата изомеризуются до рибозо5-фосфата и ксилулозо-5-фосфата. Далее под влиянием ферментов
транскетолазы и трансальдолазы происходят структурные перестройки с
образованием других моносахаридов.
31.
Все ферментыпентозофосфатного
пути локализованы в
цитозоле.
Промежуточные продукты
пентозофосфатного пути
превращения глюкозы (фруктозо6-фосфат, глицеральдегид-3фосфат) могут включаться в пути
аэробного и анаэробного
окисления и служить источником
энергии для синтеза АТР.
32.
Биологическая роль ПФПЕдинственный способ получения пентоз (для синтеза нуклеотидов)
Молекулы НАДФН также используются:
• для синтеза жирных кислот (печень, жировая ткань),
• для синтеза холестерола и других стероидов (например, желчных
кислот),
• в реакциия восстановительного аминирования для
образования глутаминовой кислоты из α-кетоглутаровой
кислоты,
• для восстановления окисленного тиоредоксина и окисленного
глутатиона в системе антиоксидантной защиты клетки от
свободно-радикального окисления,
• для реактивации фолиевой кислоты и витамина К,
• источник восстановительных эквивалентов для цитохрома Р–450,
катализирующего образование гидроксильных групп
33.
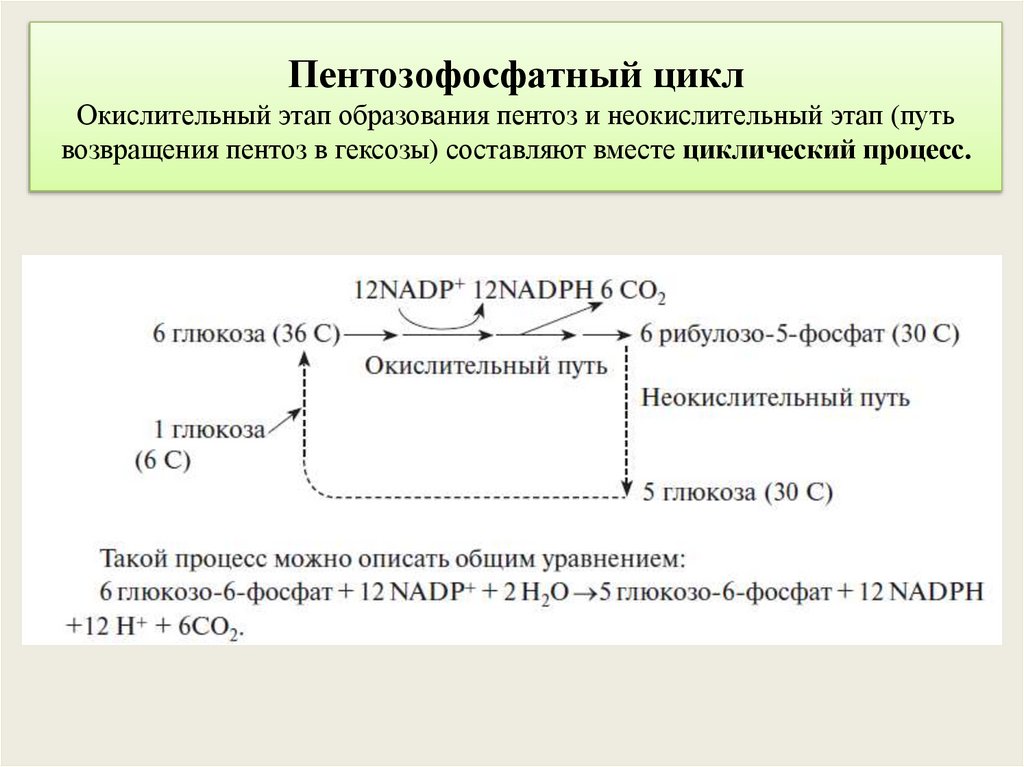
Пентозофосфатный циклОкислительный этап образования пентоз и неокислительный этап (путь
возвращения пентоз в гексозы) составляют вместе циклический процесс.
34.
Дефект глюкозо-6-фосфатдегидрогеназы в эритроцитах приводит к дефицитуNADPH + H+, снижению концентрации восстановленной формы глутатиона и
окислению SH-групп молекул гемоглобина с образованием дисульфидных связей.
Этот процесс сопровождается агрегацией протомеров гемоглобина и формированием
телец Хайнца. Эритроциты теряют пластичность, необходимую для прохождения
через капилляры, нарушается целостность мембраны, что может привести к
гемолизу.
35.
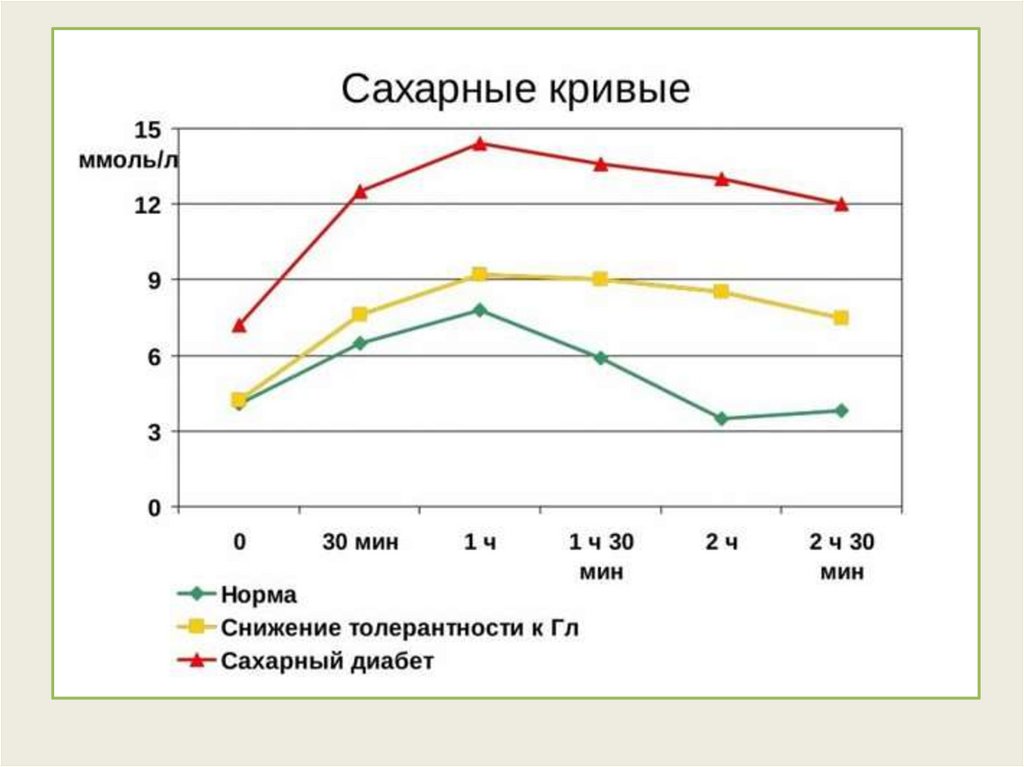
Регуляция содержания глюкозы в крови• Результатом регуляции метаболических путей
превращения глюкозы является постоянство концентрации
глюкозы в крови.
• Концентрация глюкозы в артериальной крови в течение
суток поддерживается на постоянном уровне 60-100 мг/дл
(3,3 — 5,5 ммоль/л)
36.
Источники глюкозы в крови в период пищеварения и во время голодания:1 — в период пищеварения углеводы пищи являются основным источником глюкозы в крови; 2 —
в постабсортивный период печень поставляет глюкозу в кровь за счет процессов гликогенолиза и
глюконеогенеза, причем в течение 8–12 ч уровень глюкозы в крови поддерживается в основном за
счет распада гликогена; 3 — глюконеогенез и гликоген печени участвуют в равной степени в
поддержании нормальной концентрации глюкозы; 4 — в течение суток гликоген печени
практически полностью исчерпывается и скорость глюконеогенеза увеличивается; 5 — при
длительном голодании (1 неделя и более) скорость глюконеогенеза уменьшется, но глюконеогенез
остается единственным источником глюкозы в крови
37.
38.
Общие симптомы СД1. Увеличение диуреза (полиурия);
2. Жажда (полидипсия);
3. Повышенный аппетит (полифагия);
4. Похудение;
5. Риск кетоацидотической комы;
6. Снижение иммунитета;
7. Кожный зуд;
8. Сонливость;
9. Снижение трудоспособности;
10.Слабость.
39.
Сахарный диабет I типаВозникает вследствие абсолютного дефицита инсулина, вызванного
аутоиммунным разрушением β-клеток поджелудочной железы или
вирусной инфекцией.
Сопровождается тяжелыми метаболическими нарушениями, возможно
развитие кетоацидотической комы.
В основном поражает детей, подростков, молодых людей до 30 лет.
Редко является семейным заболеванием.
На его долю приходится примерно 10-15% всех случаев
СД.
40.
Причины:1) Генетическая предрасположенность.
• Часто обусловленно дефектами генов 6-й хромосомы. Существует
предрасположенность к аутоиммунной агрессии к β-клеткам
поджелудочной железы.
• Генетические дефекты β-клеток поджелудочной железы
2) Действие цитотропных факторов.
• Вирусы (оспы, краснухи, кори, паротита)
• токсические вещества
• Стресс
• Аутоиммунные заболевания
• Онкология
41.
Острые клинические проявления СД IКетоацидотическая кома возникает когда концентрация кетоновых тел
становится выше 100 мг/дл (до 400-500мг/дл).
Гиперкетонемия приводит к:
1. ацидозу, который блокирует активность большинства ферментов,
в первую дыхательных, что вызывает гипоксию и снижение синтеза АТФ.
2. гиперосмолярности, которая приводит к дегидратации тканей и
нарушению водно-электролитного равновесия, с потерей ионов калия,
натрия, фосфора, магния, кальция, бикарбонатов.
3. Возникающая гипокалиемия ведет к гипотонии гладкой и
поперечно-полосатой мускулатуры, снижению тонуса сосудов, падению
АД, сердечной аритмии, гипотонии дыхательной мускулатуры с развитием
острой дыхательной недостаточности; атонии ЖКТ с парезом желудка и
развитием кишечной непроходимости и развитием острой почечной
недостаточности
42.
Сахарный диабет II типаИнсулинонезависимый, не склонен к кетоацидотической коме, не имеет
антител к β-клеткам.
На его долю приходится 85-90% всех случаев СД.
Страдают люди старше 40 лет, характеризуется высокой частотой семейных
форм.
Причины СД II типа:
• генетические дефекты рецепторов инсулина, у них снижается
чувствительность к инсулину;
• синтез дефектного инсулина с низкой биологической активностью;
• нарушение превращения проинсулина в инсулин;
• нарушение секреции инсулина;
• повреждение инсулина и его рецепторов антителами;
• повышение скорости катаболизма инсулина;
• действие контринсулярных гормонов;
• нарушение глюкозочувствительного механизма клеток и т.д.
43.
Сахарный диабет II типаМетаболизм СД II типа
Нарушения в углеводном обмене протекают аналогично нарушениям при СД
I типа.
Особенности метаболизма наблюдаются в нарушении липидного обмена,
так как жировая ткань содержит большое количество рецепторов к инсулину.
Высокий уровень глюкозы в крови стимулирует секрецию инсулина из βклеток, вызывая гиперинсулинемию.
Основным провоцирующим фактором, а также одним из ранних проявлений
СД II типа является ожирение.
44.
Острые осложнения СД1.
Гиперосмолярная кома – характерна для СД II типа и
наблюдается при высокой гипергликемии. Кома развивается
медленно, в течение нескольких дней, когда содержание глюкозы
в крови достигает 30-50 ммоль/л.
2.
Лактоацидотическая кома – характерна для СД II типа и возникает
при накоплении лактата, что ведёт к снижению чувствительности
адренорецепторов к катехоламинам (адреналин, норадреналин) и
развитию необратимого шока.
45.
Отдаленные осложнения СД• Превращение глюкозы в сорбитол (катаракта, нейропатии)
• Неферментативное гликозилирование различных белков, изменение
их свойств и активация их синтеза (HbA1C, микроангиопатии,
макроангиопатии, атеросклероз)
• Основную угрозу для больных представляют макрососудистые
осложнения (прежде всего инфаркт миокарда (ИМ)), доля которых в
структуре общей смертности достигает 65%).
46.
47.
Лабораторная диагностика сахарного диабета1. Глюкоза в крови и моче.
2. Кетоновые тела в крови и моче
3. Гликозилированный гемоглобин
цельной крови
4. Инсулин (проинсулин, С-пептид)
5. Антитела к инсулину
6. Антитела к β-клеткам
поджелудочной железы
7. Теста толерантности к глюкозе (ТТГ)
48.
49.
Лечение сахарного диабетаЛечение сахарного диабета зависит от его типа (I или II) и должно быть
комплексным.
Основные направления лечения:
1. Диетотерапия
2. Применение сахаропонижаюших средств
3. Инсулинотерапия
4. Лечение осложнений сахарного диабета
5. Патогенетическая терапия (Метформин улучшает чувствительность
периферических тканей к инсулину.↓ гиперинсулинемии)
Инсулинотерапия обязательна при лечении СД I типа.
Перспективные методы лечения:
1. Трансплантация изолированных -клеток или островков
поджелудочной железы;
2. Стимуляция регенерации панкреатических островков.
50.
Задание для самостоятельнойработы
• Используя материал учебника и
образовательные ресурсы интернета
подготовьте ответ на вопрос:
• «Влияние углеводов на состояние мягких и
твердых тканей полости рта. Проявление
сахарного диабета в ротовой полости»
51.
ЗаключениеИтак, изложенные сведения позволяют сделать вывод о том, что координация
скоростей гликолиза, глюконеогенеза, синтеза и распада гликогена с участием
гормонов обеспечивает:
• предотвращение чрезмерного повышения концентрации глюкозы в крови
после приема пищи;
• запасание гликогена и его использование в промежутках между приемами
пищи;
• снабжение глюкозой мышц, потребность которых в энергии быстро возрастает при мышечной работе;
• снабжение глюкозой клеток, которые при голодании в качестве источника
энергии используют преимущественно глюкозу (нервные клетки, эритроциты,
мозговое вещество почек, семенники).