




























 biology
biologySimilar presentations:


")
")
")

")

")

Генетика популяцій
1. Генетика популяцій
1.2.
3.
4.
5.
6.
7.
План
Популяція — одиниця еволюційного процесу
Частоти генотипів та частоти алелів
Закон Харді — Вайнберга
Проблема генетичної гетерогенності природних
популяцій
Методи вивчення структури популяцій
Елементарна еволюційна подія — зміна частот
алелей у популяції
Добір, мутаційний процес та ізоляція
2. Популяція — одиниця еволюційного процесу
Біологічна еволюція — це процес змін і дивергенції біологічних форм у плинічасу. Елементарною одиницею біологічної еволюції є популяція. Такою
одиницею не може бути окремий організм, бо він представляє лише одне
покоління і не може спадково змінюватись у часі, який в еволюції вимірюється
багатьма поколіннями. Індивідуальні варіації, навіть спадкові, можуть не
виявлятись у даної особини і навіть у цілого покоління у відповідності з
домінантно-рецесивними співвідношеннями алелів генів та з інших причин. Види
також не можуть бути елементарними еволюційними структурами, бо вони
нерівномірно розподілені у просторі, найчастіше у вигляді локальних популяцій,
кожна з яких має свої генетичні особливості і еволюціонує на свій лад. Таким
чином, лише популяція відповідає всім вимогам елементарної одиниці
еволюційного процесу, і генетичні зміни, що в ній відбуваються протягом
поколінь, складають так звану мікроеволюцІю.
Будь-яку спадкову зміну в популяції називають елементарною еволюційною
подією.
Популяцією називають певним способом відокремлену від інших сукупність
особин одного виду, яких об'єднує спільність походження, місця існування, а
також механізмів розмноження і адаптації. Отже, популяція — це група
організмів одного виду, що заселяє певну територію і розмножується
ізольовано від інших популяцій того ж виду.
3.
Генетики користуються поняттями ідеальних і реальних популяцій.Ідеальна популяція складається із безмежної кількості особин, між
якими можливі вільні випадкові схрещування без будь-яких
обмежень, тобто в такій популяції здійснюється панміксія. Така
популяція повністю ізольована і на неї не діють ніякі чинники, що
можуть змінювати її структуру. Ідеальних популяцій у природі
немає.
Природні популяції називають реальними. Саме вони і є об'єктом
безпосереднього вивчення як генетиками, так і іншими
спеціалістами Ці популяції в залежності від способу розмноження
особин поділяють на три типи:
1) популяції самозапильних рослин і автогамних тварин;
2) популяції перехреснозапильних рослин і алогамних тварин;
3) популяції" форм, що розмножуються вегетативно (апогамне
розмноження).
Популяції, що розмножуються статевим способом і в межах
яких здійснюються вільні випадкові схрещування, називаються
панміктичними або менделівськими.
4. Частоти генотипів та частоти алелів
Головна відмінність методології популяційної генетики від уже звичноїметодології генетичного аналізу полягає в тому, що вона має справу не із
чистими лініями і індивідуальними схрещуваннями, а зі успадкуванням у
більших сукупностях організмів, гетерогенних по своєму генетичному
складу. Вся сукупність генів популяції називається її генофондом і
визначається як 2N, де N — число особин. Таким чином, у кожному
розглянутому локусі є 2N генів, якщо мова йде про популяцію диплоїдних
організмів. Це ствердження справедливо для всіх генів, крім тих, які
перебувають у Х-хромосомі при гетерогаменості однієї із статей.
Найважливішою характеристикою популяції є частоти аллелей (генів) і
генотипів. Генофонд популяції втілюється в значеннях частот
генотипів, які визначаються на репрезентативних (досить великих)
вибірках, які повинні робитися випадково для виключення суб'єктивних
помилок експериментатора.
Використовують також показник, пов'язаний із частотою генотипів,
— частоту аллелей. Ця величина складається із частоти гомозигот по
даної аллели й половини частоти гетерозигот.
5. Закон Харді — Вайнберга
Закон Харді-Вайнберга свідчить про те, що спадкування як таке не міняє частоти алелей упопуляції.
Якщо позначити частоту алелі А через р, а частоту алелі а через q, то при наявності по
даному локусі тільки двох алелей у популяції: рА + qа = 1. Співвідношення генотипів у
такому випадку буде: (рА + qa)2 = р2AА + 2pqAa+q2aa = 1, у чому легко переконатися,
скориставшись решіткою Пеннета:
pA
qa
pA
p2AA
pqAa
qa
pqAa
q2aa
Якщо в популяції для даного гена присутні три алелі із частотами р, q і r, то частоти
генотипів також відповідають формулі біноміального розподілу:
(p + q + r)2 = р2 + q2 + r2 + 2pq + 2pr + 2qr=1
і т.д. при більшому числі алелей.
Представимо, що алелі А и а зустрічаються із частотами 0,5, тоді в F1 частоти генотипів
будуть:
0,5 A
0,5 a
0,5 A
0,25 AA
0,25 Aa
0,5 a
0,25 Aa
0,25 aa
Таким чином, 0,25AA + 0,50Aa + 0,25aа =1.
6.
Неважко побачити, що при цьому зберігається вихідна частота алелей: 0,5А и 0,5а.У наступному поколінні одержують те ж з відношення генотипів, як і в наступних
поколіннях. Користуючись цим прийомом і приймаючи інші значення p і q, легко
переконатися, що вже в першому поколінні в популяції встановлюється рівновага,
що зберігається й у всіх наступних. Ця закономірність була очевидна для
математика X. Харді, і він хотів, щоб вона стала відома біологам, що вважали, що
частота домінантної алелі в змішаній популяції може автоматично зростати.
У точному значенні закон Харді — Вайнберга відноситься до нескінченно більших
популяцій, у яких здійснюється панміксія й на які не діють ніякі зовнішні
фактори. Тільки при цих умовах популяція перебуває в рівновазі, тобто частоти
алелей і генотипів у ній постійні. Очевидно, що це ідеальне подання про
популяцію ніколи не реалізується в природних умовах. На популяцію постійно
діють численні зовнішні й внутрішні фактори, що порушують генетичну рівновагу.
Тому що ж визначити частоту аллелей у популяції при повнім домінуванні?
Оскільки, відповідно до формули Харді-Вайнберга, (рА + qa)2 = р2AА + 2pqAa+q2aa
= 1, то, знаючи частоту рецесивних гомозигот, досить обрахувати квадратний
корінь із отриманої величини, і ми знайдемо частоту рецесивної алелі q. Частота
домінантної алелі складе р = 1 — q. Визначивши в такий спосіб частоти алелей A і
а, можна з'ясувати частоти всіх генотипів у популяції.
Важливим наслідком закону Харді-Вайнберга є те, що рідкі алелі в популяції
присутні переважно в гетерозиготі. Наприклад, якщо алель а зустрічається із
частотою 0,01, а алель
А — відповідно із частотою 0,99, то частота гетерозигот складе 0,0198, тобто
близько 0,02, а рецесивних гомозигот — 0,0001. Таким чином, у гетерозиготному
стані рецесивна алель зустрічається більш ніж в 100 разів частіше, ніж у
гомозиготному.
7.
Модель Харди-Вайнберга, показує, що відбувається з алелем і згенотипом в «ідеальній» популяції (позбавленої всіх складних
особливостей, властивих реальним популяціям), якщо прийняти
ряд простих допущень:
1.
2.
3.
4.
5.
Особини з будь-яким генотипом мають однакову частоту
виживання й однаковим репродуктивним успіхом — іншими
словами, на популяцію не діє добір.
Нові алелі не виникають, а наявні алелі не перетворюються друг у
друга в результаті мутацій.
Не відбувається міграції особин ні в популяцію, ні з популяції.
Популяція нескінченно велика (тобто на практиці, досить велика,
щоб можна було зневажити помилками вибірки й інших
випадкових ефектів).
Особини в популяції схрещуються випадковим чином.
Відповідно до закону Харді-Вайнберга, «ідеальна» популяція (для
якої виконуються перераховані умови) має наступні властивості:
1.
2.
Частота алелей не змінюється з покоління в покоління (іншими
словами, популяція не еволюціонує).
Після одного покоління випадкового схрещування частоти
генотипів можна пророчити виходячи з вихідних частот алелей.
8. Застосування закону Харди-Вайнберга: розрахунок частоти гетерозигот
Одне із застосувань закону Харді-Вайнберга — оцінка частоти гетерозигот у популяції. Частоту рецесивногоознаки звичайно можна визначити, підрахувавши особин, що володіють цією ознакою, у вибірці особин з
досліджуваної популяції. За допомогою цієї інформації й закону Харді- Вайнберга можна розрахувати частоти
алелей і генотипів.
Муковісцидоз, аутосомна рецесивна ознака, зустрічається із частотою близько 1/2500 = 0,0004 у людей, що
походять із північноєвропейських популяцій. Людей, що страждають муковісцидозом, легко розпізнати по
таких симптомах як солоний піт, надмірна кількість слизу в легенях і сприйнятливість до бактеріальних
інфекцій. Тому що це рецесивна ознака, хворі люди повинні бути гомозиготами. Їхня частота в популяції
становить q2, якщо в попередніх поколіннях схрещування було випадковим.
Таким чином, частота рецессивного аллеля становить
q q 2 0, 0004 0, 02
Так як р + q = 1, то частота домінантного алеля дорівнює
p=1 – q = 1 - 0,02 = 0,98.
За рівнянням Харді-Вайнберга частота гетерозігот визначається як 2рq.
2pq = 2 * 0,98 * 0,02 = 0,04 = 4% або 1/25.
У підсумку, гетерозиготи по гену муковісцидозу зустрічаються в популяціях порівняно часто (4%),
незважаючи на те, що рецесивні гомозиготи зустрічаються дуже рідко (1/2500 або 0,04%).
У цілому, частоти всіх трьох генотипів можна оцінити, визначивши частоту кожного з алелей і допустивши
виконання умов Харди-Вайнберга.
9. Співвідношення частот алелей і генотипів, що випливає з рівняння Харді-Вайнберга
10. Проблема генетичної гетерогенності природних популяцій
Значення генетичної гетерогенності природних популяцій упершепо достоїнству оцінив С. С. Четвериков у своїй класичній роботі
«Про деякі моменти еволюційного процесу, з погляду сучасної
генетики» (1926). С. С. Четвериков передбачив подальший
розвиток популяційно-генетичних досліджень, пророчивши, що
природні популяції повинні бути у високому ступені генетично
гетерогенні, що «вид, як губка, усмоктує в себе гетерозиготні
геновариації (так С. С. Четвериков називав мутації), сам
залишаючись
при
цьому увесь час зовні (фенотипово)
однорідним» . Дійсно, більшість виникаючих мутацій рецесивні і
при високій чисельності популяції ці рецесивні мутації як би
розчиняються, виявляючись у гетерозиготі. При цьому ймовірність
їх гомозиготизації обернено пропорційна чисельності загальної
сукупності організмів.
11.
В 40— 50-х роках розглядалися в основному дві точки зору на структуруприродних популяцій: класична модель і балансова модель. Відповідно до
класичної моделі природні популяції представлені в основному
гомозиготами по домінантним алелям. Частота домінантних алелей близька
до одиниці за винятком незначної частки шкідливих рецесивних мутацій. Із
цього погляду еволюційне зрушення в популяції ґрунтується на доборі рідких
сприятливих алелей.
Відповідно до балансової моделі популяції віддавалася перевага
поліморфізму адаптивного «дикого» типу, тобто для кожного гена не існує
однієї алелі дикого типу. Скоріше більшість, якщо не всі локуси, може бути
представлено серіями алелей з різними частотами в популяції. Таким чином,
не існує якогось стандартного «дикого» типу. Балансова модель виходила
також із широкого поширення в природних популяціях ефекту гетерозису,
тобто переваги гетерозигот у порівнянні з гомозиготами по тимі ж алелям. У
цьому випадку еволюційне зрушення в популяції ґрунтується на доборі не по
одному гені, а по багатьом генам, алелі яким перебувають у балансі один з
одним. При цьому оптимальне для адаптації вираження кожної алелі
коадаптовано з іншими генами і їх алелями.
Обидві моделі популяційної структури ґрунтувалися на уявленні про те, що
більшість знову виникаючих мутацій шкідливі.
12. Методи вивчення структури популяцій
Одним із методів, який широко використовується в популяційній генетиці, єелектрофоретичний розподіл (електрофорез) білків, нуклеїнових кислот та їх
фрагментів у поліакриламідному, крохмальному або іншому гелі. Ідея цього
методичного підходу полягає в тому, що за наявності в генотипі мутантного
гена змінюється структура відповідного фрагмента ДНК І відповідних генних
продуктів, внаслідок чого змінюються і їх фізико-хімічні властивості.
Якщо досліджуваний фермент - мономер, тобто не має четвертинної
структури, то в гетерозиготі по ізозімам виявляються дві смуги - два
електрофоретичних варіанти. Якщо фермент складається із двох ідентичних
субодиниць, то електрофоретичних варіантів буде три, з огляду на вільну
комбінаторику субодиниць. При більшому числі субодиниць відповідно
збільшується й число смуг при електрофорезі. Зустрічаються й більше складні
випадки, коли білки побудовані з ідентичних і неідентичних субодиниць,
тобто контролюються двома різними генами, наприклад гемоглобін αα ββ.
13.
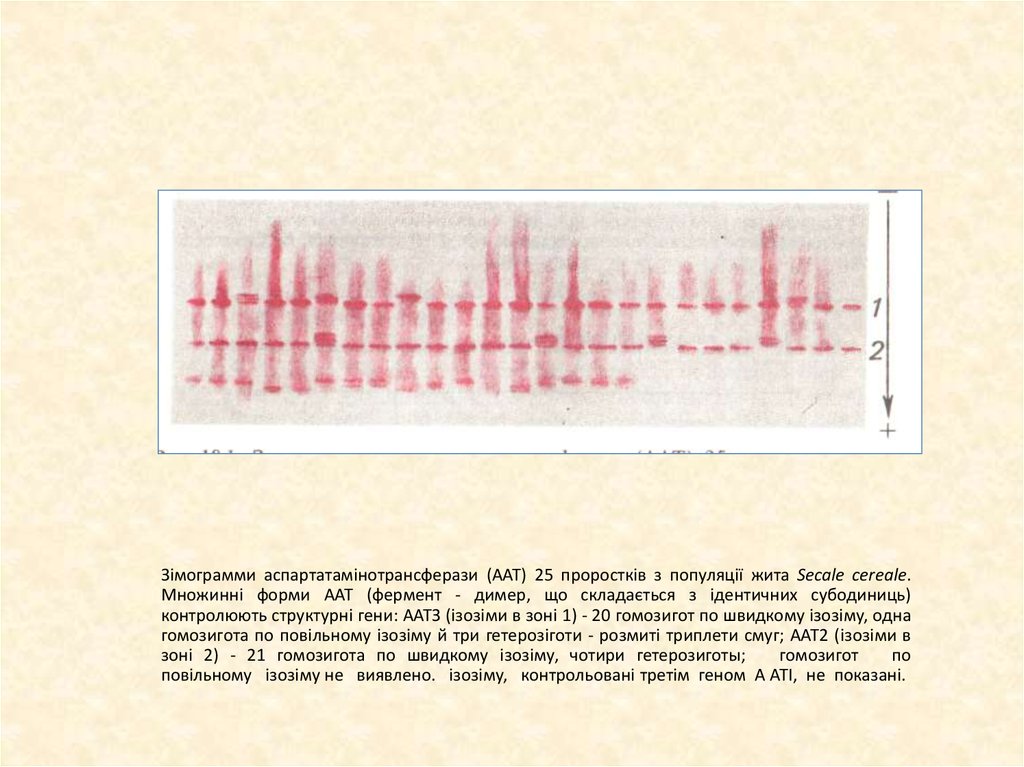
Зімограмми аспартатамінотрансферази (ААТ) 25 проростків з популяції жита Secale cereale.Множинні форми ААТ (фермент - димер, що складається з ідентичних субодиниць)
контролюють структурні гени: ААТЗ (ізозіми в зоні 1) - 20 гомозигот по швидкому ізозіму, одна
гомозигота по повільному ізозіму й три гетерозіготи - розмиті триплети смуг; ААТ2 (ізозіми в
зоні 2) - 21 гомозигота по швидкому ізозіму, чотири гетерозиготы;
гомозигот
по
повільному ізозіму не виявлено. ізозіму, контрольовані третім геном A ATI, не показані.
14.
Поліморфізм (Р) популяції оцінюється часткою поліморфнихлокусів із всіх досліджених. Так, з 30 локусів, вивчених у
каліфорнійського морського хробака Phoronopsis viridis, в 18
виявлені алельні варіанти, а в 12 варіацій не зареєстровано. У
цьому випадку: Р= 18:30= 0,60.
На матеріалі декількох популяцій одного виду можна обчислити
середній поліморфізм. Наприклад, величина Р була визначена
ще для трьох популяцій P. viridis і становила відповідно 0,50;
0,53; 0,47. Тоді середній поліморфізм буде оцінюватися як
0, 60 0,50 0,53 0, 47
Pср
0,525
4
Гетерозиготність (Н) по даному локусі визначається як
відношення гетерозигот до загального числа досліджених
особин популяції. На основі даних по окремих локусах
визначають середню гетерозиготність популяції Нср.
15.
Оскільки популяції, безсумнівно, розрізняються по ступені панмиксії (наприклад,перехреснозапильники і самозапильники), а також по селективній цінності
гетерозигот, то і реальна гетерозиготність у них буде різною, навіть при однакових
частотах алелей. З урахуванням цієї обставини обчислюють так звану очікувану
гетерозиготність на основі умовного припущення, що в популяціях завжди
здійснюється повна панміксия. Тоді при наявності чотирьох алелей одного гена із
частотами f1, f2, f3, f4 частота їх гомозигот очікується як f12, f22, f32, f42 а очікувана
гетерозиготность буде становити:
Hex = 1 – (f12, f22, f32, f42 )
Середній поліморфізм (Р) і середня гетерозиготність (Н) визначені для багатьох видів.
Безхребетні в середньому більш генетично мінливі, ніж хребетні. Hср= 13,4% для
перших і 6,0 % для других. Рослини перехреснозапильники значно мінливіші, чим
самозапильники (Hср = відповідно 19 % і 6%). По даним електрофорезу, середня
гетерозиготність людських популяцій 6,7 %. Цікаво відмітити, що означає ця величина
для людини. Приймемо, що геном людини включає 30 000 структурних генів, і ця
величина, мабуть, занижена. Тоді деякий середній індивідуум є гетерозиготним не
менш чим по 30 000 X 0,067 = 2010 генам. Отже, потенційно така полиігетерозигота
могла б утворити 22010, або близько 1065 різних типів гамет. Ця потенційна
комбінативна мінливість ніколи не реалізується ні в окремої людини, ні у всього
людства. Для порівняння скажемо, що за оцінкою фізиків число протонів і нейтронів у
Всесвіті становить близько 1076.
16.
17. Елементарна еволюційна подія — зміна частот алелей у популяції
Зміна й дивергенція біологічних форм у часі ґрунтуються на двохголовних явищах: мінливості і зміні частот алелей і генотипів.
Зміна частот алелей і генотипів у популяції становить сутність
елементарної еволюційної події.
Сам по собі добір у чистих лініях, тобто під час відсутності
генетичної гетерогенності, не створює нових форм. Разом з тим і
спадкування в гетерогенній популяції саме по собі не змінює
частот алелей. До зміни частот алелей і генотипів у генетично
гетерогенній популяції приводить добір. Зміна частот алелей і
генотипів можливо не тільки внаслідок добору, але й у результаті
мутацій, міграції особин, випадкового дрейфу генів, ізоляції, а
також виборчого, або асортативного, схрещування. Всі ці фактори,
що діють у популяціях, називають факторами динаміки
популяцій.
18. Внутривидова дівергенція або утворення видів
Процес видоутворення, що пов’язаний із просторовим розєднанням популяції єалопатричним (алос – різний ) видоутворенням коли нові форми утвоюються без розєднаяя
це називається симпатричним видоутворенням.
Наприклад екологічна спеціалізація в звяку із просторовим розєднанням може утворити
колові види. Наприклад ареали сріблястої чайки та чайкі-клуші західної Європи: чайки (їх
географічні раси) розселялися на захід та схід. Це посилювалось спеціалізацією географічних
рас. У них відбувалися різні інверсії, транслокації та ін. І коли вони знову зустрілися це вже
були нові біологічні види, які не давали фертильних нащадків. У оз. Байкал, де мешкають
більш 300 видів гаммарусів-бокоплавів, але вони не зхрещуюютья, тому що вони існують на
різній глибині та харчуються різною їжею, тому мають різну будову ротового апарату і різну
фізіологіні особливості травлення. Наприкінці це призводить до того, що така морфологічна
та фізіологічна ізоляція закінчується генетичною ізоляцією. Спарювання між ними не
можливо внаслідок різної будови статевого апарату. Таким чином відбувається сипатричне
видоутворення, коли початкові морфологічні та фізіологічні особливості призводять до
генетичної ізоляції.
19. Добір
Генетична гетерогенність, широко розповсюджена в природних популяціях, становитьоснову ефективності дарвінівського природного добору.
Пряму кореляцію між ступенем гетерогенності популяції й швидкістю еволюційної зміни
внаслідок природного добору математично обґрунтував Р. А. Фішер (1930) у своїй основній
теоремі природного добору: швидкість збільшення пристосованості якої-небудь
популяції в будь-який відрізок часу дорівнює її генетичної мінливості по
пристосованості в цей же час. У цьому випадку під пристосованістю розуміють відносну
швидкість відтворення.
Природний добір діє
на різні групи організмів у популяції залежно від їхньої
пристосованості (W). Порівнюючи пристосованість декількох груп особин, найбільшу
приймають за одиницю, а пристосованість інших груп виражають у частках одиниці.
Наприклад, якщо пристосованість для гомозигот АА й гетерозигот Аа дорівнює 1, а для
гомозигот аа — 0,9, то інтенсивність природного добору, або коефіцієнт добору,
обчислюється як
S = WAA — Waa= 1 — 0,9 = 0,1.
Тоді в популяції диплоїдных організмів за умови повного домінування можна розрахувати
частоти алелей у наступному поколінні після початку дії добору.
20.
Частота алелі А у F1 після добору буде:p02 p0 q0
p1
1 Sq02
Зміна частоти алелі А за покоління буде складати:
p02 p0 q0
Sp0 q02
p p1 p0
po
2
1 Sq0
1 Sq02
При малих значеннях Sq2
p
Spq 2
Наприклад, якщо р0 = 0,9, а q0 = 0,1 тоді при S = 0,1
p 0,1 0,9 0,12 0, 0009;
p1 0,9009
21. При крайніх значеннях частот алелей добір діє найменш ефективно. Найбільше ефективним є добір при середніх значеннях р и q. Цю закономірніс
При крайніх значеннях частот алелей добір діє найменш ефективно. Найбільше ефективним єдобір при середніх значеннях р и q. Цю закономірність ілюструють криві, що показують зміни
частоти алелі а(Δq) при доборі проти рецесивных гомозигот залежно від величини q, а також
проти
однієї
з
алелей
під
час
відсутності
домінування.
22.
Результати дії стабілізуючого, спрямованого й дизруптивного добору. Укожному з показаних випадків х0 - середнє арифметичне значення ознаки
для вихідної популяції (чорні криві), a xs - середнє значення ознаки в
популяціях, на яких діяв добір (червоні криві).
23.
Як відзначалося, добір діє на фенотипи, а непрямопозначається на зміні частот алелей. Залежно від того який
вплив робить добір на ознаки, розрізняють три типи
добору:
1) стабілізуючому, сприятливому збереженню середнього значення
ознаки
(теорія
стабілізуючого
добору
розроблена.
І.І.
Шмальгаузеном);
2) дизруптивний, або такий що розсікає, сприятливої стабілізації
крайніх значень ознаки;
3)
спрямований, або рушійний, сприятливий безперервній зміні
ознаки в певному напрямку.
Д. К. Бєляєв увів також поняття «дестабілізуючий добір». Цим
терміном він позначив підвищення розмаїтості в популяції
при різкій зміні напрямку добору, зокрема, у популяціях
тварин при їхньому одомашнюванні.
24. Мутаційний процес
Мутаційний процес — основа виникнення гетерогенності популяції. Черезнаявність мутаційного процесу важко говорити про існування істинно чистих —
гомозиготних ліній — протягом тривалого часу. Проте можна представити, що
існує абсолютно гомозиготна (АА) сукупність (р = 1), у якій відбувається
мутаційний процес А а із частотою u на гамету за одне покоління. Тоді в
наступному поколінні алелі А и а будуть зустрічатися із частотами: р = 1 — і й q =
і. Однак відбуваються не тільки прямі мутації, але й зворотні із частотою v, а в
популяції є алелі як А, так і а із частотами відповідно р и q. Частина алелей а буде
перетворюватися в А із частотою v. Тоді зміна частоти алелі А під впливом
мутаційного тиску за одне покоління можна виразити як Δр = vq — up. Частоти
алелей можуть змінюватися за рахунок мутаційного процесу тільки доти, поки vq
не стане рівним up. У такій ситуації наступає стан рівноваги. Значення р, при
якому наступає рівновага, можна знайти з вираження vq — up (стан рівноваги).
Перетворивши це рівняння, одержимо ир= v(1 - р), оскільки р + q = 1. Тоді p(u +
v) = v, а р = v/(u + v). Точно так само й q = і/ (і + v).
Якщо знову виникаюча мутація має селективну перевагу в 1 %, то ймовірність її
збереження дорівнює приблизно 2 %. Таким чином, сам по собі мутаційний
процес, мутаційний тиск недостатні для поширення рецесивної мутації.
25. Потік генів
Потік генів, або міграції особин, являє собою обмін генами між популяціями. Якщопопуляції мають різні частоти алелей, то міграція може приводити до зміни частот алелей,
що привносяться особинами — іммігрантами. Безпосередні результати такої події подібні з
наслідками виникнення мутацій, однак міграція змінює частоти алелей значно швидше, ніж
мутацій. Для обчислення ефекту міграції використовують ті ж рівняння, що й для
мутаційного процесу. Вплив потоку генів на динаміку популяцій тих або інших організмів
залежить від швидкості поширення гамет і відстані між локальними популяціями. Одні
види, наприклад людина, дуже рухливі, інші, наприклад деякі метелики, дуже обмежені у
своїх міграціях.
Міграції. Види нерідко бувають розділені на популяції, у тім або іншому ступені ізольовані
географічно. Різні фактори еволюції, у тому числі природний добір, можуть приводити до
розходження частот алелей у таких популяціях. Переміщення особин з однієї популяції в
іншу називають міграціями. Уявимо собі вид, у якого в деякому локусі є два алелі, А и а. Є
дві популяції даного виду, одна на континенті, а одна на острові. Частота алеля А на
континенті становить рm , а частота цього алелі на острові становить рi . Особини мігрують із
континенту на острів, і в наступному поколінні частота алелі А на острові складе
pi’= (1 – m)pi + mpm
де m — швидкість міграції, тобто частка особин, що мігрували, з континентальної популяції в
даному поколінні острівної популяції.
26. Дрейф генів, або генетико-автоматичні процеси, які впливають на частоти алелей у популяціях
Дрейф генів (або просто дрейф) — це абсолютно випадковий генетичноавтоматичний процес, за якого частота того чи іншого алеля в популяції різкозменшується або, навпаки, дуже зростає. Як приклад розглянемо дуже маленьку
панміктичну популяцію, в якій генотипи представлені у співвідношенні 1АА + 2Аа
= 1аа, тобто у відповідності з формулою Харді—Вайнберга.
Неважко переконатися, що Ймовірність злучення однакових гомозигот (АА х АА
або аа х аа) у даному випадку складає 1/16. Однак, із-за тієї чи іншої випадковості
це злучення може і не відбутися. Вірогідність відсутності такого злучення в
розрахунку на існуючі два класи гомозигот складе 1/16 +1/16 - 1/8 на одне
покоління. Отже, можна чекати, то за випадкового незлучення гомозигот певного
класу всього через вісім поколінь у популяції зовсім зникне відповідний алель (А
або а). Цей процес зміни частот генів у малочисельних популяціях, зобов'язаний
випадковому злученню пар за розмноження, називають дрейфом генів або генетичним дрейфом.
27.
Іноді, особливо в ізолятах чисельність популяції може бути дуже низькою. Зазвичай унормальних популяціях бувають періоди мінімальної та максимальної чисельності — т.з.
“хвилі життя”. Крім того може мати місце неріномірний розподіл особин у популяції. Тому
іноді генотип кожного наступного покоління формується вибіркою особин батьківського
покоління. Він має мінливість і похибку виборки. Якщо вибірка мала, то похибка велика, чим
менша вибірка, тим більше коливання частот алелів. Це можуть бути випадкові зміни, які не
є наслідком добору. Ці випадкові зміни можуть привести до елімінації алелів, яка
обумовлює найбільше пристосування, та навпаки, може бути закріплена алель, яка
обумовлює нижчий рівень пристосування. Це зрозуміло, якщо наприклад є 100 000 особин
у популяції. Тоді якщо є два алелі то це відношення алелей буде приблизно 50 до 50. але
якщо взяти 10 особин, тоді два алелі можуть бути 3 домінантних, 7 рецисивних. Може бути
так, що ген який обумовлює пристосованість зовсім зникне. Це цілком автоматичний
процес.
Може бути навпаки, популяція заснована особинами із дуже великою
пристосованісттю до зовнишнього середовища. Майер назвав це явище, «генетичною
революцією». Він вважав, що нові види виникають завдяки цієй «генетичній революції».
Якщо нова популяція виникає за рахунок імміграції малого числа особин, то більша частина
материнської популяції втрачається. Вплив початкового обмеженного різноманіття
генофонду на подальшу судьбу популяції назано “ефектом першозасновника”.
28. Інбридинг
Кінцева чисельність реальних популяцій приводить ще до одного важливого наслідку.Розглянемо його на прикладі людини. У кожного з нас двоє батьків, четверо бабусь і дідусів,
вісім прабабів і прадідів і т.д. На п поколінь у минулому кожний з нас має 2n предків. За 20
останніх поколінь кожний мав більше 700 тис. предків. Приймемо, що на кожне сторіччя
доводяться чотири покоління. У цьому випадку на 20 поколінь потрібен час у п'ять сторіч.
Якби все населення такої країни, як США, не зв'язане безпосереднім спорідненням (тобто
один з кожних 20 чоловік), мало абсолютно незалежні набори предків, то в XV столітті в країні
повинне було б жити 3 трильйони людей. Ця цифра на кілька порядків перевищує все
населення земної кулі в цей час. Ситуація зовсім нереальна й, отже, споріднення між людьми
набагато більше, ніж це може здаватися на перший погляд.
Аналогічні міркування застосовні до будь-якої популяції й у такий спосіб у популяціях можна
виміряти середній ступінь кревного споріднення особин
(α):α= ΣPiFi,
де рi, — частота особин з коефіцієнтом інбридингу Fi,
29. Інбридинг має кілька наслідків для популяції:
1. Підвищення гомозиготності.2. Прояв рецесивных алелей.
3. При звичайно негативному ефекті рецесивных
аллелей інбридинг спричиняє ослаблення особин
(інбредна депресія).
4. Підвищення фенотипової мінливості внаслідок
виходу в гомозиготу багатьох алелей.
30. Ізоляція
Ізоляцію забезпечують географічні, або територіально-механичні, ібіологічні фактори. Біологічні фактори ізоляції в остаточному підсумку
засновані на генетичних факторах. Навіть поведінкові, або етологічні,
фактори ізоляції базуються на генетичних розходженнях особин. У той же
час варто виділити й властиво генетичні фактори ізоляції, такі, як:
1)
2)
3)
4)
поліплоїдія,
хромосомні перебудови,
ядерно-цитоплазматична несумісність,
несумісність експресії окремих генів внаслідок їхніх мутаційних змін.
Генетичні, так само як і інші фактори ізоляції, збільшують імовірність
схрещування між родинними особинами й тим самим підвищують
ступінь інбридингу в популяціях.
Ті або інші форми ізоляції лежать в основі видоутворення й приводять до
різної екологічної спеціалізації біологічних форм, до освоєння ними
нових екологічних ніш. Екологічна ізоляція є результатом розселення.