 в клетках печени, кишечника, жировой ткани, в состоянии покоя или после еды")
")
")


")



 biology
biology chemistry
chemistrySimilar presentations:



")

")




Синтез липидов
1. СИНТЕЗ ЖИРНЫХ КИСЛОТ (В ЦИТОЗОЛЕ) в клетках печени, кишечника, жировой ткани, в состоянии покоя или после еды
Транспортные пути из митохондрии в цитозольСИНТЕЗ ЖИРНЫХ КИСЛОТ (В ЦИТОЗОЛЕ)
в клетках печени, кишечника, жировой ткани, в
состоянии покоя или после еды
1. Цитратный путь
Митохондрии
цитратсинтаза
Ацетил-КоА + Оксалоацетат
Декарбоксилирование ПВК ,
окисление ЖК,
из кетогенных аминокислот
Малат-аспартатный
челнок
Декарбоксилируется в
пируват
Цитозоль
Цитрат
(выходит из матрикса Мt в цитозоль)
цитратлиаза
Оксалоацетат + Ацетил-КоА
Малатдегидрогеназа
(цитозоль)
НАДН+Н+
НАД+
Малат (цитозоль)
Синтез жирных
кислот
2. Карнитиновый путь (как и при переносе высших ЖК)
Ацетил-КоА + карнитин
Митохондрии
Ацетилкарнитин
(выходит из матрикса в цитозоль, где распадается, а карнитин
возвращается в Mt)
2. СИНТЕЗ МАЛОНИЛА-КоА ИЗ АЦЕТИЛА (реакции карбоксилирования ацетил-КоА катализирует фермент Ацетил-КоА-карбоксилаза)
I этап - АТФ-зависимая реакция карбоксилирования биотинаСО2 + АТФ + биотин-энзим
Н3РО4 + АДФ + карбоксибиотин-энзим
II этап - перенос карбоксильной группы на ацетил-КоА
карбоксибиотин-энзим + СН3-СО~S-KoA
ацетил-КоА
Суммарное уравнение
образования
малонил КоА
HOOС-CH2-CO~S-KoA + биотин-энзим
малонил-КоА
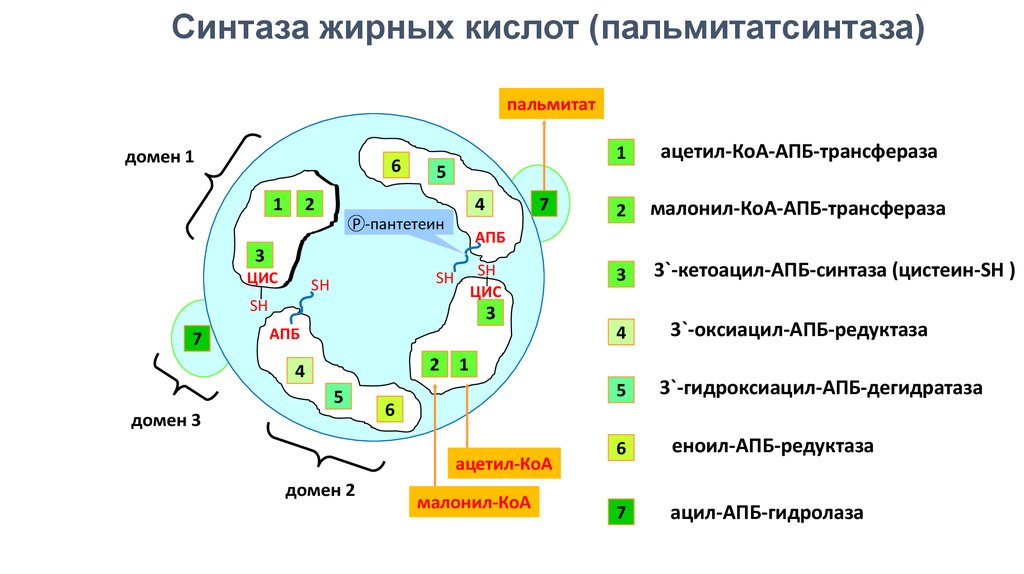
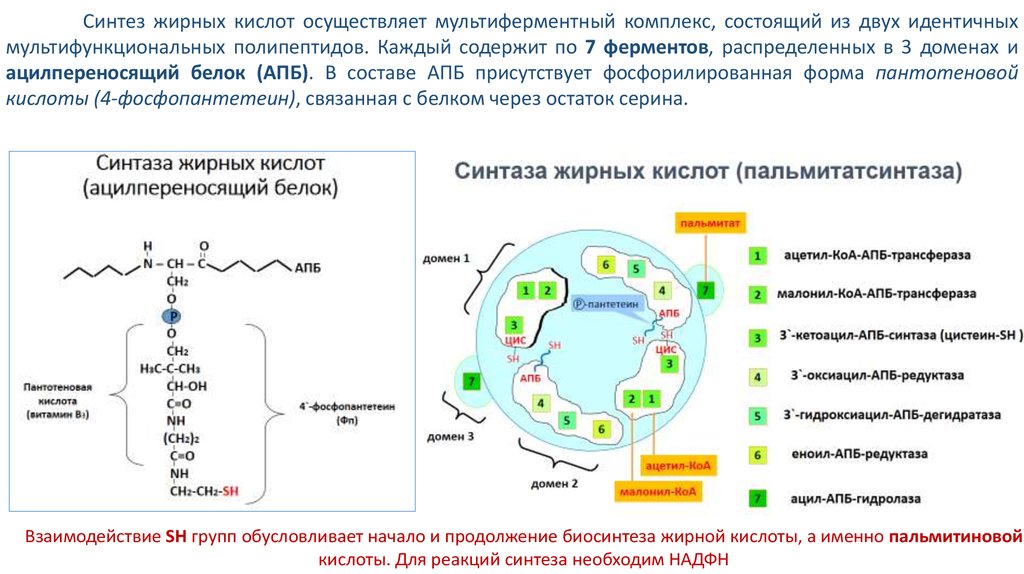
3. Синтаза жирных кислот (ацилпереносящий белок)
Пантотеноваякислота
(витамин В3)
Н
O
N – CH – C
CH2
O
P
O
CH2
H3C-C-CH3
CH-OH
C=O
NH
(CH2)2
C=O
NH
CH2-CH2-SH
АПБ
4`-фосфопантетеин
(Фп)
3
4.
Синтаза жирных кислот (пальмитатсинтаза)пальмитат
домен 1
6
11
4
Р -пантетеин
ЦИС
ЦИС
SH
7
7
2
малонил-КоА-АПБ-трансфераза
3
3`-кетоацил-АПБ-синтаза (цистеин-SH )
АПБ
SH
ЦИС
SH
SH
ацетил-КоА-АПБ-трансфераза
5
22
33
1
3
4
АПБ
2
4
5
домен 3
1
5
6
ацетил-КоА
домен 2
3`-оксиацил-АПБ-редуктаза
малонил-КоА
3`-гидроксиацил-АПБ-дегидратаза
6
еноил-АПБ-редуктаза
7
ацил-АПБ-гидролаза
5.
Синтез жирных кислот осуществляет мультиферментный комплекс, состоящий из двух идентичныхмультифункциональных полипептидов. Каждый содержит по 7 ферментов, распределенных в 3 доменах и
ацилпереносящий белок (АПБ). В составе АПБ присутствует фосфорилированная форма пантотеновой
кислоты (4-фосфопантетеин), связанная с белком через остаток серина.
Взаимодействие SH групп обусловливает начало и продолжение биосинтеза жирной кислоты, а именно пальмитиновой
кислоты. Для реакций синтеза необходим НАДФН
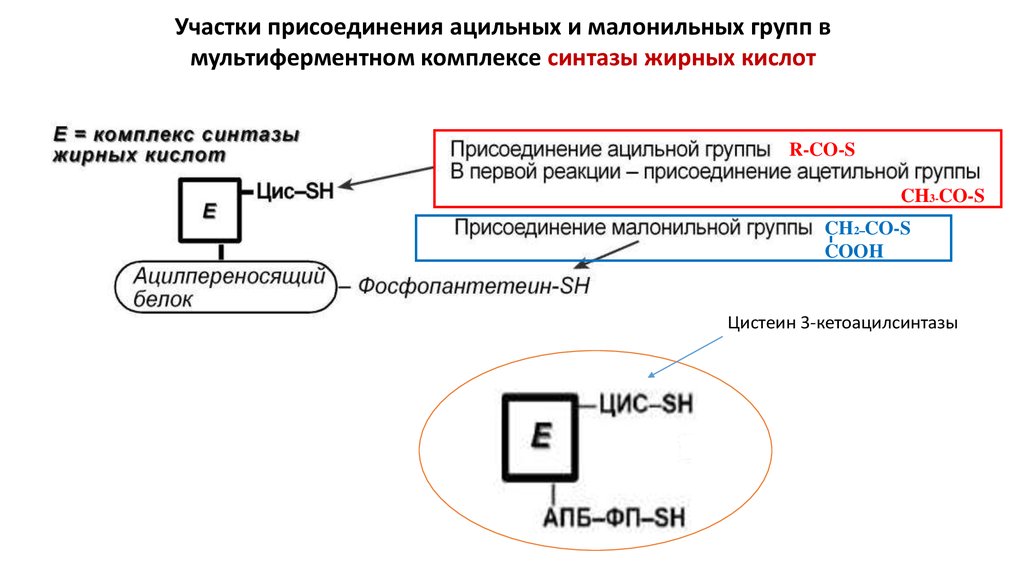
6.
Участки присоединения ацильных и малонильных групп вмультиферментном комплексе синтазы жирных кислот
R-CO-S
CH3-CO-S
CH2 ̶ CO-S
COOH
Цистеин 3-кетоацилсинтазы
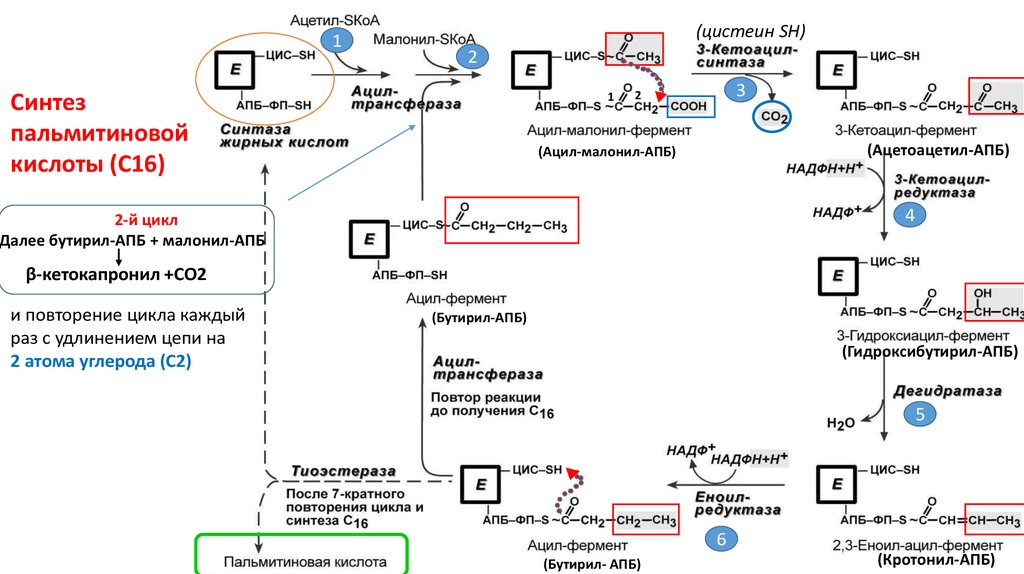
7.
1(цистеин SH)
2
Синтез
пальмитиновой
кислоты (С16)
1
3
2
(Ацетоацетил-АПБ)
(Ацил-малонил-АПБ)
4
2-й цикл
Далее бутирил-АПБ + малонил-АПБ
β-кетокапронил +СО2
и повторение цикла каждый
раз с удлинением цепи на
2 атома углерода (С2)
(Бутирил-АПБ)
(Гидроксибутирил-АПБ)
5
6
(Бутирил- АПБ)
(Кротонил-АПБ)
8.
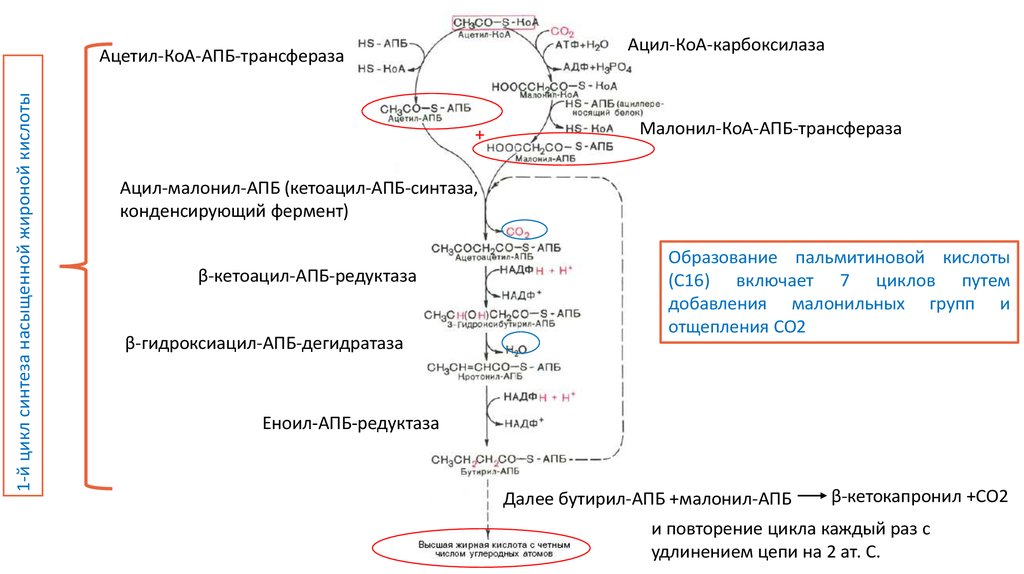
Ацил-КоА-карбоксилаза1-й цикл синтеза насыщенной жироной кислоты
Ацетил-КоА-АПБ-трансфераза
+
Малонил-КоА-АПБ-трансфераза
Ацил-малонил-АПБ (кетоацил-АПБ-синтаза,
конденсирующий фермент)
β-кетоацил-АПБ-редуктаза
β-гидроксиацил-АПБ-дегидратаза
Образование пальмитиновой кислоты
(С16) включает 7 циклов путем
добавления малонильных групп и
отщепления СО2
Еноил-АПБ-редуктаза
Далее бутирил-АПБ +малонил-АПБ
β-кетокапронил +СО2
и повторение цикла каждый раз с
удлинением цепи на 2 ат. С.
9.
Завершается синтез жирной кислоты отщеплением HS-АПБ от ацил-АПБ под влиянием фермента деацилазы(тиоэстераза)
Суммарное уравнение синтеза пальмитиновой кислоты
Индуктором биосинтеза пальмитатсинтазы и ацетил-SКоА-карбоксилазы ферментов является
инсулин. Усиление синтеза жирных кислот также достигается при поступлении цитрата и при наличии
малонил-SКоА.
Ингибируют синтез ЖК глюкагон и адреналин. Они вызывают липолиз (или поступление жирных
кислот извне), накопление ацил-SКоА в цитозоле, что автоматически уменьшает количество цитрата и
малонил-SКоА и препятствует синтезу новых жирных кислот.
10. УДЛИНЕНИЕ ЖИРНЫХ КИСЛОТ
H3C (CН 2)14 СО ~ SKoAпальмитоил-КоА
HOOC CН 2 СО ~ SKoA
малонил-КоА
НАДФН+Н+
конденсация
элонгаза
(ферментный
комплекс)
восстановление
дегидратация
восстановление
H3C
(CН 2)16 СО ~ SKoA
стеарил-КоА
деацилаза
Н2О
НS-КоА
H3C (CН 2)16 COOH
стеариновая кислота (стеарат)
Удлинение углеводородной цепи ЖК
происходит в эндоплазматическом ретикулуме
(ЭПР) с помощью присоединения малонил-КоА,
или в матриксе митохондрий путем
присоединения ацетил-КоА
11. БИОСИНТЕЗ НЕПРЕДЕЛЬНЫХ ЖИРНЫХ КИСЛОТ (в микросомах клеток печени и жировой ткани)
H3C(CН 2)14 СО ~ SKoA
пальмитоил-КоА
О2 + НАДФН+Н+
десатураза
2 Н2О + НАДФ+
H3C (CН 2)5 СН
СН
(CН 2)7 СО ~ SKoA
пальмитоолеил-КоА
Н2О
деацилаза
НS-КоА
H3C (CН 2)5 СН
СН
(CН 2)7 COOH
пальмитоолеиновая кислота
(пальмитоолеат)
Двойная связь животными клетками вводится не далее 9 атома углерода, поэтому -3 и -6-полиненасыщенные ЖК
синтезируются только из соответствующих предшественников. Например, арахидоновая кислота может образоваться в клетке только
при наличии линоленовой или линолевой кислот.
12.
13.
Глицерофосфатный путьпочки, стенка кишечника,
печень
Дигидроксиацетонфосфатный путь
жировая ткань, мышцы, печень
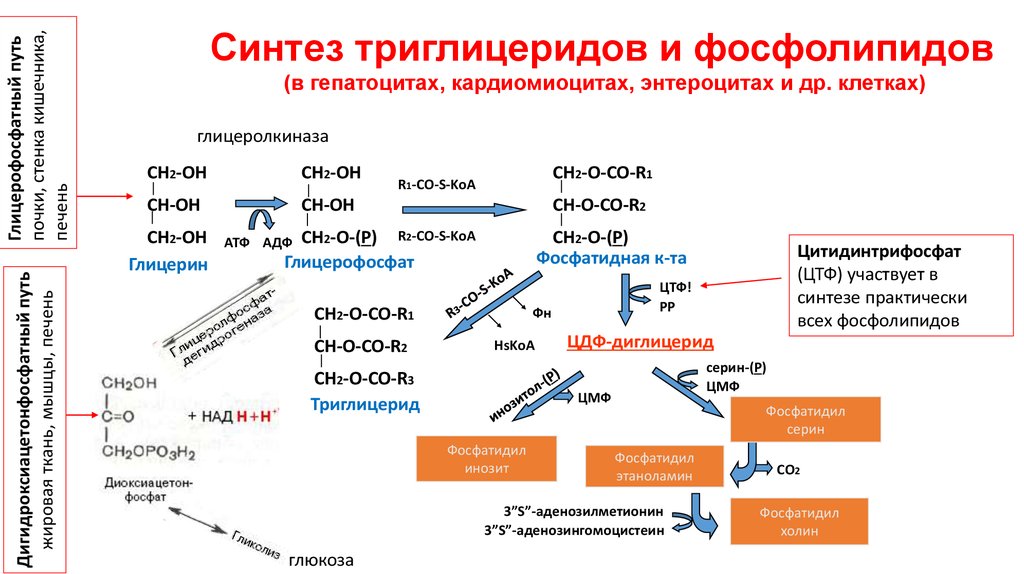
Синтез триглицеридов и фосфолипидов
(в гепатоцитах, кардиомиоцитах, энтероцитах и др. клетках)
глицеролкиназа
СН2-ОН
СН2-ОН
CH-OН
CH-OН
CH2-OН
Глицерин
СН2-О-СО-R1
R1-CO-S-KoA
CH-O-CO-R2
CH2-O-(Р) R2-CO-S-KoA
Глицерофосфат
CH2-O-(Р)
Фосфатидная к-та
АТФ АДФ
СН2-О-СО-R1
CH-O-CO-R2
ЦТФ!
РР
Фн
HsKoA
CH2-O-CO-R3
Триглицерид
ЦДФ-диглицерид
серин-(Р)
ЦМФ
ЦМФ
Фосфатидил
инозит
Фосфатидил
серин
Фосфатидил
этаноламин
3”S”-аденозилметионин
3”S”-аденозингомоцистеин
глюкоза
Цитидинтрифосфат
(ЦТФ) участвует в
синтезе практически
всех фосфолипидов
СО2
Фосфатидил
холин
14.
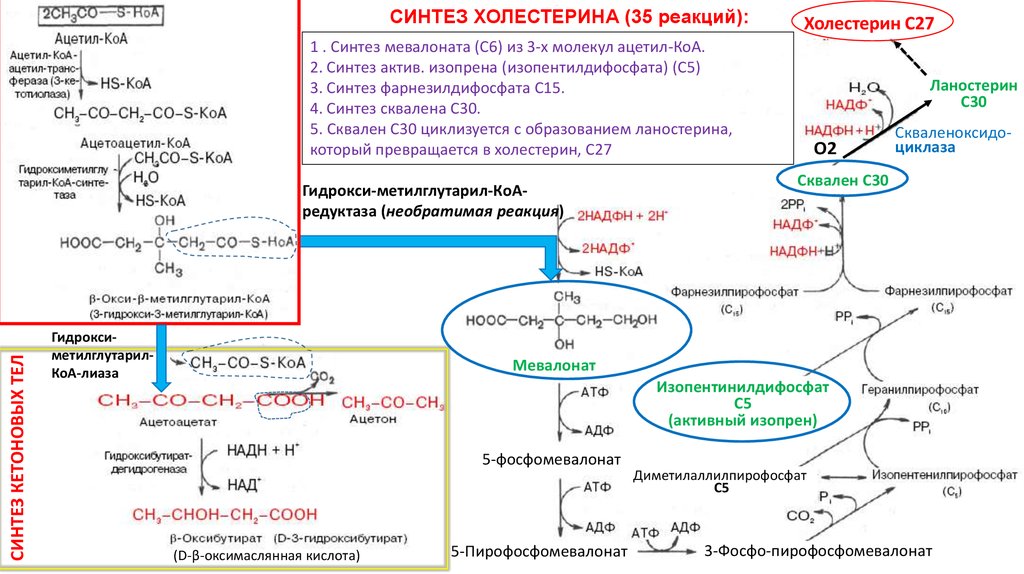
СИНТЕЗ ХОЛЕСТЕРИНА (35 реакций):Холестерин С27
1 . Синтез мевалоната (С6) из 3-х молекул ацетил-КоА.
2. Синтез актив. изопрена (изопентилдифосфата) (С5)
3. Синтез фарнезилдифосфата С15.
4. Синтез сквалена С30.
5. Сквален С30 циклизуется с образованием ланостерина,
который превращается в холестерин, С27
СИНТЕЗ КЕТОНОВЫХ ТЕЛ
Гидрокси-метилглутарил-КоАредуктаза (необратимая реакция)
ГидроксиметилглутарилКоА-лиаза
Ланостерин
С30
О2
Скваленоксидоциклаза
Сквален С30
Мевалонат
Изопентинилдифосфат
С5
(активный изопрен)
5-фосфомевалонат
(D-β-оксимаслянная кислота)
5-Пирофосфомевалонат
Диметилаллилпирофосфат
С5
3-Фосфо-пирофосфомевалонат
15.
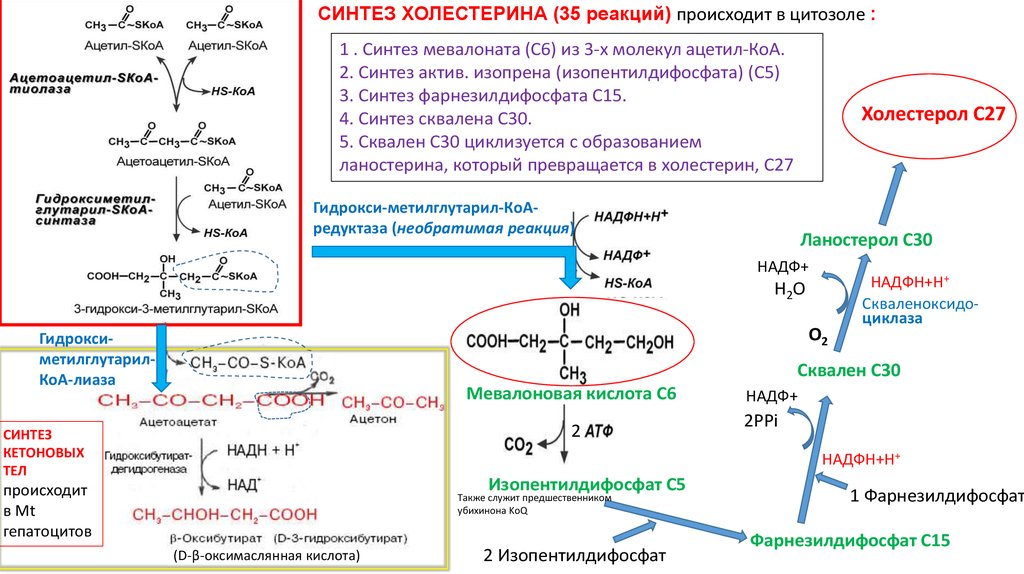
СИНТЕЗ ХОЛЕСТЕРИНА (35 реакций) происходит в цитозоле :1 . Синтез мевалоната (С6) из 3-х молекул ацетил-КоА.
2. Синтез актив. изопрена (изопентилдифосфата) (С5)
3. Синтез фарнезилдифосфата С15.
4. Синтез сквалена С30.
5. Сквален С30 циклизуется с образованием
ланостерина, который превращается в холестерин, С27
Холестерол С27
Гидрокси-метилглутарил-КоАредуктаза (необратимая реакция)
Ланостерол С30
НАДФ+
Н2О
О2
ГидроксиметилглутарилКоА-лиаза
НАДФН+Н+
Скваленоксидоциклаза
Сквален С30
Мевалоновая кислота С6
2
СИНТЕЗ
КЕТОНОВЫХ
ТЕЛ
НАДФ+
2РРi
НАДФН+Н+
Изопентилдифосфат С5
происходит
в Mt
гепатоцитов
Также служит предшественником
убихинона KoQ
(D-β-оксимаслянная кислота)
2 Изопентилдифосфат
1 Фарнезилдифосфат
Фарнезилдифосфат С15
16.
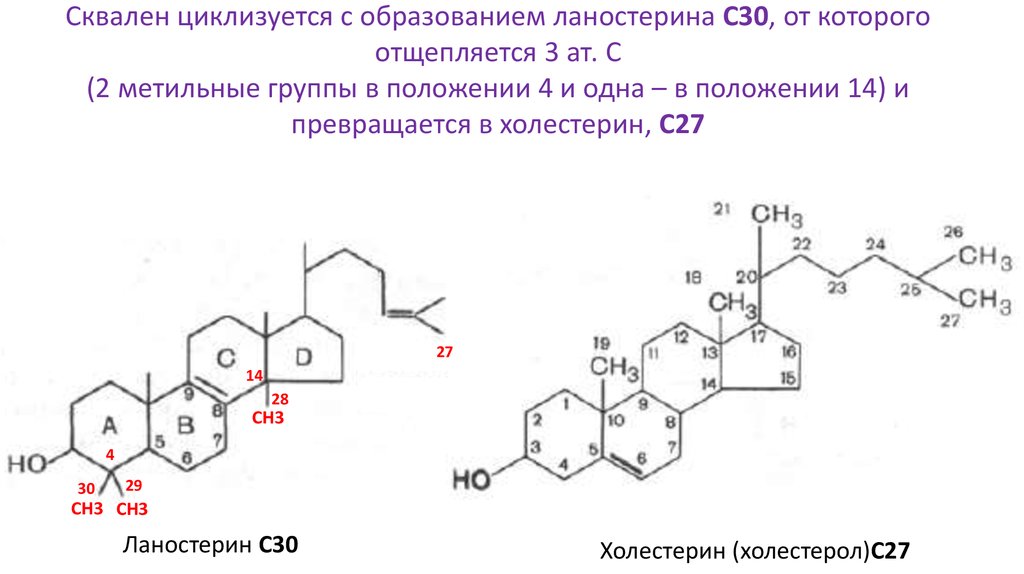
Сквален циклизуется с образованием ланостерина С30, от которогоотщепляется 3 ат. С
(2 метильные группы в положении 4 и одна – в положении 14) и
превращается в холестерин, С27
27
14
28
СН3
4
30
29
СН3 СН3
Ланостерин С30
Холестерин (холестерол)С27
17.
состав липопротеинов, %липопротеин
ТАГ Х + ЭХ апопротеины ФЛ
ХМ
88
5-7
2
3
ЛПОНП
55
17
10
18
ЛППП
26
38
11
25
ЛПНП
7
50
22
21
ЛПВП
3
20
50
27
18.
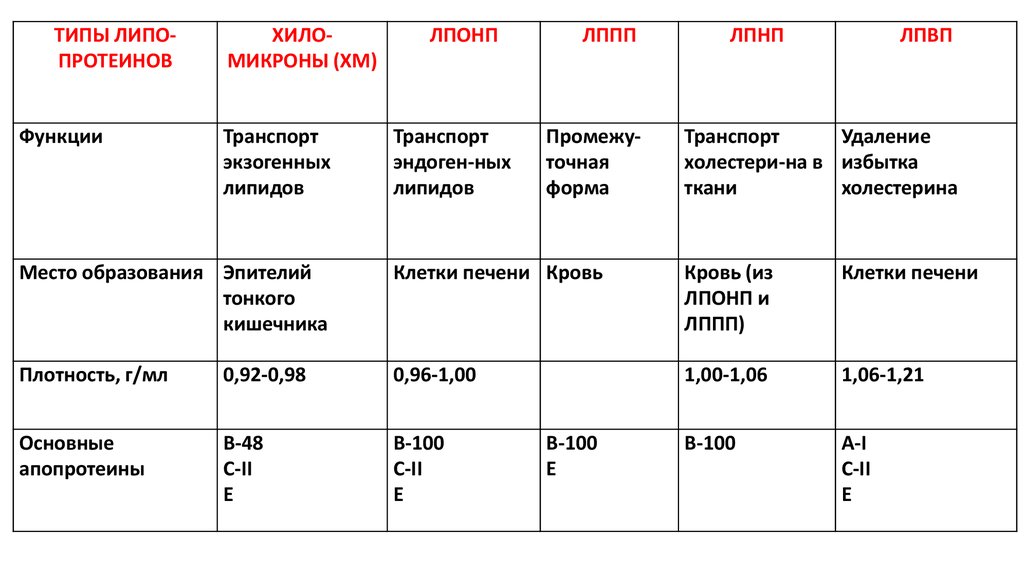
ТИПЫ ЛИПОПРОТЕИНОВФункции
ХИЛОМИКРОНЫ (ХМ)
Транспорт
экзогенных
липидов
ЛПОНП
Транспорт
эндоген-ных
липидов
ЛППП
Промежуточная
форма
ЛПНП
ЛПВП
Транспорт
Удаление
холестери-на в избытка
ткани
холестерина
Место образования Эпителий
тонкого
кишечника
Клетки печени Кровь
Кровь (из
ЛПОНП и
ЛППП)
Клетки печени
Плотность, г/мл
0,92-0,98
0,96-1,00
1,00-1,06
1,06-1,21
Основные
апопротеины
В-48
С-II
E
В-100
С-II
E
В-100
А-I
С-II
E
В-100
E
19.
В ЛПВП активно протекает реакция при участии лецитин:холестерол-ацилтрансферазы (ЛХАТреакция). В этой реакции остаток ненасыщенной жирной кислоты переносится от фосфатидилхолина (ФХ) насвободный холестерол (ХС) с образованием лизофосфатидилхолина и эфиров ХС.
3
20.
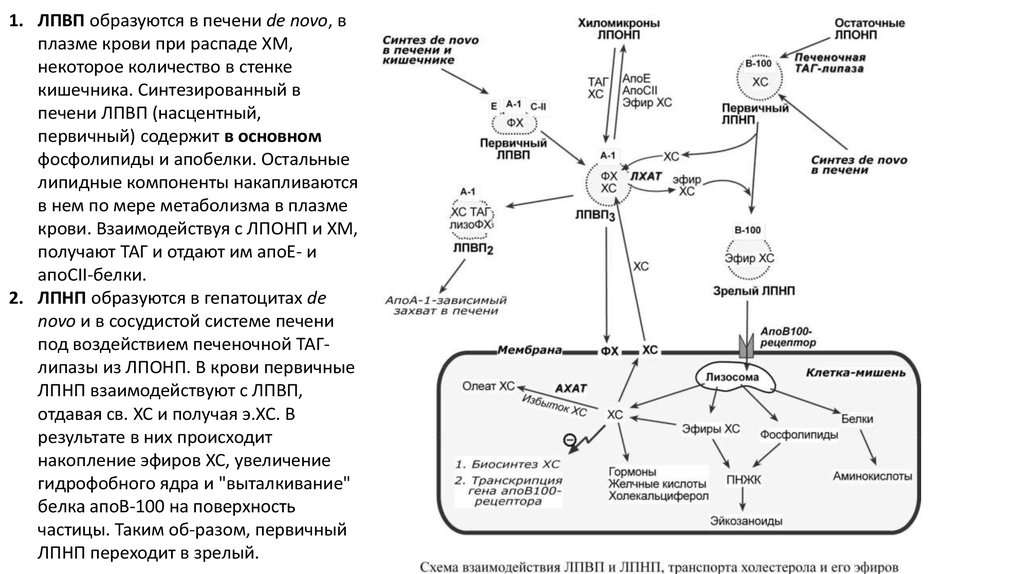
1. ЛПВП образуются в печени de novo, вплазме крови при распаде ХМ,
некоторое количество в стенке
кишечника. Синтезированный в
печени ЛПВП (насцентный,
первичный) содержит в основном
фосфолипиды и апобелки. Остальные
липидные компоненты накапливаются
в нем по мере метаболизма в плазме
крови. Взаимодействуя с ЛПОНП и ХМ,
получают ТАГ и отдают им апоЕ- и
апоСII-белки.
2. ЛПНП образуются в гепатоцитах de
novo и в сосудистой системе печени
под воздействием печеночной ТАГлипазы из ЛПОНП. В крови первичные
ЛПНП взаимодействуют с ЛПВП,
отдавая св. ХС и получая э.ХС. В
результате в них происходит
накопление эфиров ХС, увеличение
гидрофобного ядра и "выталкивание"
белка апоВ-100 на поверхность
частицы. Таким об-разом, первичный
ЛПНП переходит в зрелый.