










")






")











































")


































")






 генотипов потомков при панмиксии")










































")















")











 medicine
medicine biology
biologySimilar presentations:






")



Формальная генетика в медицине
1. Формальная генетика в медицине
кафедра медицинской генетикиСЗГМУ им. И.И. Мечникова
2.
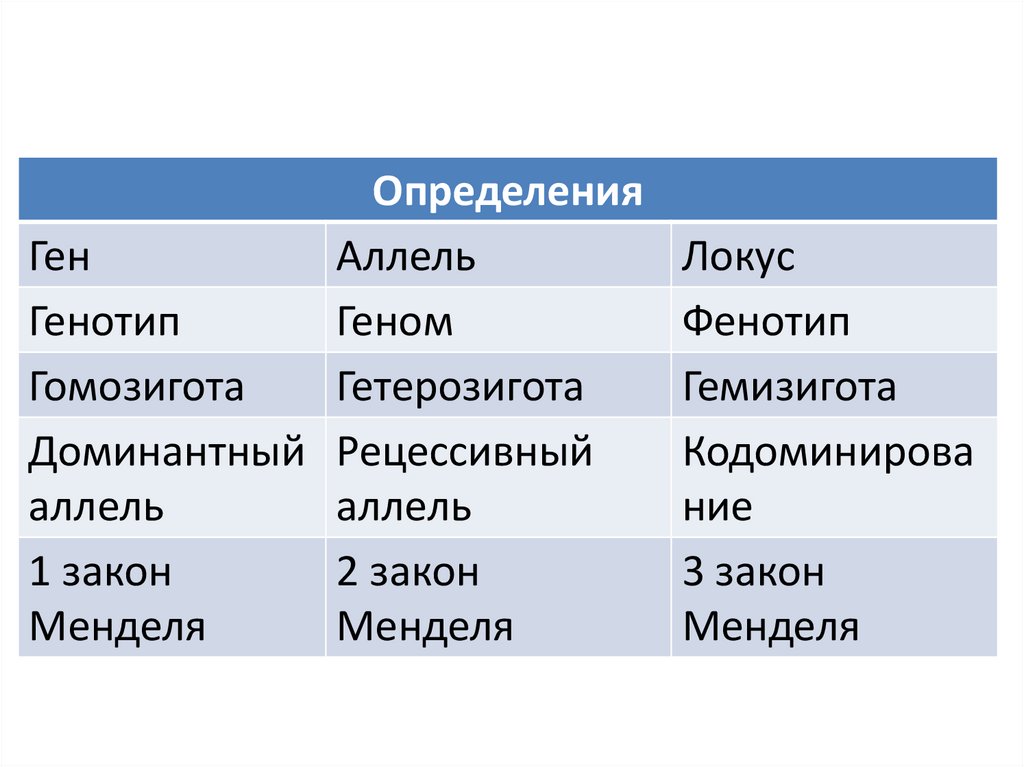
ОпределенияГен
Аллель
Генотип
Геном
Гомозигота
Гетерозигота
Доминантный Рецессивный
аллель
аллель
1 закон
2 закон
Менделя
Менделя
Локус
Фенотип
Гемизигота
Кодоминирова
ние
3 закон
Менделя
3.
Часть 1.История.
Общий обзор нуклеиновых кислот.
4. Изучение генетики на организменном уровне.
В 1865 году чешский ученыйЯн Грегор Мендель
•Выявил «факторы» (законы)
наследственности
•Заложил основы дискретности –
«дискретные задатки» –
наследственного вещества и его
распределения в половых клетках.
В 1866 году опубликовал доклад
«Опыты над растительными
гибридами».
5. Открытие ДНК
Иоганн Фридрих МишерВ 1869 году работал в университете
города Тюбинген (Германия).
Он работал с гноем больных, выделяя
из него лейкоциты и проводя
экстракцию ядер щелочными
растворами, затем добавлял уксусную
кислоту и получал преципитат (ДНК).
Вначале новое вещество получило
название нуклеин, а позже, когда
Мишер определил, что это вещество
обладает кислотными свойствами,
вещество получило название
нуклеиновая кислота.
6. Изучение генетики на клеточном уровне.
Теодор Бовери, немецкий цитолог иэмбриолог.
• В 1887 году выдвинул теорию
индивидуальности хромосом,
• установил роль центросомы как
кинетического центра клетки,
• заложил основы учения об «органообразующих веществах»,
• описал различие между хроматином
половых и соматических клеток.
7.
Изучение генетики на клеточномуровне.
• Гуго де Фриз (Голландия)
• Карл Эрих Корренс (Германия)
• Эрих Чермак (Австрия)
В 1900 году, повторно открыли
законы наследования
признаков (законы Менделя),
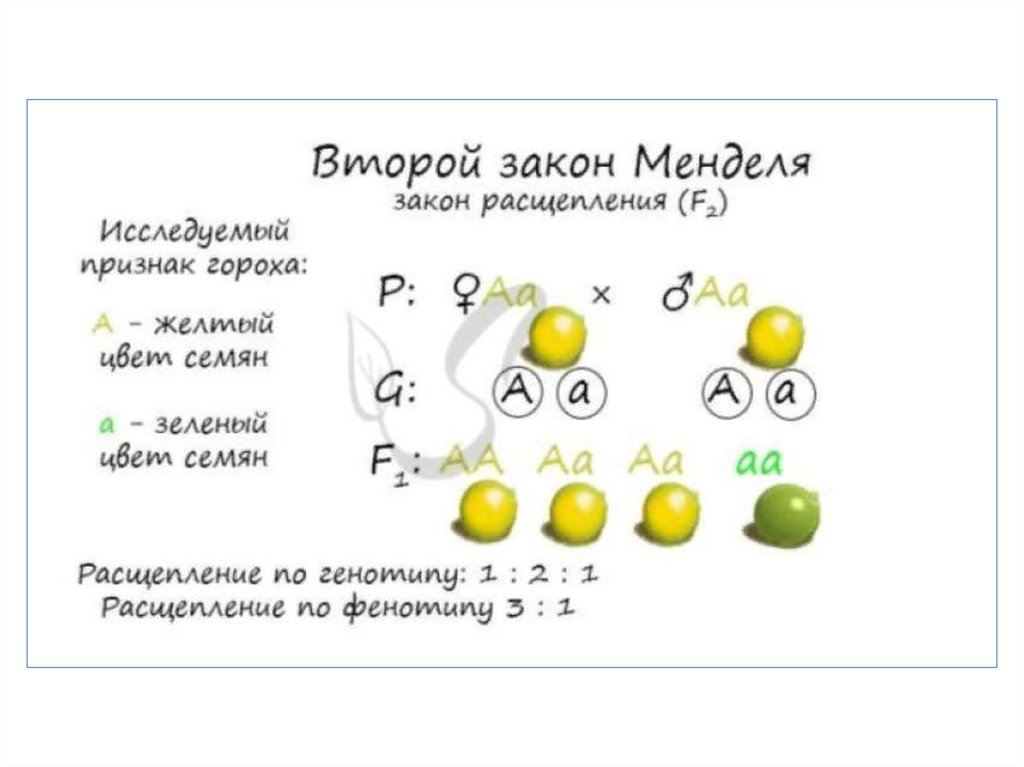
что дало мощный толчок к
развитию цитогенетики.
8.
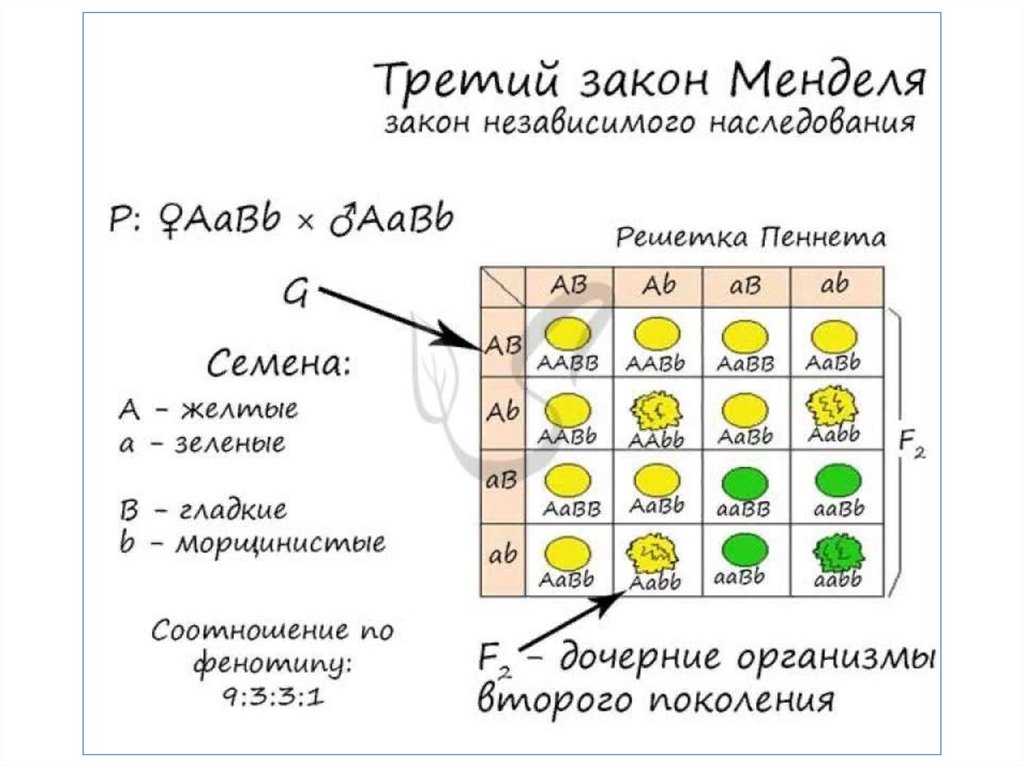
Изучение генетики на клеточномуровне.
Вильгельм Людвиг Иогансен
датский биолог.
• Создал учение о чистых линиях,
• заложил основы принципов
селекции.
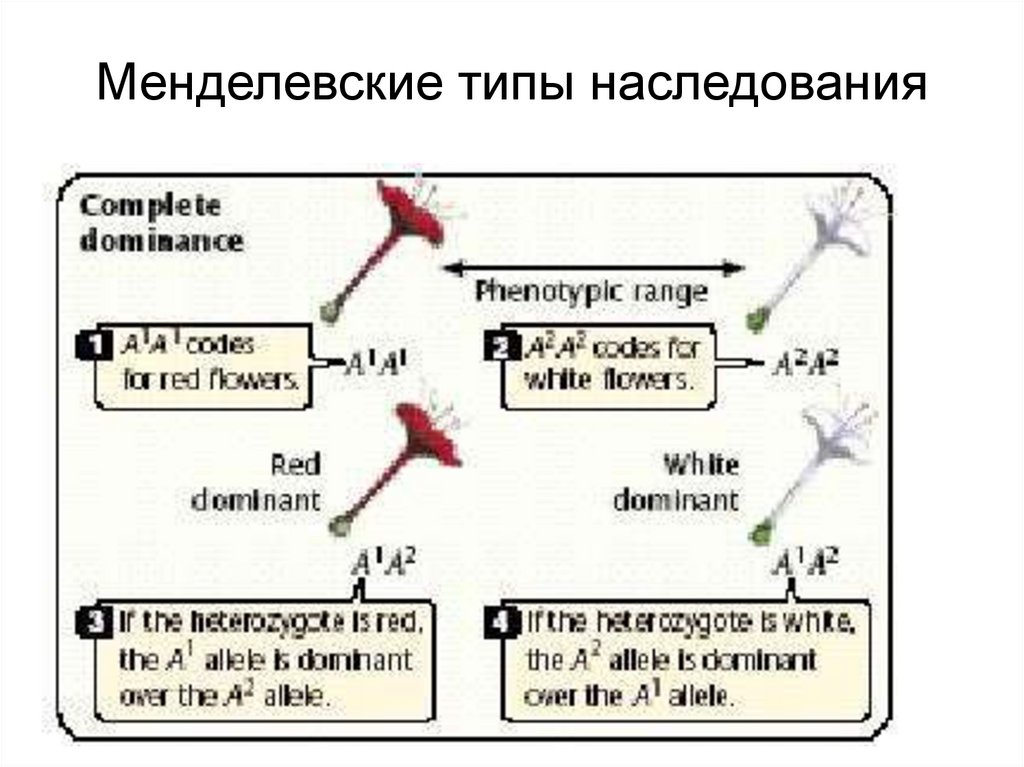
Ввел (1909) термины: «ген»,
«генотип» и «фенотип».
9.
Изучение генетики на клеточномуровне.
Томас Морган.
•Ген – единица наследственной
информации (линейно расположен в
хромосоме)
•Гены в пределах одной хромосомы
наследуются совместно (сцеплено)
•Сцепление генов может нарушаться в
результате кроссинговера
•Гены, на негомологичных хромосомах,
наследуются независимо друг от друга и
образуют различные комбинации
1910 г
10.
Изучение генетики на молекулярномуровне.
Получение первых индуцированных
мутаций
Надсон Г.А. (1925 г., СССР) на грибах
Г. Меллер (1927 г, США) на дрозофиле
И.Л. Стадлер (1928г) на кукурузе
11.
Изучение генетики на молекулярномуровне.
•Н.К. Кольцов в 1928 г высказывает
мысль о связи генов с определенным
химическим веществом (белком???).
•С.С.Четвериков основоположник
генетики популяций, концепции
«мутационного давления», создал
понятие «генотипическая среда».
• А.С.Серебровский ввел термин
«генофонд», открыл «эффект
положения» генов.
12.
Изучение генетики на молекулярномуровне.
Барбара Мак-Клинток американский генетик.
В 1940 году открыла
существование «прыгающих»
генов. Транспозиция генов
сопровождалась изменением
свойств «гена-соседа».
Последующая транспозиция
прежнее место приводила к
восстановлению функции
мутировавшего гена.
13. Эрвин Чаргафф (1)
1950-1952 гг – первое правило(для двойной цепи ДНК)
Erwin Chargaff
Т=А, G=C
14. Начало 50-х годов
Maurice H F WilkinsRosalind Franklin
15.
James Dewey WatsonFrancis Crick
16.
17.
18.
Номинация на премию19.
Становление молекулярно-генетической диагностики• Роберт Холли, Хар Гобинд Хорана, Хайнрих Маттеи, Маршалл
Ниренберг раскрыли генетический код (1961–1966)
• Генетический код — свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности
нуклеотидов.
20. Эрвин Чаргафф (2)
1967г – второе правило(для одной цепи ДНК)
Erwin Chargaff
Т≈А, G≈C
≥70-100 т.п.о.
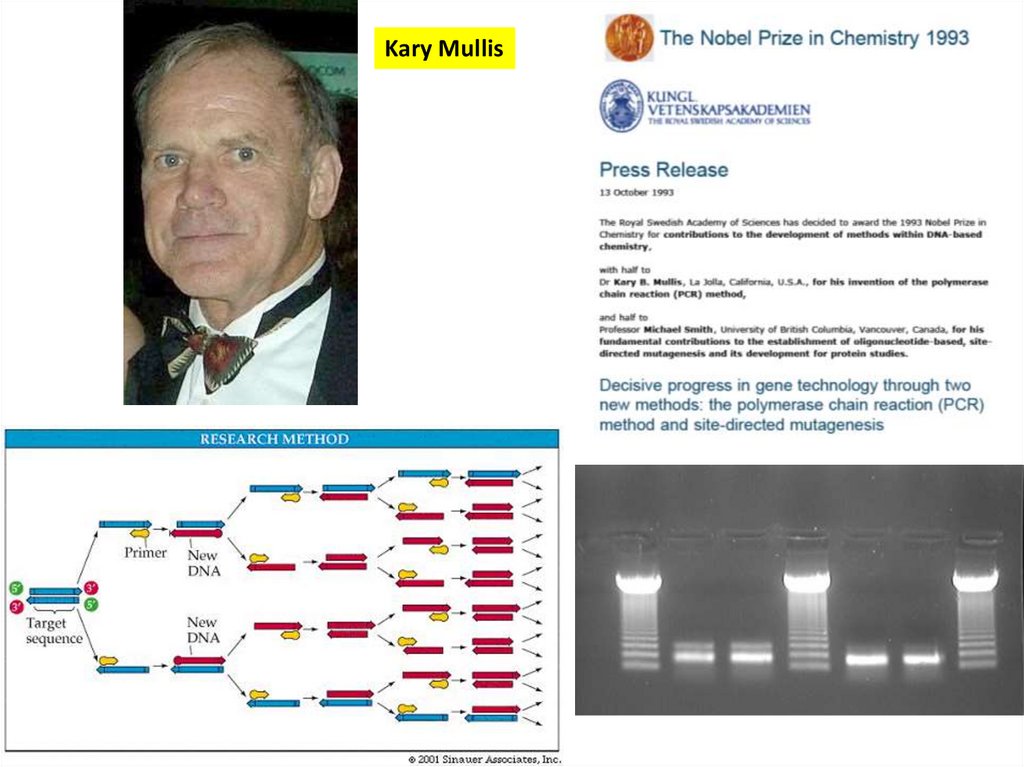
21.
Kary Mullis22. Ген – мишень для исследований
Дж. БидлД. Ледерберг
«Один ген – один фермент»
(начало 40-х годов)
В. Иогансен
1909 ген
Промотер
Э. Тейтем
Экзон 1
ГЕН
Интрон 1
Экзон 2
Терм
23. Структура генома человека
?24. Структура генома человека
Jiri Sana, 201225. Концепция «мусорной» ДНК 70-х
Ohno Susumo, 197226. Ген – мишень для исследований
«Прерывистость гена»(начало 80-х годов)
Ф. Шарп
Промотер
Р. Робертс
Экзон 1
Интрон 1
Экзон 2
Терм
27. Проект «Геном человека» 1990-2000
Цели проекта:1. Идентификация 20 000–25 000 генов
ДНК;
2. Определение последовательности 3
млрд. пар химических оснований,
составляющих ДНК человека, и
сохранение этой информации в базе
данных;
3. Усовершенствование приборов для
анализа данных;
4. Внедрение новейших технологий в
область частного использования;
5. Исследование этических, правовых и
социальных вопросов, возникающих при
расшифровке генома.
28. Проект «Геном человека» 1990-2000
В результате исполнения проекта «Геномчеловека» был:
1. Создан открытый банк генокода.
2. Проведен анализ сходства в
последовательностях ДНК различных
организмов.
3. Проведено межвидовое сравнение.
4. Подтверждено, что 98,5% ДНК не кодирует
белки, но эта часть отвечает за ее
функционирование.
5. Ген — это единица наследственности,
которую нельзя понимать как просто участок
ДНК, кодирующий белки.
6. Лишь около 60 генов претерпевают
спонтанную мутацию каждое поколение. При
этом мутировавшие гены могут вызвать
различные заболевания.
7. Появилась возможность разработки первых
генетические лекарства для блокирования
эффекта генетических отклонений
29.
30.
31.
1х1014м = 1х1011кмДлина
Кол-во раз
Земля
40 075 км
2 500 000
Земля – Луна
384 400 км
260 000
149 600 000 км
700
Земля – Солнце
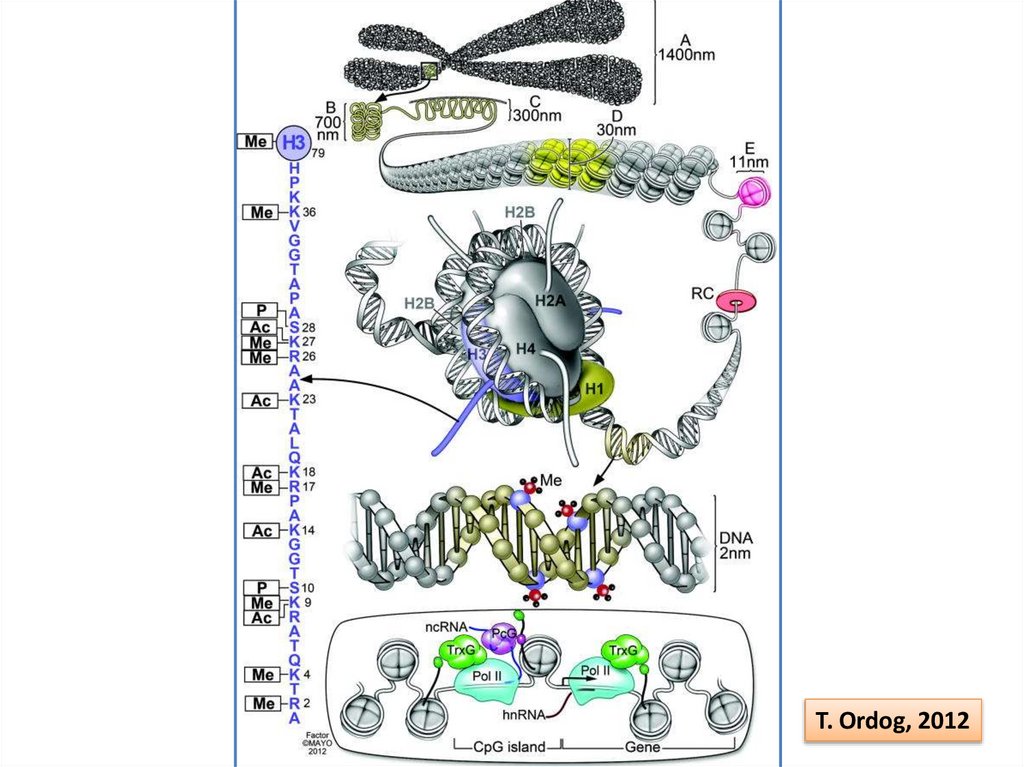
32.
T. Ordog, 201233. Структура генома человека
34.
Часть 2.Строение нуклеиновых кислот.
35. Терминология
ДНК – Дезоксирибонуклеиновая кислотаРНК – Рибонуклеиновая кислота
Транскрипция – ферментативный синтез РНК на ДНК матрице
Трансляция – ферментативная полимеризация аминокислот в полипептидную цепь
36. Основная догма молекулярной генетики
ДНКТранскрипция
мРНК
рРНК
Рибосомы
Трансляция
Полипептид
тРНК
37. Строение азотистых оснований
Пиримидиновое кольцоЦитозин
Пуриновое кольцо
Гуанин
Урацил
Тимин
Аденин
38. Строение моносахаридов
Рибоза2-дезоксирибоза
39. Нуклеозид и нуклеотид
НуклеозидУридин
Нуклеотид
Дезоксиадениловая кислота
40. Номенклатура НК.
Азотистыеоснования
Цитозин
Нуклеозиды
Нуклеотиды
Цитидин
(Дезокси)Цитидиловая кислота
Урацил
Уридин
(Дезокси)Уридиловая кислота
Тимин
Тимидин
(Дезокси)Тимидиловая кислота
Гуанин
Гуанозин
(Дезокси)Гуаниловая кислота
Аденин
Аденозин
(Дезокси)Адениловая кислота
41. Фосфатные группы
ДезоксинуклеозиддифосфатДезокситимидиндифосфат
Нуклеозидтрифосфат
Аденозинтрифосфат
42. Фосфодиэфирная связь – основа одноцепочечной структуры
3’ – 5’Фосфорнодиэфирная
связь
43. Фосфодиэфирная связь – основа одноцепочечной структуры
44. Комплиментарность
45. Двуспиральная структура ДНК
Диаметр 2АОдин
полный
виток
34 А
Большая
бороздка
22 А
Малая
бороздка
12 А
Сахарофосфатный
остов
Пары
азотистых
оснований
Центральная ось молекулы
46.
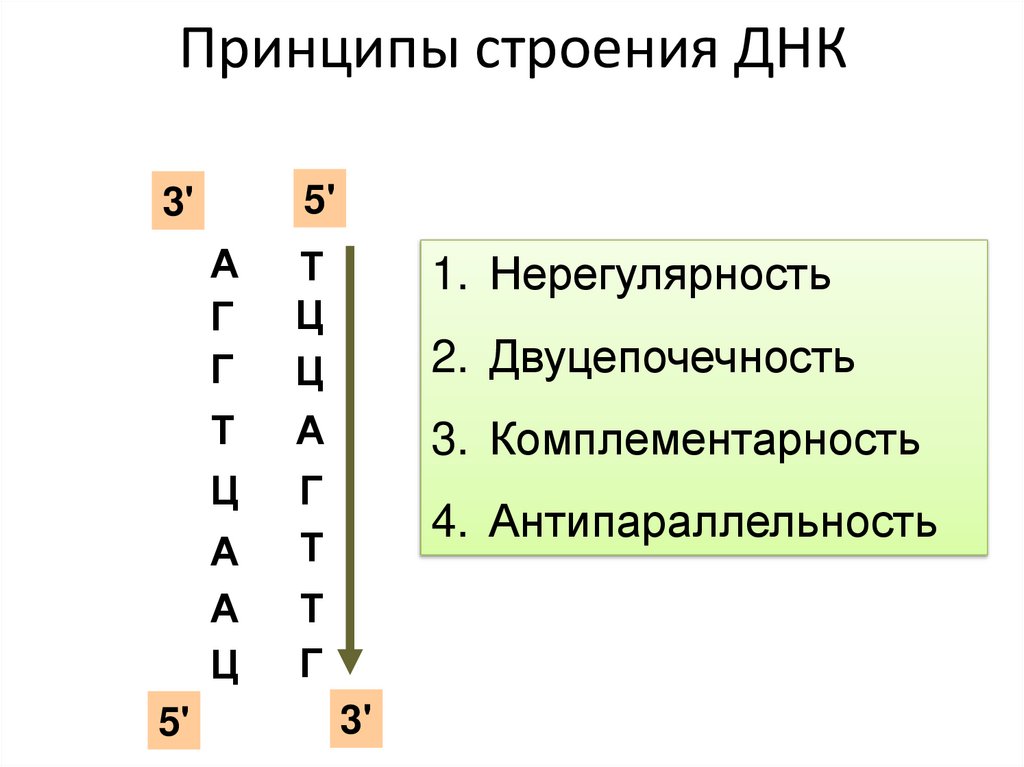
Принципы строения ДНК5'
3'
А
Г
Г
Т
Ц
А
А
Ц
5'
Т
Ц
Ц
А
1. Нерегулярность
2. Двуцепочечность
3. Комплементарность
Г
Т
4. Антипараллельность
Т
Г
3'
47. Неканоническая комплиментарность
48. Формы ДНК.
49. В-форма ДНК.
Молекула ДНК высокой влажности получиланазвание В-формы. При физиологических условиях
(низкая концентрация соли, высокая степерь
гидратации) доминирующим структурным типом ДНК
является В-форма (основная форма двухцепочечной
ДНК - модель Уотсона-Крика). Шаг спирали такой
молекулы равен 3,4 нм. На виток приходится 10
комплементарных пар в виде скрученных стопок
"монет" - азотистых оснований. Стопки удерживаются
водородными связями между двумя
противолежащими "монетами" стопок, и "обмотаны"
двумя лентами фосфодиэфирного остова,
закрученными в правую спираль. Плоскости азотистых
оснований перпендикулярны оси спирали. Соседние
комплементарные пары повернуты друг относительно
друга на 36°. Диаметр спирали 20Å, причем
пуриновый нуклеотид занимает 12Å, а
пиримидиновый - 8Å.
50. А-форма ДНК.
Молекула ДНК более низкой влажности получиланазвание А-формы. А-форма образуется в условиях
менее высокой гидратации и при более высоком
содержании ионов Na+ или К+. Эта более широкая
правоспиральная конформация имеет 11 пар азотистых
оснований на виток. Плоскости азотистых
оснований имеют более сильный наклон к оси спирали,
они отклонены от нормали к оси спирали на 20°. Отсюда
следует наличие внутренней пустоты диаметром 5Å.
Расстояние между соседними нуклеотидами составляет
0,23 нм, длина витка – 2,5 нм, диаметр спирали – 2,3 нм.
А-форму имеет спираль РНК-ДНК в комплексе матрицазатравка, а также спираль РНК-РНК и шпилечные
структуры РНК. А-форма ДНК обнаружена в спорах.
Установлено, что А-форма ДНК в 10 раз устойчивее к
действию УФ-лучей, чем В-форма.
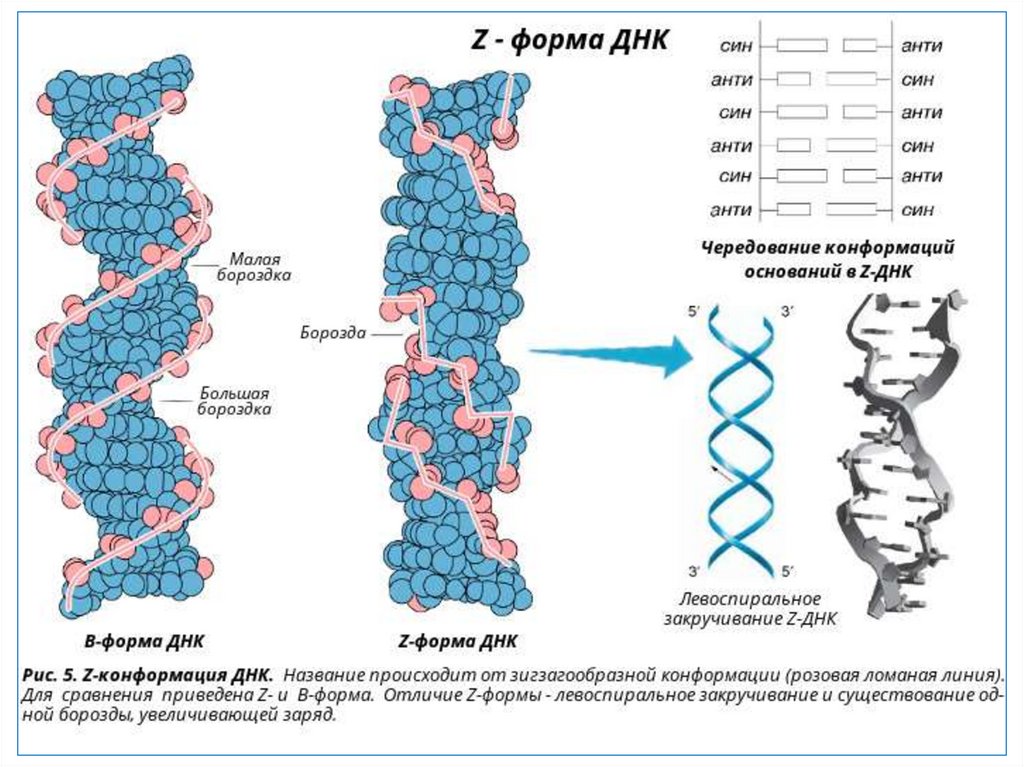
51. Z-форма ДНК.
Z-форма ДНК была открыта в 1979 году при изучениигексануклеотида d(CG)3 - [CG-CG-CG]. Ее открыл
профессор Массачусетского технологического
института Александр Рич с сотрудниками. Z-форма
стала одним из важнейших структурных элементов
ДНК в связи с тем, что ее образование наблюдалось в
участках ДНК, где пурины чередуются с пиримидинами
(например, 5’-ГЦГЦГЦ-3’), или в повторах 5’-ЦГЦГЦГ-3’,
содержащих метилированный цитозин. Существенным
условием образования и стабилизации Z-ДНК
являлось присутствие в ней пуриновых нуклеотидов в
син-конформации, чередующихся с пиримидиновыми
основаниями в анти-конформации.
Z-форма ДНК левозакручена.
52. Син- и анти- конформации.
Син-конформацияпуриновых
оснований
Анти-конформация
пуриновых
оснований
Син-конформация
пиримидиновых
оснований
Анти-конформация
пиримидиновых
оснований
53.
54. Формы ДНК. А, В и Z.
55. Структурные образования ДНК.
На молекуле ДНК выделяют следующиеструктурно-функциональные
образования:
• Смысловые последовательности
• Гены
последовательности
• Несмысловые
• Повторяющиеся
последовательности
• Псевдогены
фрагменты
• Генные
• Транспозоны
56. Повторяющиеся последовательности ДНК.
Длинные повторыСателлитная ДНК
Средние повторы
Диспергированные
по геному
транспозоны
Тандемные повторы
Копии генов
Минисателлиты
Макросателлиты
SINEs
LINE
Гены рРНК
Тандемные
повторы
Динуклеотидные повторы
Alu-повторы
L1-повторы
57. Характеристика повторяющихся последовательности ДНК.
1. Сателлитная ДНК – тандемные повторы с высокой степенью копийности,расположенные группами в гетерохроматиновых областях хромосом,
фланкирующих центромеры.
2. Вариабельные по числу тандемные повторы (минисателлиты) – области повторов
длиной 15-100 п.н., расположенные внутри и вне генов.). STRs (short tandem
repeats).
3. Короткие тандемные повторы (микросателлиты) – ди-, три-, тетра- и
пентануклеотидные последовательности с копийностью 5-50 раз. Количество
повторов уникально. VNTRs (variable number tandem repeats).
4. SINE (short interspersed elements) – последовательности длиной около 500 п.н.,
копийность не менее 500 000.
5. LINE (long interspersed elements) – последовательности длиной около 6 000 п.н.,
копийность не менее 850.
58. Псевдогены
Псевдоген – нефункциональный ген, имеющий высокуюгомологию со структурным геном, который присутствует в
другом месте генома.
От функциональной версии отличается вставки или удаления и
по наличию фланкирующих последовательностей с прямым
повторением 10–20 нуклеотидов.
Исходный ген
Исходный ген
Исходный ген
Дупликация
Мутация
Псевдоген
59. Псевдогены
60. Транспозоны
Kristian Alsbjerg Skipper, 2013C. Sandeep Kumar, 2013
61.
ГЕН????????????62. Краткая характеристика гена
Ген - структурно-функциональнаяединица ДНК, кодирующая
как минимум одну
полипептидную цепь и транскрибируемая с
образованием единой РНК.
63. Краткая характеристика гена
12
3
2
2
4
1. Промоторная область – область инициации
считывания генетической информации.
2. Экзонные области – области, содержащие
информацию о последовательности нуклеотидов в
полипептиде.
3. Интронные области – области, содержащие
регуляторные элементы.
4. Прерыватель транскрипции – область конечной
модификации РНК
64. Классификации генов
65.
Часть 2.Основные понятия.
66. Такая разная…
• Генетика – биологическая наука, изучаетзаконы передачи наследственных
признаков у всего живого
• Генетика человека – часть генетики
млекопитающих, изучает прицельно
Homo Sapiens
• Медицинская генетика – в качестве
признаков рассматривает только болезни
67. Формальная генетика
• Изучает законы передачи признаков изпоколения в поколение (наследование):
– сохранение (наследственность) и
– варьирование (изменчивость)
• Имеет дело с парой особей и их
потомством после скрещивания.
Пара нередко формируется искусственно
(гибридологический анализ)
• Использует два основных метода:
описательный и статистический
68. Фенотип
• Совокупность всех признаков организма• Применим к отдельным наборам признаков,
например марфаноидный фенотип
• Признак:
– Количественный либо качественный
– Простой либо сложный
• Противоположный термин - генотип
69. Генотип
• Характеристика всех генов конкретногочеловека (количественная и качественная)
• Ген – структурно-функциональная единица
наследственной информации
• Количественная характеристика
определяется кариотипом (проще)
• Качественная характеристика – наличием
или отсутствием мутаций в генах
• Нередко только кариотип или конкретные
мутации в отдельных генах
70. Геном (не путать)
• Имеет видовую принадлежность: геномчеловека, мыши, кузнечика, овса и т.п.
• Геном человека – совокупность генов,
необходимых для формирования homo
sapiens
• Каждый изученный ген имеет название
(CFTR, AR, PAH), локализацию (13q12.2),
известны длина, число экзонов, нормальная
нуклеотидная последовательность
71. Аллелль
• Аллель – состояние (вариант) гена• Природное (эволюционное) состояние гена
называется дикий аллель
• Популяция только здоровых людей
аналогична чистой линии
• Мутация в гене приводит к образованию
мутантного аллеля (нас интересуют те, что
приводят к болезням)
• Наличие обоих аллелей в популяции
усложняет запись и генофонда и генотипа
72. Аллельные гены
Аллель (от греч. ἀλλήλων — друг друга, взаимно) —различные формы одного и того же гена, расположенные
в одинаковых участках (локусах) гомологичных хромосом и
определяющие альтернативные варианты реализации одного
и того же признака
72
73. Аллельный полиморфизм
Аллельный полиморфизм — различия в нуклеотиднойпоследовательности аллельных генов
ACCGTAGGCCTAAGCTGACTGCA
ACCGTAGGCCTGAGCTGACTGCA
Регуляторный
• Промоторный
• Интронный
Функциональный
• Экзонный
А/А
гомозигота,
если А/G —
гетерозигота
73
74. Запись генотипа
• Гомозиготное состояние – аллели одногогена совпадают: АА, аа
• Гетерозиготное состояние – аллели одного
гена не совпадают: Аа
• Доминирование – способность аллеля
проявляться в гетерозиготном состоянии
• Запись усложняется при локализации генов
в половых хромосомах, при числовых
аномалиях хромосом и множ. аллелизме
75.
Часть 3.Наследование признаков.
76. Основные типы наследования признака
Доминантный ген обеспечивает проявление признака как в гомозиготном, так и вгетерозиготном состоянии. Пример: А/А или А/a
Рецессивный ген обеспечивает проявление признака только в гомозиготном
состоянии. Пример: a/a
77.
Законы наследования МенделяСкрещивание особей, гомозиготных по разным аллелям,
дает генетически однородное потомство (поколение F1),
все особи которого гетерозиготны по этим аллелям. При
этом, какая из двух гомозиготных особей мужского пола, а
какая женского, значения не имеет – закон однородности
и реципрокности. Свойство реципрокности справедливо
только для аутосомных генов.
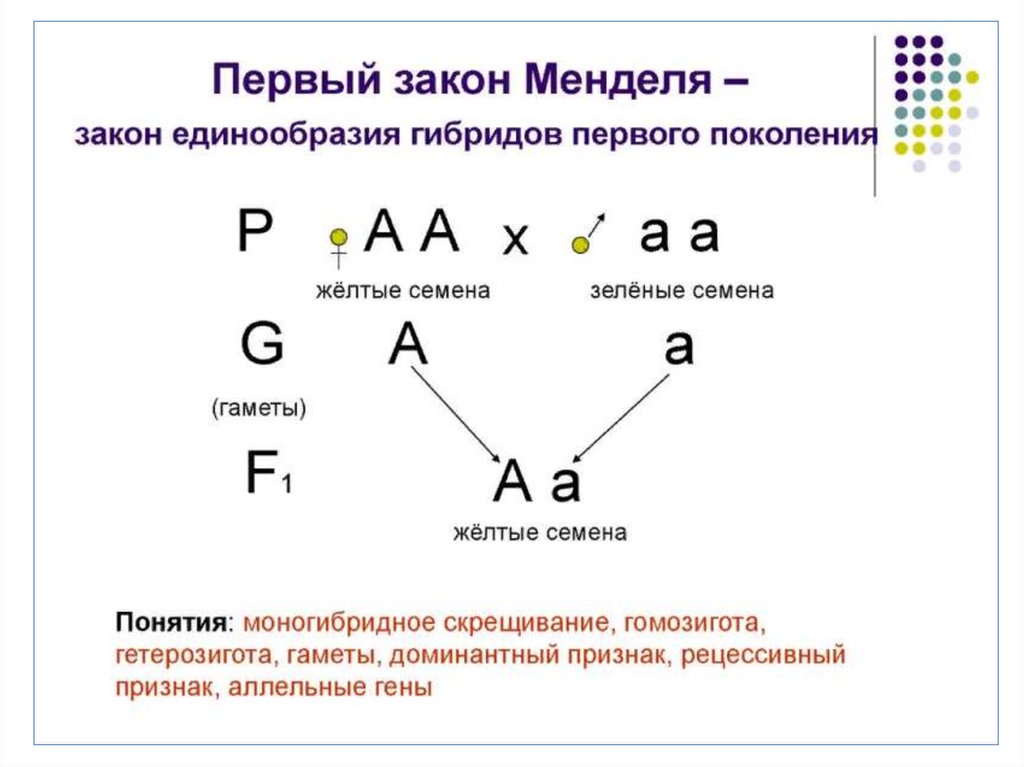
78. Первый закон Менделя –
закон единообразия гибридов первогопоколения
1-й закон Менделя подразумевает, что при
скрещивании двух гомозиготных организмов,
относящихся к чистым линиям и отличающихся друг от
друга одной парой альтернативных проявлений
определенного признака, первое поколение гибридов
(F1) будет одинаковым и будет нести проявление
признака лишь одного из родителей.
79.
закон единообразия гибридов первогопоколения
80.
Законы наследования МенделяПри скрещивании гетерозигот поколения F1 между собой
(интеркросс) выщепляются разные генотипы: половина из них
снова оказываются гетерозиготами, а гомозиготные потомки
каждого из двух родительских типов составляют по одной
четверти. И в следующих поколениях при скрещивании
гетерозигот повторяется такое же отщепление 1:2:1, тогда как в
скрещиваниях
одинаковых
гомозигот
расщепления
нет.
Мендель правильно объяснил этот результат, предположив, что
у гетерозигот образуются зародышевые клетки двух типов в
отношении 1:1 - закон расщепления и закон чистоты гамет.
81. Второй закон Менделя –
В результате скрещивания двух гетерозиготныхпотомств первого поколения между собой, можно
наблюдать расщепление во втором поколении. Это
расщепление имеет определенное числовое
соотношение по фенотипу — 3:1, а по генотипу —
1:2:1.
82.
83.
Законы наследования МенделяПри скрещивании особей, различающихся по двум и
более
парам
генов,
каждая
пара
расщепляется
независимо. Наблюдаемые сегрегационные отношения
определяются статистическим законом независимого
комбинирования.
Этот закон справедлив только при
отсутствии сцепления.
84. Третий закон Менделя – закон независимого расщепления
Третий закон наследственности гласит, что прискрещивании двух особей, отличающихся одна от
другой по нескольким парам альтернативных
признаков (двум и более), происходит независимое
наследование генов и соответствующих им признаков,
а также комбинирование во всех доступных сочетаниях
(как при моногибридном скрещивании).
85.
86.
Менделевские типы наследования• Если мы знаем характер генетического контроля признака, то
мы можем предсказать с определенной вероятностью
фенотип на основе генотипа (если он известен).
• Если мы знаем связь между генотипом и фенотипом, то мы
можем сделать определенные предсказания о развитии
признака (например, болезни) и, в некоторых случаях,
предпринять действия полезные индивиду.
87.
Менделевские типы наследования• На основании своих опытов Мендель пришел к выводу, что далеко
не каждому генотипу соответствует четко отличающийся фенотип:
часто гетерозиготы в той или иной степени сходны с одной из
гомозигот.
• Аллель, который определяет фенотип гетерозиготы, Мендель
назвал
доминантным,
а
другой,
не
гетерозиготном состоянии, - рецессивным.
проявляющийся
в
88.
Менделевские типы наследованияВ рамках моногенного наследования выделяют:
• аутосомно-доминантный тип (на одной из двух аутосом
расположен доминантный ген);
• аутосомно-рецессивный тип (на одной из двух аутосом
расположен рецессивный ген);
• Х-сцепленный доминантный тип (на Х-хромосоме
расположен доминантный ген);
• Х-сцепленный рецессивный тип (на Х-хромосоме
расположен рецессивный ген);
• Y-сцепленный тип или голандрическое наследование (ген
расположен на Y-хромосоме).
89.
Менделевские типы наследования• Типы доминирования. Мендель наблюдал, что при
объединении задатков двух признаков в одном организме
может быть проявление только одного признака. Второй
задаток
не
проявляется.
Такой
тип
доминирования
называется полным.
• В тех случаях, когда фенотип гетерозиготы является
промежуточным между фенотипами гомозигот, говорят о
промежуточном, или неполном доминировании.
90.
Менделевские типы наследования91.
Менделевские типы наследования• В последние годы в связи с разработкой методов,
позволяющих проводить анализ на уровне, более близком к
первичному эффекту генов, обнаруживается все больше
примеров, когда каждому генотипу соответствует свой
фенотип, отличный от остальных. Этот тип наследования
иногда называют кодоминированием. Он очень популярен в
последнее время в суде.
Фенотипическое проявление кодоминантного
взаимодействия генов на примере цветка
рододендрона (Rhododendron).
92.
Менделевские типы наследования вих приложении к человеку
• Аутосомно-рецессивный
тип
наследования.
Тип
наследования при котором гетерозигота фенотипически не
отличается от нормальной гомозиготы.
• В отличие от доминантного наследования, при котором
почти все пораженные потомки происходят от браков
гетерозигот со здоровыми гомозиготами, большинство
браков, наблюдаемых при рецессивных заболеваниях
происходит
между
фенотипически
нормальными
гетерозиготами.
• В потомстве такого брака генотипы АА, Аа и аа будут
представлены в отношении 1:2:1 и вероятность того, что
ребенок окажется пораженным, составляет 25%.
93.
Менделевские типы наследования вих приложении к человеку
• В популяции, хотя и довольно редко, встречаются браки гетерозигот и
пораженных гомозигот. В таком браке среди детей ожидается
сегрегационное отношение 1:1. Поскольку отношение 1:1 характерно
и при доминантном наследовании, то эта ситуация называется
«псевдодоминированием». К счастью, браки между гетерозиготами
и пораженными гомозиготами редки.
94.
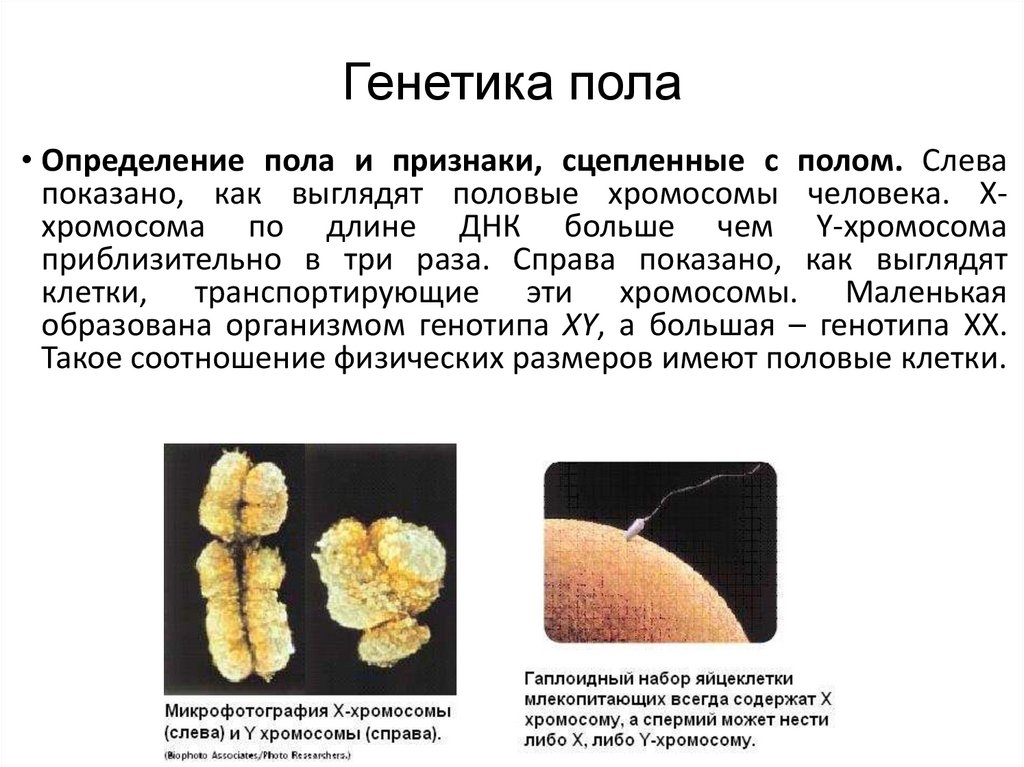
Генетика пола• Определение пола и признаки, сцепленные с полом. Слева
показано, как выглядят половые хромосомы человека. Ххромосома по длине ДНК больше чем Y-хромосома
приблизительно в три раза. Справа показано, как выглядят
клетки, транспортирующие эти хромосомы. Маленькая
образована организмом генотипа XY, а большая – генотипа ХХ.
Такое соотношение физических размеров имеют половые клетки.
95.
Генетика пола• На слайде показано, как выглядит человек после того, как он уже
начал развиваться: это игла, и на ее конце находится несколько
поделившихся клеток, полученных из оплодотворенной
яйцеклетки. Таков наш размер на стадии 4-5-ого деления.
96.
Генетика пола• Не у всех организмов, в отличие от человека, пол
однозначно определяется при слиянии гамет. У некоторых
пресмыкающихся мужской пол не формируется при
повышенной температуре (в частности, у крокодилов),
образуются только женские организмы.
• У птиц и ряда других групп организмов гомогаметный пол
(ХХ) – мужской, а гетерогаметный пол (XY) – женский, что
противоположно тому типу хромосомного определения
пола, который оказался зафиксирован эволюцией в линии
человека и других млекопитающих.
97.
Генетика пола• На рисунке представлен случай
отклонения по составу половых
хромосом – синдром Тернера:
индивид генотипа Х0 (Yхромосомы нет, а Х-хромосома
одна). Фенотип индивида женский.
Отклонений
в
поведении нет, хотя при
большем
дисбалансе
в
генотипе
наблюдаются
проблемы
в
умственном
развитии.
Другой случай отклонения по составу половых хромосом - это синдром
Клайнфельтера: генотип XXY, фенотип мужской. Хотя две Х-хромосомы
присутствуют, как в норме у женщин. То есть для определения пола у
человека важно, чтобы присутствовала Y-хромосома.
98.
Генетика пола• Чтобы сбалансировать действие генов Х хромосомы у мужчин
(где Х-хромосома одна) и у женщин (где Х-хромосомы две)
природа предусмотрела инактивацию одной из Х-хромосом в
каждой клетке женского организма. У женщин одна Ххромосома инактивирована, то есть гены в ней не работают, а
работают только в другой хромосоме – не инактивированной.
Инактивиация вызвана компактизацией хромосомы даже в
интерфазе. Поэтому компактизированная хромосома видна
при окрашивании – в виде так называемого тельца Барра. Это
один из диагностических признаков женского пола у
млекопитающих,
что
у
человека,
что
у
слона.
Некомпактизованная Х-хромосома, как и все аутосомы, не
окрашивается и не видна в интерфазе.
99.
Генетика пола• Х-сцепленные типы наследования. У человека каждый
брак можно рассматривать как менделевский бэккросс
(возвратное скрещивание) в отношении Х- и Y-xpoмосомы:
100.
Генетика пола• Гены, локализованные в Y-хромосоме. Y-хромосома несет сравнительно
мало генов (по последним данным международного каталога генов OMIM,
всего около 40). Небольшая часть таких генов гомологична генам Xхромосомы, остальные присутствующие только у мужчин, участвуют в
контроле детерминации пола и сперматогенеза. Так, на Y-хромосоме
находятся гены SRY и AZF, ответственные за программу половой
дифференцировки. Мутации в любом из этих генов приводят к нарушениям
развития яичек и блоку сперматогенеза, что выражается в азооспермии.
Такие мужчины страдают бесплодием, и потому их заболевание не
наследуется.
101.
Качественные и количественныепризнаки
• Они различаются по характеру изменчивости и особенности
наследования. Качественные признаки характеризуются
прерывистой,
а
количественные
—
непрерывной
изменчивостью.
• Первые из них дают четкие границы при расщеплении на
доминантные или рецессивные признаки. Это связано с тем,
что каждый из них обычно контролируется одним аллельным
геном.
• Количественные признаки не дают четких границ расщепления
при разных вариантах скрещивания, хотя отличаются от
качественных более высокой степенью изменчивости.
102.
Качественные и количественныепризнаки
• Особенностью количественных признаков является сложный
характер наследования. Каждый из них детерминируется не
одним, а множеством локусов в хромосомах.
• Такой тип наследования, когда один признак обусловливается
многими генами, носит название полигенного.
• Уровень развития количественного признака зависит от
соотношения доминантных и рецессивных генов, других
генетических факторов и степени модифицирующего действия
факторов внешней среды.
103.
Полигенные признаки• ПРИМЕР:
• Рассмотрим это на примере слуха. Чтобы человек слышал,
нужны многие структуры уха: молоточки, наковаленки,
барабанные перепонки, и если не будет какого-то белка,
нужного, чтобы сформировать эти структуры, то человек не
будет слышать. Поэтому глухота у человека может возникать
из-за дефектов в десятках известных генов. При этом разные
генотипы могут приводить к одному фенотипу (глухота).
104.
Наследование полигенных признаков• Отдельный ген может влиять на несколько признаков, в
развитие которых он вовлечен. Такой эффект действия гена
называется плейотропным (множественным).
• Например, альбинизм вызывается дефектом одного гена,
контролирующего образование меланина. При этом меняются
одновременно такие признаки как цвет волос (белый), цвет
кожи (белый) и цвет глаз, (он будет красным от цвета
кровеносных сосудов, лежащих за радужной оболочкой глаза).
105.
Наследование полигенных признаков• Часть ядерных генов наследуется только однородительски. Это
гены Y-хромосомы, которая наследуется только от отца к сыну.
Те гены в Y-хромосоме, которые не имеют пары в Х-хромосоме,
наследуются однородительски – от отца к сыну. У человека
один из таких генов определяет признак «волосатые уши» – у
женщин такой признак не проявляется – гена этого нет, нет и
его аллеля, из-за которого уши волосатые.
106.
Наследование полигенных признаков• Проявление признака может зависеть также от пола индивида.
Существуют признаки, гены которых могут находиться в
аутосомах
или
половых
хромосомах
обоих
полов,
но
проявляются лишь у одного из них. Такие признаки называют
ограниченными полом.
• Например, гены казеинов – белков молока – это случай
наиболее строгого контроля транскрипции ограниченной
полом. Разница уровней экспрессии между ними составляет 1
на 100 000.
107.
Модели наследования признаков• При создании модели наследования признака для выяснения
его генетического контроля начинают с проверки типа
наследования по базовой, простейшей схеме - менделевской
и, если соответствия нет, то добавляют к этой схеме
дополнительные условия, понятные на сегодня:
– неравную вероятность образования и сочетания гамет и выживания
потомства;
– однородительский тип наследования (Y – волосатые уши, мтДНК болезнь Лебера)
– взаимодействие аллелей: наличие доминирования;
– взаимодействие генов;
– сцепление генов;
– ограничения признака полом;
– вероятность проявления признака при данном генотипе;
– влияние среды (не всегда формализуемое).
108.
Закон Харди–Вайнберга и егоприложения
• Формулировка закона Харди—Вайнберга. До сих пор
применение законов Менделя к наследованию признаков у
человека мы рассматривали с точки зрения изучения отдельных
семей. Каковы, однако, следствия из этих законов для
генетической структуры популяций? Область науки, в которой
решаются такие проблемы, называется популяционной
генетикой.
• Закон Харди—Вайнберга был сформулирован в 1908 г.
независимо английским математиком Годфри Гаральдом Харди
и немецким врачом Вильгельмом Вайнбергом. Следует
заметить, что еще в 1904 г. Пирсон, пытаясь применить
менделевские правила к количественным признакам, говорил
об этой закономерности для специального случая равных частот
двух аллелей.
109.
Годфри Харолд Харди (англ.Godfrey Harold Hardy; 1877 1947, Кембридж,
Великобритания) —
английский математик.
Известен работами в теории
чисел и математическом
анализе.
Вильгельм Вайнберг (нем.
Wilhelm Weinberg, 1862,
Штутгарт — 1937, Тюбинген)
— немецкий врач. Основатель
Штутгартского общества
расовой гигиены.
110. Закон Харди –Вайнберга (1908)
При определенных условиях, не изменяющих частоты аллелей ,популяция имеет определенное соотношение особей с
доминантными и рецессивными признаками, при этом частоты
аллелей остаются неизменными в ряду поколений
111. Частоты генотипов
Аллели: А,аГенотипы:
АА= n1
Аа= n2
аа = n3
N (общее число особей )= n1+n2+n3
Доля генотипа АА (частота )= n1/N
Доля генотипа Аа(частота)= n2/N
Доля генотипа аа (частота)= n3/N
112. Частоты генов
Аллели: А,аГенотипы:
АА= n1
Аа= n2
аа = n3
N (общее число особей )= n1+n2+n3
Доля гамет, содержащих аллель «А»
(частота гена А ) p= n1 +1/2n2
N
Доля гамет, содержащих аллель «а»
(частота гена а ) q= n3 +1/2n2
N
113. Распределение индивидов в различных выборках по группам крови системы «MN» и частоты аллелей
ГородСанктПетер
бург
Число
Число индивидов
обследо
МN
NN
ванных ММ
701
225
327
149
Число
аллелей , %
LМ
LN
?
?
114. Для Санкт-Петербурга
Доля гамет, содержащих аллель «А»(частота гена А ) p= n1 +1/2n2
225+ 327/2
701
N
Доля гамет, содержащих аллель «а»
(частота гена а ) q= n3 +1/2n2
N
327/2+ 149
701
0, 554
(55.4%)
0, 446
(44,6%)
115. Распределение индивидов в различных выборках по группам крови системы «MN» и частоты аллелей
ГородЧисло индивидов
Число
обследо
МN
NN
ванных ММ
СанктПетер 701
бург
225
327
149
Число аллелей ,
%
LМ
LN
55,42
44,58
116. Закон Харди-Вайнберга
Фонд отцов(Рсп +qсп)
=
Фонд матерей
(Ряц +qяц)
2
2
2
(P+q) =P +2Pq +q
117. Частота (вероятность) генотипов потомков при панмиксии
Гены ивероятности
А /p
а/q
А/р
АА/р2
Аа/pq
а/q
Аа/pq
аа/q2
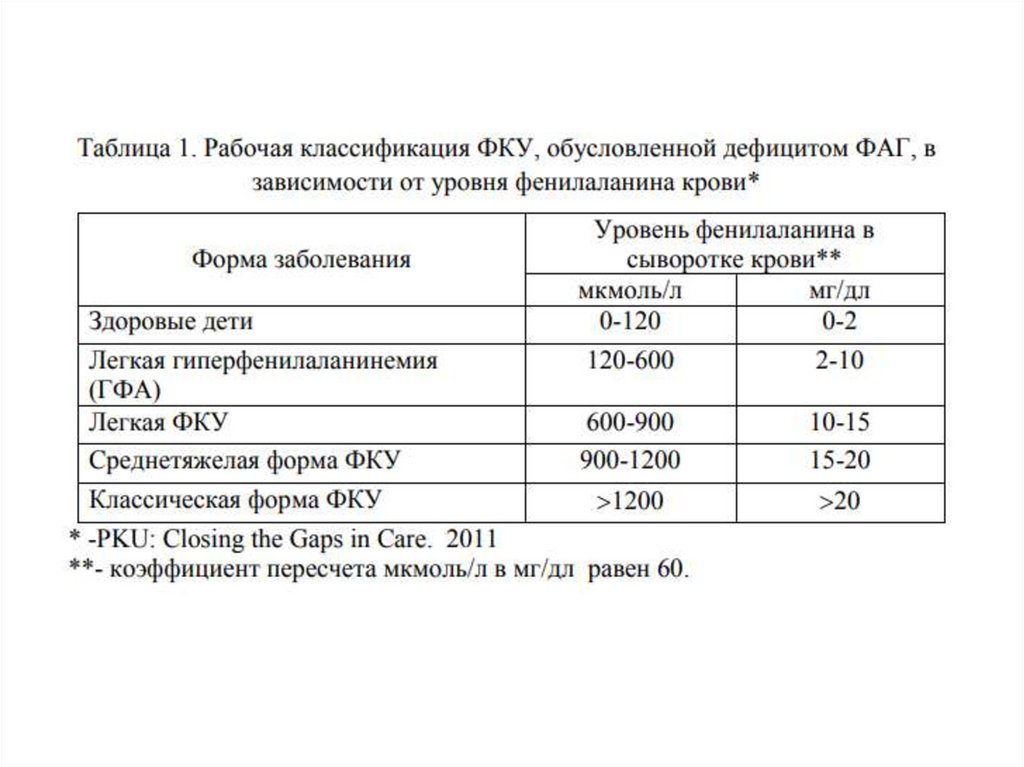
118. ФЕНИЛКЕТОНУРИЯ
Фенилкетонури́я (фенилпировинограднаяолигофрения) — наследственное заболевание
группы ферментопатий, связанное с нарушением
метаболизма аминокислот, главным образом
фенилаланина. Сопровождается накоплением
фенилаланина и его токсических продуктов, что
приводит к тяжёлому поражению ЦНС,
проявляющемуся, в частности, в виде нарушения
умственного развития.
119. Фенилкетонурия
Тип наследования – Аутосомно-рецессивныйПатогенез – наследственное заболевание, связанное с накоплением в
организме аминокислоты фенилаланина и ее токсическим действием на ЦНС.
Заболеваемость – Распространенность ФКУ варьируется во всем мире. У
Caucasians распространенность составляет примерно 1:10 000 живорожденных, а
в России - 1: 7000.
Этиология – мутации гена Phenylalanine hydroxylase (PAH), приводящие к
дефициту белка. Локус 12q23.2
120. Фенилкетонурия
5,350,9
Yajie Su, 2019
121.
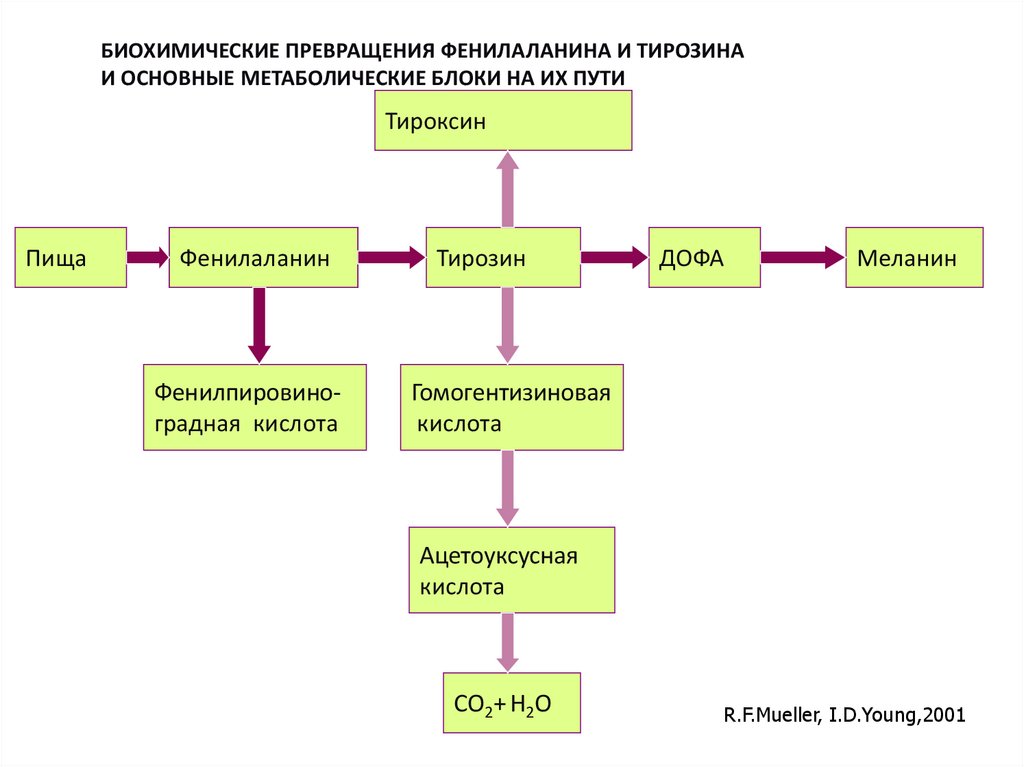
БИОХИМИЧЕСКИЕ ПРЕВРАЩЕНИЯ ФЕНИЛАЛАНИНА И ТИРОЗИНАИ ОСНОВНЫЕ МЕТАБОЛИЧЕСКИЕ БЛОКИ НА ИХ ПУТИ
Тироксин
Пища
Фенилаланин
Фенилпировиноградная кислота
Тирозин
ДОФА
Меланин
Гомогентизиновая
кислота
Ацетоуксусная
кислота
СО2+ Н2О
R.F.Mueller, I.D.Young,2001
122.
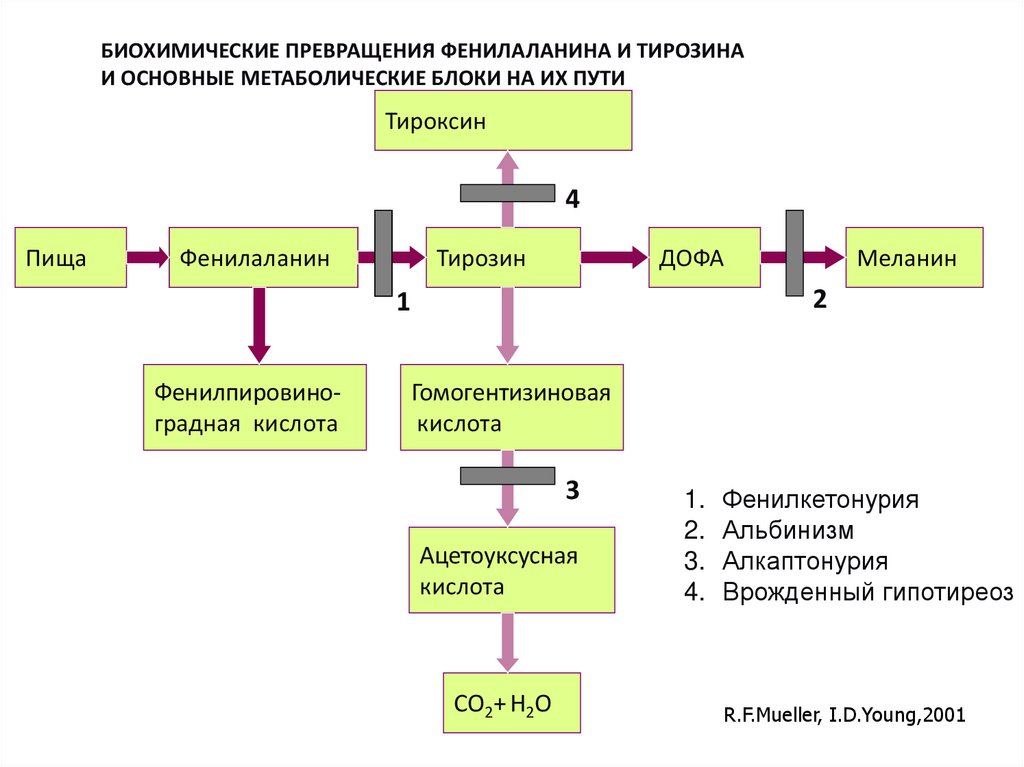
БИОХИМИЧЕСКИЕ ПРЕВРАЩЕНИЯ ФЕНИЛАЛАНИНА И ТИРОЗИНАИ ОСНОВНЫЕ МЕТАБОЛИЧЕСКИЕ БЛОКИ НА ИХ ПУТИ
Тироксин
4
Пища
Фенилаланин
Тирозин
ДОФА
2
1
Фенилпировиноградная кислота
Меланин
Гомогентизиновая
кислота
3
Ацетоуксусная
кислота
1. Фенилкетонурия
2. Альбинизм
3. Алкаптонурия
4. Врожденный гипотиреоз
СО2+ Н2О
R.F.Mueller, I.D.Young,2001
123.
124. ДИАГНОСТИКА
Производится полуколичественным тестом иликоличественным определением фенилаланина в
крови.
При нелеченных случаях возможно выявление
продуктов распада фенилаланина в моче (не ранее
10-12 дня жизни ребенка).
Также возможно определение активности фермента
фенилаланингидроксилазы в биоптате печени
поиск мутаций в гене фенилаланингидроксилазы
125. Клиническая картина
• С 2-6 мес жизни• Нарушения вскармливания: рвоты, поносы, отказ от
еды
• Гипотония мышц, судороги, раздражительность,
вялость
• Эпилепсии, плохо поддающиеся терапии
• «Мышиный» запах мочи
• Аллергический дерматит, экзема
• Светлые кожные покровы, волосы, радужная
оболочка глаз
• Задержка психомоторного развития
• Снижение интеллекта (вплоть до идиотии)
126. ЛЕЧЕНИЕ
При своевременной диагностике патологических измененийможно полностью избежать, если с рождения и до полового
созревания ограничить поступление в организм фенилаланина с
пищей.
Позднее начало лечения хотя и даёт определённый эффект, но не
устраняет развившихся ранее необратимых изменений ткани
мозга
Лечение проводится в виде строгой диеты от обнаружения
заболевания как минимум до полового созревания, многие
авторы придерживаются мнения о необходимости пожизненной
диеты. Диета исключает мясные, рыбные, молочные продукты и
другие продукты, содержащие животный и, частично,
растительный белок. Дефицит белка восполняется
аминокислотными смесями без фенилаланина.
127. Лечение
Диет-питание с ограничением продуктов, богатых фенилаланином;
специализированные аминокислотные смеси без фенилаланина.
Симптоматическое.
128.
Лечение• Пищевой светофор
129.
Лечение130. Задача
Частота = 1 ребенок на 10000 новорожденныхq2= 1/10000
q=√ 1/10000= 0,01
P=1-q=0,99
2pq= 2*0,99*0,01=0,0198
N=25000
Количество гетерозигот= 0,0198*25000=495
(чел)
Фенилкетонурия
131.
Закон Харди–Вайнберга и егоприложения
• Значение равновесия Харди—Вайнберга. Если наблюдаемые
доли генотипов в популяции соответствуют ожидаемым из
закона
Харди—Вайнберга,
то
говорят,
что
популяция
находится в равновесии Харди—Вайнберга.
• Равновесие
Харди—Вайнберга
—
это
равновесие
распределения генов в популяции (пула генов) между разными
генотипами. Если равновесие было нарушено какими-либо
силами, то при случайном скрещивании оно восстановится за
одно поколение.
132.
Закон Харди—Вайнбергасправедлив при условиях:
1.Скрещивания в отношении изучаемых фенотипов
должны быть случайными.
2. Отклонение от случайного скрещивания может
быть вызвано, в частности, кровнородственными
браками: если в популяции высокий уровень
кровного родства, то следует ожидать увеличения
количества гомозигот. Можно даже вычислить
частоту кровнородственных браков в популяции на
основе отклонений от соотношений Харди—
Вайнберга.
3. Соотношения Харди—Вайнберга могут быть
нарушены миграциями.
133. Факторы, нарушающие равновесие
• Инбридинг• Дрейф генов (с частными вариантами)
• Поток генов
• Мутационный процесс
• Естественный отбор
134. Инбридинг
• Родственный брак – у супругов есть общийпредок или два предка
• Потомство называется инбрендным
• Рассчитывается коэффициент инбридинга
для пары и для популяции
• Работает чаще на мутантный аллель
135. Дрейф генов
• Работает в малых популяциях с хорошейизоляцией
• Эффективная часть популяции –
способная дать потомство в ближайшие
годы
• Обеспечивает случайный сдвиг в пользу
любого аллеля при смене поколений
136. Эффект основателя – частный вар.
• явление снижения исмещения
генетического
разнообразия при
заселении малым
количеством
представителей
рассматриваемого
вида новой
географической
территории.
• Термин ввёл Э.
Майр;
137. Поток генов
• Миграционный процесс• Измеряется путём отслеживания
популяционной частоты носительства
аллелей (изменения по годам)
• Зависит от видов и степени изоляции,
для человека – социальные и
политические факторы
• Работает на выравнивание
межпопуляционной разницы
138. Мутационный процесс
• Весьма редкое событие 10 в -5 -6• Работает на мутантный аллель
• Почему-то варьирует для разных генов
• Может исчисляться специальным
индексом (изучается на сперматогензе)
139. Естественный отбор
• Работает, как правило, на повышениечастоты дикого аллеля
• Специальный индекс обозначает
возможность доживания до
репродуктивного возраста и фертильности
• При индексе 0,0 мутация поддерживается
только за счёт de novo
140.
Закон Харди–Вайнберга и егоприложения
Иногда в качестве фактора, нарушающего соотношение Харди—
Вайнберга, упоминается отбор. Это справедливо, но не
обязательно. Как правило, отбор изменяет генные частоты.
Однако отбор, действующий до репродуктивного возраста,
например в пренатальном периоде или позже, включая детство и
юность, совсем не влияет на соотношения Харди—Вайнберга у
взрослых. Отклонения могут наблюдаться в субпопуляции детей в
зависимости от конкретного типа отбора. Кроме того, даже при
сильном отборе в соответствующей возрастной группе
регистрация статистически значимых отклонений от соотношений
Харди-Вайнберга требует выборок большого объема — больше,
чем обычно имеется в распоряжении исследователя. Иногда
вывод об отсутствии значимого отбора формулируется исходя из
наблюдаемого соответствия популяции соотношениям Харди—
Вайнберга.
141.
Закон Харди–Вайнберга и егоприложения
Формально отклонение от закона Харди—Вайнберга
может наблюдаться в том случае, если популяция
представляет собой смесь субпопуляций, лишь
частично скрещивающихся между собой (случайное
скрещивание происходит только внутри субпопуляций,
и, следовательно, генные частоты в этих субпопуляциях
различны).
142.
Закон Харди–Вайнберга и егоприложения
Другой причиной отклонения может служить
существование пока не выявленного («немого»)
аллеля, в связи с чем гетерозиготные носители этого
аллеля неотличимы от гомозигот по обычному аллелю.
Однако Смит (1970) указал, что генетически немой
аллель может вызывать значимое отклонение от
закона Харди—Вайнберга только тогда, когда
гомозиготы по этому аллелю имеют частоту,
достаточно высокую, чтобы быть выявленными.
143.
Статистические методы формальнойгенетики: анализ сегрегационных
отношений
• Сегрегационные отношения как вероятности. В ходе мейоза
при отсутствии каких-либо нарушений различные гаметы
образуются в точно таких относительных пропорциях, какие
ожидаются
из
законов
Менделя.
Из
диплоидного
гетерозиготного сперматоцита с аллелями А и а образуются
гаплоидные сперматозоиды: два с аллелем А и два с
аллелями а. Если бы все сперматозоиды данного индивида
участвовали в оплодотворении и если бы ни одна из зигот не
погибла до рождения, то сегрегационное отношение среди
его потомства было бы в точности 1:1.
144.
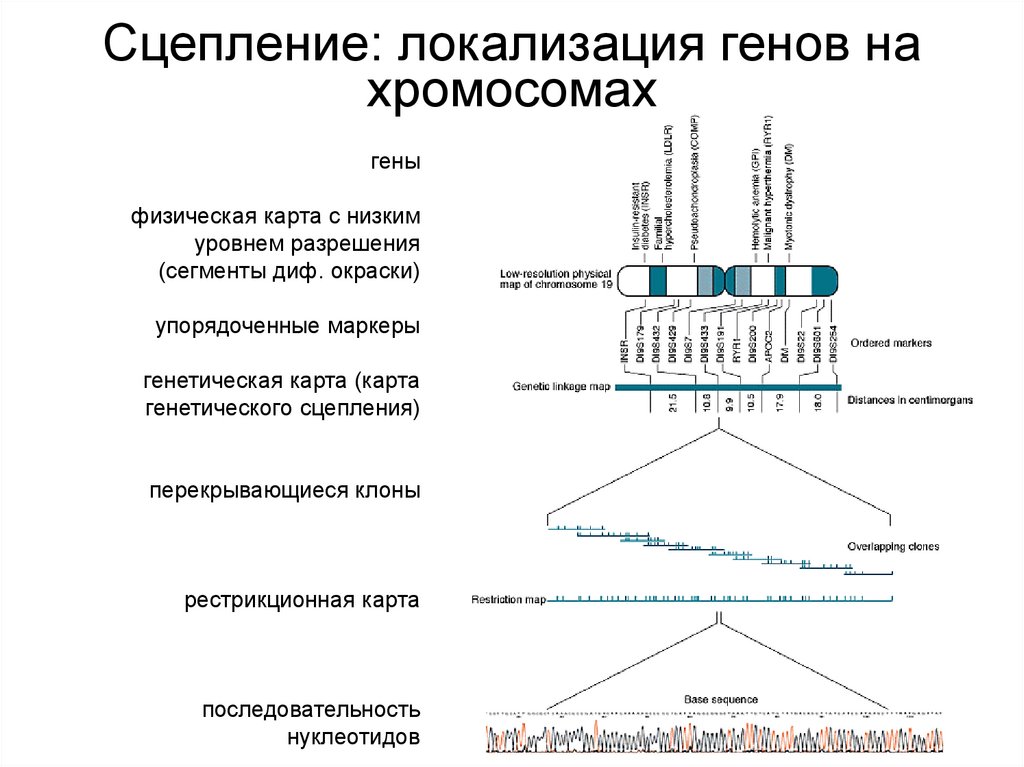
Сцепление: локализация генов нахромосомах
• Картирование генов. Гены расположены в
хромосомах в линейном порядке. Отсюда следует,
что, во-первых, гены, локализованные в одной
хромосоме, передаются совместно, а во-вторых,
сегрегация сцепленных генов не является
независимой. С другой стороны, известно, что во
время первого мейотического деления образуются
хиазмы и гомологичные хромосомы обмениваются
между
собой
определенными
сегментами
(кроссинговер).
Таким
образом,
гены,
расположенные в одной хромосоме, не всегда
передаются вместе. Вероятность совместной
передачи двух сцепленных генов зависит от
расстояния между ними и от того, насколько часто
они разделяются кроссинговером.
145.
146.
Сцепление: локализация генов нахромосомах
• Первый ген человека был локализован на Ххромосоме в 1911 г., а первый аутосомный ген –
только в 1968 г. К середине 1970-х гг. было
локализовано менее 100 генов, значительная часть из
них располагалась в Х-хромосоме.
• Карта генетического сцепления у человека составляет
около 2809 сМ для мужчин и 4782 сМ для женщин.
Меньший "размер" мужского генома объясняется тем,
что частота рекомбинации в сперматогенезе меньше,
чем в оогенезе. Средняя длина генома человека в
единицах генетического расстояния составляет около
3300 сМ.
147.
Сцепление: локализация генов нахромосомах
•Методы картирования.
•Генетическое картирование - это картирование, основанное на
методах классической генетики - определении групп сцепления,
частоты рекомбинации и построении генетических карт, где единицей
измерения служат проценты рекомбинации, или сантиморганы (сМ).
•Цитогенетическое картирование – применение методов
цитогенетики, когда для локализации каких-либо нуклеотидных
последовательностей и определения их взаимного расположения
используются цитологические препараты.
•Физическое картирование - это обширная группа методов,
позволяющая строить карты генома (обычно их называют
физическими) высокого уровня разрешения и определять расстояния
между локализуемыми нуклеотидными последовательностями с
точностью от нескольких десятков тысяч п.н. до одной нуклеотидной
пары.
148.
Сцепление: локализация генов нахромосомах
гены
физическая карта с низким
уровнем разрешения
(сегменты диф. окраски)
упорядоченные маркеры
генетическая карта (карта
генетического сцепления)
перекрывающиеся клоны
рестрикционная карта
последовательность
нуклеотидов
149.
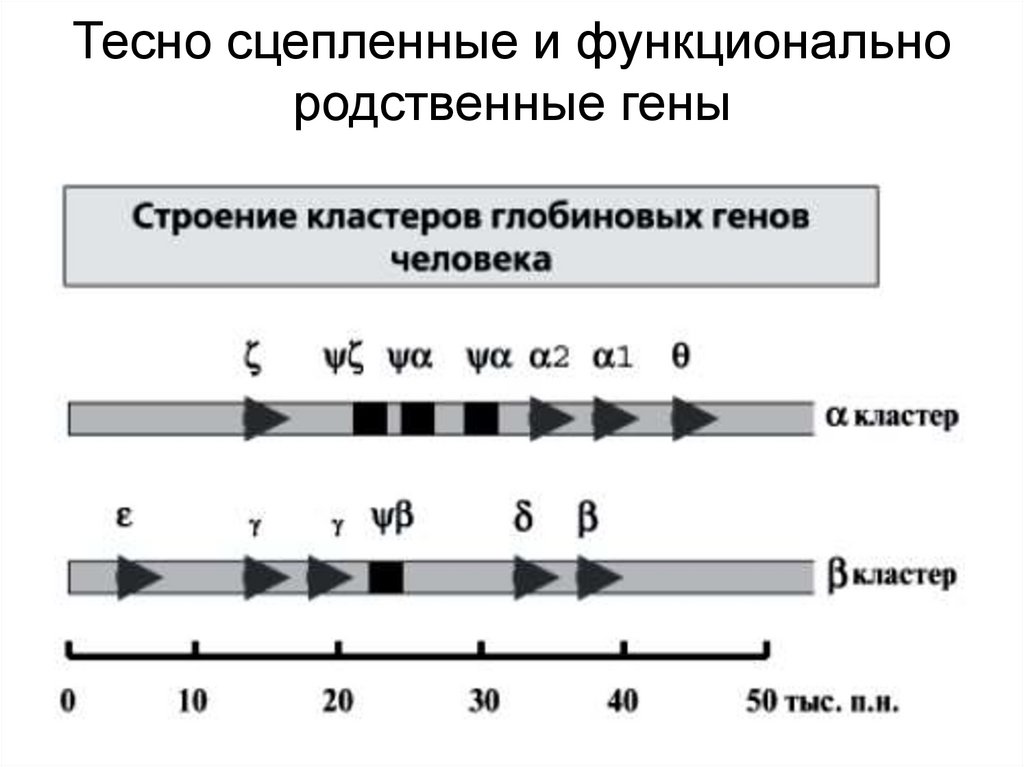
Тесно сцепленные и функциональнородственные гены
• Генные кластеры – результат эволюционного процесса. В
некоторых случаях кластеризация генов отражает историю
эволюционного развития.
• Допустим, на ранних этапах эволюции существовал один
локус, затем произошла дупликация гена и появилась
возможность функционального расхождения.
• Первая дупликация подготовила почву для последующих
дупликаций на основе механизма неравного кроссинговера
и, следовательно для дальнейшей функциональной
специализации. Такие дупликации в некоторых случаях
происходят не одноразово, а многократно. Сейчас стало
ясно, что дупликации играют очень важную роль в эволюции.
Подсчитано, что в геноме человека в сумме дуплицировано
около 3,6% нуклеотидных последовательностей размером в
1000 и более п.н.
150.
Тесно сцепленные и функциональнородственные гены
• Разные копии одного семейства генов могут располагаться в
геноме рядом и следовать друг за другом (это называют
тандемной дупликацией). Рассмотрим несколько примеров.
Так, гены, кодирующие рибосомную РНК (рРНК) и белки
гистоны, расположены в геноме человека в виде кластеров,
построенных из одних и тех же последовательностей,
следующих друг за другом.
151.
Тесно сцепленные и функциональнородственные гены
• Такие «гроздья» одинаковых генов обычно требуются
организму для того, чтобы нарабатывать большие количества
определенного продукта.
• рРНК составляет по массе основную часть всей клеточной
РНК, входя в состав таких важных «машин» клетки, как
рибосомы.
• Огромная масса гистонов нужна клетке для упаковки
гигантских молекул ДНК и плотной укладки их в хромосомы.
Во всех таких случаях говорят о большой «дозе» гена в
геноме, которая в конечном итоге и обеспечивает клетку
большим количеством продукта.
152.
Тесно сцепленные и функциональнородственные гены
• Однако в большинстве других случаев возникающие
в результате дупликации «копии» генов постепенно
претерпевают изменения (мутации), что приводит к
возникновению определенных различий между
ними, хотя формальное родство при этом
сохраняется.
• Такой процесс был назван дивергенцией, т.е.
расхождением. Дивергенцию обычно выражают в
процентах различий между двумя родственными
последовательностями нуклеотидов в ДНК или
последовательностями аминокислот в белках.
153.
Тесно сцепленные и функциональнородственные гены
• Псевдогены. В семействах генов встречаются иногда
такие гены, которые не утруждают себя никакой
работой. Нарушение их способности функционировать
в клетке обычно связано с накоплением опечаток в их
ДНКовом тексте. Такие гены получили название
псевдогенов. Перед их названием ставят греческую
букву ψ. Не совсем ясно, зачем геному нужны такие
гены, почему он сохранил их в эволюции, не
избавился от них.
154.
Тесно сцепленные и функциональнородственные гены
• По имеющимся оценкам, в геноме человека имеется
около 20 000 псевдогенов. В частности, в огромной
семействе генов–рецепторов обоняния из 1000 генов
около 60% являются псевдогенами. Считается, что
массивная потеря функциональных генов произошла
за последние 10 млрд. лет, и связано это со
снижением роли обоняния у человека по сравнению с
другими позвоночными организмами. Здесь, по–
видимому, действует принцип отмирания из–за
неупотребления.
155. Псевдогены
156.
Тесно сцепленные и функциональнородственные гены
• Установлено, что существует, по крайней мере, два механизма,
приводящих к образованию псевдогенов. Один из них
заключается в удвоении уже существующего гена
(дупликация), а потом дополнительная копия накапливает
мутации, которые препятствуют его работе.
• Другой механизм связан с процессом сплайсинга и обратной
транскрипции. На сформировавшейся мРНК синтезируется ее
ДНК–копия, которая в дальнейшем встраивается в геном.
Такие псевдогены не содержат ни интроны, ни промоторные
участки. Кроме того, в них со временем накапливаются
многочисленные мутации, не позволяющие этим генам
обеспечивать синтез нормального белка.
157.
Тесно сцепленные и функциональнородственные гены
158. Проявление аллеля в гемизиготе
46,XY46,XX
Здоровый
Больной
Носитель
46,XY 46,XY 45,X
46,XX 46,XX
159. Число аллелей
• 46,XY и 45,Х – гемизиготны, генотип А или а• 47,XXY – мужчины с 2 аллелями по Х
• 47,ХХХ – женщины с тремя аллелями по Х
• Множественный аллелизм:
– понятие популяционное
– конкретный человек имеет так же два аллеля
– усложняется вопрос доминирования
– нередко появляются компаунды, вроде аа1
160. Ограничения законов Менделя
• Один признак определяется только однимгеном (нет действия неаллельных генов)
• Только два (альтернативных) варианта
признака: больной-здоровый
• Гендер-независимое проявление признака
• Только два аллеля в популяции
• Взаимодействие аллелей – доминирование
• Проявление доминантного аллеля –
полная пенетрантность, высокая
экспрессивность
161. Проявление доминантных аллелей у гетерозигот
• Пенетрантность – непосредственнаячастота патологического фенотипа у Аа
– Варьирует от 40 % до 100 % (таблицы)
– Средняя величина – 80 %
• Проявление А у 100 % гетерозигот –
полная пенетрантность
• Экспрессивность – варьирование тяжести
заболевания в зависимости от генного
окружения и регуляции экспрессии гена
162. Взаимодействие аллельных генов
163. Типы аллельных взаимодействий
Доминанирование - проявление у гетерозиготтолько доминантного аллеля
Кодоминирование – совместное проявление двух
аллелей
Неполное доминирование – проявление
доминантного аллеля у гетерозигот в ослабленной
форме
Сверхдоминирование - доминантный ген в
гетерозиготном состоянии проявляется сильнее,
чем в гомозиготном.
164. Полное доминирование
165. Неполное доминирование (ген АТМ)
АААa
Здоровый
человек
Повышенный
риск
онкологических
заболеваний
aa
Синдром ЛуиБар
166. Взаимодействие неаллельных генов
167. Варианты взаимодействия
Комплементация - для формированияпризнака необходимо действие взаимодополняющих (комплементарных) генов
Эпистаз (супрессия) - подавление действия
одного гена (гипостатического) действием
другого (эпистатического или супрессора)
Полимерия - аддитивное действие генов
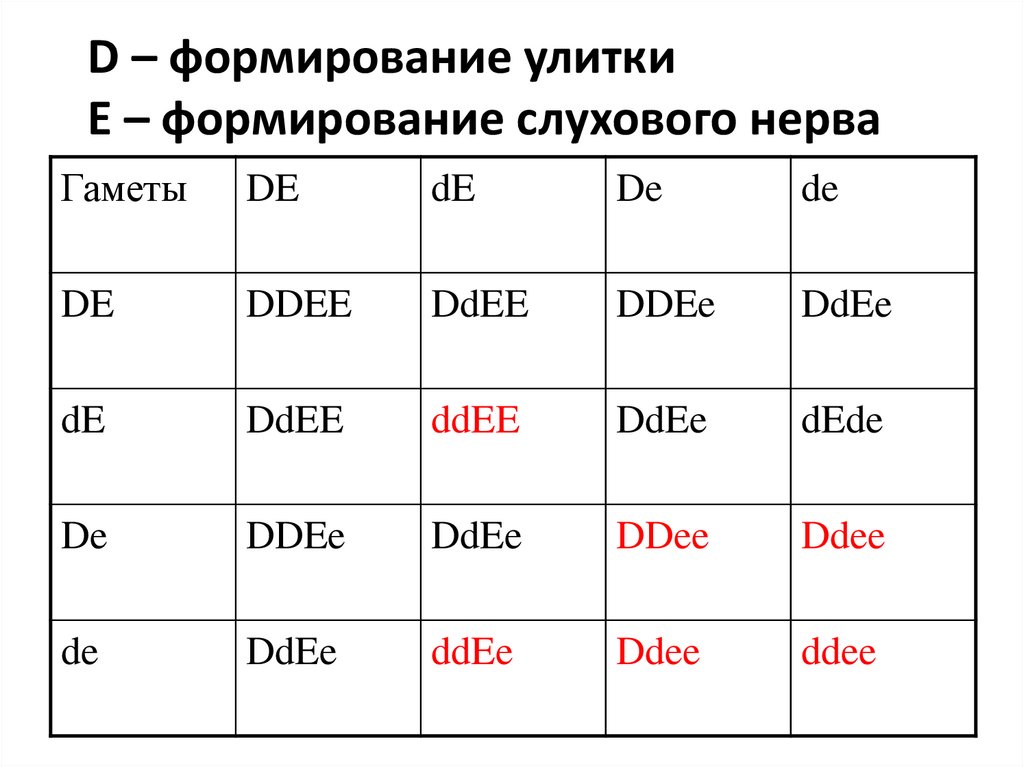
168. Комплементация
• Лат. complementum - средство пополнения,ситуация, когда для формирования признака
необходима работа нескольких генов
• Модель – груз, подвешенный на цепи. Разрыв
любого звена – падение груза (заболевание)
• У человека нормальный слух обусловлен в том
числе, двумя доминантными неаллельными
генами D и E. Ген D - определяет развитие улитки,
а ген Е- развитие слухового нерва
• Гомозигота DDЕЕ и гетерозиготы DdЕе - имеют
нормальный слух; рецессивные гомозиготы даже
по одному из этих генов (ddЕЕ или DDее) - глухие
169.
D – формирование улиткиE – формирование слухового нерва
Гаметы
DE
dE
De
de
DE
DDEE
DdEE
DDEe
DdEe
dE
DdEE
ddEE
DdEe
dEde
De
DDEe
DdEe
DDee
Ddee
de
DdEe
ddEe
Ddee
ddee
170. Эпистаз
• Подавление действия одного гена действиемдругого гена. Подавляющий ген называется
геном - супрессором, или ингибитором, а
подавляемый - гипостатическим
• Похож на доминирование для аллельных генов.
Однако здесь аллель одного гена подавляет
проявление аллеля из другой аллеломорфной
пары, т. е. неаллельного гена.
• Бывает доминантным и рецессивным
171. Виды эпистаза
Доминантный –подавление
доминантным аллелем
эпистатического (А) гена
действия любых аллелей
гипостатического (В) гена
AА или Аа > ВВ, Вb, bb
Рецессивный –
подавление
рецессивным аллелем
эпистатического (А) гена
действия любых аллелей
гипостатического (В) гена
аа > ВВ, Вb, bb
172. Полимерия
• Также аддитивное взаимодействие генов –степень развития количественного признака
определяется влиянием нескольких
полимерных генов
• Модель – стопка матов. Выдёргивание одного
или нескольких матов уменьшает признак,
чтобы он исчез, нужно убрать все маты
• Полимерные гены обозначают одинаковыми
буквами, чтобы подчеркнуть их одинаковую
направленность
173. Полимерия: цвет кожи
174. Альбинизм как вариант эпистаза?
175. Взаимодействие генотипа и среды в формировании признаков
176. Среда
ГенетикаФизика
Химия
Биология
Соц.-Псих.
Хромосомы
Гены
Генные сети
прочее
177. Распространённость
• Мультифакторные болезни – от процентовдо долей процента
• Хромосомные болезни – от 1:600 до
единичных случаев в мире
• Моногенные болезни – от 1:1500 до
единичных случаев в мире
178. Устаревший постулат
• Один ген –• Один белок –
• Один признак
NB! Плейотропное действие гена
NB! Один ген – разные синдромы
179. Обратная ситуация
• Генокопии (биол.) – одинаковое проявлениемутаций разных генов
• Генетическая гетерогенность (мед.) –
свойство генетических заболеваний, при
котором они вызываются мутациями разных
генов
NB! Причина разных типов наследования
болезни в разных семьях
NB! Сложности молекулярно-генетической
диагностики
180. Синдром Барде-Бидла
• OMIM 209900 Гены 17q23, 16q21, 15q22.3q23, 14q32.1, 12q21.3, 12q21.2, 11q13, 9q31q34.1, 8q21.13-q22.1, 7p14, 4q27, 4q27,3p12-q13, 2q31, 20p12
181. Отосклероз
• MIM 166800 + ещё 6 АД форм• Пенетрантность 40 %
• Проявляется после 20 лет
• Нередко mutacio de novo
182. Общий итог (ген-болезнь)
• Клинический полиморфизм:– Варьирует возраст начала
– Варьируют диагностические признаки
– Варьируют дополнительные признаки
– Варьирует тяжесть проявлений
– Варьирует эффективность разных способов
лечения
– Варьирует продолжительность жизни
183. CFTR 7q31
МуковисцидозMIM 602421
CFTR
7q31
184. Напоследок
• Фенокопия – случай патологическогосостояния у человека, имитирующий
генетическое заболевание при
достоверном исключении генетического
дефекта
185. Популяционная генетика
186. Популяционная генетика
• Раздел генетики, изучающий распределениечастот аллелей и их изменение под влиянием
различных процессов
• Основа стабильности – равновесие ХардиВайнберга
• Сдвиг осуществляется за счёт:
– Мутагенеза
– Естественного отбора
– Инбридинга
– Дрейфа генов
– Потока генов
187. Две ветви эволюционной теории
• традиционная популяционная генетика– оперирует наборами генотипов
• биометрическая теория
– оперирует наборами фенотипов изучаемых
объектов
– используется в селекции растений и
животных
188. Этапы развития генетики популяций
• Вторая половина 20-х — конец 30-х годов XX века. В это времяпроисходило накопление данных о генетической гетерогенности
популяций. Он завершился выработкой представлений о
полиморфизме популяций.
• 40-е -середина 60-х годов XX века. Изучение механизмов поддержания
генетического полиморфизма популяций. Появление и развитие
представлений о важной роли гетерозиса в формировании
генетического полиморфизма.
• Вторая половина 60-х — конец 1970-х годов XX века. Этот этап
характеризуется широким применением белкового электрофореза для
изучения полиморфизма популяций. Формируются представления о
нейтральном характере эволюции.
• С конца 1970-х годов. Этот период характеризуется методическим
смещением в сторону применения ДНК-технологий для изучения
особенностей процессов происходящих в популяциях. Важным
моментом этого этапа (примерно с начала 1990-х годов) является
широкое применение вычислительной техники и специализированных
программ (например, PHYLIP, Clustal, Popgene) для анализа
разнообразных типов генетических данных.
189. Закон Харди — Вайнберга
• Частоты генотипов по какому-либо гену (в случае если впопуляции есть два аллеля этого гена) будут
поддерживаться постоянными из поколения в
поколение и соответствовать уравнению:
P^2 + 2pq + q^2 = 1,
где p^2 – доля гомозигот по 1 аллелю,
q^2 - доля гомозигот по альтернативному аллелю,
2pq - доля гетерозигот
• Это закон, действующий в абсолютной популяции:
– бесконечно большого размера,
– в которой не действует отбор,
– не идет мутационный процесс,
– отсутствует обмен особями с другими популяциями,
– не происходит дрейф генов,
– все скрещивания случайны
190. Закон Харди — Вайнберга для двух аллелей: по оси абсцисс показаны частоты аллелей p и q, по оси ординат — частоты генотипов.
Каждая кривая соответствует одному из трех возможных генотипов.191. Значение закона Харди — Вайнберга
• В медицинской генетике закон Харди — Вайнбергапозволяет оценить популяционный риск генетически
обусловленных заболеваний
• Каждая популяция обладает собственным
аллелофондом и, соответственно, разными частотами
неблагоприятных аллелей
• Зная частоты рождения детей с наследственными
заболеваниями, можно рассчитать структуру
аллелофонда
• Зная частоты неблагоприятных аллелей, можно
предсказать встречаемость болезни у новорожденных
192. Применение для аутосомно-рецессивных синдромов
Применение для аутосомнорецессивных синдромов• Расчёт на основе заболеваемости
• Гетерозиготы в 2 раза чаще
• Частота доминантного аллеля почти 1
• Один рецессивный требует вычисления
квадратного корня
193. Применение закона для аутосомно-доминантных синдромов
Применение закона для аутосомнодоминантных синдромов• Семейная гиперхолестеринемия (СГ) –
аутосомно-доминантное заболевание
человека.
• Типичным осложнением СГ являются
атеросклероз коронарных сосудов и
инфаркты миокарда.
• Частота гетерозиготной формы СГ в
большинстве популяций 1:500,
гомозиготной -1:1000000.