






















 chemistry
chemistrySimilar presentations:
")




")




Лекция 9. Липиды. Метаболизм
1.
ГБОУ ВПО УГМУ Минздрава РФКафедра биохимии
Дисциплина: Биохимия
ЛЕКЦИЯ № 9
Липиды.
Метаболизм
Лектор: Гаврилов И.В.
Факультет: лечебно-профилактический,
Курс: 2
Екатеринбург, 2015г
2.
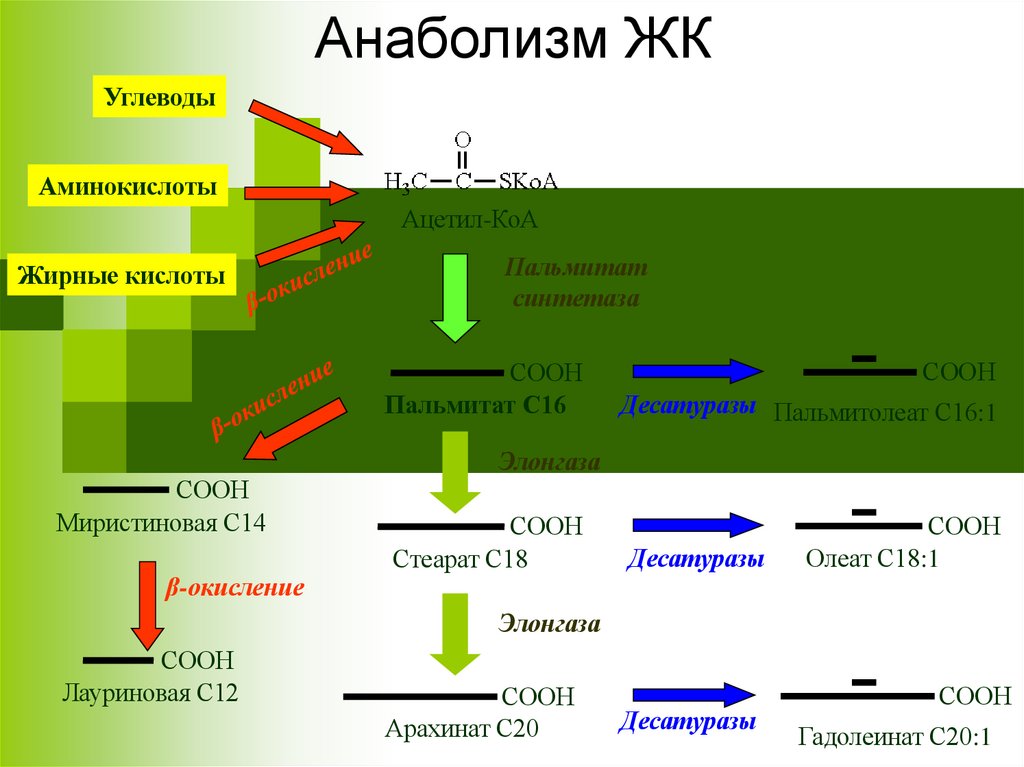
Анаболизм ЖКУглеводы
Аминокислоты
Ацетил-КоА
Жирные кислоты
Пальмитат
синтетаза
СООН
Пальмитат С16
СООН
Миристиновая С14
β-окисление
СООН
Десатуразы Пальмитолеат С16:1
Элонгаза
СООН
Стеарат С18
Десатуразы
СООН
Олеат С18:1
Элонгаза
СООН
Лауриновая С12
СООН
Арахинат С20
Десатуразы
СООН
Гадолеинат С20:1
3.
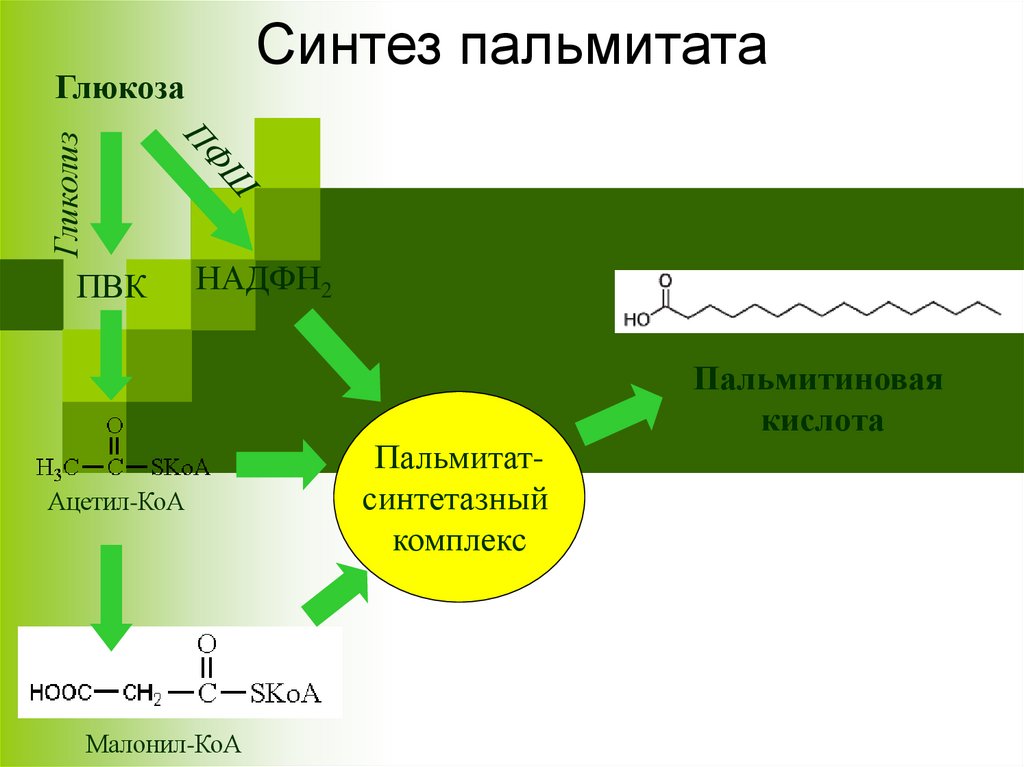
Синтез пальмитатаГликолиз
Глюкоза
ПВК
НАДФН2
Ацетил-КоА
Малонил-КоА
Пальмитатсинтетазный
комплекс
Пальмитиновая
кислота
4.
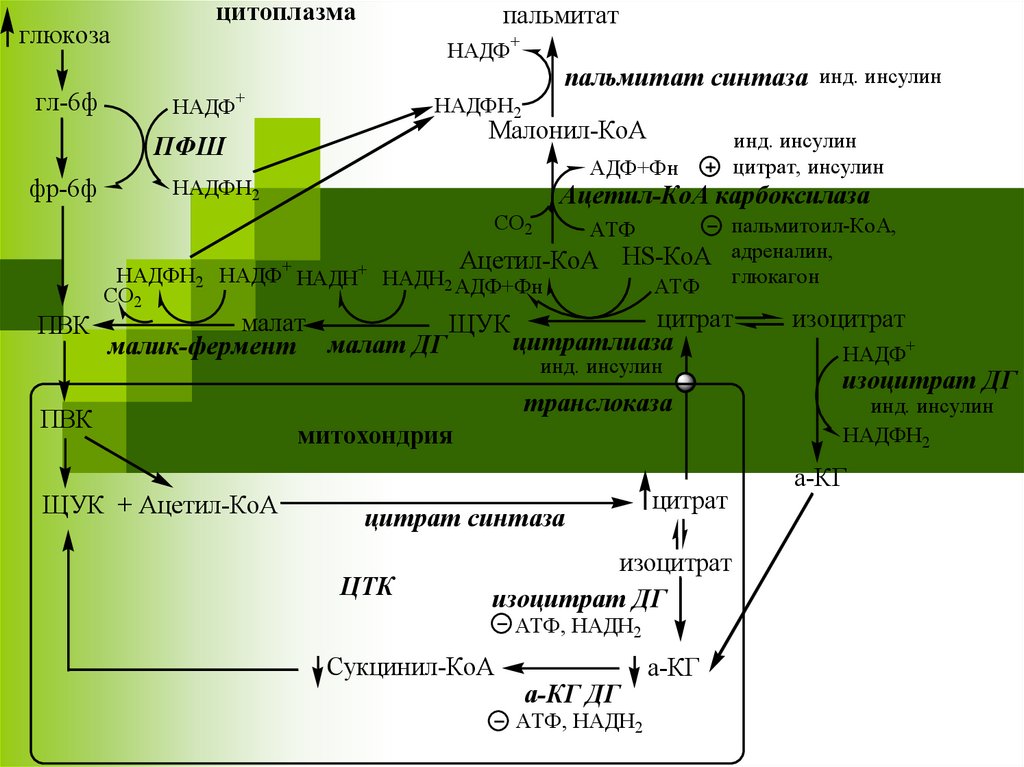
глюкозагл-6ф
цитоплазма
пальмитат
НАДФ+
пальмитат синтаза инд. инсулин
НАДФ+
НАДФН2
Малонил-КоА
ПФШ
фр-6ф
АДФ+Фн
НАДФН2
Ацетил-КоА карбоксилаза
СО2
пальмитоил-КоА,
HS-КоА адреналин,
АТФ глюкагон
АТФ
Ацетил-КоА
ПВК
инд. инсулин
цитрат, инсулин
НАДФН2 НАДФ+ НАДН+ НАДН2 АДФ+Фн
СО2
цитрат
малат
ЩУК
цитратлиаза
малик-фермент малат ДГ
инд. инсулин
ПВК
ЩУК + Ацетил-КоА
транслоказа
митохондрия
цитрат синтаза
ЦТК
цитрат
изоцитрат
изоцитрат ДГ
АТФ, НАДН2
Сукцинил-КоА
а-КГ ДГ
АТФ, НАДН2
а-КГ
изоцитрат
НАДФ+
изоцитрат ДГ
инд. инсулин
НАДФН2
а-КГ
5.
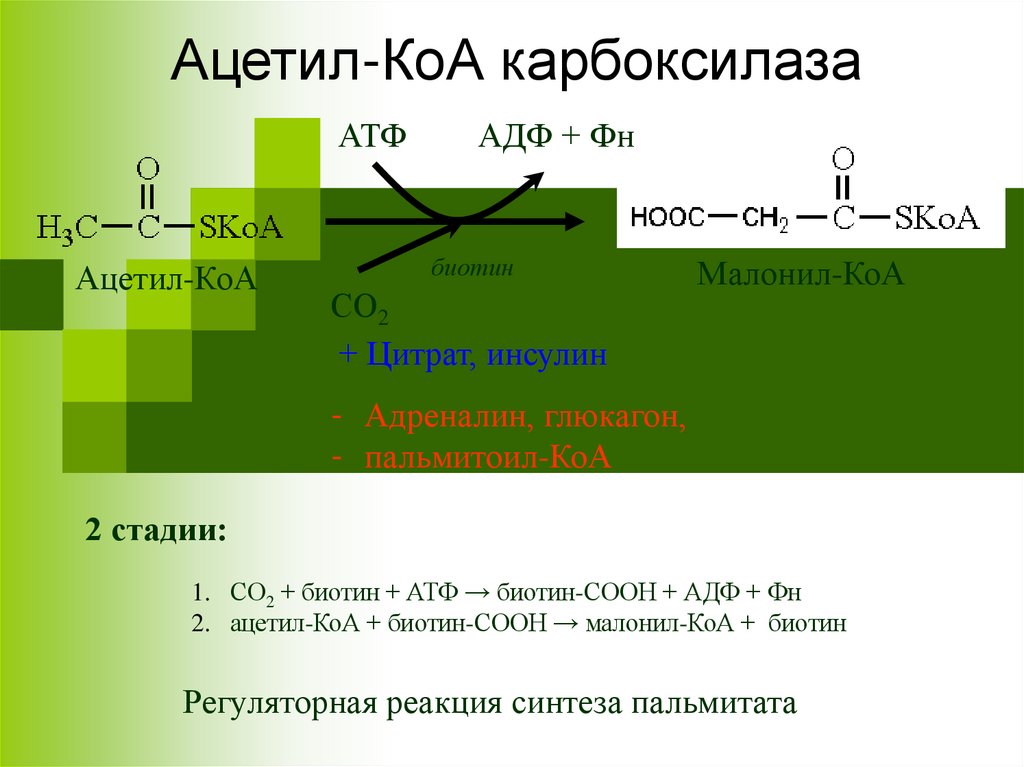
Ацетил-КоА карбоксилазаАТФ
Ацетил-КоА
АДФ + Фн
биотин
СО2
+ Цитрат, инсулин
Малонил-КоА
- Адреналин, глюкагон,
- пальмитоил-КоА
2 стадии:
1. СО2 + биотин + АТФ → биотин-СООН + АДФ + Фн
2. ацетил-КоА + биотин-СООН → малонил-КоА + биотин
Регуляторная реакция синтеза пальмитата
6.
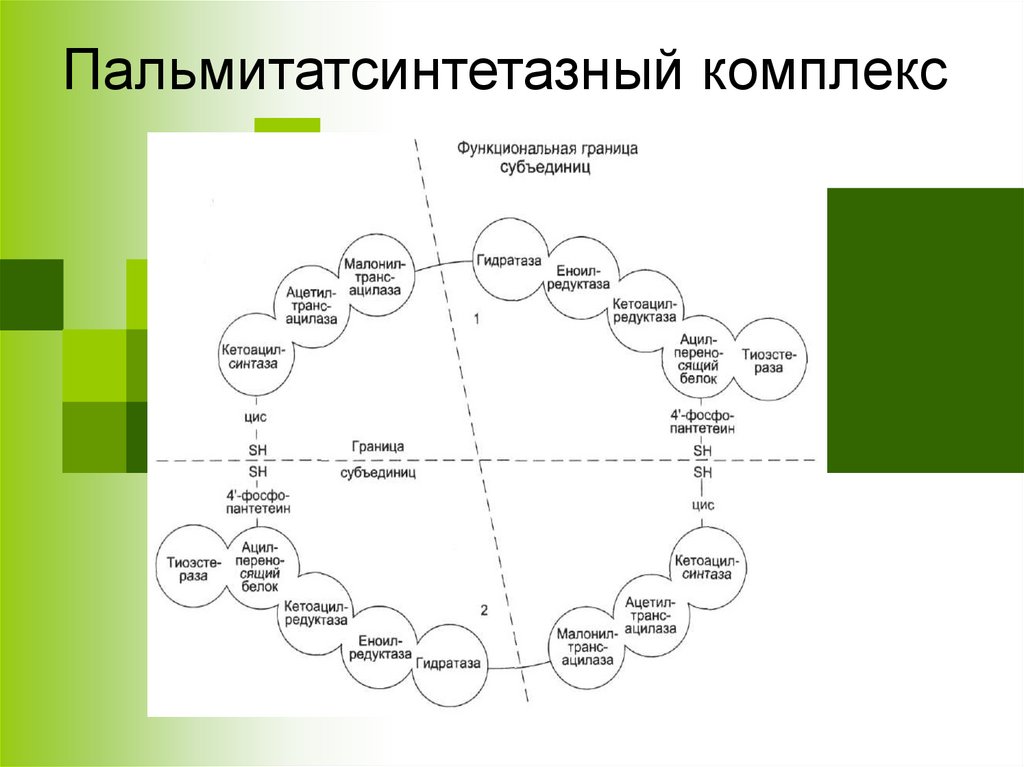
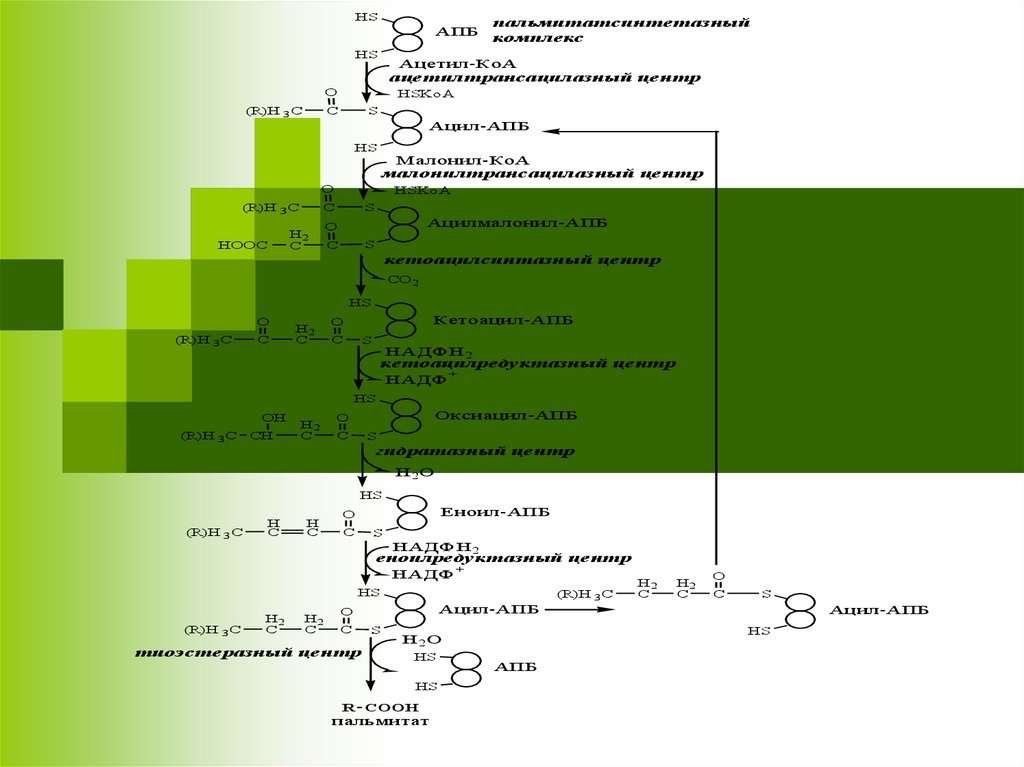
Пальмитатсинтетазный комплекс7.
HSАПБ
HS
Ацетил-КоА
ацетилтрансацилазный центр
O
(R)H 3C
НSKoA
C
S
Ацил-АПБ
HS
Малонил-КоА
малонилтрансацилазный центр
O
(R)H 3C
HOOC
пальмитатсинтетазный
комплекс
НSKoA
C
H2
C
S
Ацилмалонил-АПБ
O
S
C
кетоацилсинтазный центр
СО 2
HS
О
(R)H 3C
C
H2
C
Кетоацил-АПБ
O
C
S
НАДФН2
кетоацилредуктазный центр
НАДФ+
HS
ОH
(R)H 3C
CH
H2
C
Оксиацил-АПБ
O
C
S
гидратазный центр
Н2О
HS
(R)H 3C
H
C
H
C
Еноил-АПБ
O
C
S
НАДФН2
еноилредуктазный центр
НАДФ+
HS
(R)H 3C
H2
C
H2
C
Ацил-АПБ
O
C
тиоэстеразный центр
S
HS
R CООН
пальмитат
H2
C
H2
C
O
C
S
Ацил-АПБ
HS
Н2О
HS
(R)H 3C
АПБ
8.
HSАПБ
HS
Ацетил-КоА
ацетилтрансацилазный центр
O
(R)H 3C
НSKoA
C
S
Ацил-АПБ
HS
Малонил-КоА
малонилтрансацилазный центр
O
(R)H 3C
HOOC
пальмитатсинтетазный
комплекс
НSKoA
C
H2
C
S
Ацилмалонил-АПБ
O
S
C
кетоацилсинтазный центр
СО 2
HS
О
(R)H 3C
C
H2
C
Кетоацил-АПБ
O
C
S
НАДФН2
кетоацилредуктазный центр
НАДФ+
HS
ОH
(R)H 3C
CH
H2
C
Оксиацил-АПБ
O
C
S
гидратазный центр
Н2О
HS
(R)H 3C
H
C
H
C
Еноил-АПБ
O
C
S
НАДФН2
еноилредуктазный центр
НАДФ+
HS
(R)H 3C
H2
C
H2
C
Ацил-АПБ
O
C
тиоэстеразный центр
S
HS
R
CООН
пальмитат
H2
C
H2
C
O
C
S
Ацил-АПБ
HS
Н2О
HS
(R)H 3C
АПБ
9.
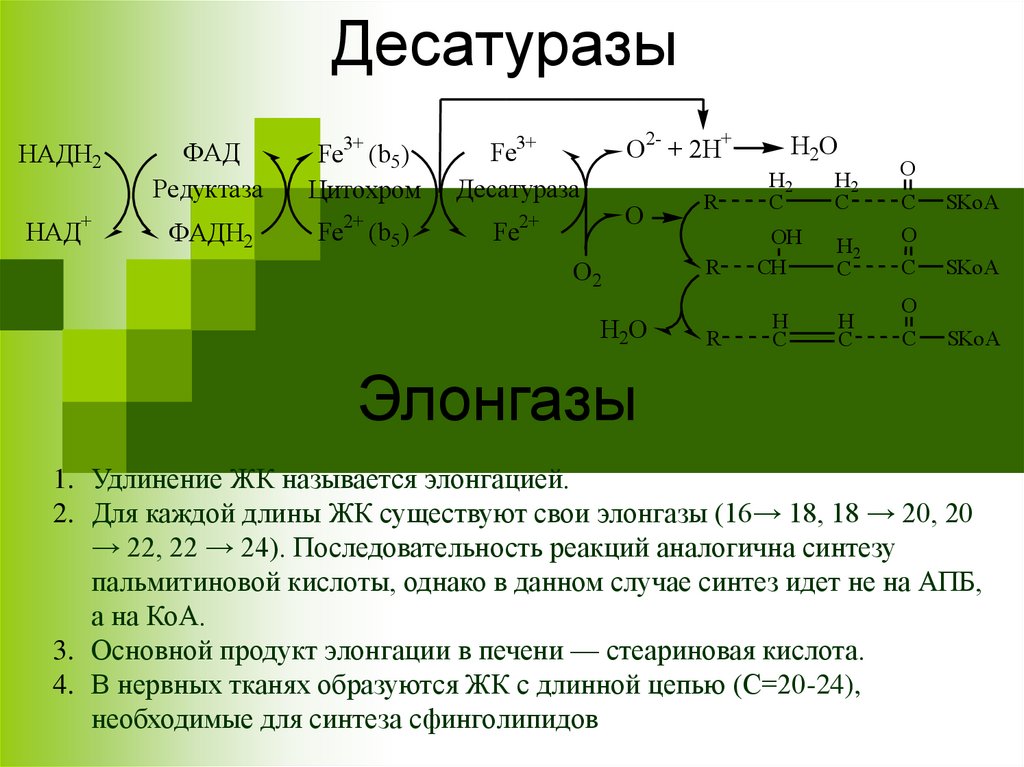
ДесатуразыНАДН2
НАД+
ФАД
Редуктаза
Fe3+ (b5)
Цитохром
Fe3+
Десатураза
ФАДН2
Fe2+ (b5)
Fe2+
O2- + 2H+
O
O2
H2O
H2O
R
H2
C
H2
C
R
OH
CH
R
H
C
O
C
SKoA
H2
C
O
C
SKoA
H
C
O
C
SKoA
Элонгазы
1. Удлинение ЖК называется элонгацией.
2. Для каждой длины ЖК существуют свои элонгазы (16→ 18, 18 → 20, 20
→ 22, 22 → 24). Последовательность реакций аналогична синтезу
пальмитиновой кислоты, однако в данном случае синтез идет не на АПБ,
а на КоА.
3. Основной продукт элонгации в печени — стеариновая кислота.
4. В нервных тканях образуются ЖК с длинной цепью (С=20-24),
необходимые для синтеза сфинголипидов
10.
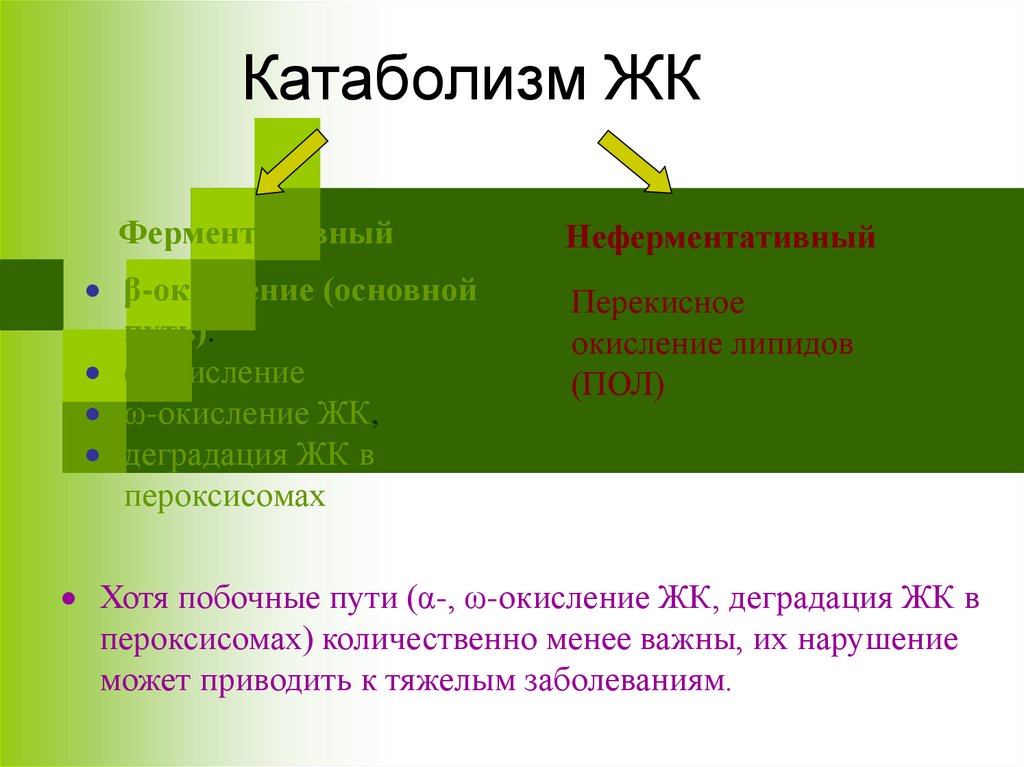
Катаболизм ЖКФерментативный
β-окисление (основной
путь).
α-окисление
ω-окисление ЖК,
деградация ЖК в
пероксисомах
Неферментативный
Перекисное
окисление липидов
(ПОЛ)
Хотя побочные пути (α-, ω-окисление ЖК, деградация ЖК в
пероксисомах) количественно менее важны, их нарушение
может приводить к тяжелым заболеваниям.
11.
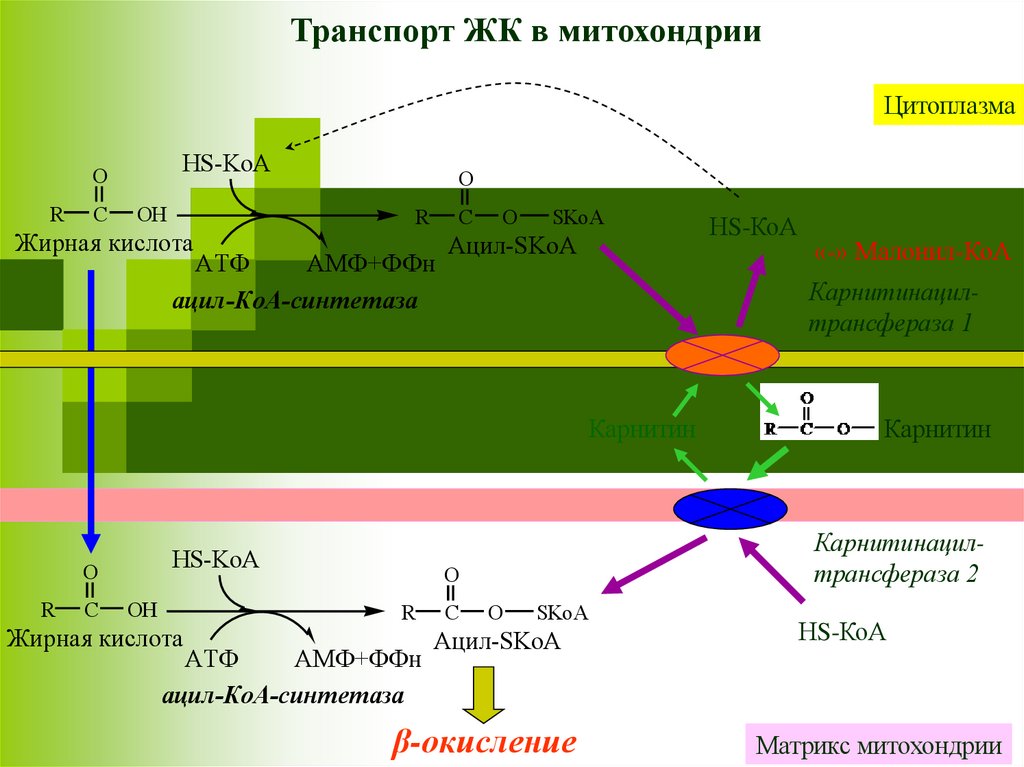
Транспорт ЖК в митохондрииЖК с короткой и средней цепью (от 4 до 12 атомов С) могут
проникать в матрикс митохондрий путём диффузии. Активация
этих ЖК происходит ацил-КоА синтетазами в матриксе
митохондрий.
ЖК с длинной цепью, сначала активируются в цитозоле (ацилКоА синтетазами на внешней мембране митохондрий), а затем
переносятся в матрикс митохондрий специальной транспортной
системой с помощью карнитина.
Карнитин поступает с пищей или
синтезируется из лизина и
метионина с участием витамина
С.
L-карнитин
12.
Транспорт ЖК в митохондрииЦитоплазма
HS-KoA
O
R
C
OH
Жирная кислота
O
R
C
АТФ
АМФ+ФФн
ацил-КоА-синтетаза
O
SKoA
Ацил-SKoA
O
R
C
OH
Жирная кислота
АТФ
АМФ+ФФн
ацил-КоА-синтетаза
C
Карнитин
Карнитинацилтрансфераза 2
O
R
«-» Малонил-КоА
Карнитинацилтрансфераза 1
Карнитин
HS-KoA
НS-КоА
O
SKoA
Ацил-SKoA
β-окисление
НS-КоА
Матрикс митохондрии
13.
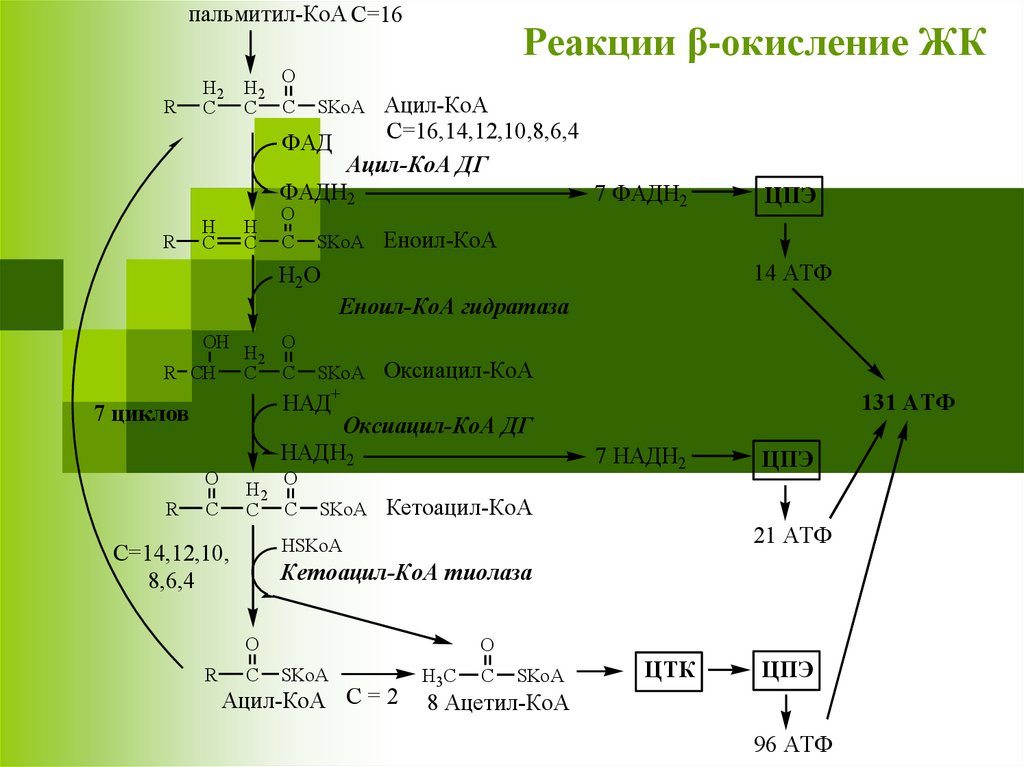
пальмитил-КоА C=16R
R
H2
C
O
H2
C C
H
C
O
C
Реакции β-окисление ЖК
Ацил-КоА
C=16,14,12,10,8,6,4
ФАД
Ацил-КоА ДГ
ФАДН2
7 ФАДН2
H
C
SKoA
SKoA
ЦПЭ
Еноил-КоА
14 АТФ
Н2О
Еноил-КоА гидратаза
ОН
R CН
O
H2
C C
Оксиацил-КоА
НАД+
7 циклов
Оксиацил-КоА ДГ
НАДН2
О
R
SKoA
O
H2
C C
C
SKoA
7 НАДН2
ЦПЭ
Кетоацил-КоА
21 АТФ
НSKoA
C=14,12,10,
8,6,4
Кетоацил-КоА тиолаза
O
R
131 АТФ
C
O
SKoA
Ацил-КоА C = 2
H3C
C
SKoA
ЦТК
ЦПЭ
8 Ацетил-КоА
96 АТФ
14.
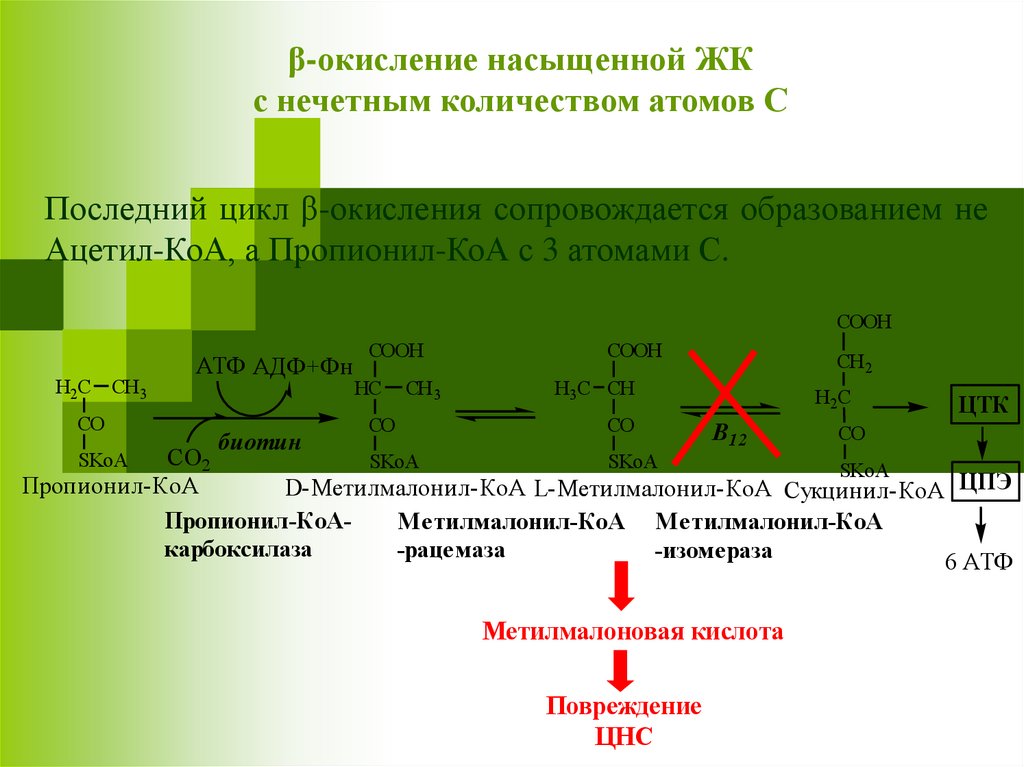
β-окисление насыщенной ЖКс нечетным количеством атомов С
Последний цикл β-окисления сопровождается образованием не
Ацетил-КоА, а Пропионил-КоА с 3 атомами С.
COOH
H2C
CH 3
CO
АТФ АДФ+Фн
биотин
COOH
HC
CO
CH 3
COOH
CH 2
H3C CH
CO
H2C
В12
CO
ЦТК
CO2
SKoA
SKoA
SKoA
Пропионил-КоА
D-Метилмалонил-КоА L-Метилмалонил-КоА Сукцинил-КоА ЦПЭ
Пропионил-КоАМетилмалонил-КоА Метилмалонил-КоА
карбоксилаза
-рацемаза
-изомераза
6 АТФ
SKoA
Метилмалоновая кислота
Повреждение
ЦНС
15.
Регуляцияβ-окисление активируют: глюкагон, адреналин, ЖК, НАД+,
АДФ:
Голод, физическая нагрузка → ↑ глюкагон, ↑ адреналин →
липолиз ТГ в адипоцитах → ↑ ЖК в крови → ↑ β-окисление в
аэробных условиях в мышцах, печени → ↑АТФ;
↑АТФ, ↑НАДH2
↑Ацетил-КоА → ↑ цитрат, ↑ЖК → ↓ гликолиз → ↑ экономию
глюкозы для нервной ткани, эритроцитов и т.д.
β-окисление ингибируют: инсулин, НАДH2, АТФ.
Пища → ↑ инсулин → ↑ гликолиз → ↑ Ацетил-КоА → ↑ синтез
малонил-КоА и ЖК, ↑ малонил-КоА →
↓ карнитинацилтрансферазы I в печени → ↓ транспорт ЖК в
матрикс митохондрий → ↓ ЖК в матриксе → ↓ β-окисление
ЖК
16.
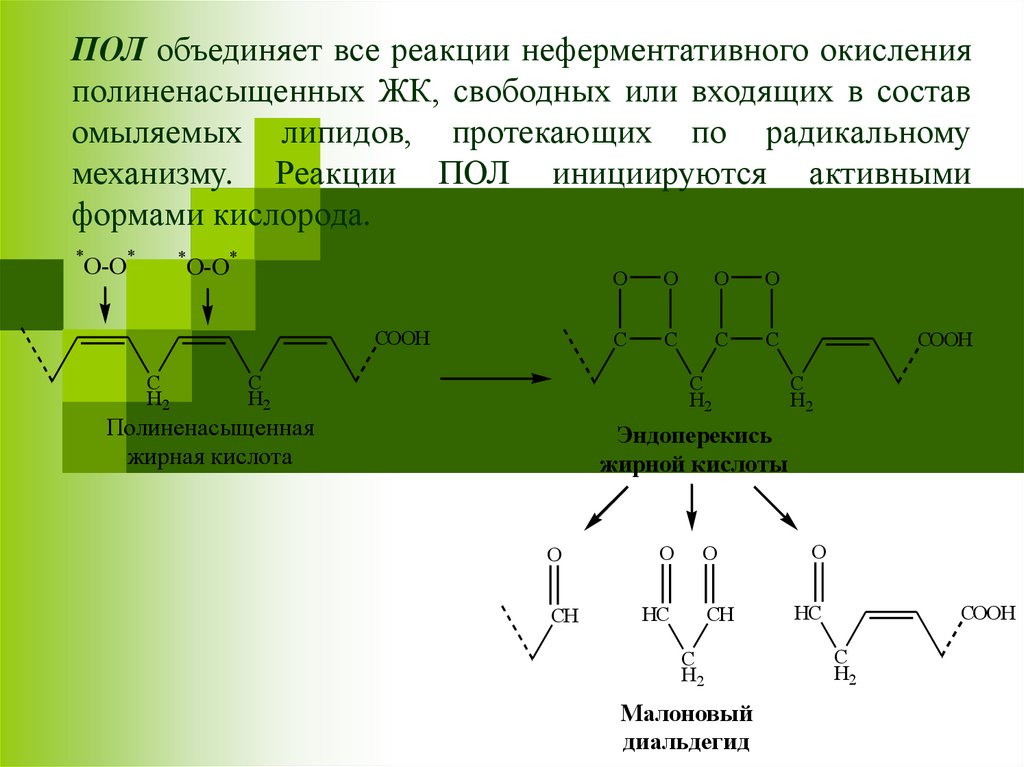
ПОЛ объединяет все реакции неферментативного окисленияполиненасыщенных ЖК, свободных или входящих в состав
омыляемых липидов, протекающих по радикальному
механизму. Реакции ПОЛ инициируются активными
формами кислорода.
*
O-O*
*
O-O*
СООН
C
H2
O
O
O
O
С
С
С
С
C
H2
C
H2
Полиненасыщенная
жирная кислота
СООН
C
H2
Эндоперекись
жирной кислоты
O
O
O
O
CH
HC
СH
HС
C
H2
Малоновый
диальдегид
СООН
C
H2
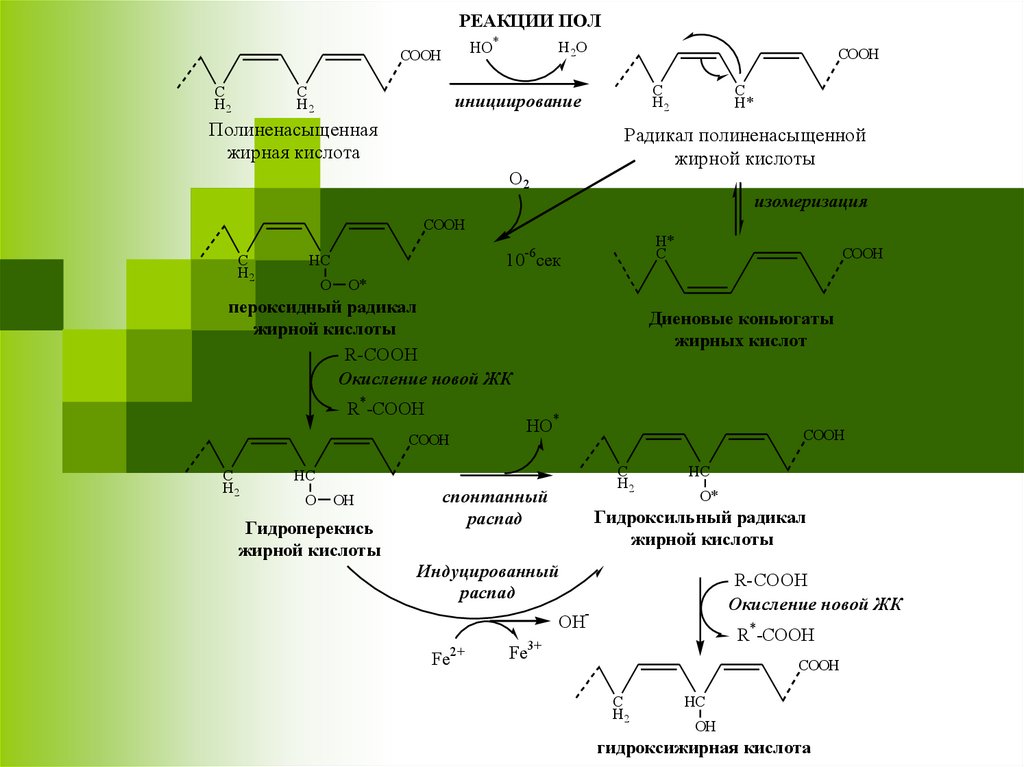
17.
РЕАКЦИИ ПОЛHО*
СООН
C
H2
C
H2
H2O
СООН
C
H2
инициирование
Полиненасыщенная
жирная кислота
Радикал полиненасыщенной
жирной кислоты
О2
изомеризация
СООН
C
H2
O
H*
C
10-6сек
HC
R*-COOH
СООН
Диеновые коньюгаты
жирных кислот
HO*
OH
Гидроперекись
жирной кислоты
СООН
C
H2
HC
O
COOH
O*
пероксидный радикал
жирной кислоты
R-COOH
Окисление новой ЖК
C
H2
C
H*
спонтанный
распад
HC
O*
Гидроксильный радикал
жирной кислоты
Индуцированный
распад
R-COOH
Окисление новой ЖК
OHFe2+
R*-COOH
Fe3+
СООН
C
H2
HC
OH
гидроксижирная кислота
18.
Регуляция ПОЛПроцессы ПОЛ усиливаются при избытке катехоламинов (стресс), гипоксии, ишемии
(при реоксигенации), повышенном содержании активных форм О2, снижении
антиоксидантной защиты, повышенном содержании ненасыщенных жирных
кислот.
Активация ПОЛ происходит при воспалении и характерна для многих заболеваний:
дистрофии мышц (болезнь Дюшенна), болезни Паркинсона, атеросклерозе,
развитии опухолей.
Физиологическое значение реакций ПОЛ:
1. модифицируют физико-химические свойства биомембран: увеличивают их
проницаемость.
2. регулируют активность мембранных ферментов, реакции окислительного
фосфорилирования.
3. участвуют в контроле клеточного деления.
4. Участвует в адаптации организма.
Повышение активности ПОЛ приводит к:
1. разрушению, фрагментации клеточных мембран, повреждению и гибели клеток.
2. модификации ЛП, особенно ЛПНП. Они становятся «липкими», легче проникают в
сосудистую стенку, хорошо захватываются макрофагами, что ускоряет развитие
атеросклероза.
3. накоплению продуктов ПОЛ, многие из которых токсичны, канцерогенены и
мутагенены (МДА).
4. ускорению процессов старения организма.
19.
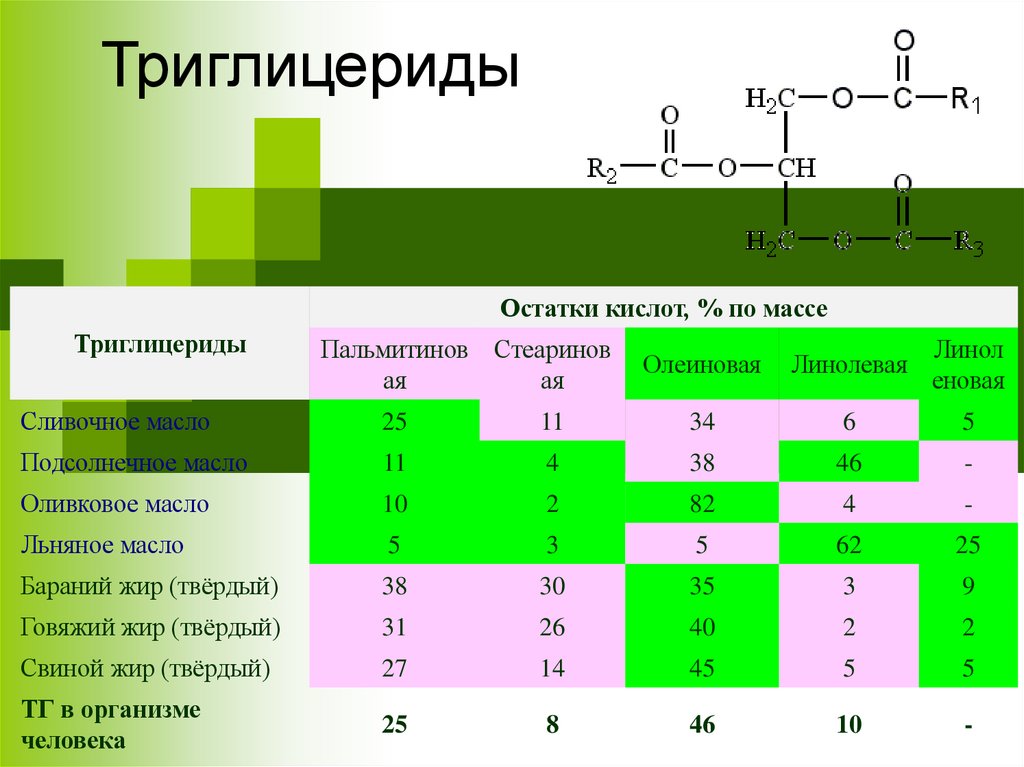
ТриглицеридыОстатки кислот, % по массе
Триглицериды
Пальмитинов Стеаринов
ая
ая
Олеиновая
Линолевая
Линол
еновая
Сливочное масло
25
11
34
6
5
Подсолнечное масло
11
4
38
46
-
Оливковое масло
10
2
82
4
-
Льняное масло
5
3
5
62
25
Бараний жир (твёрдый)
38
30
35
3
9
Говяжий жир (твёрдый)
31
26
40
2
2
Свиной жир (твёрдый)
27
14
45
5
5
ТГ в организме
человека
25
8
46
10
-
20.
Жировая тканьЖировая ткань – это разновидность соединительной ткани,
составляет 15-20% веса мужчин и 20-25% веса женщин.
Как и любая ткань, она состоит из клеток (адипоциты,
макрофаги, тучные клетки и т.д.) и межклеточного вещества.
Межклеточное вещество в жировой ткани представлено
небольшим количеством волокон (коллагена и эластина) и
основного вещества (гликозаминогликаны, протеоликаны).
Белая жировая ткань
Есть везде. Основные жировые
отложения имеются под кожей (в
подкожножировой клетчатке), и вокруг
внутренних органов, преимущественно в
области живота (висцеральный жир).
21.
функцииСинтез липидов из углеводов
запасающая (95% ТГ в жировой ткани, 5% - во
внутренних органах и костях), накопление витаминов А,
Д, Е
теплоизоляционная;
механическая защита органов и тканей;
эндокринная (секреция: лептина, эстрогенов,
ангиотензиногена, интерлейкина-6, ФНО-α);
Антибактериальная защита (комплементарные
факторы: адипсин, комплемент С3, фактор В)
Регуляция воспаления (α1-кислый гликопротеин,
гаптоглобин)
пассивное обезвреживание (депонирование)
токсичных веществ
Участие в водно-солевом обмене
Обеспечивает поддержание гомеостаза
22.
Химический состав65-85% - ТГ,
22% - вода,
5,8% белок,
15 ммоль/кг калий.
Из жирных кислот
42—51% - олеиновая (18:1),
22—31% пальмитиновая (16:0),
5—14% пальмитоолеиновая (16:1),
3—5 % миристиновая,
1—5 % линолевая
23.
ОСОБЕННОСТИ МЕТАБОЛИЗМААДИПОЦИТА БЕЛОЙ ЖИРОВОЙ ТКАНИ
Энергетический обмен медленный, анаэробный,
потребляет мало кислорода. Энергия АТФ тратится
на транспорт жирных кислот через клеточные
мембраны (с участием карнитина).
Белковый обмен низкий, белки синтезируются
адипоцитами преимущественно для собственных
нужд. На экспорт синтезируются лептин, белки
острой фазы воспаления (α1-кислый гликопротеин,
гаптоглобин), компоненты системы комплимента
(адипсин, комплемент С3, фактор В), интерлейкины.
Углеводный обмен. Невысокий, преобладает
катаболизм.
Липидный обмен - интенсивно идут реакции
липолиза и липогенеза.
24.
Бурая жировая тканьМало у взрослого человека, много у
новорожденного
Локализована около почек и
щитовидной железы.
Смешанная жировая ткань: между
лопатками, на грудной клетке и плечах.
25.
Функция бурой жировой тканитермогенез
«взрывной» липолиз
26.
ОСОБЕННОСТИ МЕТАБОЛИЗМА БУРОЙ ЖИРОВОЙ ТКАНИ1. потребляет много кислорода
2. активно окисляет глюкозу и жирные кислоты
3. энергетический обмен высокий (низкий синтез
АТФ, 2 реакции в гликолизе,1 в ц. Кребса), в
основном энергия в виде тепла
(несократительный термогенез)
4. разобщение в митохондриях белком
термогенином (РБ-1) процессов окисления и
фосфорилирования
5. Характерен феномен «взрывного липолиза»
6. Термогенез активируется при переохлаждении,
излишке липидов в крови, под действием
лептина.
27.
Липогенез – синтез липидов28.
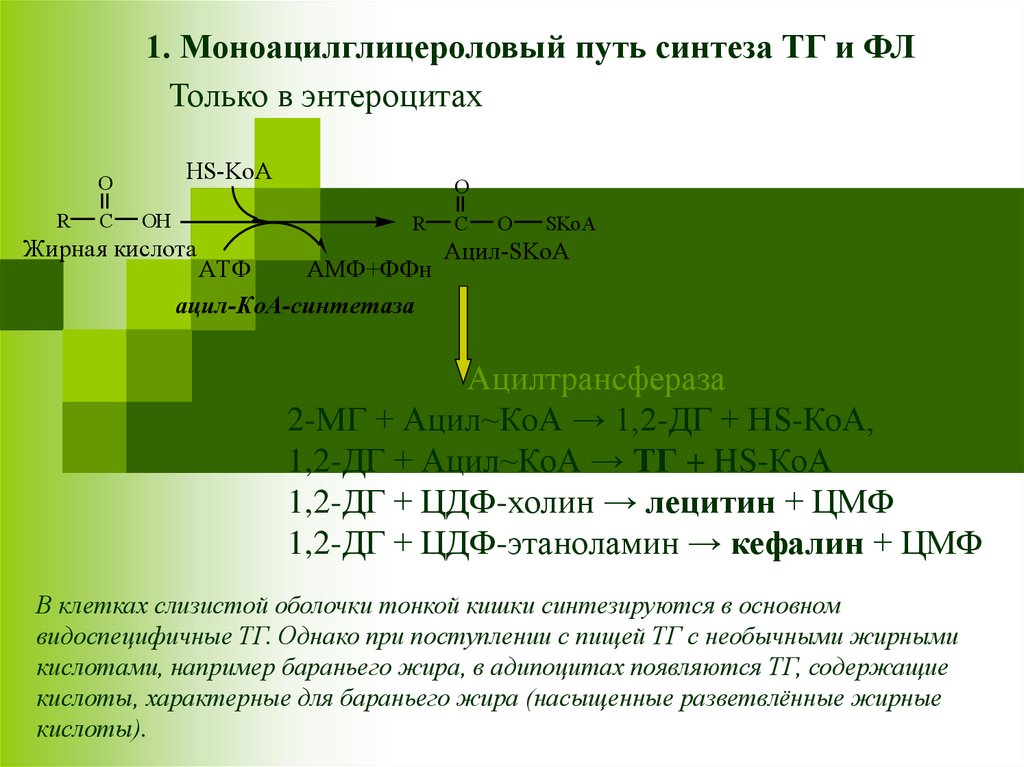
1. Моноацилглицероловый путь синтеза ТГ и ФЛТолько в энтероцитах
HS-KoA
O
R
C
OH
Жирная кислота
O
R
АТФ
АМФ+ФФн
ацил-КоА-синтетаза
C
O
SKoA
Ацил-SKoA
Ацилтрансфераза
2-МГ + Ацил~КоА → 1,2-ДГ + HS-КоА,
1,2-ДГ + Ацил~КоА → ТГ + HS-КоА
1,2-ДГ + ЦДФ-холин → лецитин + ЦМФ
1,2-ДГ + ЦДФ-этаноламин → кефалин + ЦМФ
В клетках слизистой оболочки тонкой кишки синтезируются в основном
видоспецифичные ТГ. Однако при поступлении с пищей ТГ с необычными жирными
кислотами, например бараньего жира, в адипоцитах появляются ТГ, содержащие
кислоты, характерные для бараньего жира (насыщенные разветвлённые жирные
кислоты).
29.
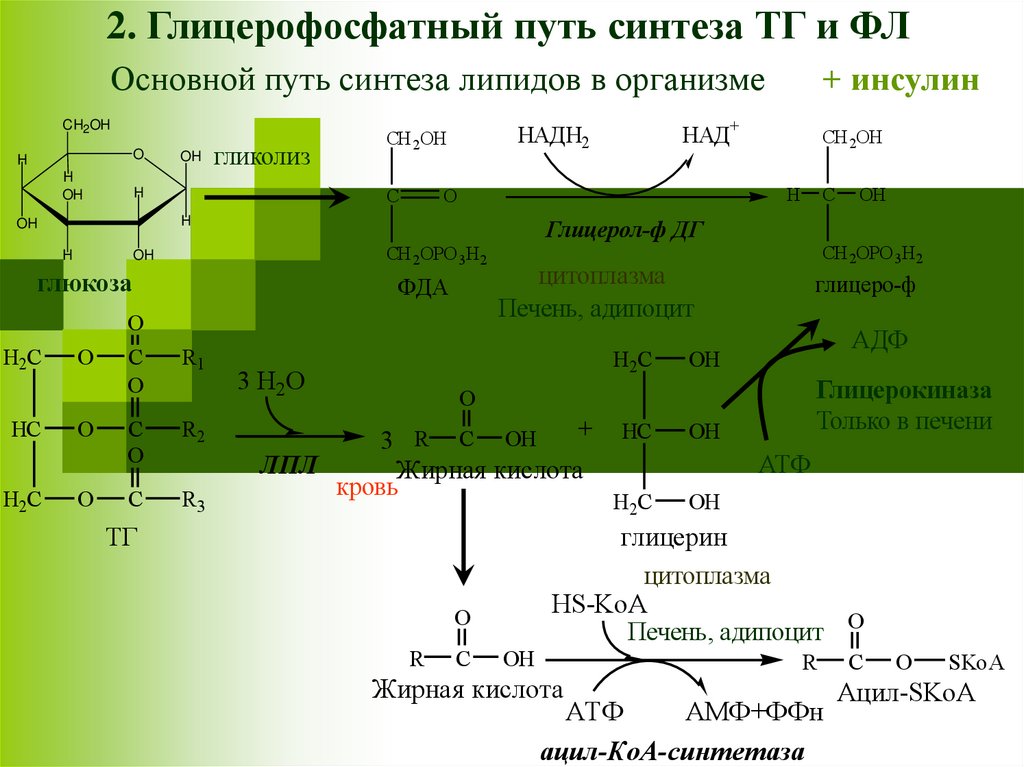
2. Глицерофосфатный путь синтеза ТГ и ФЛОсновной путь синтеза липидов в организме
CH2OH
O
H
H
OH
OH
гликолиз
С
H
H
CH 2OPO 3H2
OH
ФДА
O
HC
H2C
O
O
O
С
O
R1
С
O
R2
С
R3
С
Глицерол-ф ДГ
глюкоза
H2C
CH 2OH
H
O
H
OH
НАД+
НАДН2
CH 2OH
+ инсулин
3 Н2О
CH 2OPO 3H2
цитоплазма
Печень, адипоцит
H2C
OH
HC
OH
глицеро-ф
АДФ
Глицерокиназа
Только в печени
O
+
3 R C OH
ЛПЛ
Жирная кислота
кровь
ТГ
C
АТФ
H2C
OH
глицерин
цитоплазма
HS-KoA
Печень, адипоцит O
O
R
OH
OH
Жирная кислота
R
АТФ
АМФ+ФФн
ацил-КоА-синтетаза
C
O
SKoA
Ацил-SKoA
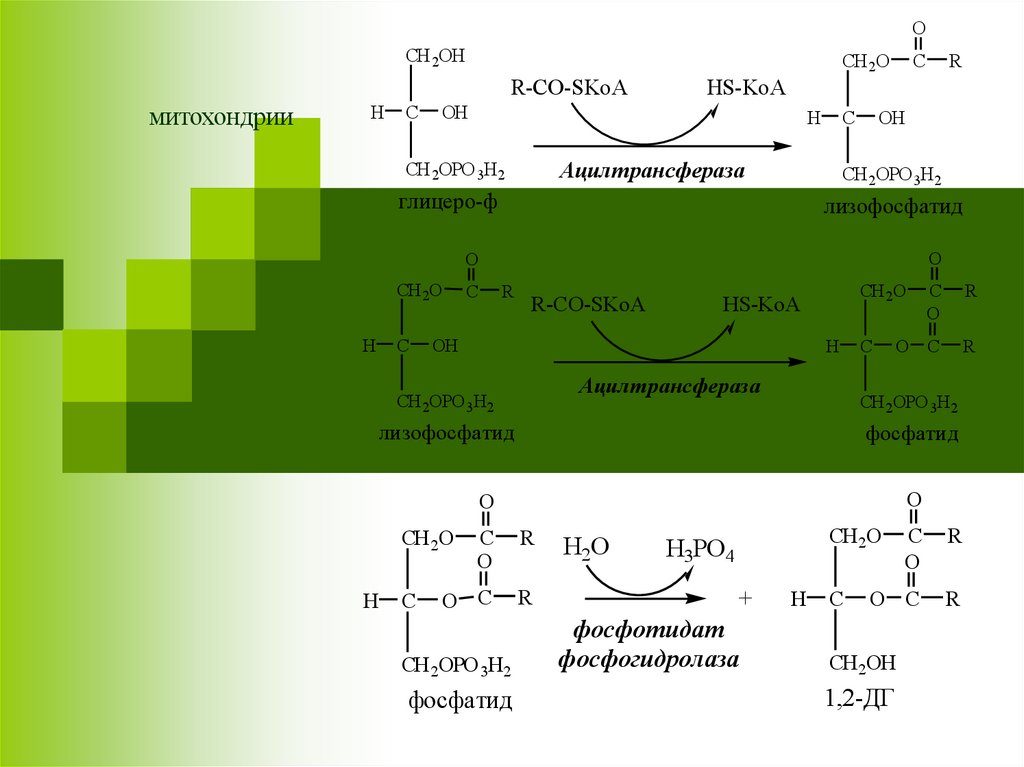
30.
OCH 2OH
R-CO-SKoA
митохондрии
H
С
CH 2O
HS-KoA
OH
H
С
Ацилтрансфераза
CH 2OPO 3H2
C
OH
CH 2OPO 3H2
глицеро-ф
лизофосфатид
O
O
CH 2O
H
С
C
R
R-CO-SKoA
CH 2O
HS-KoA
OH
H
Ацилтрансфераза
CH 2OPO 3H2
С
C
O
R
O C
R
CH 2OPO 3H2
лизофосфатид
фосфатид
O
O
CH 2O
H
С
R
C
O
R
O C
R
CH 2OPO 3H2
фосфатид
Н2О
CH 2O
Н3РО4
+
фосфотидат
фосфогидролаза
H
С
C
O
R
O C
R
CH 2OH
1,2-ДГ
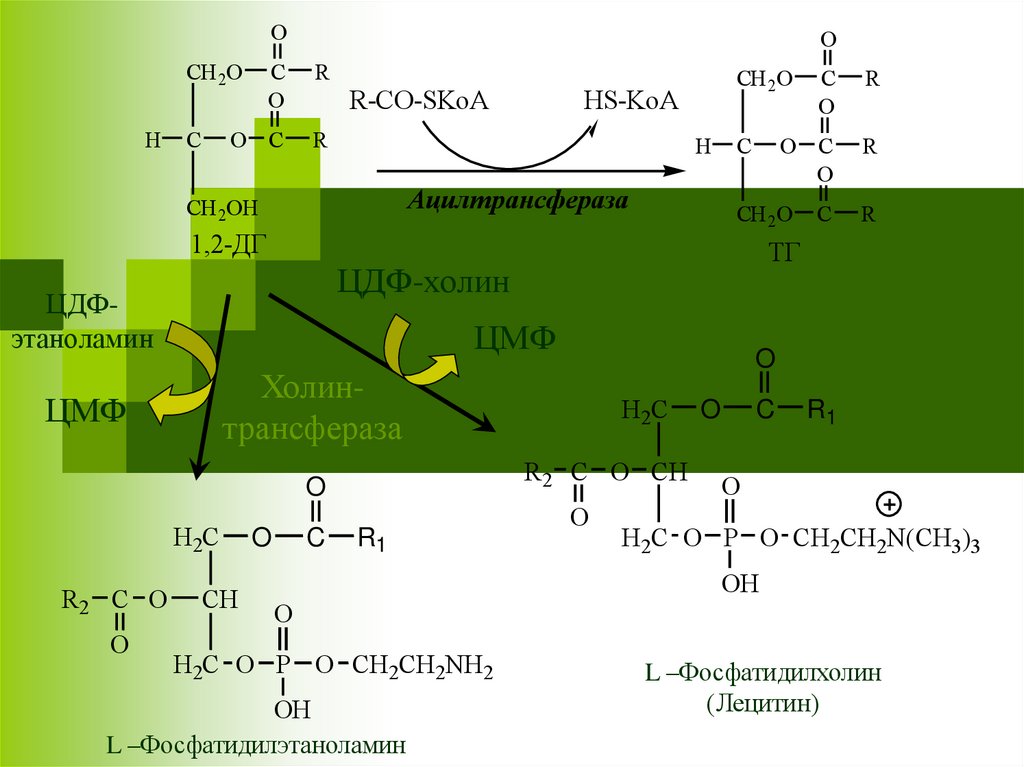
31.
OCH 2O
H
C
O
C
O
R
O C
R
R-CO-SKoA
CH 2O
HS-KoA
H
Ацилтрансфераза
CH 2OH
ЦМФ
H2C
O
CH
O
C
H2C
R2 C O CH
O
R1
R
C
R
O
Холинтрансфераза
ЦМФ
O C
O
ТГ
ЦДФ-холин
ЦДФэтаноламин
R
CH 2O
1,2-ДГ
R2 C O
C
C
O
O
O
C
R1
O
H2C O P O CH2CH2N(CH3)3
OH
O
H2C O P O CH2CH2NH2
OH
L –Фосфатидилэтаноламин
L –Фосфатидилхолин
(Лецитин)
32.
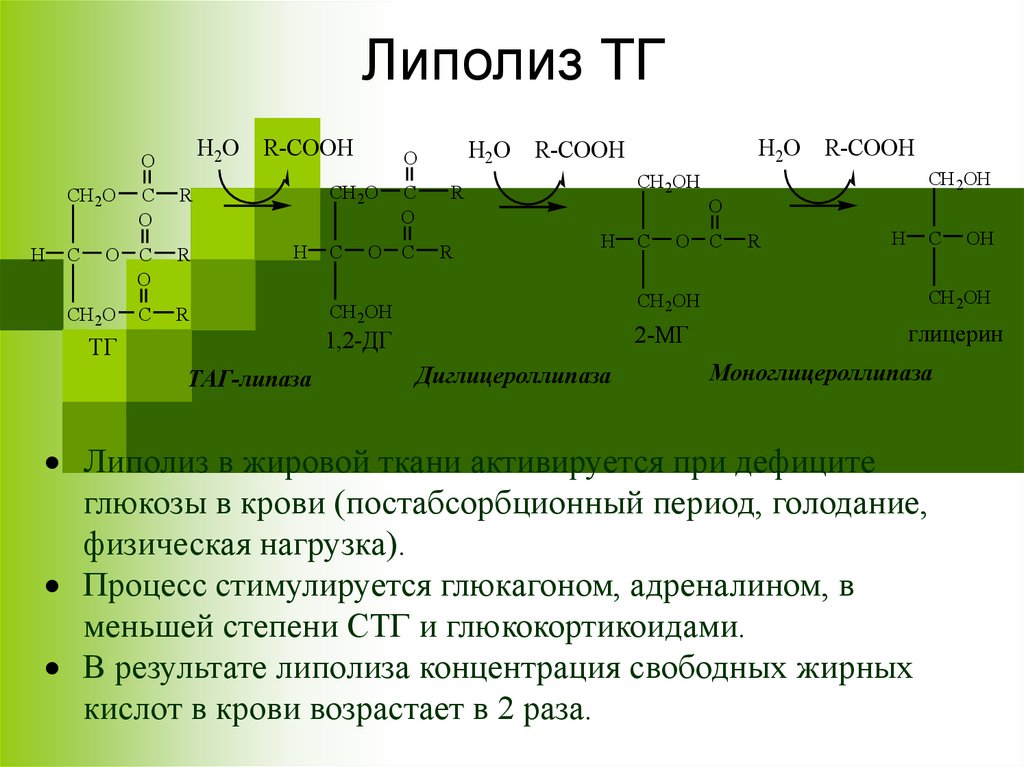
Липолиз – распад липидов33.
Липолиз ТГН2О R-COOH
O
CH 2O
H
C
C
O
R
O C
O
R
CH 2O
C
CH 2O
H
R
C
C
O
R
O C
R
CH 2OH
H
CH 2OH
1,2-ДГ
ТГ
ТАГ-липаза
Н2О R-COOH
Н2О R-COOH
O
Диглицероллипаза
C
CH 2OH
O
O C
R
H
C
OH
CH 2OH
CH 2OH
2-МГ
глицерин
Моноглицероллипаза
Липолиз в жировой ткани активируется при дефиците
глюкозы в крови (постабсорбционный период, голодание,
физическая нагрузка).
Процесс стимулируется глюкагоном, адреналином, в
меньшей степени СТГ и глюкокортикоидами.
В результате липолиза концентрация свободных жирных
кислот в крови возрастает в 2 раза.
34.
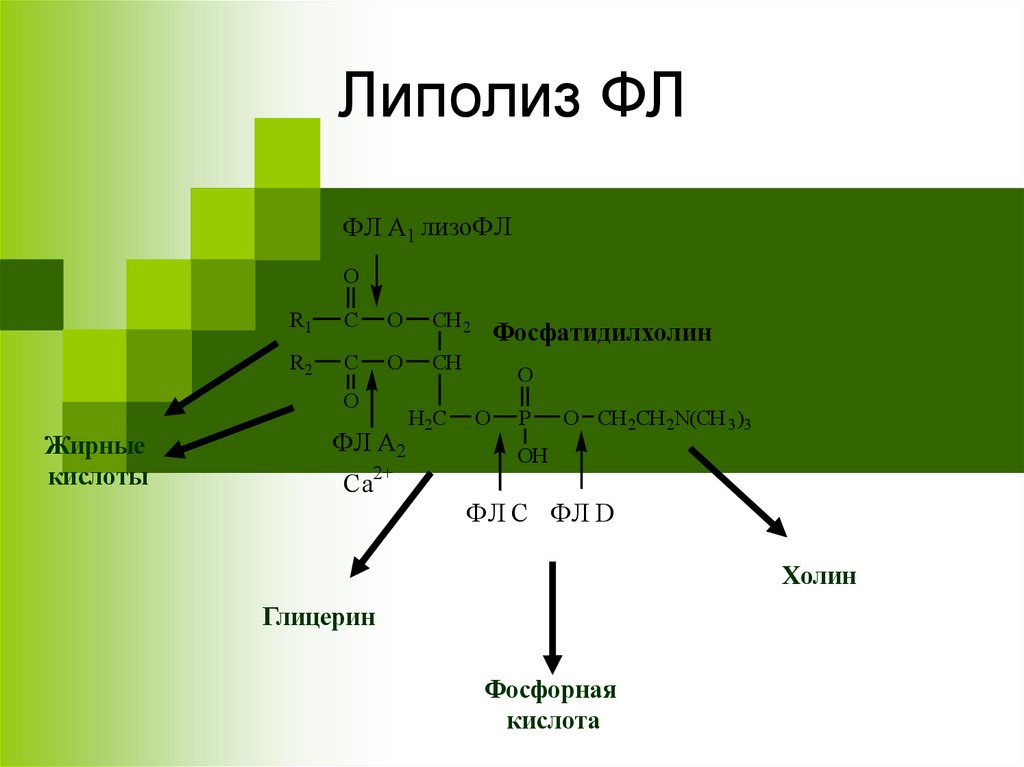
Липолиз ФЛФЛ А1 лизоФЛ
O
R1
C
O
CH 2
R2
C
O
CH
O
Жирные
кислоты
ФЛ А2
2+
H2C
Фосфатидилхолин
O
O
P
O CH 2CH 2N(CH 3)3
OH
Ca
ФЛ C ФЛ D
Холин
Глицерин
Фосфорная
кислота
35.
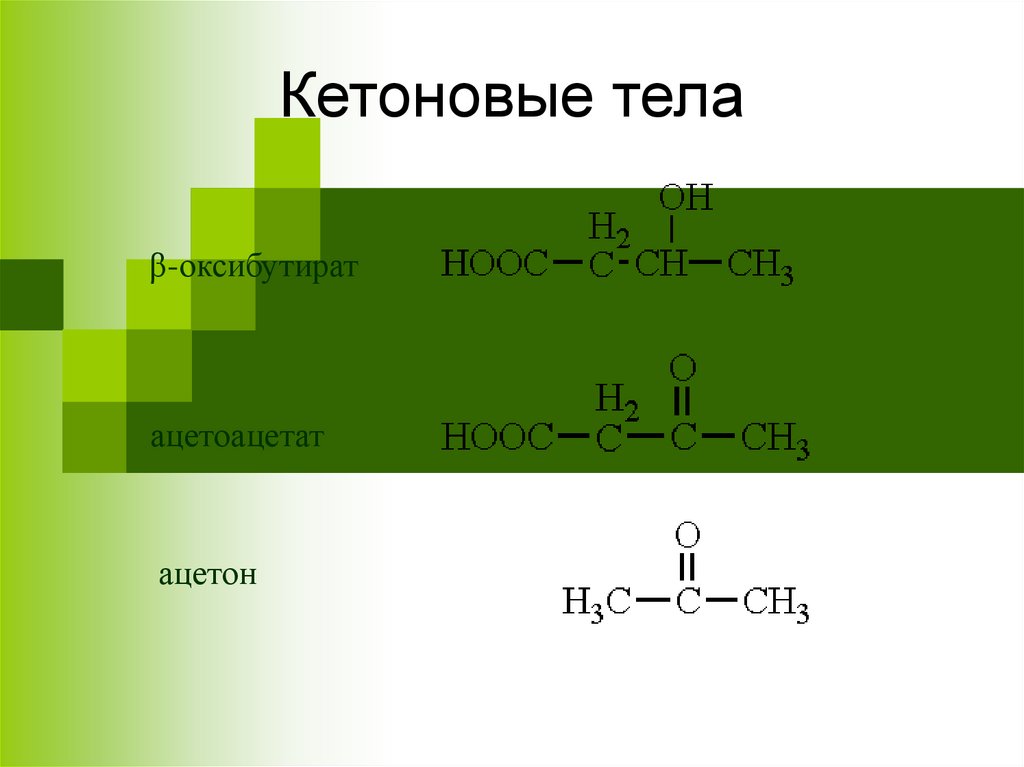
Кетоновые телаβ-оксибутират
ацетоацетат
ацетон
36.
Биологическое значение КТ1. КТ - топливные молекулы, окисление β-гидроксибутирата до
СО2 и Н2О обеспечивает синтез 26 АТФ. Окисление КТ, как и
ЖК сберегает глюкозу, что имеет большое значение в
энергоснабжении аэробных тканей при голодании и
физических нагрузках, когда возникает дефицит глюкозы.
2. Для нервной ткани КТ имеют исключительное значение, так
как в отличие от мышц и почек, нервная ткань практически не
использует ЖК в качестве источника энергии (ЖК не проходят
гематоэнцефалический барьер).
3. Преимущество КТ перед ЖК:
1) КТ водорастворимы, а ЖК – нет;
2) ЖК разобщают окислительное фосфорилирование и
усиливают синтез ТГ, а КТ – нет.
37.
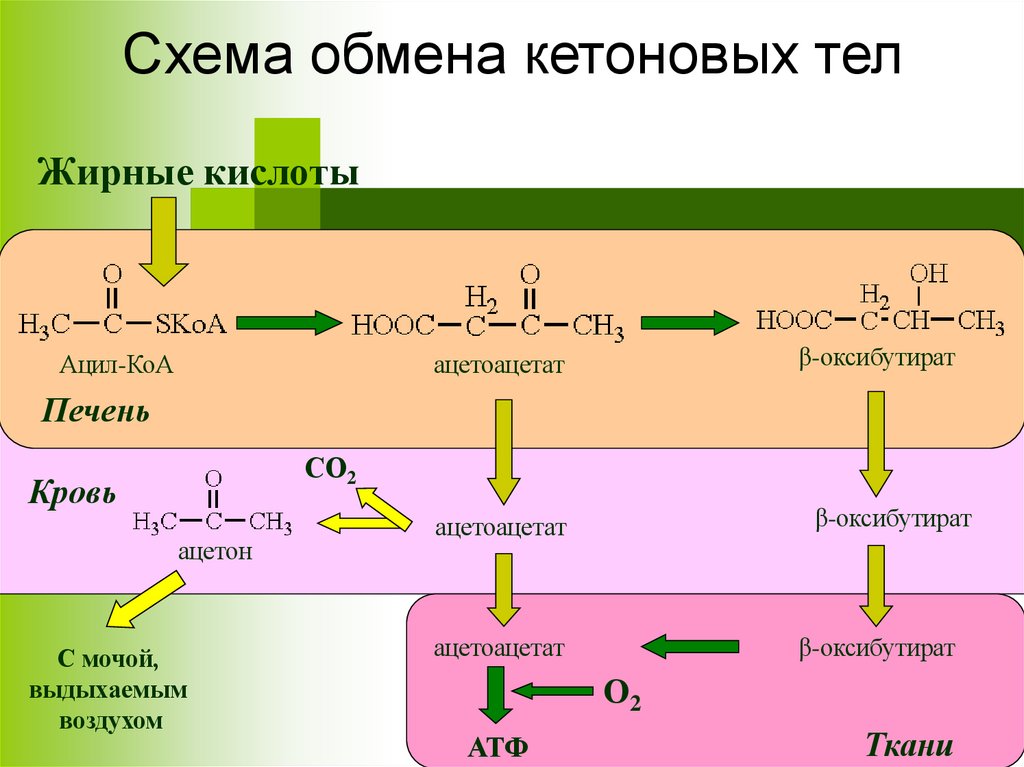
Схема обмена кетоновых телЖирные кислоты
Ацил-КоА
ацетоацетат
β-оксибутират
ацетоацетат
β-оксибутират
ацетоацетат
β-оксибутират
Печень
СО2
Кровь
ацетон
С мочой,
выдыхаемым
воздухом
О2
АТФ
Ткани
38.
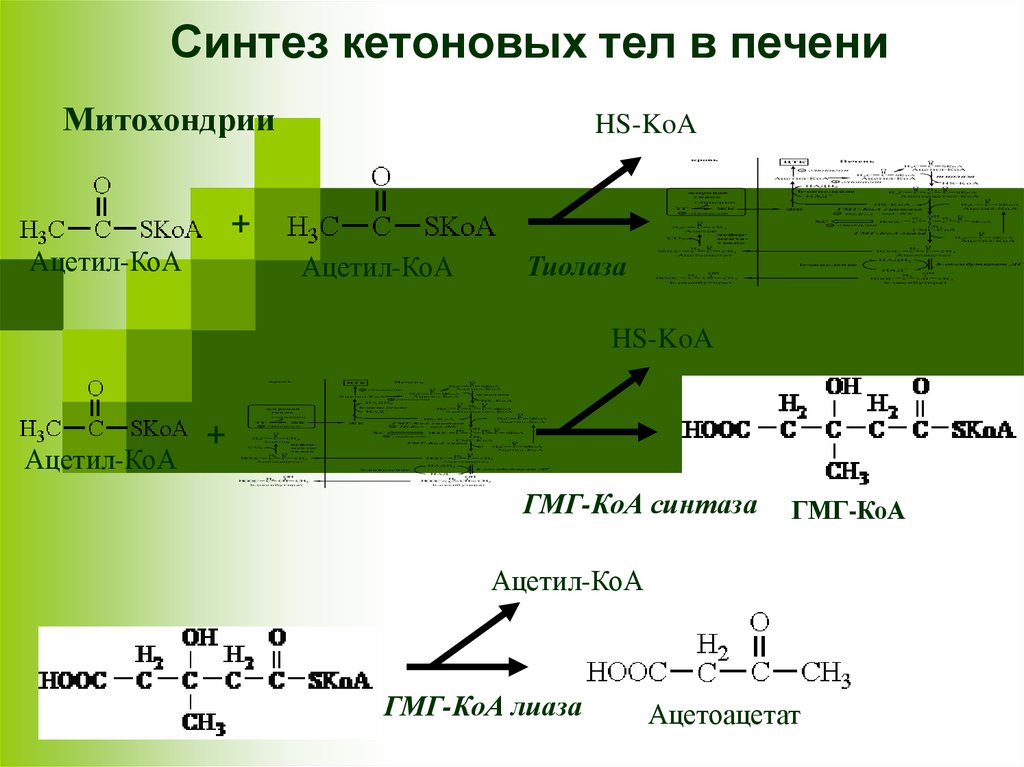
Синтез кетоновых тел в печениМитохондрии
HS-KoA
кровь
Печень
ЦТК
O
H3C
H3C
C
SKoA
ЖК
ТГ
глюкагон
+
H3C
CO2
Ацетил-КоА
Тиолаза
Ацетил-КоА
HOOC
C
HOOC
Печень
ЦТК
H3C
НАД+
CH 3
HOOC
b-оксибутират
НАДН2
жировая
ткань
глицерин
Ацетил-КоА
+
ЖК
ТГ
глюкагон
H2
C
H3C
CO2
HOOC
C
CH 3
Ацетон
неферментативно
H2
C
ЖК
H2
C
HOOC
глюкагон
CH 3
C
SKoA
Ацетил-КоА
C
O
H2
C
C
SKoA
CH 3
O
H3C
C
SKoA
Ацетил-КоА
H2
C
HOOC
O
C
CH 3
Ацетоацетат
НАДН2
b-оксибутират ДГ
НАД+
OH
CH
SKoA
H3C
OH
ГМГ-КоА
ГМГ-КоА лиаза
b-окисление
H2
C
C
O
инд. ЖК
HS-KoA
CH 3
b-оксибутират
O
Ацетоацетил-КоА
O
C
HS- KoA
C
H2
C
HS- KoA
ГМГ-КоА синтаза
Ацетоацетат
HOOC
H3C
НАД+
SKoA
тиолаза
O
b-окисление
ХС
O
C
SKoA
HOOC
H2
C
OH
CH
CH 3
b-оксибутират
ГМГ-КоА синтаза
ГМГ-КоА
Ацетил-КоА
ГМГ-КоА лиаза
C
SKoA
SKoA
O
C
SKoA
O
C
CH 3
OH
CH
CH 3
b-оксибутират
Ацетил-КоА
C
Ацетил-КоА
глюкагон
C
Ацетоацетат
НАДН2
b-оксибутират ДГ
O
H3C
O
глюкагон
Ацетил-КоА
C
Ацетил-КоА
O
H2
C
Ацетил-КоА
H2
C
HS-KoA
кровь
H3C
OH
CH 3
HOOC
OH
CH
SKoA
H3C
CH 3
b-окисление
H2
C
C
ГМГ-КоА
ГМГ-КоА лиаза
O
Ацетоацетат
HOOC
H2
C
глюкагон
CH 3
O
H2
C
O
инд. ЖК
HS-KoA
ХС
C
C
Ацетоацетил-КоА
HS- KoA
ГМГ-КоА синтаза
ЖК
Ацетон
неферментативно
H2
C
H3C
НАД+
O
HS- KoA
O
b-окисление
SKoA
тиолаза
Ацетил-КоА
глюкагон
НАДН2
жировая
ткань
глицерин
C
Ацетил-КоА
O
глюкагон
Ацетил-КоА
Ацетоацетат
39.
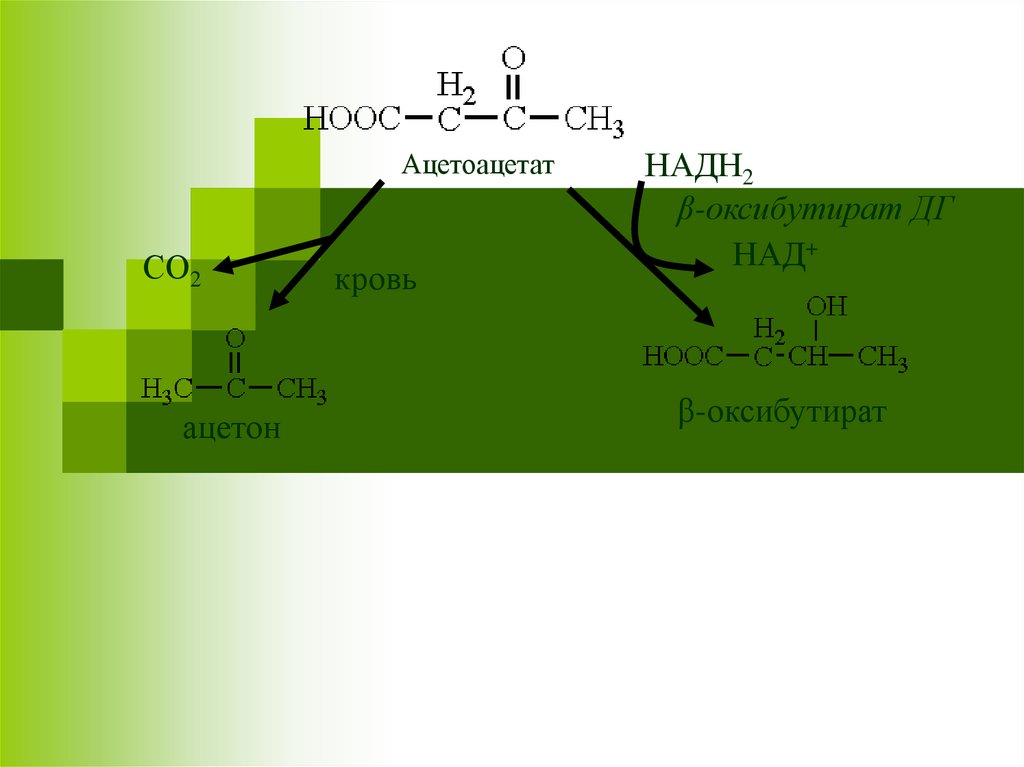
АцетоацетатСО2
ацетон
кровь
НАДН2
β-оксибутират ДГ
НАД+
β-оксибутират
40.
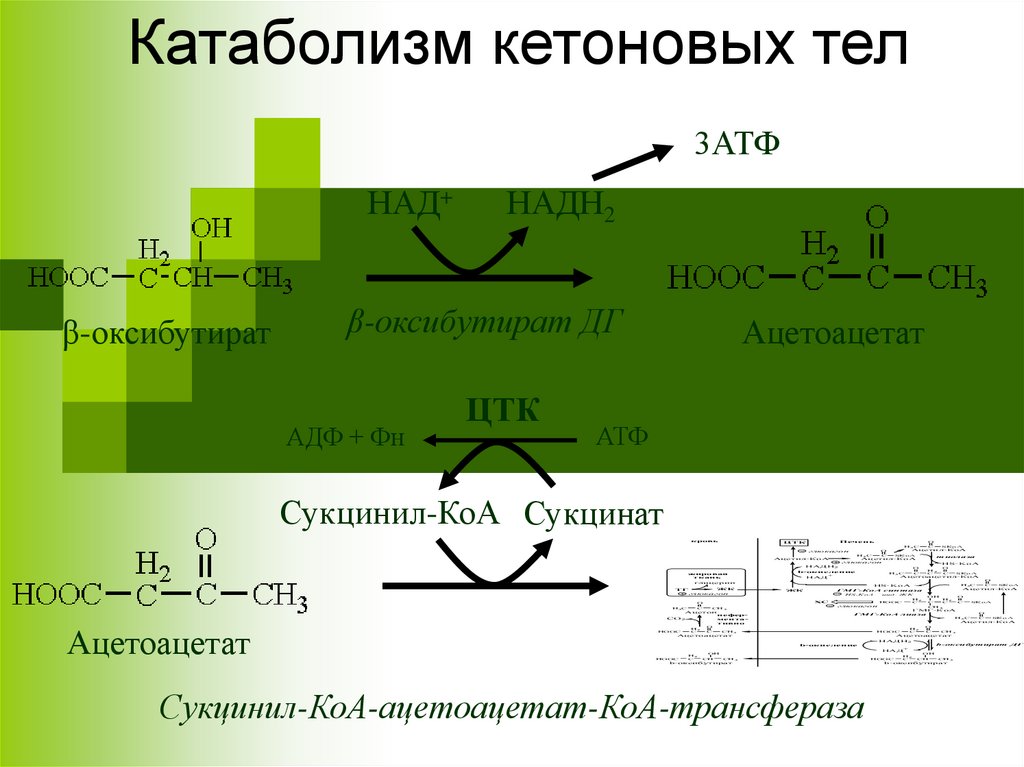
Катаболизм кетоновых тел3АТФ
НАД+
β-оксибутират
НАДН2
β-оксибутират ДГ
АДФ + Фн
ЦТК
Ацетоацетат
АТФ
Сукцинил-КоА Сукцинат
кровь
Печень
ЦТК
O
H3C
H3C
C
SKoA
ЖК
ТГ
глюкагон
Ацетоацетат
HOOC
C
CH 3
Ацетон
неферментативно
H2
C
H2
C
HOOC
глюкагон
SKoA
H3C
C
SKoA
Ацетил-КоА
OH
C
O
H2
C
C
SKoA
CH 3
ГМГ-КоА
ГМГ-КоА лиаза
O
H3C
C
SKoA
Ацетил-КоА
CH 3
H2
C
HOOC
CH 3
b-оксибутират
Сукцинил-КоА-ацетоацетат-КоА-трансфераза
O
C
CH 3
Ацетоацетат
НАДН2
b-оксибутират ДГ
НАД+
OH
CH
C
O
инд. ЖК
HS-KoA
b-окисление
H2
C
O
H2
C
HS- KoA
ГМГ-КоА синтаза
ЖК
O
C
C
Ацетоацетил-КоА
Ацетоацетат
HOOC
H3C
НАД+
ХС
O
H3C
CO2
HS- KoA
O
b-окисление
SKoA
тиолаза
Ацетил-КоА
глюкагон
НАДН2
жировая
ткань
глицерин
C
Ацетил-КоА
O
глюкагон
Ацетил-КоА
HOOC
H2
C
OH
CH
CH 3
b-оксибутират
41.
кровьПечень
ЦТК
O
H3C
H3C
C
SKoA
H3C
C
H2
C
ХС
H2
C
HOOC
C
H2
C
C
SKoA
SKoA
O
C
SKoA
O
C
CH 3
Ацетоацетат
НАДН2
b-оксибутират ДГ
НАД+
CH 3
HOOC
H2
C
OH
CH
CH 3
b-оксибутират
Тиолаза
Ацетил-КоА
О2
C
Ацетил-КоА
O
Ацетил-КоА
b-оксибутират
HS-KoA
H3C
OH
H3C
CH 3
OH
CH
SKoA
CH 3
O
b-окисление
H2
C
C
ГМГ-КоА
ГМГ-КоА лиаза
Ацетоацетат
HOOC
H2
C
HOOC
глюкагон
CH 3
C
O
H2
C
O
инд. ЖК
HS-KoA
Ацетон
неферментативно
HOOC
C
Ацетоацетил-КоА
HS- KoA
ГМГ-КоА синтаза
ЖК
O
CO2
H3C
НАД+
ЖК
ТГ
глюкагон
HS- KoA
O
b-окисление
SKoA
тиолаза
Ацетил-КоА
глюкагон
НАДН2
жировая
ткань
глицерин
C
Ацетил-КоА
O
глюкагон
Ацетил-КоА
Ацетил-КоА
О2
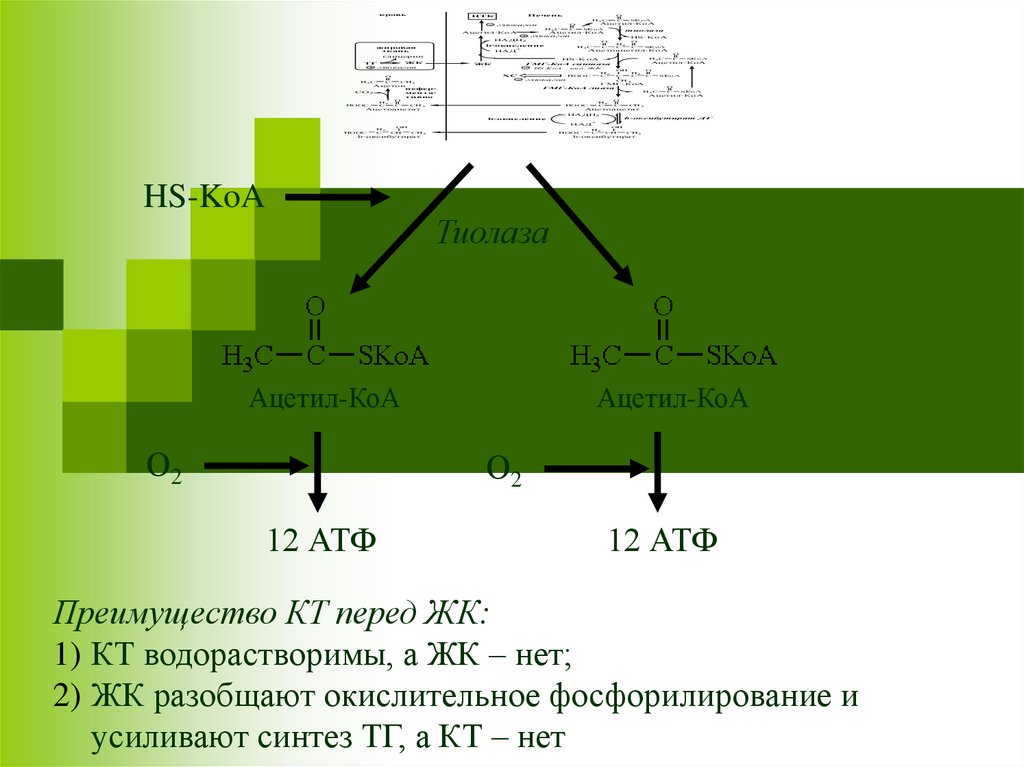
12 АТФ
12 АТФ
Преимущество КТ перед ЖК:
1) КТ водорастворимы, а ЖК – нет;
2) ЖК разобщают окислительное фосфорилирование и
усиливают синтез ТГ, а КТ – нет
42.
НормыСодержание КТ в крови:
30 мг/л – норма;
300-500 мг/л – при голодании;
3-4 г/л – при сахарном диабете;
43.
ХОЛЕСТЕРИНХолестерин (ХС) — стероид, характерный только для
животных организмов.
В сутки в организме синтезируется 1г ХС.
ХС синтезируется в печени (50%), тонком кишечнике (1520%), коже, коре надпочечников, половых железах.
С пищей поступает в сутки 0,3—0,5г ХС.
Биологическая роль ХС
входит в состав мембран клеток, увеличивает их
электроизоляционные свойства, придает жесткость и
прочность;
В мембране защищает полиненасыщенные ЖК от
окисления;
из ХС синтезируются жёлчные кислоты, стероидные
гормоны, витамин Д3
является компонентом желчи, участвует в
переваривании липидов.
44.
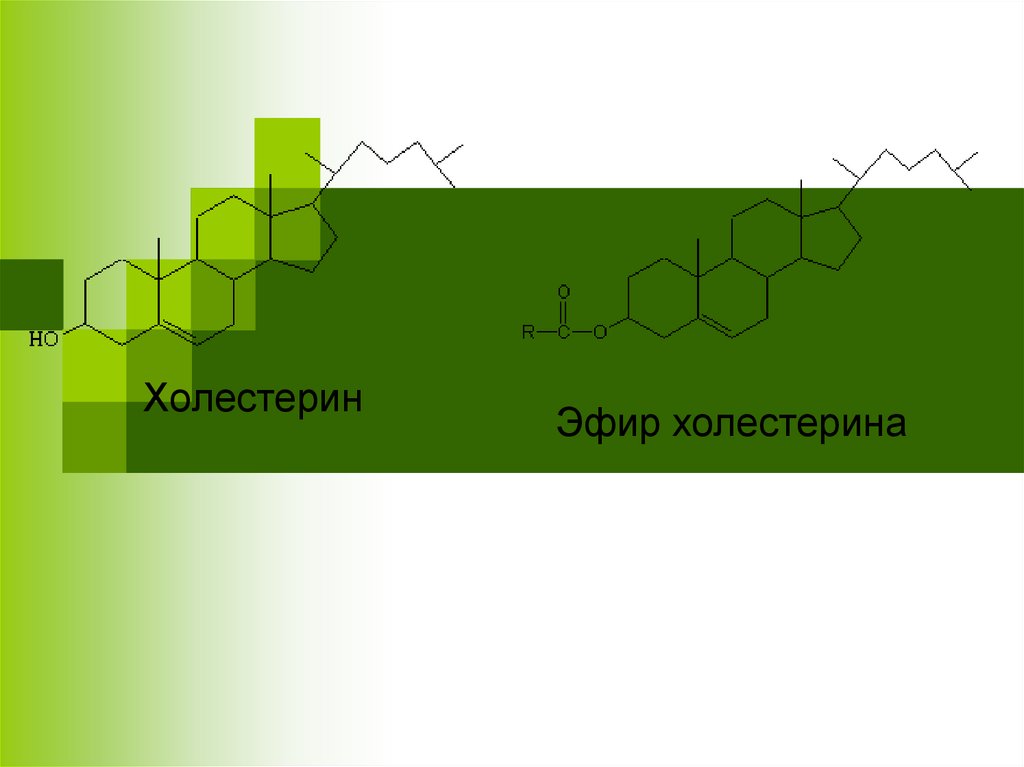
ХолестеринЭфир холестерина
45.
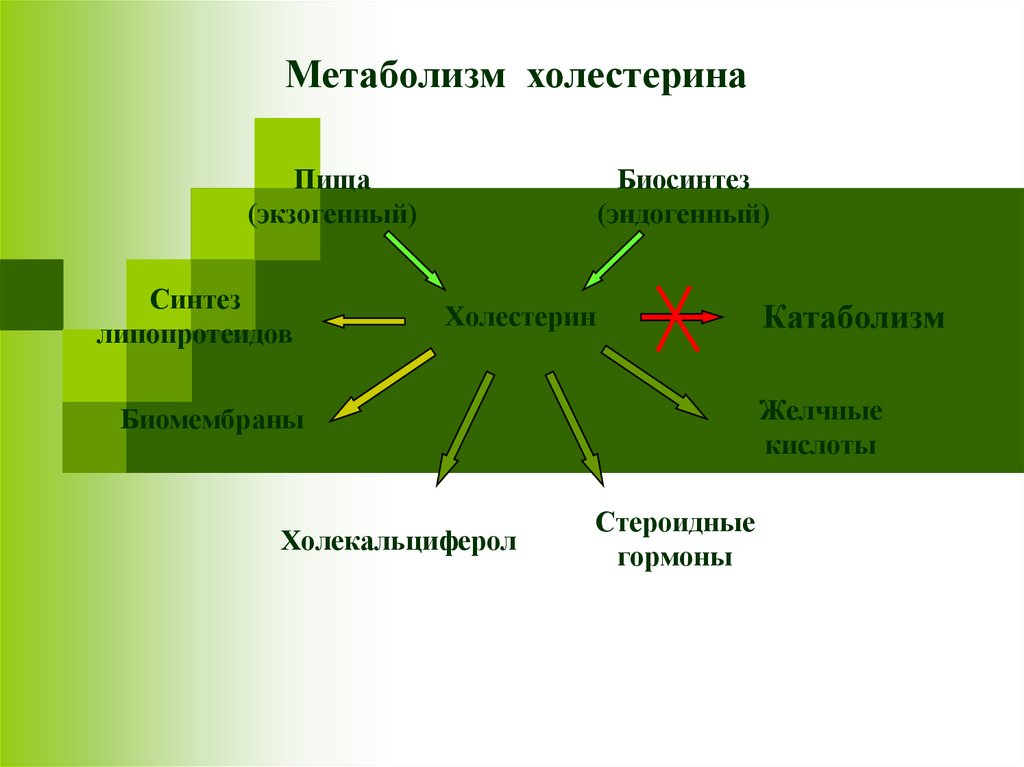
Метаболизм холестеринаПища
(экзогенный)
Синтез
липопротеидов
Биосинтез
(эндогенный)
Холестерин
Желчные
кислоты
Биомембраны
Холекальциферол
Катаболизм
Стероидные
гормоны
46.
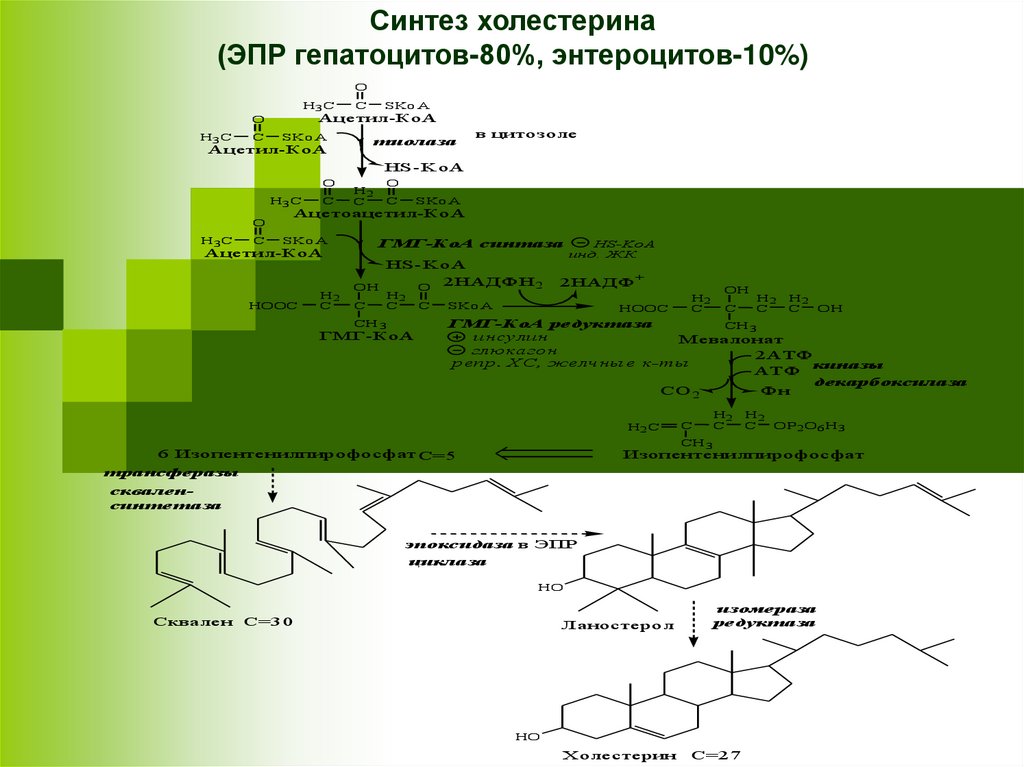
СИНТЕЗ ХСпроисходит в цитозоле и ЭПР клеток.
Это один из самых длинных метаболических путей в
организме человека (около 100 последовательных
реакций).
3 этапа:
I этап - образование мевалоната (мевалоновой
кислоты).
Две молекулы ацетил-КоА конденсируются тиолазой с
образованием ацетоацетил-КоА;
Гидроксиметилглутарил-КоА-синтаза (ГМГ-КоА)
присоединяет третий ацетильный остаток к
ацетоацетил-КоА с образованием ГМГ-КоА.
ГМГ-КоА-редуктаза восстанавливает ГМГ-КоА до
мевалоната с использованием 2 молекул НАДФH2.
47.
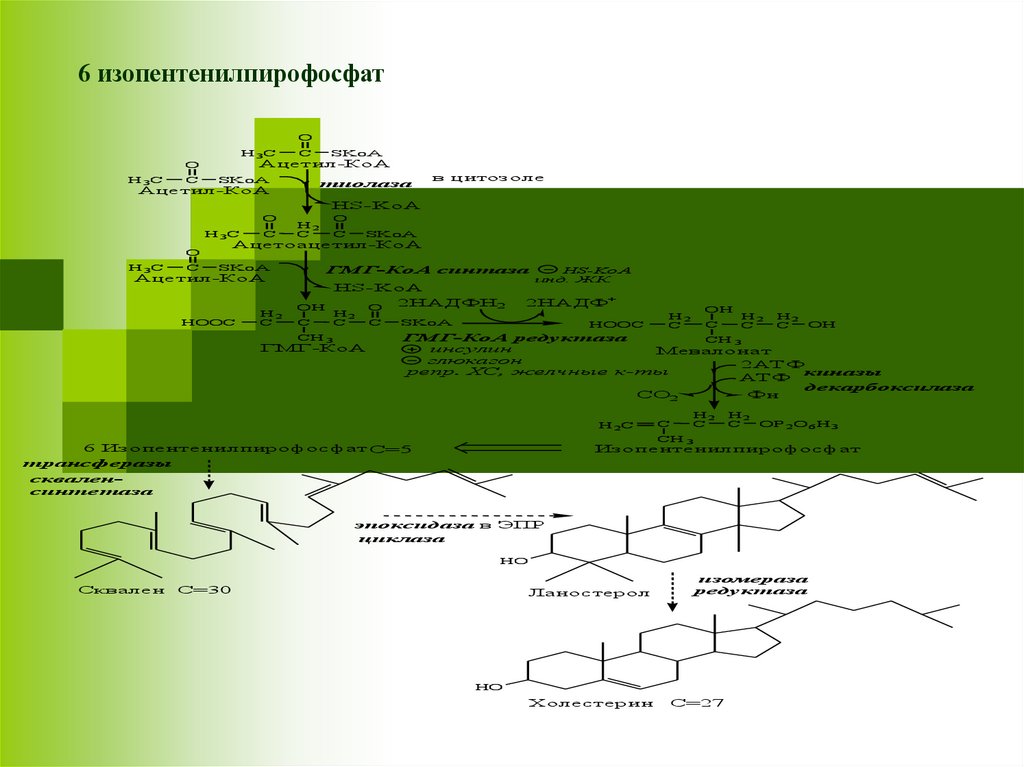
II этап - образование скваленаМевалонат превращается в изопреноидную
структуру — изопентенилпирофосфат (5
атомов С);
2 изопентенилпирофосфата конденсируются в
геранилпирофосфат (10 атомов С);
Присоединение изопентенилпирофосфата к
геранилпирофосфату дает
фарнезилпирофосфат (15 атомов С).
2 фарнезилпирофосфата конденсируются в
сквален (15 атомов С).
48.
III этап - образование ХССквален циклазой превращается в
ланостерин, (4 цикла и 30 атомов С).
Далее происходит 20
последовательных реакций,
превращающих ланостерин в ХС (27
атомов С).
49.
Синтез холестерина(ЭПР гепатоцитов-80%, энтероцитов-10%)
O
H3C
H3C
C
C
SKoA
Ацетил-КоА
O
SKoA
тиолаза
Ацетил-КоА
в цитозоле
HS-KoA
O
H3C
O
H3C
C
C
H2
C
O
C
SKoA
Ацетоацетил-КоА
SKoA
Ацетил-КоА
HOOC
H2
C
ГМГ-КоА синтаза
HS-KoA
OH
O 2НАДФН2
C
H2
C
CH 3
ГМГ-КоА
C
HS-KoA
инд. ЖК
2НАДФ+
SKoA
HOOC
OH
H2
C
C
H2
C
H2
OH
C
ГМГ-КоА редуктаза
CH 3
инсулин
Мевалонат
глюкагон
2АТФ
репр. ХС, желчные к-ты
АТФ киназы
декарбоксилаза
Фн
СО2
H2C
C
H2
C
H2
OP 2O6H3
C
CH 3
6 Изопентенилпирофосфат С=5
трансферазы
скваленсинтетаза
Изопентенилпирофосфат
эпоксидаза в ЭПР
циклаза
HO
Сквален С=30
Ланостерол
изомераза
редуктаза
HO
Холестерин
С=27
50.
6 изопентенилпирофосфатO
H3C
H3C
C
C
SKoA
Ацетил-КоА
O
SKoA
тиолаза
Ацетил-КоА
в цитозоле
HS- KoA
O
H3C
O
H3C
C
C
H2
C
O
C
SKoA
Ацетоацетил-КоА
SKoA
Ацетил-КоА
HOOC
H2
C
ГМГ-КоА синтаза
HS- KoA
2НАДФН2
OH
O
C
H2
C
CH 3
ГМГ-КоА
C
HS-KoA
инд. ЖК
2НАДФ+
SKoA
HOOC
OH
H2
C
C
H2
C
H2
C
H2C
C
H2
C
H2
C
OP 2O6H3
CH 3
6 Изопентенилпирофосфат С=5
трансферазы
скваленсинтетаза
Изопентенилпирофосфат
эпоксидаза в ЭПР
циклаза
HO
Сквален
OH
ГМГ-КоА редуктаза
CH 3
инсулин
Мевалонат
глюкагон
2АТФ
репр. ХС, желчные к-ты
АТФ киназы
декарбоксилаза
Фн
СО2
С=30
Ланостерол
изомераза
редуктаза
HO
Холестерин
С=27
51.
Этерификация ХСХС образует с ЖК сложные эфиры (ЭХС),
которые более гидрофобны, чем сам ХС.
В клетках эту реакцию катализирует
АХАТ(АцилКоА:ацилхолестеринтрансферазой):
ХС + Ацил-КоА → ЭХС + HSKoA
ЭХС формирует в цитоплазме липидные капли,
которые являются формой хранения ХС. По
мере необходимости ЭХС гидролизуются
холестеролэстеразой на ХС и ЖК.
ЭХС синтезируются в крови в ЛПВП под
действием ЛХАТ (лецитин:
холестеролацилтрансферазой):
ХС + лецитин → ЭХС + лизолецитин
52.
Регуляция ключевого фермента синтеза ХСГидроксиМетилГлутарил-КоА-редуктазы
Инсулин через дефосфорилирование активирует
фермент.
Повышение концентрации ацетил-КоА стимулирует
синтез ХС.
Синтез ХС активируется при питании углеводами и
снижается при голодании.
Глюкагон через фосфорилирование ингибирует
фермент.
ХС, желчные кислоты (в печени) репрессируют ген ГМГКоА-редуктазы.
53.
Выведение ХС из организмаВ сутки из организма выводится 1,0г - 1,3г ХС.
С желчью (0,5 - 0,7 г/сут) - в виде жёлчных
кислот.
Часть ХС в кишечнике под действием ферментов
бактерий восстанавливается по двойной связи,
образуя холестанол и копростанол.
С кожным салом в сутки выделяется 0,1г ХС.