





































 – истинные мутации")



 изменчивость")














")
















 biology
biologySimilar presentations:










Закономерности наследственности и изменчивости
1. Г е н е т и к а
Закономерности наследственностии
изменчивости
2. Законы Менделя
1 закон.Закон единообразия или закон доминирования.
У гибридов 1-го поколения проявляется признак одного из родителей в неизменном
виде – признак детерминирован
2 закон. Закон расщипления
Рецессивный признак не исчезает, он сохраняется у гибридов первого
поколения в неизменном виде и проявляется у гибридов второго
поколения в соотношении 3:1 – признак детерминирован
3 закон.
Закон независимого наследования
Признаки наследуются независимо друг от друга (каждый по отдельности
признак у гибридов 2-го поколения дает расщепление 3:1), образует
все возможные сочетания признаков в соотношении 9:3:3:1 –
наследственность дискретна.
Наследственность детерминирована и дискретна
3. Общие закономерности расщепления при скрещивании полигетерозигот
Полигетерозиготы – это организмы, гетерозиготные по многимгенам. Если число генов равно n, то при полном
доминировании наблюдаются следующие закономерности:
Число
генов
Число
типов
гамет
Число
генотипов
Расщепление
генотипу
(а при неполном
доминировании
и по фенотипу)
1
2
3
2
4
по
Число
фенотипов
Расщепление
по фенотипу
при полном
доминировании
1:2:1
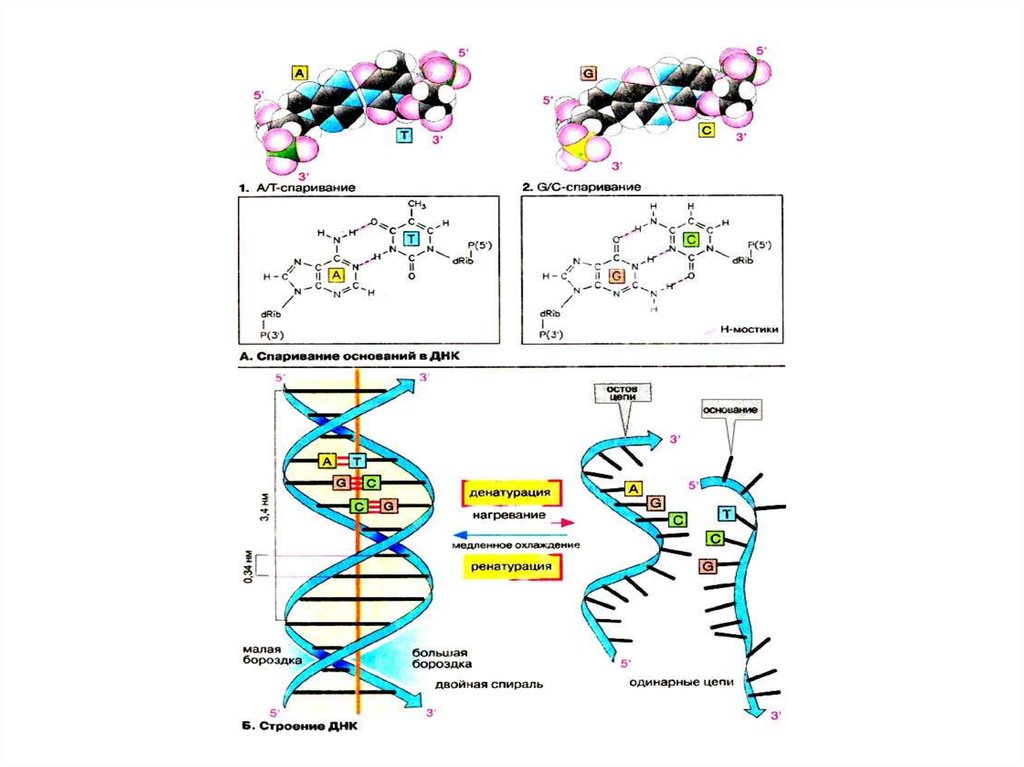
2
3:1
9
1:2:1:2:4:2:1:2:1
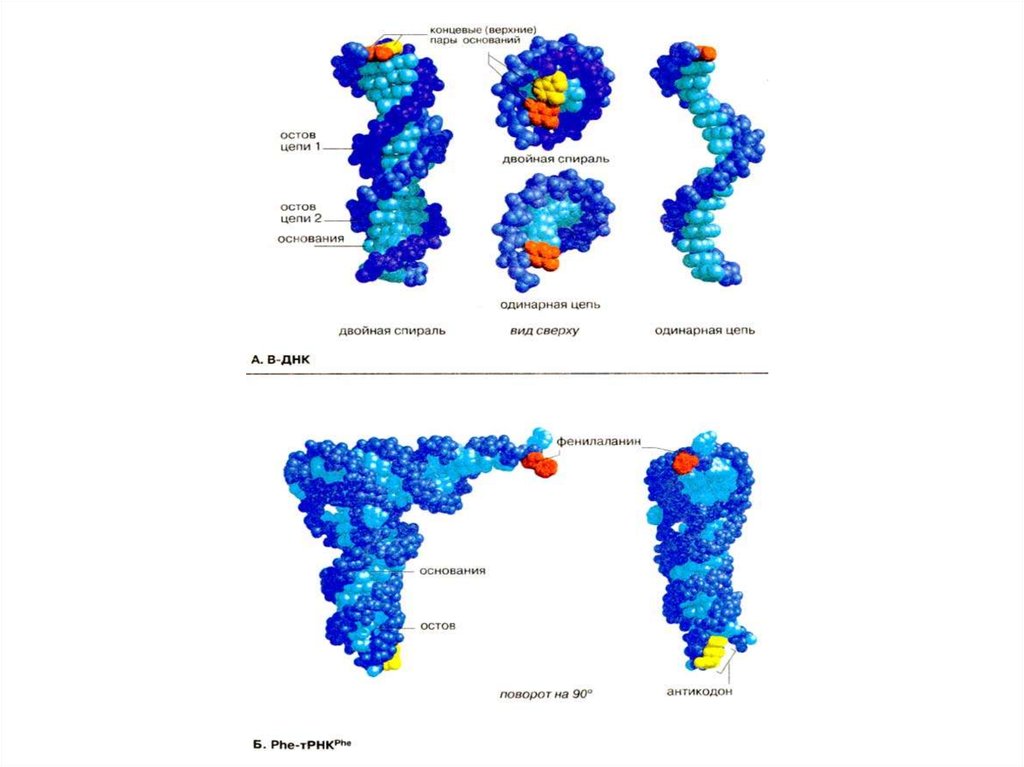
4
9:3:3:1
8
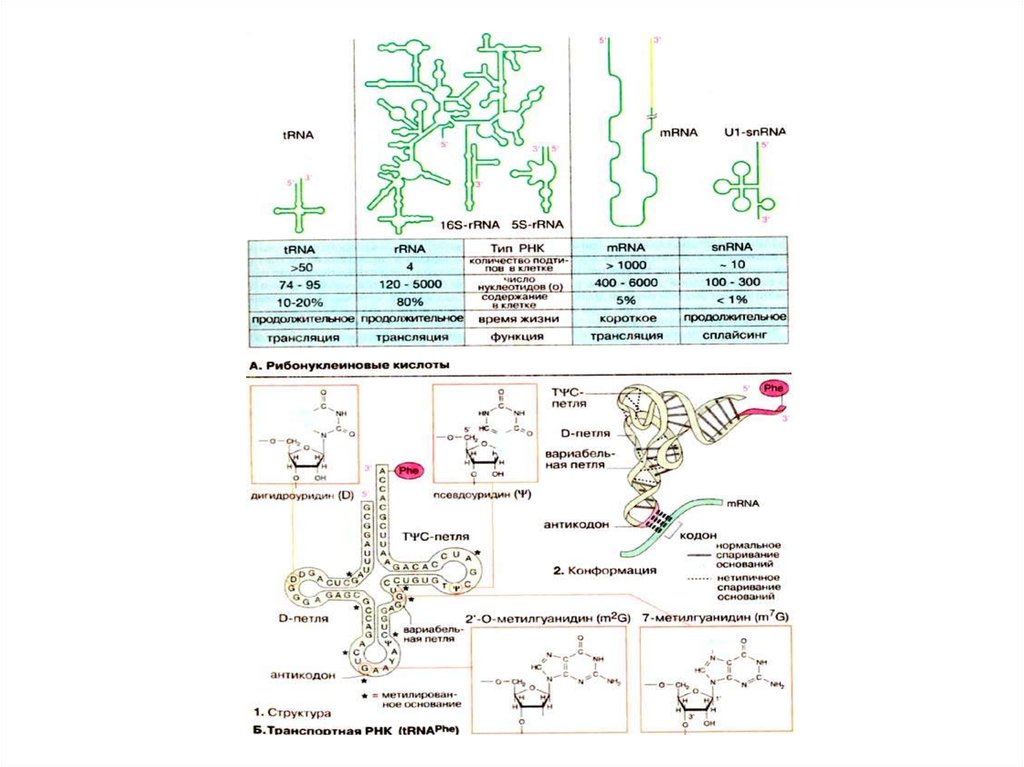
27:9:9:3:9:3:3:1
2n
(3 : 1) n
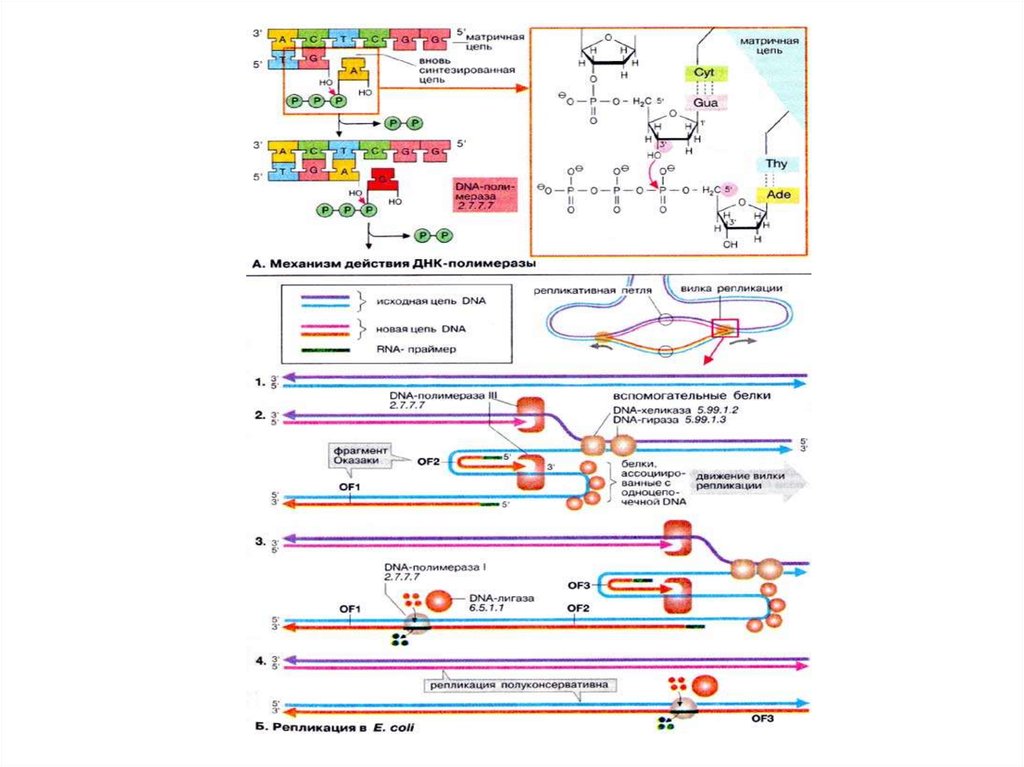
3
8
27
1:2:1:2:4:2:1:2:1:
2:4:2:4:8:4:2:4:2:
1:2:1:2:4:2:1:2:1
N
2n
3n
(1 : 2 : 1) n
4. Возвратное скрещивание
Скрещивание гибридных форм с родительскими формами(гомозиготами по доминантному или рецессивному признакам или
особями из чистых линий)
Анализирующее скрещивание позволяет выявить
гетерозиготность фенотипически доминантной форма, т.е.
определить генотип фенотипически доминантной формы. Для
этого анализируемую форму скрещивают с гомозиготой по
рецессиву. Анализирующее скрещивание один из видов
возвратного скрешивания.
АА x аа
Аа – в Fв расщепления нет
Аа x аа
1 Аа : 1 аа – расщепление по генотипу
в Fв расщепление по фенотипу 1[А-] : 1 [аа]
5.
Насыщающиескрещивания
При скрещивании чистопородных
коричневых и серых норок все гибриды
первого поколения коричневые. Гибридные
коричневые норки скрещиваются с
чистопородными коричневыми. Все их
потомки – коричневые (единообразны по
фенотипу). Однако по генотипу существует
расщепление – 1 часть гомозигот : 1 часть
гетерозигот.
Таким образом, при скрещивании
доминантных гомозигот (АА) с любыми
формами (Аа или аа) расщепление по
фенотипу отсутствует. Но при скрещивании
гетерозигот (Аа) с доминантными
гомозиготами (АА) при единообразии по
фенотипу всегда существует расщепление
по генотипу в соотношении 1:1.
Такое скрещивание используется для
подавления рецессивных признаков, для
выведения новых сортов и пород и
насыщения их доминантными аллелями –
поэтому его называют насыщающим.
6. Рецепроктное или обратное скрещивание
Данный вид скрещивания позволяет определить связанноли наследование того или иного признака с полом, т.е.
зависит ли фенотипическое проявление признака от того у
кого – самца или самки - локализован феноопределяющий
ген (аллель)
OА-ХOаа
OааХOА-
7. Сцепленное наследование
Законы Моргана (хромосомная теориинаследования):
Гены наследуются совместно образуя группу сцепления или
хромосому
Локусы генов располагаются в хромосоме линейно
Хромосомы способны конъюгировать и обмениваться
гомологичными участками (кроссинговер), образуя
рекомбинанты. Сила сцепления между локусами зависит от
расстояния между ними.
- абсолютное сцепление- гены расположены рядом
- относительное сцепление- возможны рекомбинации(до 46%)
- практически независимое наследование - количество
рекомбинантов около 50%
Сила сцепления обратно пропорциональна расстоянию между
локусами.
Вероятность кроссинговера прямо пропорционально расстоянию
между локусами и обратно пропорциональна силе сцепления
8. Расстояние между локусами
Морган предложил измерять расстояниемежду локусами в процентах
рекомбинантных форм в потомстве.
1% = 1 М
Пример: в потомстве из 180 мух 45 имели новые
сочитания 2-х признаков, т.е. отличались сочитанием
признаков от родительских форм – были
рекомбинантами
расстояние между локусами 2 анализируемых генов
находится так:
180 - 100%
45 - х %
х = 25%
9. Полигенное наследование
В большинстве случаев один признак контролируется множествомразных (неаллельных) генов. Такое наследование признаков, при
котором за формирование одного признака отвечает два и более
неаллельных гена, называется полигенным. При полигенном
наследовании гены неравноценны по степени воздействия на
признак. Например, можно выделить следующие группы генов:
Олигогены – это главные гены, которые в наибольшей степени
ответственны за формирование признака. Обеспечивают
дискретную (прерывистую) изменчивость качественных признаков.
Супрессоры – гены, подавляющие действие других генов.
Бустеры – гены, усиливающие действие других генов.
Модификаторы – гены, оказывающие незначительное влияние на
формирование признака, уточняющие действие других генов.
Полигены – множество генов, оказывающих сходное действие.
Обеспечивают непрерывную изменчивость количественных
10. Основные типы взаимодействия генов : комплементарность, эпистаз и полимерия.
1. Комплементарность. Новообразования прискрещивании
Комплементарность – это такой тип полигенного
наследования, при котором неаллельные гены взаимно
дополняют друг друга. В простейшем случае
рассматривается два гена, каждый из которых
представлен двумя аллелями, причем, аллель А
полностью доминирует над аллелем а, а аллель В
полностью доминирует над аллелем b. Если два
комплементарных гена не сцеплены между собой, то во
втором поколении возможны следующие расщепления
по фенотипу: 9:3:3:1, 9:6:1, 9:7
11. 2. Э п и с т а з
Эпистаз – это такой способ взаимодействия генов, прикотором действие одного гена подавляется действием
другого, неаллельного гена. При этом ген-подавитель
называется эпистатическим геном, а подавляемый ген –
гипостатическим.
Различают рецессивный и доминантный эпистаз.
Обычно рассматривают случаи полного доминирования
для каждого неаллельного гена, причем эпистатический и
гипостатический гены не сцеплены между собой. Тогда
при рецессивном эпистазе в F2 наблюдаются расщепления
9 : 3 : 4 или 9 : 7, а при доминантном эпистазе – 13:3 или
12:3:1.
12. 3. Полимерия
Полимерия – это такой тип полигенного наследования,при котором признак определяется взаимодействием
нескольких пар неаллельных генов со сходным действием.
Такие гены называют гомологичными и обозначают
сходными символами, например, А1, А2, А3 и т.д.
Некумулятивная полимерия. Для качественных
признаков характерна некумулятивная полимерия с полным
доминированием и расщеплением 15 : 1 (в случае действия
двух пар аллелей)
Кумулятивная полимерия. Для количественных
признаков характерна кумулятивная полимерия с неполным
доминированием и расщеплением 1 : 4 : 6 : 4 : 1 (в случае
действия двух пар аллелей).
13. Межаллельные взаимодействия
Неполное доминирование – транскрипция рецессивногогена
не
подавляется
действием
доминантного.
В
гетерозиготном состоянии у гибридов фенотипически
появляется промежуточное значение признака.
Кодоминирование
–
совместное
действие
генов
с
одинаковой фенотипической экспрессией. В результате
гибриды имеют новый признак.
Множественный аллелизм – существование в популяции 3
и более аллелей одного гена (серии генов), возникших в
результате многократных мутаций в одном и том же
локусе.
Аллели в серии проявляют или соотносительное доминирование, или кодоминирование. У
Сверхдоминирование
–
в
гетерозиготном
состоянии
признак проявляется с большей фенотипической силой
чем у гомозигот. Такое явление (гетерозисная сила)
проявляется
при
скрещивании
(гетерозисе)
особей
инбридных
линий.
Гетерозисная
сила
объясняется
возрастанием общей жизнестойкости и жизнеспособности
гибридов в следствии перевода в гетерозиготное состояние
большинства рецессивных «вредных» генов.
14. Наследование признаков, сцепленных с полом, у млекопитающих
Наследование признаков, сцепленных сполом, у млекопитающих
У млекопитающих самки являются гомогаметным полом (их
половые хромосомы представлены двумя Х–хромосомами), а
самцы – гетерогаметным полом (их половые хромосомы
представлены одной Х–хромосомой и одной Y–хромосомой).
Пол у млекопитающих определяется наличием Y–хромосомы. В
состав Y-хромосомы входит H–Y–антиген, который запускает
экспрессию генов, определяющих формирование основных
мужских половых признаков. В большинстве случаев у самцов
млекопитающих Y–хромосома практически не несет генов. Однако
некоторые аномальные признаки самцов (например, волосатые
уши у мужчин, ихтиодермия или развитие у них перепонок между
вторым и третьим пальцами ног) обусловлены изменениями в Y–
хромосоме. Подобные аномалии стопроцентно наследуются по
мужской линии от отца ко всем его сыновьям и далее. Такие
признаки называются голандрическими.
15.
Несмотря на то, что женские особи млекопитающихимеют две Х-хромосомы, а мужские – только одну,
экспрессия генов Х-хромосомы происходит на одном
и том же уровне у обоих полов. Это обусловлено тем,
что в каждой клетке самки функционирует лишь одна
из двух Х–хромосом. Вторая Х–хромосома на ранних
стадиях эмбрионального развития (в период
имплантации) инактивируется и превращается в
генетически инертное тельце Барра (половой
хроматин). При этом в разных клетках отцовская и
материнская Х-хромосомы выключаются случайно.
Состояние инактивации данной Х-хромосомы
наследуется в ряду клеточных делений. Таким
образом, женские особи, гетерозиготные по генам
половых хромосом, представляют собой мозаики.
16. ГЕНЕТИКИ ПОПУЛЯЦИЙ
С точки зрения генетики,
популяция – это генетическая система, обладающая исторически
сложившейся генетической структурой. Основные положения
популяционной генетики сложились на основании изучения природных и
модельных популяций высших раздельнополых животных (моллюсков,
насекомых, позвоночных), которые воспроизводят себя с помощью
нормального полового размножения – амфимиксиса, или объединения
женских и мужских гамет. В таких случаях группировка особей, способных
скрещиваться между собой и производить полноценное (т.е.
жизнеспособное и плодовитое) потомство, называется генетической, или
менделевской популяцией. В свою очередь, потомки, достигшие
половозрелости, также должны скрещиваться между собой и производить
полноценное потомство, то есть популяция должна существовать
длительное число поколений.
Таким образом, с точки зрения генетики, популяция представляет собой
множество особей, объединенных достаточно высокой степенью родства.
В рамках генетического подхода выделяется представление об идеальной
популяции.
17. Характеристика идеальной популяции
• Идеальная популяция – это абстрактное понятие,которое широко используется в моделировании
микроэволюционных процессов. При описании
систем скрещивания в идеальной популяции широко
используется понятие панмиксии – случайного
свободного скрещивания, при котором вероятность
встречи гамет не зависит ни от генотипа, ни от
возраста скрещивающихся особей. Если исключить
половой отбор, то к панмиктической популяции
применима концепция гаметного резервуара,
согласно которой в популяции в период размножения
формируется гаметный резервуар (генный пул),
включающий банк женских гамет и банк мужских
гамет. Если члены популяции равноудалены друг от
друга, то встреча гамет и формирование зигот
происходят случайным образом.
18. Условия существования идеальной популяции
1. Рост численности популяции ничем не ограничен2. Миграция (обмен генетическим материалом между
популяциями) отсутствует
3. Особи со всеми видами генотипа обладают одинаковой
жизнеспособностью. Естественный отбор не оказывает заметного
влияния на частоту большинства аллелей
4. Особи с разными генотипами имеют возможность
скрещиваться с равной вероятностью
5. Скрещивание особей равновероятно
6. Скорость прямых мутаций равна скорости обратных (нет
мутационных изменений генотипа и генофонда)
7. Отсутствуют межаллельные и межгенные взаимодействия
19.
Реальные популяции в большей или меньшей степени отличаются от
идеальной. Одним из наиболее существенных отличий является множество
способов воспроизведения.
По способу воспроизведения различают следующие типы популяций:
амфимиктические – основным способом размножения является нормальное
половое воспроизведение;
амфимиктические панмиктические – при формировании брачных пар
наблюдается панмиксия (свободное скрещивание);
амфимиктические инбредные – при формирование брачных пар
наблюдается близкородственное скрещивание (инбридинг, инцухт, инцест);
крайним случаем близкородственного скрещивания является
самооплодотворение;
апомиктические – наблюдаются различные отклонения от нормального
полового процесса, например, апомиксис, партеногенез, гиногенез, андрогенез;
наблюдается у агамных (бесполых) форм;
клональные – при отсутствии полового процесса и размножении только
вегетативным путем или с помощью спор бесполого размножения (например,
конидий); частным случаем клонирования является полиэмбриония – развитие
нескольких зародышей из одной зиготы:
комбинированные – например, клонально-амфимиктические при метагенезе у
кишечнополостных (чередовании бесполого и полового размножения) и
гетерогонии (чередовании партеногенетического и амфимиктического поколений
у червей, некоторых членистоногих и низших хордовых).
20. Определения
Панмиксия(свободное
скрещивание)
означает,
что
на
формирование брачных пар не влияет генотип или возраст особей,
участвующих в размножении. Фактически это означает, что
рассматриваемый признак не оказывает заметного влияния на
формирование брачных пар.
Инбридинг – близкородственное скрещивание у животных; инцухт
–
близкородственное
скрещивание
у
растений;
инцест
(кровосмешение) – при которых не происходит объединения двух
клеток. Обычно этот термин используют по отношению к растениям.
При
апомиксисе
новый
организм
может
развиваться
из
неоплодотворенной яйцеклетки (см. партеногенез), а также из какойлибо другой специализированной клетки зародышевого мешка
(например, из клеток–антипод или синергид), реже – непосредственно
из клеток нуцеллуса или покровов семязачатка. Близкородственное
скрещивание у человека.
Апомиксис – это множество форм образования зародышей.
Примеры растений–апомиктов: ястребинки, одуванчики, манжетки.
21.
Партеногенез – это девиантная форма полового процесса, при которойновый организм развивается из неоплодотворенной яйцеклетки без
участия мужских гамет. Различают нередуцированный партеногенез с
развитием зародыша из диплоидной клетки и редуцированный
партеногенез с развитием зародыша из гаплоидной яйцеклетки. Как
правило, партеногенез чередуется с нормальным половым размножением
(при цикломорфозе у коловраток, дафний, тлей).
Гиногенез – это девиантная форма полового процесса, при которой
мужские гаметы служат для стимуляции развития нового организма из
яйцеклетки, но оплодотворения не происходит, и мужское ядро
(пронуклеус) погибает. В этом случае у дочернего организма сохраняются
только материнские хромосомы. Гиногенез встречается у гибридов рыб,
земноводных, а также в бессамцовых популяциях.
Андрогенез – это девиантная форма полового процесса, при которой
происходит оплодотворение, но затем женское ядро (пронуклеус)
погибает, а мужское ядро замещает его в качестве ядра зиготы. В этом
случае у дочернего организма сохраняются только отцовские хромосомы.
Андрогенез обычно наблюдается в лабораторных условиях.
Агамные формы – организмы, у которых отсутствует нормальный
половой процесс
22. Генетическая структура популяций
• Каждая популяция обладает собственнойгенетической структурой. Генетическая
структура популяций определяется исходным
соотношением аллелей, естественным
отбором и элементарными эволюционными
факторами (мутационный процесс и
давление мутаций, изоляция, популяционные
волны, генетико-автоматические процессы,
эффект основателя, миграции и др.). Для
описания генетической структуры популяций
используются понятия «аллелофонд» и
«генофонд».
23. Аллелофонд
• . Аллелофонд популяции – это совокупность аллелейв популяции. Если рассматриваются два аллеля
одного гена: А и а, то структура аллелофонда
описывается уравнением: pA + qa = 1. В этом
уравнении символом pA обозначается
относительная частота аллеля А, символом qa –
относительная частота аллеля а.
• Популяции, в которых структура аллелофонда
остается относительно постоянной в течение
длительного времени, называются стационарными.
• Если рассматриваются три аллеля одного гена: а1,
а2,, а3, то структура аллелофонда описывается
уравнением: p а1 + q а2 + r а3 = 1. В этом уравнении
символами p, q, r обозначаются соответствующие
частоты аллелей.
24. Генофонд.
Термин генофонд употребляется в разных значениях. Основоположник
учения о генофонде и геногеографии Александр Сергеевич
Серебровский называл генофондом «совокупность всех генов данного
вида..., чтобы подчеркнуть мысль о том, что в лице генофонда мы
имеем такие же национальные богатства, как и в лице наших запасов
угля, скрытых в наших недрах» (1928). Однако это выражение в
настоящее время используется для определения генетического
потенциала, а генофондом называют совокупность всех генотипов в
популяции.
При изучении природных популяций часто приходится сталкиваться с
полным доминированием: фенотипы гомозигот АА и гетерозигот Аа
неразличимы. Кроме того, в природе широко распространено
полигенное определение признаков, причем типы взаимодействия
неаллельных генов (комплементарность, эпистаз, полимерия) не
всегда известны. Поэтому на практике часто изучают не генофонд, а
фенофонд популяций, то есть соотношение фенотипов. В настоящее
время развивается раздел генетики популяций, который называется
фенетика популяций.
25. Закон Харди–Вайнберга – основной закон популяционной генетики
Структура генофонда в панмиктической стационарной популяции
описывается основным законом популяционной генетики – законом
Харди-Вайнберга, который гласит, что в идеальной популяции
существует постоянное соотношение относительных частот
аллелей и генотипов, которое описывается уравнением: (p A +
q a)2 = р2 АА + 2∙р∙q Aa + q2 aa = 1
Если известны относительные частоты аллелей p и q и общая
численность популяции Nобщ, то можно рассчитать ожидаемую, или
расчетную абсолютную частоту (то есть численность особей) каждого
генотипа. Для этого каждый член уравнения нужно умножить на Nобщ:
p2 AA · Nобщ + 2·p·q Aa · Nобщ + q2 aa · Nобщ = Nобщ
В данном уравнении:
p2 AA · Nобщ – ожидаемая абсолютная частота (численность)
доминантных гомозигот АА
2·p·q Aa · Nобщ – ожидаемая абсолютная частота (численность)
гетерозигот Аа
q2 aa · Nобщ – ожидаемая абсолютная частота (численность)
рецессивных гомозигот аа
26. Действие закона Харди-Вайнберга при полном доминировании
Известно, что черная окраска шерсти у кошек определяется генотипом аа. При этомчерная окраска может быть или сплошной, или частичной. Генотипы АА и Аа
обусловливают все остальное разнообразие типов окраски, но черный цвет при
этом полностью отсутствует.
Предположим, что в одной из городских популяций кошек на о. Сахалин из 100
просмотренных животных полную или частичную черную окраску имели 36
животных.
Прямой расчет структуры аллелофонда популяции в этом случае невозможен из-за
полного доминирования: гомозиготы АА и гетерозиготы Аа фенотипически
неразличимы. Согласно уравнению Харди-Вайнберга частота черных кошек
составляет q2 аа. Тогда можно рассчитать частоты аллелей:
q2aa = 36/100 = 0,36; qa = 0,36 –1/2 =0,6; pA = 1 – 0,6 = 0,4
Таким образом, структура аллелофонда данной популяции описывается
соотношением: р А + q a = 0,4 + 0,6 = 1. Частота рецессивного аллеля оказалась
выше, чем частота доминантного.
Рассчитаем частоты генотипов:
р2 АА = 0,42 = 0,16; 2 pq Аа = 2 х 0,4 х 0,6 = 0,48; q2aa = 0,62 = 0,36
Рассчитаем число особей с разными генотипами:
n AA= 0,16 х100=16 особей;
n Aa= 0,48х100=48 особей;
п aa = 0,36х 100=36 особей.
27. Практическое значение закона Харди–Вайнберга
1. В здравоохранении – позволяет оценить популяционный риск генетически
обусловленных заболеваний, поскольку каждая популяция обладает
собственным аллелофондом и, соответственно, разными частотами
неблагоприятных аллелей. Зная частоты рождения детей с наследственными
заболеваниями, можно рассчитать структуру аллелофонда. В то же время, зная
частоты неблагоприятных аллелей, можно предсказать риск рождения больного
ребенка.
Пример 1. Известно, что альбинизм – это аутосомно-рецессивное заболевание.
Установлено, что в большинстве европейских популяций частота рождения
детей-альбиносов составляет 1 на 20 тысяч новорожденных. Следовательно,
q2aa = 1/20000 = 0,00005; qa = 0,00005–1/2 = 0,007; pA = 1 – 0,007 = 0,993 ≈ 1
Поскольку для редких заболеваний рА ≈ 1, то частоту гетерозиготных носителей
можно рассчитать по формуле 2·q. В данной популяции частота гетерозиготных
носителей аллеля альбинизма составляет 2 q Аа = 2 х 0,007 = 0,014, или
примерно каждый семидесятый член популяции.
Пример 2. Пусть в одной из популяций у 1% населения выявлен рецессивный
аллель, который не встречается в гомозиготном состоянии (можно
предположить, что в гомозиготном состоянии этот аллель летален). Тогда 2 q Аа
= 0,01, следовательно, qa = 0,01:2 = 0,005. Зная частоту рецессивного аллеля,
можно установить частоту гибели зародышей–гомозигот: q2aa = 0,0052 =
0,000025 (25 на миллион, или 1 на 40 тысяч).
28.
2. В селекции – позволяет выявить генетический потенциал исходногоматериала (природных популяций, а также сортов и пород народной
селекции), поскольку разные сорта и породы характеризуются собственными
аллелофондами, которые могут быть рассчитаны с помощью закона ХардиВайнберга. Если в исходном материале выявлена высокая частота
требуемого аллеля, то можно ожидать быстрого получения желаемого
результата при отборе. Если же частота требуемого аллеля низка, то нужно
или искать другой исходный материал, или вводить требуемый аллель из
других популяций (сортов и пород).
3. В экологии – позволяет выявить влияние самых разнообразных
факторов на популяции. Дело в том, что, оставаясь фенотипически
однородной, популяция может существенно изменять свою генетическую
структуру под воздействием ионизирующего излучения, электромагнитных
полей и других неблагоприятных факторов. По отклонениям фактических
частот генотипов от расчетных величин можно установить эффект действия
экологических факторов. (При этом нужно строго соблюдать принцип
единственного различия. Пусть изучается влияние содержания тяжелых
металлов в почве на генетическую структуру популяций определенного вида
растений. Тогда должны сравниваться две популяции, обитающие в крайне
сходных условиях. Единственное различие в условиях обитания должно
заключаться в различном содержании определенного металла в почве).
29. Генетические параметры популяции
При описании популяций или их сравнении между собой используютцелый ряд генетических характеристик.
• Полиморфизм. Популяция называется полиморфной по данному
локусу, если в ней встречается два или большее число аллелей. Если
локус представлен единственным аллелем, говорят о мономорфизме.
Исследуя много локусов, можно определить среди них долю
полиморфных, т.е. оценить степень полиморфизма, которая является
показателем генетического разнообразия популяции.
• Гетерозиготность. Важной генетической характеристикой
популяции является гетерозиготность – частота гетерозиготных особей
в популяции. Она отражает также генетическое разнообразие.
• Коэффициент инбридинга. С помощью этого коэффициента
оценивают распространенность близкородственных скрещиваний в
популяции.
• Ассоциация генов. Частоты аллелей разных генов могут зависеть
друг от друга, что характеризуется коэффициентами ассоциации.
• Генетические расстояния. Разные популяции отличаются друг от
друга по частоте аллелей. Для количественной оценки этих различий
предложены показатели, называемые генетическими расстояниями.
30.
Различные популяционно-генетические процессы по-разномувлияют на эти параметры: инбридинг приводит к уменьшению
доли гетерозиготных особей; мутации и миграции увеличивают,
а дрейф уменьшает генетическое разнообразие популяций;
отбор изменяет частоты генов и генотипов; генный дрейф
увеличивает, а миграции уменьшают генетические расстояния и
т.д. Зная эти закономерности, можно количественно
исследовать генетическую структуру популяций и
прогнозировать ее возможные изменения. Этому способствует
солидная теоретическая база популяционной генетики –
популяционно-генетические процессы математически
формализованы и описаны уравнениями динамики. Для
проверки различных гипотез о генетических процессах в
популяциях разработаны статистические модели и критерии.
Прилагая эти подходы и методы к исследованию популяций
человека, животных, растений и микроорганизмов, можно
решить многие проблемы эволюции, экологии, медицины,
селекции и др. Рассмотрим несколько примеров,
демонстрирующих связь популяционной генетики с другими
науками.
31. ИЗМЕНЧИВОСТЬ
32.
• Изменчивость -- это возникновениеиндивидуальных различий. На основе
изменчивости организмов появляется
генетическое разнообразие форм,
которые в результате действия
естественного отбора преобразуются в
новые подвиды и виды. Различают
изменчивость фенотипическую и
мутационную, или генотипическую.
33.
Классификация форм изменчивостьФенотипическая (ненаследуемая - паратипическая)
Модификационная
Фенокопии – фенотипическое копирование мутаций при критическом
изменении действующего фактора среды
Случайная – не обусловлена воздействием типичных факторов,
обычно носит травматический характер
Генотипическая (наследуемая)
Комбинативная
Мутационная: генные (точечные), хромосомные, геномные.
Соотносительная (коррелятивная)
34. Модификационная изменчивость
Модификационная изменчивость не вызывает изменений
генотипа, она связана с реакцией данного, одного и того же
генотипа на изменение внешней среды: в оптимальных условиях
выявляется максимум возможностей, присущих данному генотипу.
В этом случае все особи с одинаковым генотипом отвечают на
внешние условия одинаково (Ч. Дарвин этот тип изменчивости
назвал определенной изменчивостью).
Модификационная изменчивость обычно колеблется в
определенных пределах. Степень варьирования признака у
организма, то есть пределы модификационной изменчивости,
называется нормой реакции.
Широкая норма реакции свойственна таким признакам, как удои
молока, размеры листьев, окраска у некоторых бабочек;
узкая норма реакции -- жирности молока, яйценоскости у кур,
интенсивности окраски венчиков у цветков и другое.
Фенотип формируется в результате взаимодействий генотипа и
факторов среды. Фенотипические признаки не передаются от
родителей потомкам, наследуется лишь норма реакции, то есть
характер реагирования на изменение окружающих условий. У
гетерозиготных организмов при изменении условий среды можно
вызвать различные проявления данного признака
35.
Свойства модификаций:
1) ненаследуемость;
2) групповой характер изменений;
3) соотнесение изменений действию
определенного фактора среды;
4) обусловленность пределов
изменчивости генотипом.
36. Генотипическая изменчивость
Генотипическая изменчивость подразделяется на мутационную икомбинативную.
Мутациями называются скачкообразные и устойчивые изменения единиц
наследственности - генов, влекущие за собой изменения наследственных
признаков. Термин “мутация” был впервые введен де Фризом. Мутации
обязательно вызывают изменения генотипа, которые наследуются
потомством и не связаны со скрещиванием и рекомбинацией генов.
Классификация мутаций.
Мутации можно объединять, в группы -классифицировать по характеру
проявления, по месту или, по уровню их возникновения.
• Мутации по характеру проявления бывают доминантными и
рецессивными. Мутации нередко понижают жизнеспособность или
плодовитость. Мутации, резко снижающие жизнеспособность, частично
или полностью останавливающие развитие, называют полулетальными а
несовместимые с жизнью - летальными.
Мутации подразделяют по месту их возникновения. Мутация, возникшая в
половых клетках, не влияет на признаки данного организма, а проявляется
только в следующем поколении. Такие мутации называют
генеративными. Если изменяются гены в соматических клетках, такие
мутации проявляются у данного организма и не передаются потомству при
половом размножении. Но при бесполом размножении, если организм
развивается из клетки или группы клеток, имеющих изменившийся -мутировавший -- ген, мутации могут передаваться потомству. Такие
мутации называют соматическими.
37.
Мутации классифицируют по уровню их возникновения.Геномные - изменение кариотипа (изменение числа хромосом).
Полиплоидия -- увеличение числа хромосом, кратное гаплоидному набору. В
соответствии с этим у растений различают триплоиды (Зп), тетраплоиды (4п) и т.д. В
растениеводстве известно более 500 полиплоидов (сахарная свекла, виноград,
гречиха, мята, редис, лук и др.). Все они выделяются большой вегетативной массой и
имеют большую хозяйственную ценность.Полиплоиды получают в результате
воздействия на растения температуры, ионизирующей радиации, химических
веществ (колхицин), которые разрушают веретено деления клетки. У таких растений
гаметы диплоидны, а при слиянии с гаплоидными половыми клетками партнера в
зиготе возникает триплоидный набор хромосом (2п + п = Зп). Такие триплоиды не
образуют семян, они бесплодны, но высокоурожайны. Четные полиплоиды образуют
семена.
Гетероплоидия - изменение числа хромосом, не кратное гаплоидному набору. При
этом набор хромосом в клетке может быть увеличен на одну, две, три хромосомы (2п
+ 1; 2п + 2; 2п + 3) или уменьшен на одну хромосому (2л-1). Например, у человека с
снндромом Дауна оказывается одна лишняя хромосома по 21-й паре и кариотип
такого человека составляет 47 хромосом. У людей с синдромом Шерешевского -Тернера (2п-1) отсутствует одна Х-хромосома и в кариотипе остается 45 хромосом.
Эти и другие подобные отклонения числовых отношений в кариотипе человека
сопровождаются расстройством здоровья, нарушением психики и телосложения,
снижением жизнеспособности и др.
38.
• Хромосомные мутации связаны с изменением структурыхромосом.
Существуют следующие виды перестроек хромосом:
делеции - отрыв различных участков хромосомы,
дупликации - удвоение отдельных фрагментов,
инверсии - поворот участка хромосомы на 180°
транслокации - присоединение отдельного участка
хромосомы к другой хромосоме.
транспозиция - перенос участка хромосомы в другой
участок той же хромосомы.
Подобное изменение влечет за собой нарушение
генетического гомеостаза, функции генов в хромосоме и
наследственных свойств организма, а иногда и его гибель.
39. Генные мутации (точечные) – истинные мутации
Затрагивают структуру самого гена и влекут за собой изменение свойстворганизма (гемофилия, дальтонизм, альбинизм, окраска венчиков
цветков и т.д.). Они могут быть доминантными и рецессивными.
Первые проявляются как у гомозигот так и у гетерозигот, вторые -только у гомозигот. Одни мутации оказывают на организм
положительное действие, другие безразличны, а третьи вредны,
вызывая либо гибель организма, либо ослабление его
жизнеспособности (например, серповидноклеточная анемия,
гемофилия у человека).
Существуют следующие виды генных мутаций:
делеции - потеря участков генов,
дупликации - удвоение отдельных фрагментов гена,
инверсии - поворот участка гена на 180°
транслокации – встраивания в ген участка не гомологичного гена.
транспозиция - перенос участка гена в другой участок того же гена.
Потеря или добавление участка гена вызывает изменение рамки
считывание и нарушает последовательность нуклеотидов, а
следовательно и изменение качества триплетов. Данный вид генных
мутаций наиболее разрушителен.
40.
При выведении новых сортов растений и штаммов микроорганизмов
используют индуцированные мутации, искусственно вызываемые
теми или иными мутагенными факторами – мутагенами (рентгеновские
или ультрафиолетовые лучи, химические вещества).
В природе мутации носят спонтанный характер
Мутации могут быть:
ядерные
внеядерные (цитоплазматические) – возникают в клеточных
структурах имеющих собственное ДНК (митохондрии, пластиды,
плазмиды и т.д.), передаются по материнской линии, т.к. при
оплодотворении цитоплазматические структуры сперматозоида в
формировании зиготы не участвуют.
Способность к мутированию - одно из свойств гена. Каждая
отдельная мутация вызывается какой-то причиной, но в
большинстве случаев эти причины неизвестны. Мутации
связаны с изменениями во внешней среде. Это убедительно
доказывается тем, что путем воздействия внешними
факторами удается резко повысить их число.
41. Свойства мутаций.
• 1. Мутации возникают внезапно, скачкообразно.• 2. Мутации наследственны, то есть стойко
передаются из поколения в поколение.
• 3. Мутации ненаправленные - мутировать может
любой локус, вызывая изменения как
незначительных, так и жизненно важных признаков.
• 4. Одни и те же мутации могут возникать повторно.
• 5. По своему проявлению мутации могут быть
полезными и вредными, доминантными и
рецессивными.
42. Комбинативная изменчивость
Комбинативная наследственная изменчивость возникает в результате1)
обмена гомологичными участками гомологичных хромосом в процессе
мейоза;
2) как следствие независимого расхождения хромосом при мейозе;
3) случайного сочетания хромосом при скрещивании.
Изменчивость может быть обусловлена не только мутациями, но и
сочетаниями отдельных генов и хромосом, новая комбинация которых при
размножении приводит к изменению определенных признаков и свойств
организма. Такой тип изменчивости называют комбинативной
наследственной изменчивостью. Новые комбинации генов возникают:
• 1) при кроссинговере, во время профазы первого мейотического деления;
• 2) во время независимого расхождения гомологичных хромосом в анафазе
первого мейотического деления;
• 3) во время независимого расхождения дочерних хромосом в анафазе
второго мейотического деления
• 4) при слиянии разных половых клеток.
Сочетание в зиготе рекомбинированных генов может привести к
объединению признаков разных пород и сортов.
Комбинативная изменчивость обеспечивает появление особей с наиболее
благоприятным сочетанием признаков.
43. Соотносительная (коррелятивная) изменчивость
• Возникает в результате свойства генов влиять наформирование не одного, а двух и более признаков
(плейотропное действие генов)
• Обеспечивает постоянство взаимосвязанных
признаков, целостность организма как системы
• Пример:
Длинноногие животные имеют длинную шею. У
столовых сортов свеклы согласованно изменяется
окраска корнеплода, черешков и жилок листа
44. Закон гомологических рядов
В селекции важное значение имеет закон гомологических рядов наследственнойизменчивости, сформулированный советским ученым Н. И. Вавиловым.
Он гласит:
• Внутри разных видов и родов, генетически близких (т. е. имеющих
единое происхождение), наблюдаются сходные ряды наследственной
изменчивости.
Такой характер изменчивости выявлен у многих злаков (рис, пшеница, овес, просо
и др.), у которых сходно варьируют окраска и консистенция зерна,
холодостойкость и иные качества.
Зная характер наследственных изменений у одних сортов, можно предвидеть
сходные изменения у родственных видов и, воздействуя на них мутагенами,
вызывать у них подобные полезные изменения, что значительно облегчает
получение хозяйственно ценных форм.
Известны многие примеры гомологической изменчивости и у человека; например,
альбинизм (дефект синтеза клетками красящего вещества) обнаружен у
европейцев, негров и индейцев; среди млекопитающих -- у грызунов, хищных,
приматов; малорослые темнокожие люди - пигмеи встречаются в тропических
лесах экваториальной Африки, на Филиппинских островах и в джунглях
полуострова Малакки; некоторые наследственные дефекты и уродства,
присущие человеку, отмечены и у животных. Таких животных используют в
качестве модели для изучения аналогичных дефектов у человека. Например,
катаракта глаза бывает у мыши, крысы, собаки, лошади; гемофилия - у мыши и
кошки, диабет - у крысы; врожденная глухота - у морской свинки, мыши, собаки;
заячья губа - у мыши, собаки, свиньи и т. д. Эти наследственные дефекты -убедительное подтверждение закона гомологических рядов наследственной
изменчивости Н. И. Вавилова.
45. Помимо вышерассмотренных различают изменчивость:
индивидуальную и групповую;прерывистую (дискретную) и непрерывную;
качественную и количественную;
независимую изменчивость разных признаков и
коррелятивную (соотносительную);
направленную (определенную, по Ч.Дарвину) и
ненаправленную (неопределенную, по Ч.Дарвину);
адаптивную (приспособительную) и неадаптивную.
При решении общих проблем биологии и особенно эволюции
наиболее существенно подразделение изменчивости, с одной
стороны, на наследственную и ненаследственную, а с другой на индивидуальную и групповую. Все категории изменчивости
могут встречаться в наследственной и ненаследственной,
групповой и индивидуальной изменчивости.
46. Методы генетики
Биохимический – позволяет по продукту (белку) определить
нарушения (мутации) в структуре гена
Кариотипирование – позволяет обнаружить геномные и хромосомные
мутации
Генеологический – позволяет выявить характер наследования
признака (аутосомно-доминантный, аутосомно- рецессивный,
сцепленный с Х-хромосомой доминантный или рецессивный,
сцепленный с У-хромосомой)
Близнецовый –позволяет определить роль наследственности и среды
в формирование фенотипа
Популяционно-статистический – определить соотношение генотипов
в популяции, концентрацию различных аллелей
Гибридологический (предложен Г.Менделем) – позволяет
проанализировать генотипы особей, выявить формы взаимодействия и
взаимовлияния генов и аллелей.
Картирование – позволяет установить структуру хромосом и место
локализации генов
47. Типы наследования признаков
- Аутосомно-доминантный тип наследования:а. При достаточном числе потомков признак
обнаруживается в каждом поколении
б. Редкий признак наследуется примерно
половиной детей
в. Потомки мужского и женского пола
наследуют этот признак одинаково
г. Оба родителя в равной мере передают этот
признак детям
48.
- Аутосомно-рецессивный типнаследования:
а. Признак может передаваться через
поколение даже при достаточном числе
потомков
б. Признак может проявиться у детей в
отсутствие его у родителей. Обнаруживается
тогда в 25% случаев у детей
в. Признак наследуется всеми детьми, если
оба родителя больны
г. Признак в 50% развивается у детей, если
один из родителей болен
д. Потомки мужского и женского пола
наследуют этот признак одинаково
49.
- Наследование сцепленное с Х хромосомой, еслиген, контролирующий проявления признака рецессивный:
а. Мужчины наследуют чаще, чем женщины
б. Наследуют такой признак девочки только от отца
в. В браках, где оба супруга здоровы, могут родиться
дети, имеющие его, при этом он наследуется 50%
сыновей и 100% здоровых дочерей
г. Прослеживается чередование больных мужчин в
поколениях: где их больше, где - меньше
50.
- Наследование сцепленное с Ххромосомой, если ген, контролирующий
проявления признака, - доминантный:
а. Мужчины наследуют реже, чем женщины
б. Если признак только у супруги, то
наследуют его все дети (мать гомозиготная),
или половина детей (мать гетерозиготная)
в. Если только у супруга, то наследуют все
лица женского пола
51.
- Наследование сцепленное с Ухромосомой:
а. Страдают только сыновья,
б. проявляется в каждом поколении,
если отец болен.
52. МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ
В хранении, передаче и преобразовании генетической информациицентральное место занимают нуклеиновые кислоты. Решающим
фактором при этом является способность нуклеиновых, оснований к
специфическому (комплементарному) спариванию.
Биологическая функция нуклеиновых кислот основана главным
образом на свойстве оснований образовывать специфически
(комплементарно) связанные пары.
53. Спаривание оснований в ДНК
Первым свидетельством существования таких структур пocлyжил тот факт, что в
каждом типе ДНК (DNA) содержится примерно одинаковое количество аденина
и тимина. То же самое относится к гуанину и цитозину. Напротив, соотношение
(аденин+тимин) / (гуанин+цитозин) в различных организмах варьирует.
Предложенная в 1953 г. модель структуры ДНК позволила объяснить причину
таких соотношений: интактная ДНК состоит из двух полидезоксинуклеотидных
цепей. Каждое основание одной цепи связано с комплементарным ему
основанием другой цепи водородными мостиками. При этом аденин
комплементарен тимину, гуанин — цитозину. Таким образом, каждая пара
состоит из одного пуринового и одного пиримидинового основания
Комплементарность А и T, соответственно G и С, становится понятной, если
рассмотреть возможные водородные мостики между основаниями. В качестве
доноров (см. с. 14) выступают аминогруппы (аденина, цитозина, гуанина) и NНгруппы гетероциклов (тимина и гуанина). Возможными акцепторами являются
карбонильные группы (тимина, цитозина, гуанина) и атомы азота гетероциклов.
Пара A-T может образовывать два, а пара G-C даже три линейных и поэтому
особенно устойчивых мостика. Урацил, содержащийся в РНК вместо тимина,
ведет себя при спаривании основании подобно тимину.
54. Структура ДНК
Спаривание оснований, проиллюстрированное на схеме А, охватывает в молекуле ДНК
миллионы звеньев. Конечно, это возможно только в том случае, если полярность обеих
цепей различна, т.е. обе цепи имеют противоположные направления. Кроме того, обе цепи
должны быть закручены в виде двойной спирали. Из-за стерических ограничений,
вызванных 2'-ОН-группой остатка рибозы, РНК не мoгyт образовывать структур, подобных
двойной спирали. Поэтому РНК имеют менее регулярную структуру по сравнению с ДНК .
Преобладающая в клетке конформация ДНК (так называемая В-ДНК) представлена
схематически на схеме 1, в виде вандерваальсовой модели. На схеме 1
дезоксирибозофосфатный остов изображен в виде ленты. Основания (здесь указаны в виде
полос) расположены внутри двойной спирали. Следовательно, эта область ДНК
неполярна.
Напротив, внешняя сторона молекулы полярна и заряжена отрицательно за счет углеводных
остатков и фосфатных групп остова. Цепи ДНК на протяжении всего тяжа образуют два
желоба, которые носят названия «малая бороздка» и «большая бороздка».
Так как обе цепи связаны только нековалентными взаимодействиями, двойная спираль при
нагревании или инкубации в щелочном растворе легко распадается на отдельные цепи
(денатурирует). При медленном охлаждении ранее неупорядоченные отдельные цепи
благодаря спариванию оснований вновь образуют двойную спираль (молекула
ренатурирует). Процессы де- и ренатурации играют важную роль в генной инженерии .
В функциональном отношении две цепи ДНК не эквивалентны. Кодирующей цепью
(матричной, смысловой) является та из них, которая считывается в процессе транскрипции .
Именно эта цепь служит матрицей для PHK. Некодирующая цепь (антисмысловая) по
последовательности подобна РНК (при условии замены T на U). Общепринято давать
структуру гена в виде последовательности некодирующей цепи ДНК в направлении 5'→3'.
Если прочитать кодоны в этом направлении, то с помощью генетического кода можно
воспроизвести аминокислотную последовательность белка в принятом порядке, от N- к Сконцу.
55.
56. Молекулярные модели ДНК и тРНК
На схеме представлены вандерваальсовые модели двух небольших нуклеиновыхкислот. На схеме А изображена ДНК (DNA), построенная из 17 пар нуклеотидов,
на схеме Б — структура фенилаланинспецифичной тРНК (tRNA) дрожжей (75
нуклеотидов).
При исследовании синтетических молекул ДНК было показано, что ДНК может
принимать различные конформации. Наиболее часто встречается приведенная
здесь В-форма ДНК. Молекула состоит из двух антипараллельных
полидезоксинуклеотидных цепей, которые закручены в правую двойную
спираль.
Остов этого тяжа образован остатками дезоксирибозы и фосфатными
группировками, соединенными фосфодиэфирными связями. Остов одной
цепи выделен темно-синим цветом, другой — синим, а основания обеих цепей
— голубым. Ароматические кольца отстоят друг от друга на 0,34 нм и почти
перпендикулярны оси спирали. Каждое основание повернуто по отношению к
предыдущему на 35о. На каждый виток двойной спирали (360о) приходится
примерно 10 пар оснований, ход спирали — 3,4 нм. Между остовами двух
отдельных цепей имеются две широкие бороздки. Большая бороздка видна на
модели снизу и сверху, малая бороздка — в центре. ДНК-ассоциированные
белки взаимодействуют с наиболее доступными основаниями в области
большой бороздки. Пространственные соотношения между остовом и
основаниями более наглядно видны на модели одинарной цепи (справа). На
всех моделях аденин окрашен в желтый цвет, а комплементарный ему тимин —
в оранжевый. На виде сверху (в центре; одна цепь выше другой) основания
просматриваются наиболее полно.
57.
58.
Молекулы РНК не могут образовывать двойную спираль и поэтому не
столь высокоупорядочены по сравнению c ДНК. В качестве примера
здесь приведена молекула тРНК из дрожжей. По аналогии с другими
моделями, приведенными на схеме А, основания окрашены в голубой
цвет, остов из остатков рибозы и фосфатных группировок — в темносиний. Связанная на 3'-конце молекула фенилаланина выделена
красным цветом (в исследованной тРНК этот аминокислотный остаток
отсутствовал).
На модели видно, что тРНК свернута в компактную клинообразную
структуру. Как и в ДНК, большинство основании находятся внутри
молекулы, в то время как полярный остов — на поверхности.
Исключение составляют три основания антикодона (окрашены в
желтый цвет), который должен взаимодействовать с мРНК.
Большинство оснований принимает участие в межмолекулярном
спаривании, некоторые из образованных пар не соответствуют общим
принципам спаривания оснований, обычно соблюдаемым в ДНК (A + U,
G + С).
Рибонуклеиновые кислоты [PHK(RNA)] представляют собой полимеры
из нуклеозидфосфатных звеньев, соединенных фосфодиэфирной
связью. В качестве азотистых оснований в РНК присутствуют урацил,
цитозин, аденин и тимин. В РНК можно также встретить множество
необычных и модифицированных азотистых оснований.
59. Рибонуклеиновые кислоты
РНК принимают участие во всех стадиях процесса генной экспрессии и биосинтеза белка. Свойстванаиболее важных видов РНК приведены в таблице. Кроме того, здесь схематически показаны
вторичные структуры молекул РНК.
В отличие от ДНК, РНК не образуют двойных спиралей, но содержат короткие участки со спаренными
основаниями. Это приводит к образованию субструктур, которые при двумерном изображении
напоминают «шпильки» и петли, образующие фигуру типа «кленового листа». В таких структурах
двухцепочечные участки соединены петлями. Множество фрагментов, в которых чередуются
структуры типа шпилька—петля, содержится в высокомолекулярных РНК, таких, например, как
рибосомная 16S-рРНК (16S-rRNA) (в центре). Кроме того, эти фрагменты образуют трехмерные
структуры; следовательно, РНК подобно белкам имеют четвертичную структуру. До настоящего
времени установлена четвертичная структура небольших PHK, прежде всего тРНК (tRNA). Из
иллюстраций, приведенных на схеме Б очевидно, что трехмерная укладка структуры типа
«кленовый лист» окончательно не установлена.
PHK клетки существенно различаются по размерам, строению и продолжительности существования.
Преобладающую часть представляют рибосомные РНК [рРНК (rRNA)], которые в различных
формах составляют структурный и функциональные части рибосом . Рибосомные РНК
синтезируются в ядре в процессе транскрипции на ДНК, там же подвергаются процессингу и
ассоциируют с рибосомными белками, образуя рибосому . Приведенная на схеме А бактериальная
16S-рРНК, включающая 1542 нуклеотида, является компонентом малой рибосомной субчастицы, в
то время как небольшая 5S-рРНК (из 120 нуклеотидов) входит в состав большой субчастицы.
Матричная РНК [мРНК (mRNA)] переносит генетическую информацию из клеточного ядра в цитоплазму.
Ее транскрипты также сильно модифицируются в ядре (созревание мPHK) Так как мРНК
считывается на рибосоме кодон за кодоном она не должна складываться в стабильную третичную
структуру. Спариванию оснований препятствуют белки, ассоциированные с мРНК. Из-за различного
объема информации, которую могут нести мРНК, РНК этого типа сильно варьируют по размерам.
Для мРНК характерно короткое время жизни, так как они быстро распадаются после трансляции. В
сплайсинге предшественников мРНК принимают участие малые ядерные РНК [мяРНК (snRNA, от
англ. small nuclear RNA)]. Они ассоциированы c рядом белков, образуя «сплайсомы».
60. Транспортные РНК ( tRNAPhe )
Транспортные РНК [тРНК (tRNA)] участвуют в процессе трансляции в
качестве промежуточного связующего звена между нуклеиновыми
кислотами и белками. Это небольшие молекулы РНК из 70-90
нуклеотидов, которые с помощью своих антикодонов "узнают" за счет
спаривания оснований определенные кодоны на мРНК. На 3'-конце
(ССА-3') они несут ту аминокислоту, которая согласно генетическому
коду соответствует очередному кодону мРНК.
Последовательность оснований и третичная структура
фенилаланинспецифичной тРНК (tRNAPhe) из дрожжей являются
типичными для всех тРНК. В молекуле этой тРНК содержится
довольно много минорных и модифицированных оснований (1,
выделены темно-зеленым цветом). К ним относятся псевдоуридин (ψ),
дигидроуридин (D), тимидин (T), встречающийся обычно в ДНК, а
также множество метилированных нуклеотидов, таких, например, как 7метилгуанидин (m7G) и входящий в состав антикодона 2'-Ометилгуанидин (m2G). Конформацию молекулы стабилизируют
многочисленные пары оснований, часть из которых не соответствуют
общим принципам спаривания
61.
62. Репликация
Для передачи дочерним клеткам генетической информации в процессе
репликации ДНК (DNA) должна быть создана копия генома. Репликация
ДНК осуществляется ДНК-зависимыми ДНК-полимеразами. Эти
ферменты используют в качестве шаблона одну из цепей двойной
спирали ДНК, так называемую матрицу. На матрице, начиная с
короткой стартовой последовательности (праймера), ферменты
синтезируют комплементарную цепь и воспроизводят в итоге исходную
двухтяжевую ДНК. Субстратами ДНК-полимераз являются четыре
дезоксирибонуклеотидтрифосфата: аденозин-, гуанозин-, тимидин- и
цитозинтрифосфаты. При каждом шаге синтеза ДНК происходит
спаривание нуклеотида с соответствующим азотистым основанием
матричной цепи. Затем α-фосфатная группа связанного нуклеотида
подвергается нуклеофильной атаке со стороны 3'-ОН-группы
предыдущего нуклеотида. За этим следует удаление дифосфата и
образование новой фосфодиэфирной связи. Эти этапы повторяются
снова и снова по мере движения ДНК-полимеразы от одного основания
к следующему вдоль матрицы. В соответствии с этим механизмом
матричная цепь ДНК считывается в направлении 3'→5'.
В большинстве клеток имеется несколько ДНК-полимераз. Наряду с
ферментами, которые осуществляют собственно репликацию,
существуют полимеразы, которые включены в процессы репарации
ДНК или реплицируют митохондриальную ДНК эукариот. Большинство
ДНК-полимераз построены из множества субъединиц, роль которых до
конца не выяснена.
63.
64.
В настоящее время процесс репликации у прокариот достаточно изучен, в то время как
многие аспекты эукариотической репликации остаются неясными. Однако с большой долей
вероятности можно утверждать, что в большинстве клеток этот процесс протекает в
основном одинаково. На схеме показана простейшая схема репликации у бактерии
Escherichia coli. В бактериях репликация начинается со специфической точки в кольцевой
ДНК (область начала репликации) и продолжается в обоих направлениях. В результате
образуются две репликативные вилки, которые продвигаются в противоположных
направлениях, т. е. обе цепи реплицируются одновременно. На схеме исходная ДНК (1)
окрашена в голубой и фиолетовый цвета, а вновь синтезирующаяся — в розовый и
оранжевый. В функционировании каждой вилки принимают участие множество различных
белков, из которых здесь указаны наиболее важные. Каждая репликативная вилка (2)
включает по крайней мере две молекулы ДНК-полимеразы III, ассоциированные с
несколькими вспомогательными белками.
Поскольку матричная цепь всегда читается в направлении 3'→5', только одна из цепей
может считываться непрерывно (розовая/фиолетовая; 2). Другая цепь (голубого цвета)
считывается в направлении, противоположном движению репликативной вилки. В
результате на матрице вначале синтезируются короткие фрагменты новой цепи ДНК
(зеленый/оранжевый), так называемые фрагменты Оказаки (OF), названные так по имени
их первооткрывателя. Каждый фрагмент начинается с короткой РНК-затравки (праймера,
зеленого цвета), необходимой для функционирования ДНК-полимеразы. Праймер
синтезируется специальной РНК-полимеразой («праймаза», на схеме не показана), ДНКполимераза III достраивает этот праймер до фрагмента ДНК длиной 1000-2000
дезоксинуклеотидных звеньев (оранжевого цвета). Синтез этого фрагмента далее
прерывается, и новый синтез начинается со следующего РНК-праймера. Индивидуальные
фрагменты Оказаки первоначально не связаны друг с другом и все еще имеют РНК на 5'концах (3). На некотором расстоянии от репликативной вилки ДНК-полимераза I начинает
замещать РНК-праймер последовательностью ДНК. В завершение остающиеся
одноцепочечные разрывы репарируются ДНК-лигазой. В образованной таким образом
двойной спирали ДНК только одна из цепей синтезирована заново. Поэтому говорят, что
репликация ДНК происходит по полуконсервативному механизму
65. Транскрипция
Для того чтобы хранящаяся в ДНК информация могла быть
использована, ее необходимо переписать (транскрибировать) в
последовательность РНК. При этом ДНК служит только матрицей, т. е.
она не меняется в процессе транскрипции. Транскрибируемые
последовательности ДНК, т. е. участки ДНК, которые кодируют
определенные белки, называются генами. Установлено, что геном
млекопитающих содержит по крайней мере 50000 индивидуальных
генов, которые вместе составляют менее 20% суммарной ДНК генома.
Функция «избыточных» последовательностей ДНК до конца не
установлена.
Транскрипция осуществляется ДНК-зависимыми РНК-полимеразами.
Они действуют подобно ДНК-полимеразам, за исключением того, что
включают во вновь синтезируемую цепь РНК (RNA) рибонуклеотиды
вместо дезоксирибонуклеотидов и не нуждаются в праймерах.
Эукариотические клетки обычно содержат по крайней мере три
различных типа РНК-полимераз, РНК-полимераза I катализирует
синтез РНК с коэффициентом седиментации 45S, которая служит
предшественником трех различных рибосомных РНК. РНК-полимеразы
II синтезируют гяРНК (hnRNA), которые служат предшественниками
мРНК (mRNA) и мяРНК (snRNA). Наконец, РНК-полимераза III
транскрибирует гены, которые кодируют тРНК (tRNA), 5S- рРНК (rRNA)
и некоторые мяРНК. Эти РНК служат предшественниками
функциональных РНК, которые образуются в процессе созревания
РНК .
66. Структура β-глобинового гена
• В качестве примера организации небольшого эукариотическогогена представлена схема гена, кодирующего β-цепь
гемоглобина (146 аминокислот). Этот ген состоит из более чем
2000 п.о. (2 тыс. п.о.). Однако из них только около 450 п.о. из них
несут информацию об аминокислотной последовательности βглобина. Кроме трех кодирующих участков (экзонов), ген
включает два некодирующих (интроны, I1 и I2). На 5'-конце гена
располагается промоторный участок (розовый цвет) длиной
приблизительно 200 п.о., который имеет регуляторную функцию.
Транскрипция начинается с 3'-конца промоторного участка и
продолжается, пока не достигнет сайта полиаденилирования.
Образующийся первичный транскрипт (гяРНК) β-глобинового
гена состоит из ~1600 п.о. Во время созревания РНК
некодирующие последовательности, соответствующие
интронам, удаляются и, кроме того, оба конца гяРНК
модифицируются. Зрелая β-глобиновая мРНК включает в себе
~40% гяРНК и дополнительно 3'-концевую последовательность
из 100-200 АМФ (AMP).
• У многих генов разделение на экзоны и интроны еще более
выражено. Так, общая длина гена дигидрофолатредуктазы
составляет выше 30 тыс.п.о. Информация об аминокислотной
последовательности распределена по 6 экзонам, которые в
сумме составляют только ~6 тыс.п.о.
67.
68.
• Как уже упоминалось, РНК-полимераза II связывается с 3'концом промоторного участка. Последовательность,обеспечивающая это связывание, так называемый ТАТА-бокс,
короткий А- и Т-обогащенный участок, последовательность
которого слегка варьирует у разных генов. Типичная
последовательность (каноническая) — ...ТАТААА... . Для
взаимодействия полимеразы с этим участком необходимы
несколько белков, основных факторов транскрипции.
Дополнительные факторы могут либо стимулировать, либо
ингибировать этот процесс (контроль транскрипции) .
• После инициации синтеза (2), РНК-полимераза движется в
направлении 3'→5' матричной цепи. В процессе инициации
фермент разделяет короткий участок двойной спирали ДНК на
две отдельные цепочки. Нуклеозидтрифосфаты связываются
комплементарно на кодирующей цепочке ДНК водородными
связями и шаг за шагом присоединяются к растущей молекуле
РНК (3). Вскоре после начала элонгации 5'-конец транскрипта
защищается «кэпом» (от англ. cap). Как только транскрипция
доходит до сайта полиаденилирования (обычно это
последовательность ...ААТААА...), транскрипт отщепляется (4).
После этого полимераза прекращает транскрипцию и
диссоциирует от ДНК.
69. Созревание РНК
Большинство меток организма содержит полный набор генов, но обычно из этого набора
используется крайне незначительный объем информации. Постоянно транскрибируются
только те гены, которые кодируют структурные белки и ферменты промежуточного
метаболизма. Кроме этих постоянно необходимых генов имеется много других генов,
активных только в определенных типах клеток, при определенных метаболических условиях
или во время дифференцировки.
Контроль на уровне транскрипции
Порядок транскрибирования генов определяется регуляторной системой, которая носит
название системы регуляции транскрипции. Контроль транскрипции осуществляется
структурами двух типов. Большинство генов содержат в своем промоторном участке
несколько коротких сегментов ДНК (DNA) (регуляторные элементы, цис-действующие
элементы), с которыми могут связываться факторы транскрипции. Регуляторные элементы,
стимулирующие транскрипцию связанных с ними генов, называются энхансерами
(усилителями, от англ. enhancer). Белки, подавляющие транскрипцию. — сайленсерами
(успокоителями, от англ. silencer). Факторы транскрипции — это белки, т. е. продукты
других, независимых генов. Поэтому их называют опосредованно действующими
факторами. Для процесса транскрипции генов требуются не только РНК-полимераза, но и
другие белки, называемые основными факторами транскрипции. Установлено, что у
эукариот таким фактором является ТАТА-связывающий белок (ТСБ, англ. ТАТА-Вох
Binding Protein, TBP), который взаимодействует с основным регуляторным элементом.
ТАТА-боксом, присутствующим в большинстве генов. С этим комплексом затем
связываются другие основные факторы транскрипции и РНК-полимеразы. Дополнительные
факторы могут влиять на инициацию транскрипции, связываясь с другими регуляторными
элементами. Отсюда они взаимодействуют с основным транскрипционным комплексом,
либо активируя, либо ингибируя его. Такие факторы активируют, например, комплексы
стероидных гормонов с рецепторами. По завершении транскрипции из гяРНК вырезаются
интроны, содержащие некодирующие последовательности- процесс процессинга
70. Сплайсинг
Сплайсинг РНК катализируется комплексами белков с РНК, известными как
«малые ядерные рибонуклеопротеидные частицы» (мяРНП,). Интроны,
входящие в гяРНК (hnRNA), имеют специфические последовательности на 3'- и
5'-концах (а). На первой стадии сплайсинга ОН-группа аденозилового остатка,
расположенного в интроне, атакует (при участии мяРНП) и расщепляет
фосфодиэфирную связь на 5'-конце интрона (б). Одновременно в интроне
образуется новая связь, которая придает ему форму петли (в). На второй
стадии терминальная ОН-группа 5'-концевого интрона атакует связь в 3'-конце
интрона. В результате оба экзона соединяются, а интрон освобождается (г).
Во время сплайсинга комплексы из гяРНК и мяРНП образуют сплайсому.
Полагают, что мяРНК в сплайсоме образуют канонические пары друг с другом и
с гяРНК и таким образом фиксируют и ориентируют их реакционные группы.
Собственно катализ обусловлен РНК-составляющей сплайсомы Такие
каталитические РНК носят название рибозимов.
В. 5'- и 3'-Концевые модификации мРНК
У эукариот после завершения собственно транскрипции 5'-конец растущей
молекулы РНК блокируется структурой, которая называется кэп (от англ. cap). В
случае мРНК кэп состоит из 7'-метил-ГТФ и защищает РНК от гидролиза 5'экзонуклеазами. В конце транскрипции к 3'-концу присоединяется
полиадениловая последовательность, которая может включать до 200
звеньев АМФ (AMP). Только после этого созревшая мРНК (mRNA) покидает
ядро.
71.
72. Генетический код, активация аминокислот
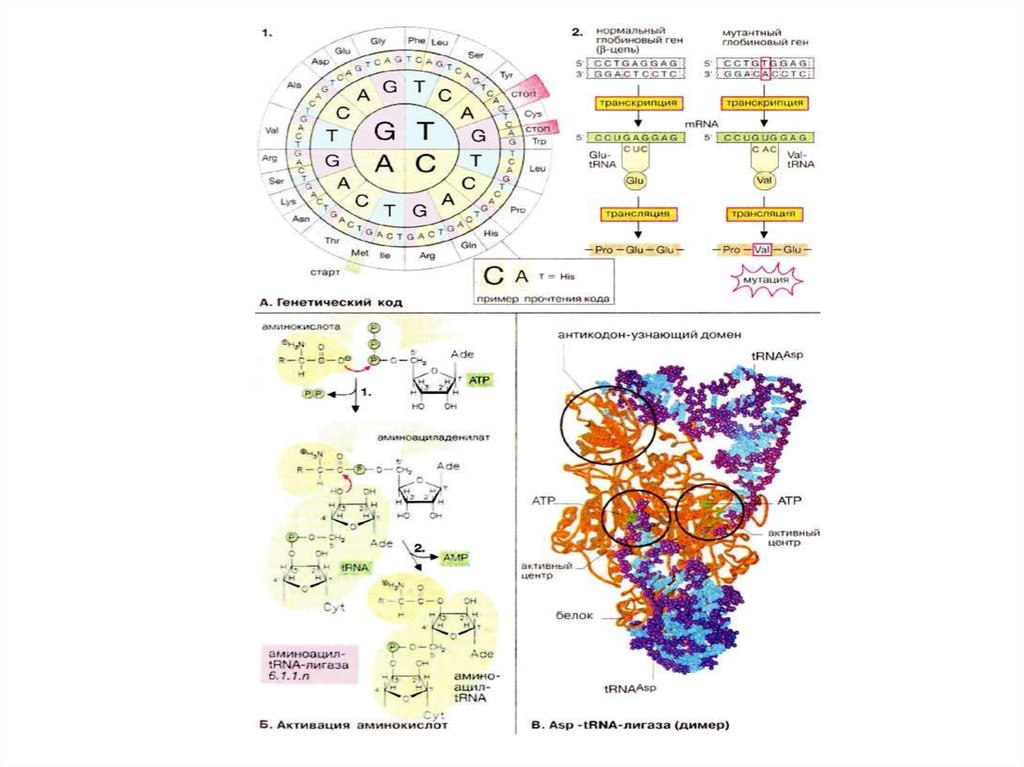
Большая часть генетической информации, содержащейся в ДНК, кодирует
последовательность аминокислот. Процесс экспрессии генетической
информации включает транскрипцию «текста», записанного на «языке
нуклеиновой кислоты», в текст, записанный на «языке белков». Таково
происхождение термина трансляция (дословно — перевод), используемого для
обозначения процесса биосинтеза белков. Правила, которым следует
трансляция, называют генетическим кодом.
Поскольку в биосинтезе участвуют 20 аминокислот, называемых
протеиногенными, «язык» нуклеиновых кислот должен содержать по крайней
мере 20 слов (кодонов) Однако в аминокислотном «алфавите» имеется только
четыре «буквы» (А, Г, Ц и У или Т [или в англ. транскрипции: A, G, С и U или Т*]),
так что для получения 20 различных слов каждое должно состоять по крайней
мере из трех букв. Кодоны действительно включают три азотистых основания
(триплет нуклеотидов). На схеме 1 представлен стандартный код ДНК
(последовательность триплетов в некодирующей цепи), изображенный в
виде круга. Схема читается от центра наружу, так что, например, триплет CAT
кодирует аминокислоту гистидин. ДНК-кодоны идентичны таковым в мРНК
(mRNA), за исключением того, что в мРНК вместо урацила (U), характерного для
ДНК, стоит тимин (Т).
В триплетном генетическом коде для 20 аминокислот потенциально существует
43 = 64 кодона. Таким образом, большинство аминокислот записывается
несколькими кодонами, т. е. генетический код является вырожденным. Кроме
того, имеются три триплета, которые обозначают конец транскрипции (стопкодоны). Еще один специальный кодон, стартовый (инициирующий) кодон,
маркирует начало трансляции. Генетический код, показанный на рисунке,
является почти универсальным. Этому стандарту не полностью соответствуют
только митохондрии и некоторые микроорганизмы.
73.
74. Активация аминокислот
Для каждой из 20 аминокислот имеется соответствующая аминоацил-тРНК-лигаза,которая в цитоплазме соединяет аминокислоту с тPHK(tRNA) (см. с. 88). Этот
процесс активации аминокислот осуществляется в две стадии. Сначала
аминокислота связывается с ферментом и реагирует с АТФ (АТР), образуя
макроэргический смешанный ангидрид — аминоациладенилат. Затем
аминоацильный остаток переносится на концевую 3'-ОН-группу концевого
остатка рибозы тРНК (другой группой лигаз аминоацил переносится на 2'-ОНгруппу). В аминоацил-тРНК карбоксильная группа аминокислотного остатка
этерифицируется остатком рибозы 3'-концевого остатка аденозина, входящего в
последовательность ...ССА-3'.
• Точность трансляции зависит, прежде всего, от субстратной специфичности
аминоацил-тРНК-лигаз. Корректирующий механизм активного центра лигазы
обеспечивает немедленное удаление ошибочно присоединенных
аминокислотных остатков. В среднем встречается только одна ошибка на 1300
аминокислотных остатков — поразительно высокая точность «работы», если
представить, насколько близки структуры некоторых аминокислот.
• Процесс активации аминокислот представлен на примере лигазы, специфичной
для аспарагиновой кислоты. Молекулы фермента (окрашены в оранжевый цвет)
связаны между собой в димер, причем каждая субъединица ассоциирована с
одной молекулой тРНК (окрашены в голубой цвет). В активном центре
присутствует остаток АТФ (окрашен в зеленый цвет), связанный с 3'-концом
тРНК Другой домен белка (слева вверху) отвечает за «узнавание» антикодона
тРНК.
75. Характеристика генетического кода
Принцип кодировки1.
2.
3.
4.
5.
6.
Триплетность
Вырожденность
(упорядоченность,
экономность)
Однозначность
Надежность
Видоспецифичность
Линейность
Принцип чтения
1.
2.
3.
4.
5.
6.
Однонаправленность
Непрерывность
Неперекрываемость
Универсальность
(относительная)
Смысловые и
бессмысленные триплеты
Коллениарность
76. Классификация генов у эукариот
1. Акцепторные – регулируют деятельность структурных генов
2. Структурные – несут информацию о первичной структуре белка.
Подразделяются на 2 категории:
1) по структуре организации:
а) уникальные гены. Представлены в единственном числе и не
имеют копий. Если в них происходят мутации. То они сразу
проявляются фенотипически
б) умеренно повторяющиеся, имеет до 70 копий, кодируют синтез
специфических белков (тубулина, миозина)
в) высокоповторяющиеся более 70 копий, входит в состав
сателлитной ДНК.
2) псевдогены – в ходе мутаций утратили свою информативность,
но так как они нейтральны, то не исчезают.
3) спейсеры –это гены связывающие акцепторные и структурные
гены между собой.
77. Классификация генов у эукариот
• 1) домашнего хозяйства• 2) роскошного синтеза
• 3) гамиозисные гены –активируются в
период эмбриогенеза
• 4) гены стресса – в норме неактивны,
функционируютв экстремальной
ситуации
• 5) прото-онкогены – в каждой клетке до
70 онкогенов