








 biology
biologySimilar presentations:






")



Анатомия и физиология человека. Лекция 1
1.
АНАТОМИЯ И ФИЗИОЛОГИЯ ЧЕЛОВЕКАЛЕКЦИЯ 1
•Потенциал покоя
Гайдуков Александр Евгеньевич
МФТИ 2025
2.
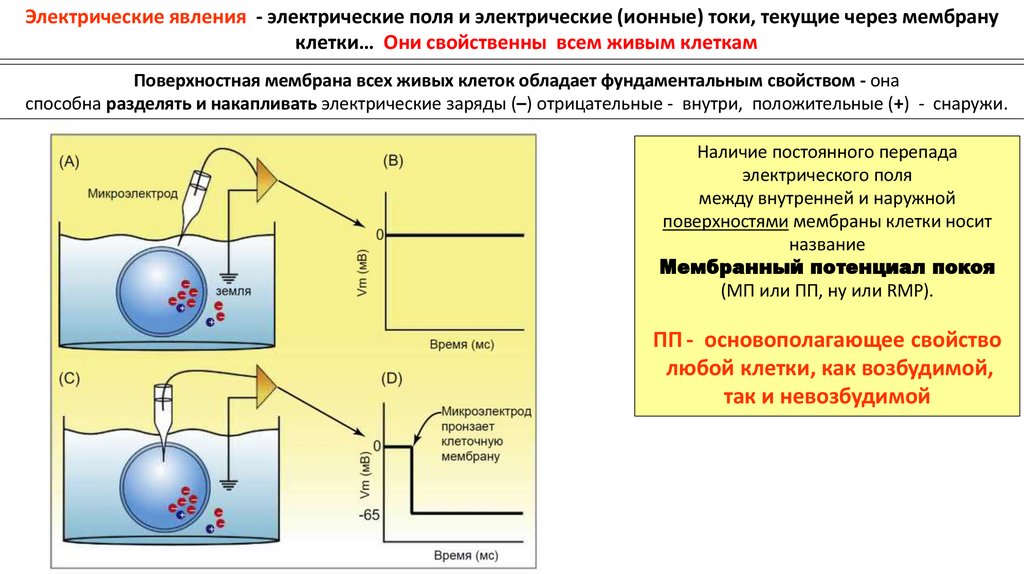
Электрические явления - электрические поля и электрические (ионные) токи, текущие через мембрануклетки… Они свойственны всем живым клеткам
Поверхностная мембрана всех живых клеток обладает фундаментальным свойством - она
способна разделять и накапливать электрические заряды (–) отрицательные - внутри, положительные (+) - снаружи.
Наличие постоянного перепада
электрического поля
между внутренней и наружной
поверхностями мембраны клетки носит
название
Мембранный потенциал покоя
(МП или ПП, ну или RMP).
ПП - основополагающее свойство
любой клетки, как возбудимой,
так и невозбудимой
3.
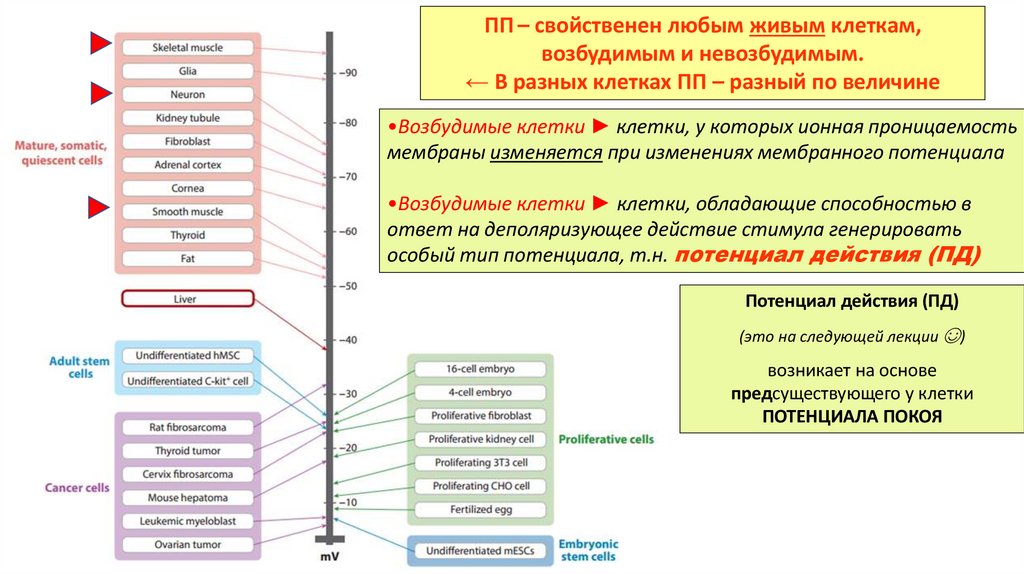
ПП – свойственен любым живым клеткам,возбудимым и невозбудимым.
← В разных клетках ПП – разный по величине
•Возбудимые клетки ► клетки, у которых ионная проницаемость
мембраны изменяется при изменениях мембранного потенциала
•Возбудимые клетки ► клетки, обладающие способностью в
ответ на деполяризующее действие стимула генерировать
особый тип потенциала, т.н. потенциал действия (ПД)
Потенциал действия (ПД)
(это на следующей лекции ☺)
возникает на основе
предсуществующего у клетки
ПОТЕНЦИАЛА ПОКОЯ
4.
Ключевой Вопрос: Как объяснить пространственное разделение электрических зарядовсосредоточение положительных зарядов снаружи мембран клеток, а отрицательных – внутри,
т.е. наличие разности потенциалов между наружной и внутренней поверхностью мембраны клеток?
Всё это похоже на диффузионный потенциал
Именно его изучал Вальтер Нернст
Возникает в растворе электролита при диффузии анионов и катионов солей
Диффузия заряженных частиц – противоионов происходит с разной скоростью, в соответствии с их подвижностью (U) в растворе электролита
Анионы
Катионы
Соль
К+А-
то, что заставляет
диффундировать ионы
RT u1 u 2 [Ci ]2
Vдиф
ln
zF u1 u 2 [Ci ]1
C1
С2
Фронт анионов
… пропорционально
логарифму отношения
концентраций
U - подвижность иона
cм/сек : В/см = см2 / В·cек
5.
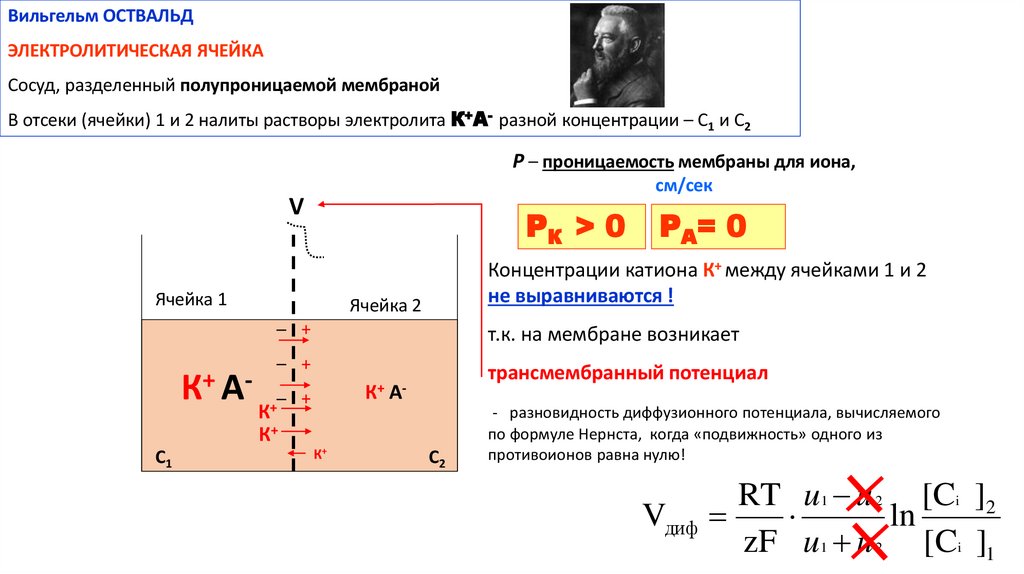
Вильгельм ОСТВАЛЬДЭЛЕКТРОЛИТИЧЕСКАЯ ЯЧЕЙКА
Сосуд, разделенный полупроницаемой мембраной
В отсеки (ячейки) 1 и 2 налиты растворы электролита К+А- разной концентрации – С1 и С2
Р – проницаемость мембраны для иона,
см/сек
V
РК > 0
Ячейка 1
Концентрации катиона К+ между ячейками 1 и 2
не выравниваются !
Ячейка 2
– +
К + А-
т.к. на мембране возникает
– +
трансмембранный потенциал
К+ А-
– +
К+
К+
С1
PА= 0
К+
С2
- разновидность диффузионного потенциала, вычисляемого
по формуле Нернста, когда «подвижность» одного из
противоионов равна нулю!
RT u1 u 2 [Ci ]2
Vдиф
ln
zF u1 u 2 [Ci ]1
6.
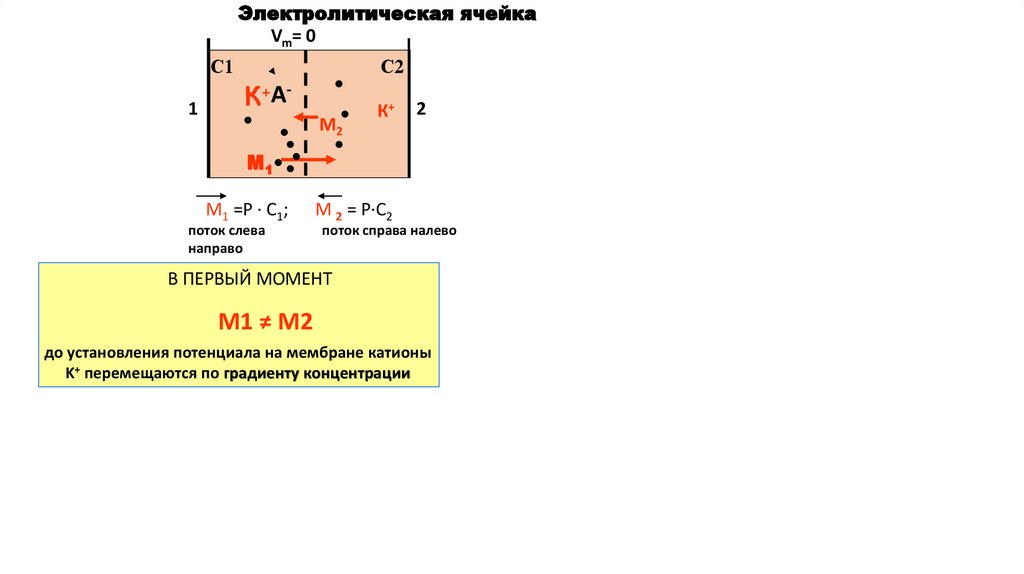
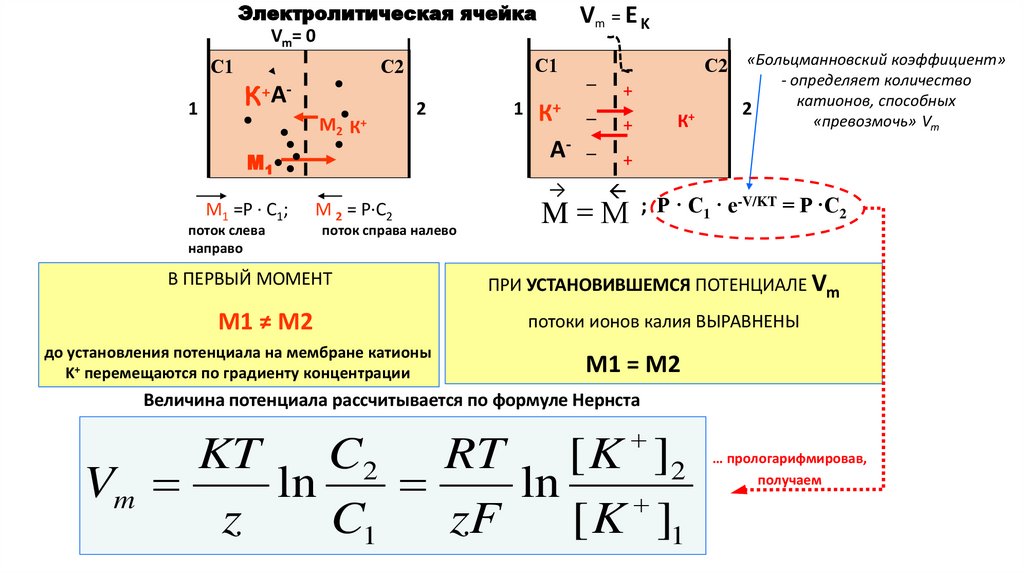
Электролитическая ячейкаVm= 0
С1
1
С2
К+АМ2
К+
2
М1
М1 =Р · С1;
поток слева
направо
М 2 = Р·С2
поток справа налево
В ПЕРВЫЙ МОМЕНТ
М1 ≠ М2
до установления потенциала на мембране катионы
K+ перемещаются по градиенту концентрации
7.
Электролитическая ячейкаVm= 0
С1
1
С1
С2
К+АМ2 К+
2
М1
М1 =Р · С1;
поток слева
направо
Vm = Е K
М 2 = Р·С2
поток справа налево
В ПЕРВЫЙ МОМЕНТ
М1 ≠ М2
до установления потенциала на мембране катионы
K+ перемещаются по градиенту концентрации
1
К+ –
А- –
+
+
+
+
+
+
→
–
С2 «Больцманновский коэффициент»
К+
2
- определяет количество
катионов, способных
«превозмочь» Vm
; Р · С1 · е-V/KT = Р ·C2
M М
ПРИ УСТАНОВИВШЕМСЯ ПОТЕНЦИАЛЕ Vm
потоки ионов калия ВЫРАВНЕНЫ
М1 = М2
Величина потенциала рассчитывается по формуле Нернста
KT C2 RT [ K ]2
Vm
ln
ln
z
C1
zF
[ K ]1
… прологарифмировав,
получаем
8.
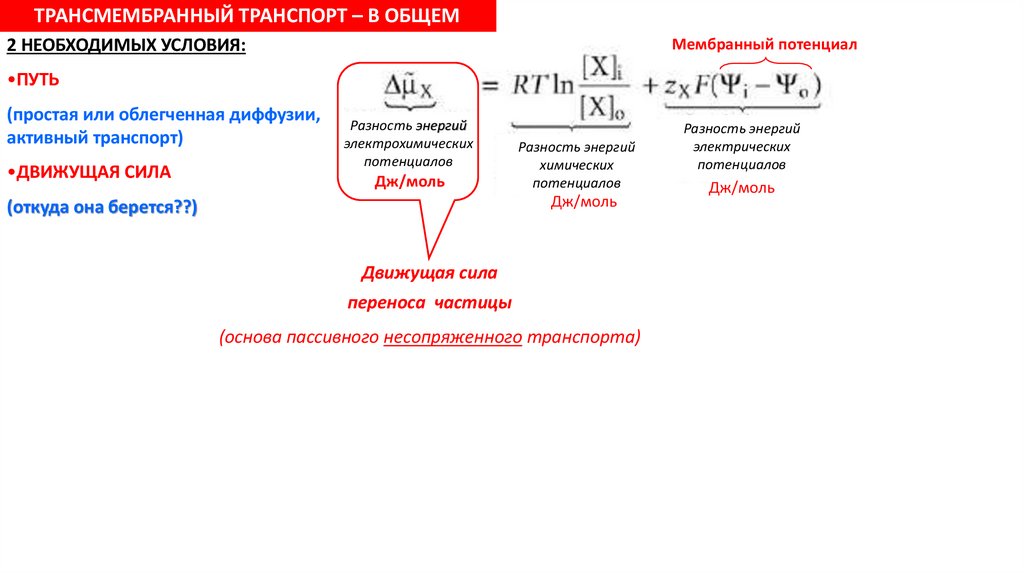
ТРАНСМЕМБРАННЫЙ ТРАНСПОРТ – В ОБЩЕМ2 НЕОБХОДИМЫХ УСЛОВИЯ:
•ПУТЬ
(простая или облегченная диффузии,
активный транспорт)
•ДВИЖУЩАЯ СИЛА
(откуда она берется??)
9.
ТРАНСМЕМБРАННЫЙ ТРАНСПОРТ – В ОБЩЕМ2 НЕОБХОДИМЫХ УСЛОВИЯ:
Мембранный потенциал
•ПУТЬ
(простая или облегченная диффузии,
активный транспорт)
•ДВИЖУЩАЯ СИЛА
Разность энергий
электрохимических
потенциалов
Дж/моль
Разность энергий
химических
потенциалов
Дж/моль
(откуда она берется??)
Движущая сила
переноса частицы
(основа пассивного несопряженного транспорта)
Разность энергий
электрических
потенциалов
Дж/моль
10.
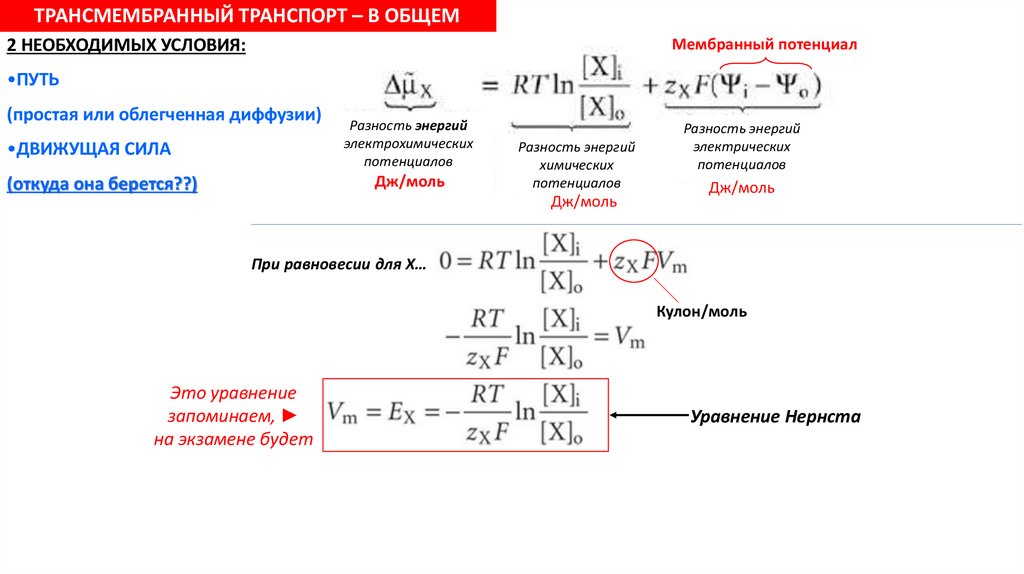
ТРАНСМЕМБРАННЫЙ ТРАНСПОРТ – В ОБЩЕМ2 НЕОБХОДИМЫХ УСЛОВИЯ:
Мембранный потенциал
•ПУТЬ
(простая или облегченная диффузии)
•ДВИЖУЩАЯ СИЛА
Разность энергий
электрохимических
потенциалов
Дж/моль
(откуда она берется??)
Разность энергий
химических
потенциалов
Дж/моль
Разность энергий
электрических
потенциалов
Дж/моль
При равновесии для Х…
Кулон/моль
Это уравнение
запоминаем,
на экзамене будет
Уравнение Нернста
11.
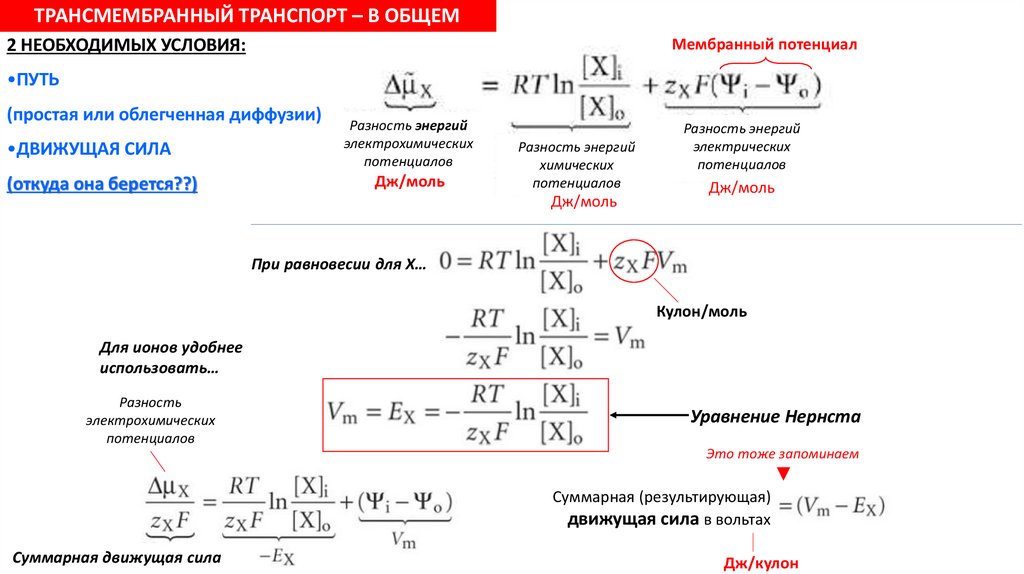
ТРАНСМЕМБРАННЫЙ ТРАНСПОРТ – В ОБЩЕМ2 НЕОБХОДИМЫХ УСЛОВИЯ:
Мембранный потенциал
•ПУТЬ
(простая или облегченная диффузии)
•ДВИЖУЩАЯ СИЛА
(откуда она берется??)
Разность энергий
электрохимических
потенциалов
Дж/моль
Разность энергий
химических
потенциалов
Дж/моль
Разность энергий
электрических
потенциалов
Дж/моль
При равновесии для Х…
Кулон/моль
Для ионов удобнее
использовать…
Разность
электрохимических
потенциалов
Уравнение Нернста
Это тоже запоминаем
▼
Суммарная (результирующая)
движущая сила в вольтах
Суммарная движущая сила
Дж/кулон
12.
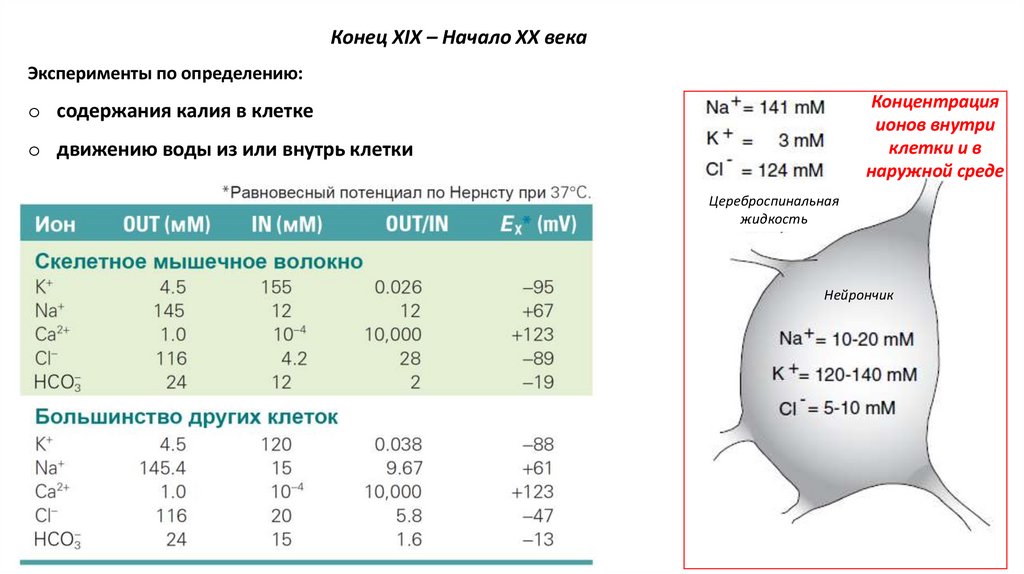
Конец XIX – Начало XX векаЭксперименты по определению:
Концентрация
ионов внутри
клетки и в
наружной среде
o содержания калия в клетке
o движению воды из или внутрь клетки
Цереброспинальная
жидкость
Нейрончик
13.
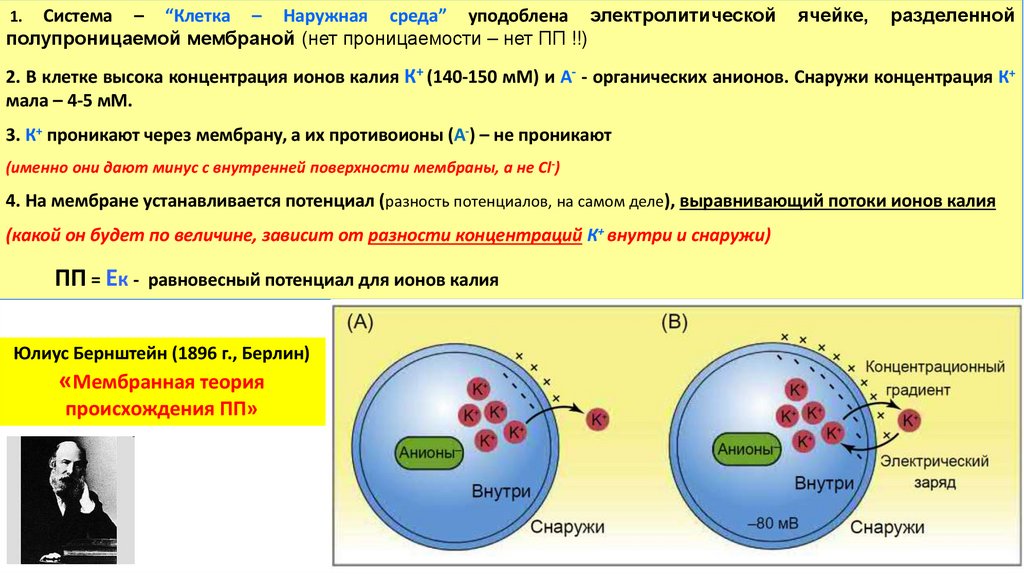
Система – “Клетка – Наружная среда” уподоблена электролитическойполупроницаемой мембраной (нет проницаемости – нет ПП !!)
1.
ячейке,
разделенной
2. В клетке высока концентрация ионов калия К+ (140-150 мМ) и А- - органических анионов. Снаружи концентрация К+
мала – 4-5 мМ.
3. К+ проникают через мембрану, а их противоионы (А-) – не проникают
(именно они дают минус с внутренней поверхности мембраны, а не Cl-)
4. На мембране устанавливается потенциал (разность потенциалов, на самом деле), выравнивающий потоки ионов калия
(какой он будет по величине, зависит от разности концентраций К+ внутри и снаружи)
ПП = Ек - равновесный потенциал для ионов калия
Юлиус Бернштейн (1896 г., Берлин)
«Мембранная теория
происхождения ПП»
14.
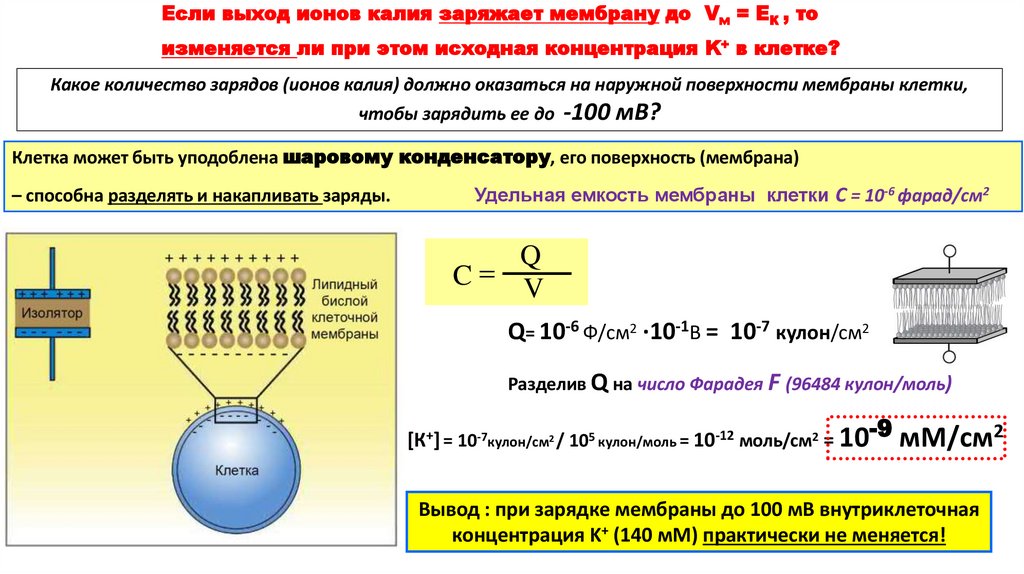
Если выход ионов калия заряжает мембрану до Vм = ЕК , тоизменяется ли при этом исходная концентрация K+ в клетке?
Какое количество зарядов (ионов калия) должно оказаться на наружной поверхности мембраны клетки,
чтобы зарядить ее до -100 мВ?
15.
Если выход ионов калия заряжает мембрану до Vм = ЕК , тоизменяется ли при этом исходная концентрация K+ в клетке?
Какое количество зарядов (ионов калия) должно оказаться на наружной поверхности мембраны клетки,
чтобы зарядить ее до -100 мВ?
Клетка может быть уподоблена шаровому конденсатору, его поверхность (мембрана)
– способна разделять и накапливать заряды.
Удельная емкость мембраны клетки С = 10-6 фарад/см2
Q
C V
Q= 10-6 Ф/cм2 ·10-1В = 10-7 кулон/cм2
Разделив Q на число Фарадея F (96484 кулон/моль)
[К+] = 10-7кулон/см2 / 105 кулон/моль = 10-12 моль/cм2 = 10-9 мМ/см2
Вывод : при зарядке мембраны до 100 мВ внутриклеточная
концентрация K+ (140 мМ) практически не меняется!
16.
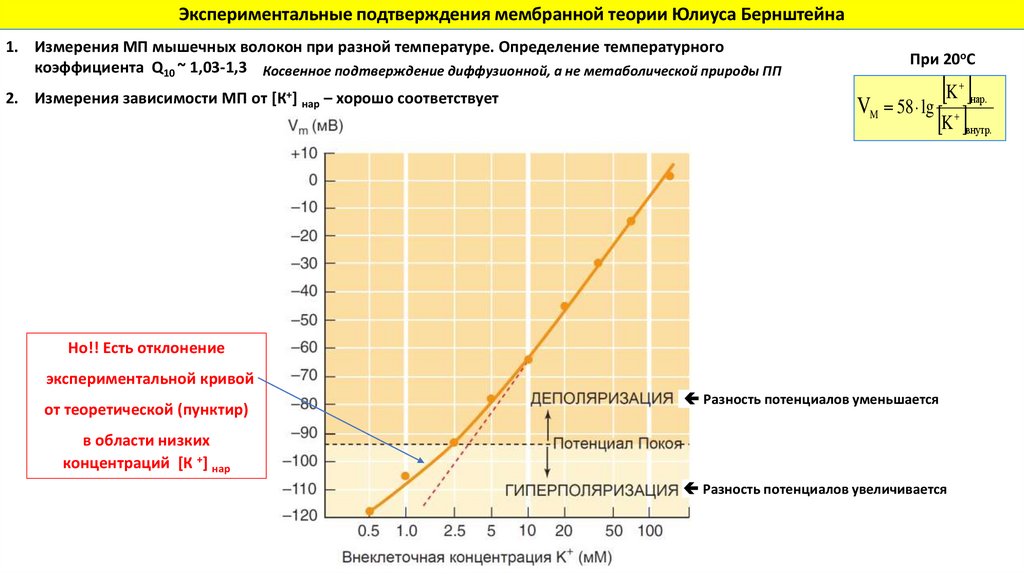
Экспериментальные подтверждения мембранной теории Юлиуса Бернштейна1. Измерения МП мышечных волокон при разной температуре. Определение температурного
коэффициента Q10 ~ 1,03-1,3 Косвенное подтверждение диффузионной, а не метаболической природы ПП
2. Измерения зависимости МП от [К+] нар – хорошо соответствует
При 20оС
K
V 58 lg
K
нар.
M
внутр.
Но!! Есть отклонение
экспериментальной кривой
от теоретической (пунктир)
Разность потенциалов уменьшается
в области низких
концентраций [К +] нар
Разность потенциалов увеличивается
17.
40-е годы XX векаМодель мембраны, проницаемой к трем ионам – K+, Cl- и Na+
ДАНО:
ПП = - 80 мВ; ЕNa ≈ +30 мВ, а мембрана клетки пропускает Na+
Если мембрана клетки проницаема и к Na+, то МП не может быть равновесным для трансмембранных
потоков ионов натрия и других проникающих ионов – калия и хлора
ПП ≠ Еравн К ≠ Еравн Cl ≠ Еравн Na
Вопрос: как же
формируется и
удерживается
стабильный ПП
порядка –80мВ?
18.
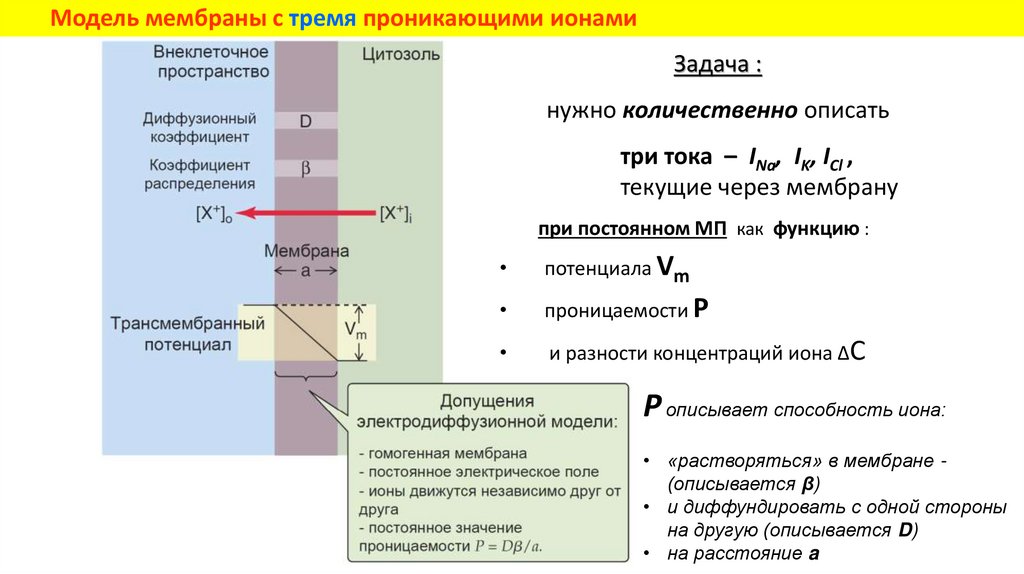
Модель мембраны с тремя проникающими ионамиЗадача :
нужно количественно описать
три тока – INa, IK, ICl ,
текущие через мембрану
при постоянном МП как функцию :
потенциала Vm
проницаемости P
и разности концентраций иона ΔC
P описывает способность иона:
• «растворяться» в мембране (описывается β)
• и диффундировать с одной стороны
на другую (описывается D)
• на расстояние a
19.
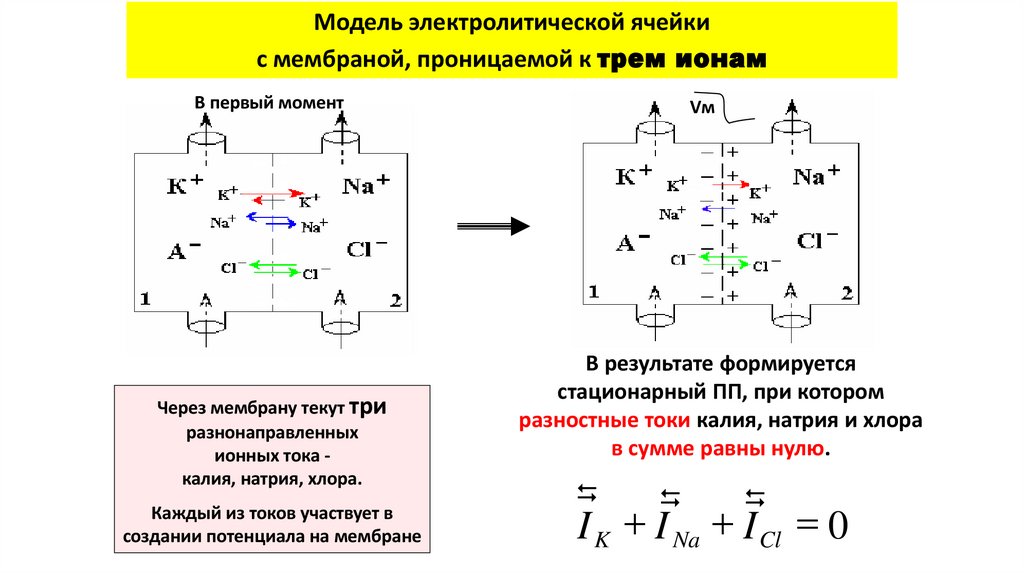
Модель электролитической ячейкис мембраной, проницаемой к трем ионам
В первый момент
Через мембрану текут три
разнонаправленных
ионных тока калия, натрия, хлора.
Каждый из токов участвует в
создании потенциала на мембране
Vм
В результате формируется
стационарный ПП, при котором
разностные токи калия, натрия и хлора
в сумме равны нулю.
I K I Na I Cl 0
20.
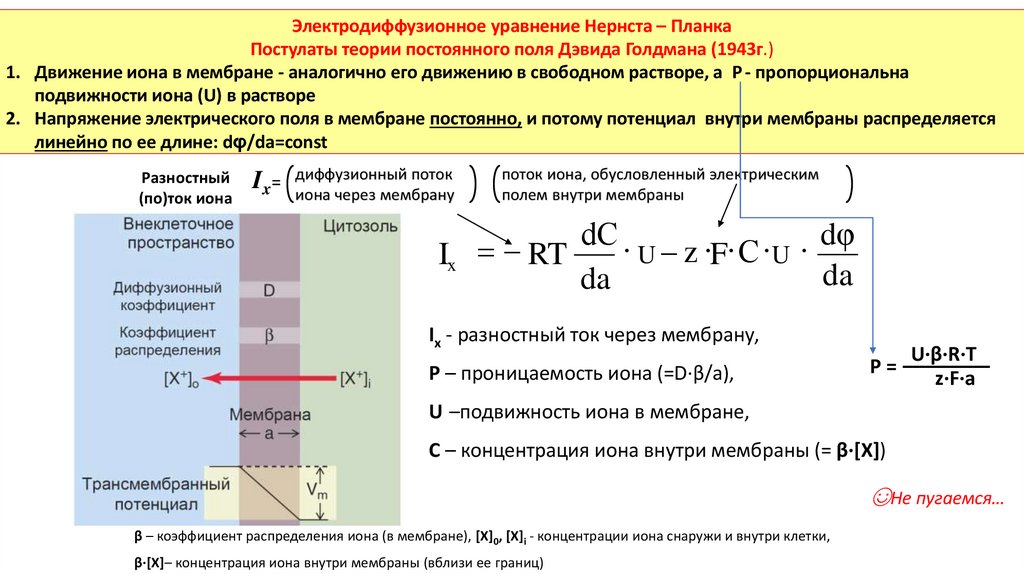
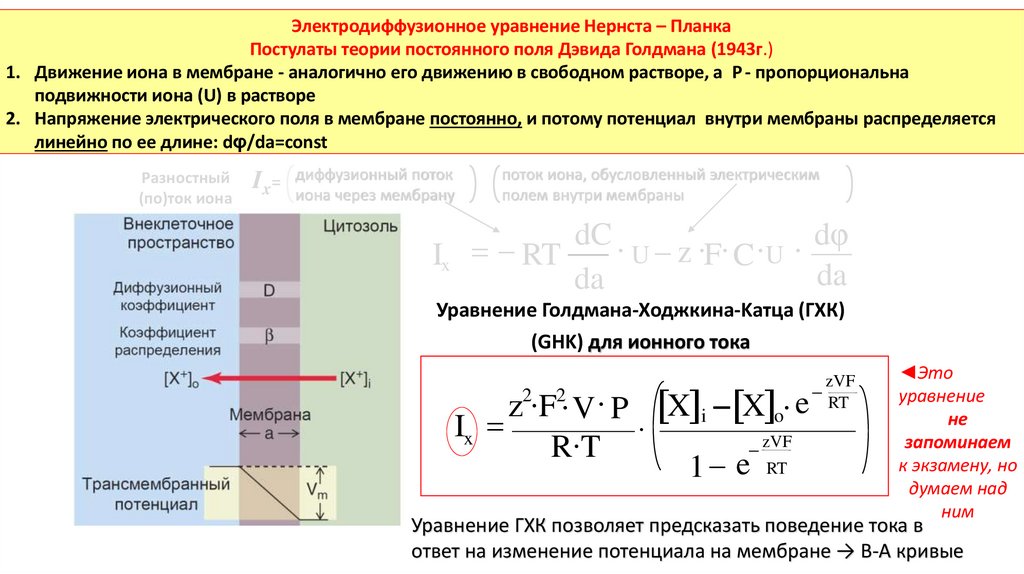
Электродиффузионное уравнение Нернста – ПланкаПостулаты теории постоянного поля Дэвида Голдмана (1943г.)
1. Движение иона в мембране - аналогично его движению в свободном растворе, а Р - пропорциональна
подвижности иона (U) в растворе
2. Напряжение электрического поля в мембране постоянно, и потому потенциал внутри мембраны распределяется
линейно по ее длине: dφ/da=сonst
Разностный
(по)ток иона
поток
Ix= диффузионный
иона через мембрану
поток иона, обусловленный электрическим
полем внутри мембраны
dС
dφ
∙ U z ∙F∙ С ∙U ∙
Ix RT
da
da
Ix - разностный ток через мембрану,
P – проницаемость иона (=D·β/a),
P=
U·β·R·T
z·F·a
U –подвижность иона в мембране,
С – концентрация иона внутри мембраны (= β·[X])
☺Не пугаемся…
β – коэффициент распределения иона (в мембране), [X]0, [X]i - концентрации иона снаружи и внутри клетки,
β·[X]– концентрация иона внутри мембраны (вблизи ее границ)
21.
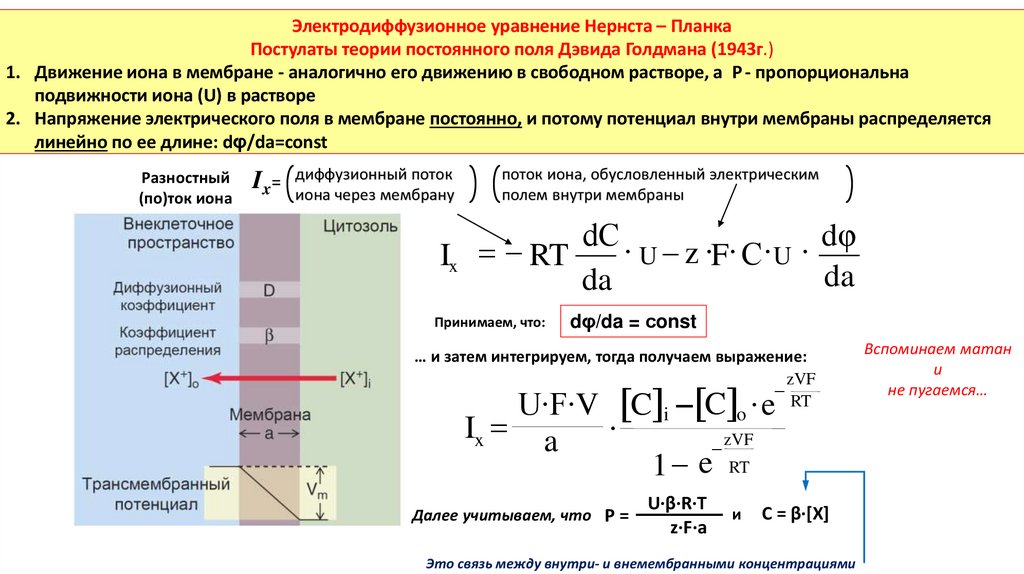
Электродиффузионное уравнение Нернста – ПланкаПостулаты теории постоянного поля Дэвида Голдмана (1943г.)
1. Движение иона в мембране - аналогично его движению в свободном растворе, а Р - пропорциональна
подвижности иона (U) в растворе
2. Напряжение электрического поля в мембране постоянно, и потому потенциал внутри мембраны распределяется
линейно по ее длине: dφ/da=сonst
Разностный
(по)ток иона
поток
Ix= диффузионный
иона через мембрану
поток иона, обусловленный электрическим
полем внутри мембраны
dС
dφ
∙ U z ∙F∙ С ∙U ∙
Ix RT
da
da
Принимаем, что:
dφ/da = сonst
… и затем интегрируем, тогда получаем выражение:
zVF
RT
U∙F∙V C i C o ∙ e
Ix a
∙
zVF
1 e RT
Далее учитываем, что P =
U·β·R·T
z·F·a
и
C = β·[X]
Это связь между внутри- и внемембранными концентрациями
Вспоминаем матан
и
не пугаемся…
22.
Электродиффузионное уравнение Нернста – ПланкаПостулаты теории постоянного поля Дэвида Голдмана (1943г.)
1. Движение иона в мембране - аналогично его движению в свободном растворе, а Р - пропорциональна
подвижности иона (U) в растворе
2. Напряжение электрического поля в мембране постоянно, и потому потенциал внутри мембраны распределяется
линейно по ее длине: dφ/da=сonst
Разностный
(по)ток иона
поток
Ix= диффузионный
иона через мембрану
поток иона, обусловленный электрическим
полем внутри мембраны
dC
dφ
∙ U z ∙F∙ C ∙U ∙
Ix RT
da
da
Уравнение Голдмана-Ходжкина-Kатца (ГХК)
(GHK) для ионного тока
z2∙F2∙ V ∙ P X i X o∙ e
Ix
∙
zVF
R∙T
1 e RT
zVF
RT
◄Это
уравнение
не
запоминаем
к экзамену, но
думаем над
ним
Уравнение ГХК позволяет предсказать поведение тока в
ответ на изменение потенциала на мембране → В-А кривые
23.
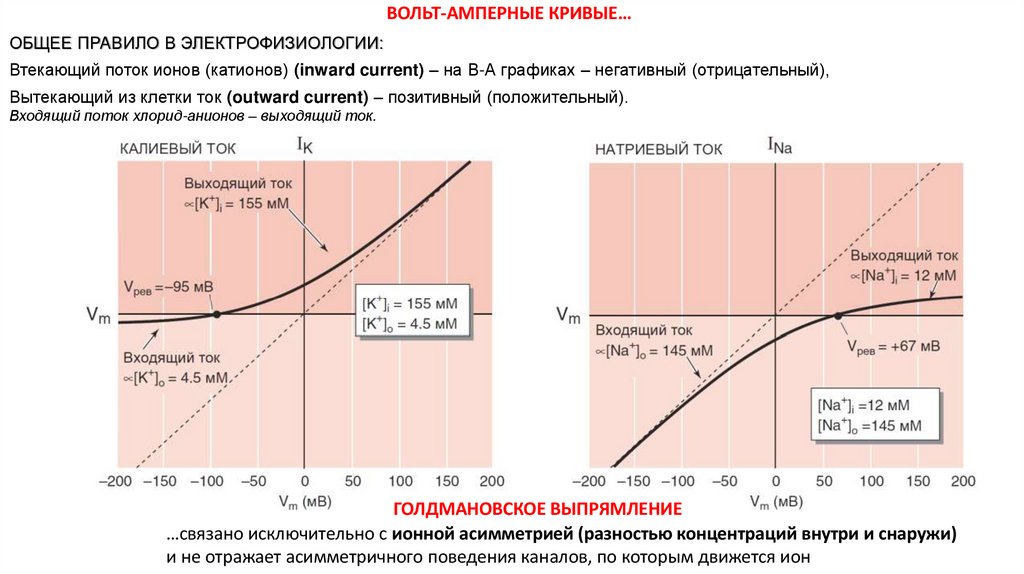
ВОЛЬТ-АМПЕРНЫЕ КРИВЫЕ…ОБЩЕЕ ПРАВИЛО В ЭЛЕКТРОФИЗИОЛОГИИ:
Втекающий поток ионов (катионов) (inward current) – на В-А графиках – негативный (отрицательный),
Вытекающий из клетки ток (outward current) – позитивный (положительный).
Входящий поток хлорид-анионов – выходящий ток.
ГОЛДМАНОВСКОЕ ВЫПРЯМЛЕНИЕ
…связано исключительно с ионной асимметрией (разностью концентраций внутри и снаружи)
и не отражает асимметричного поведения каналов, по которым движется ион
24.
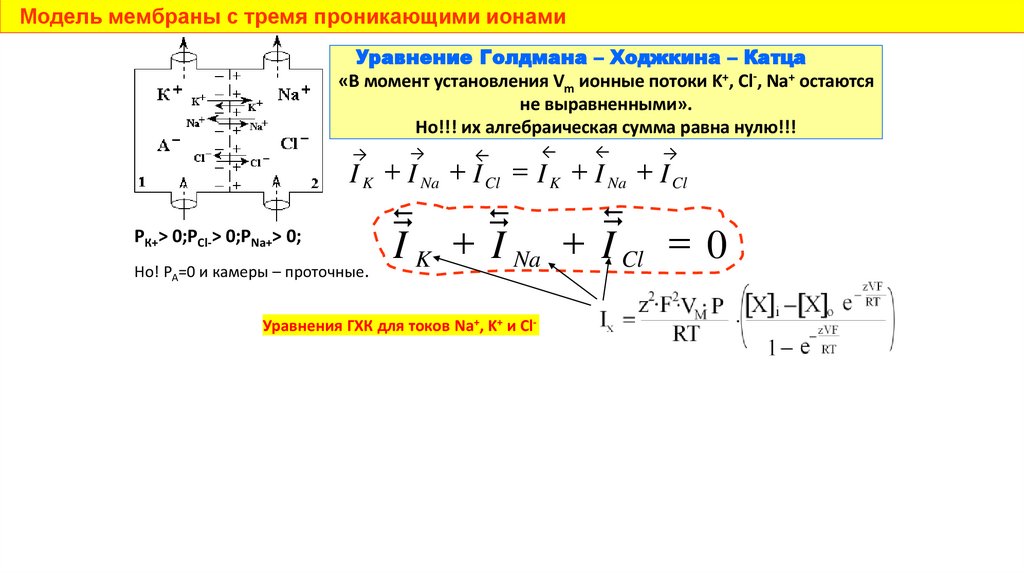
Модель мембраны с тремя проникающими ионамиУравнение Голдмана – Ходжкина – Катца
«В момент установления Vm ионные потоки K+, Cl-, Na+ остаются
не выравненными».
Но!!! их алгебраическая сумма равна нулю!!!
→
→
→
→
→
→
I K I Na I Cl I K I Na I Cl
I Na I Cl 0
I
K
Но! Р =0 и камеры – проточные.
РК+> 0;РCl-> 0;РNa+> 0;
А
25.
Модель мембраны с тремя проникающими ионамиУравнение Голдмана – Ходжкина – Катца
«В момент установления Vm ионные потоки K+, Cl-, Na+ остаются
не выравненными».
Но!!! их алгебраическая сумма равна нулю!!!
→
→
→
→
→
→
I K I Na I Cl I K I Na I Cl
I Na I Cl 0
I
K
Но! Р =0 и камеры – проточные.
РК+> 0;РCl-> 0;РNa+> 0;
А
Уравнения ГХК для токов Na+, K+ и Сl-
26.
Модель мембраны с тремя проникающими ионамиУравнение Голдмана – Ходжкина – Катца
«В момент установления VМ ионные потоки K+, Cl-, Na+ остаются
не выровненными».
Но!!! их алгебраическая сумма равна нулю!!!
→
→
→
→
→
→
I K I Na I Cl I K I Na I Cl
I Na I Cl 0
I
K
Но! Р =0 и камеры – проточные.
РК+> 0;РCl-> 0;РNa+> 0;
А
Уравнения ГХК для токов Na+, K+ и Сl-
PK K 1 e
e
V
RT
V
RT
V
RT
V
RT
P Cl e P K P Na P Cl
( P K P Na P Cl ) P K P Na P Cl
PNa Na 1 e
K
Решаем для Vm,
и, прологарифмировав,
получаем
Na
2
Cl
1
1
Cl
2
2
Na
2
K
2
K
Na
Cl
1
21
Cl
1
PK K 2 PNa Na 2 PCl Cl 1
RT
VМ
ln
F
PK K 1 PNa Na 1 PCl Cl 2
27.
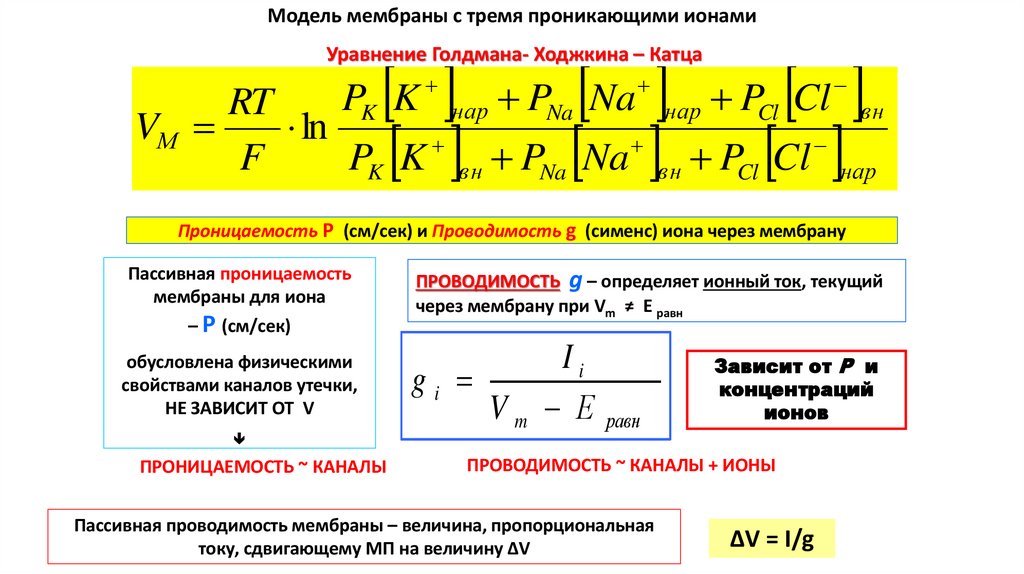
МОДЕЛЬ МЕМБРАНЫ С ТРЕМЯ ПРОНИКАЮЩИМИ ИОНАМИУравнение Голдмана-Ходжкина-Катца (ГХК)
Это уравнение
запоминаем
PK K нар PNa Na нар PCl Cl вн
RT
VМ
ln
F
PK K вн PNa Na вн PCl Cl нар
В числителе у анионов Cl- введена внутриклеточная концентрация, а не наружная, как у ионов К+ и Na+.
Это связано с отрицательным зарядом анионов Cl- ► их ток имеет противоположное направление.
Важно, как соотносятся величины проницаемостей
РK : PNa : PCl между собой при ПП
Чем больше будет проницаемость мембраны для конкретного иона,
тем легче он будет мембрану преодолевать,
и тем больший заряд он будет переносить, участвуя в формировании ПП.
28.
Модель мембраны с тремя проникающими ионамиУравнение Голдмана- Ходжкина – Катца
PK K нар PNa Na нар PCl Cl вн
RT
VМ
ln
F
PK K вн PNa Na вн PCl Cl нар
Проницаемость Р (см/сек) и Проводимость g (сименс) иона через мембрану
Пассивная проницаемость
мембраны для иона
– Р (см/сек)
обусловлена физическими
свойствами каналов утечки,
НЕ ЗАВИСИТ ОТ V
ПРОВОДИМОСТЬ g – определяет ионный ток, текущий
через мембрану при Vm ≠ Е равн
Ii
gi
V m Е равн
Зависит от Р и
концентраций
ионов
ПРОНИЦАЕМОСТЬ ~ КАНАЛЫ
ПРОВОДИМОСТЬ ~ КАНАЛЫ + ИОНЫ
Пассивная проводимость мембраны – величина, пропорциональная
току, сдвигающему МП на величину ∆V
ΔV = I/g
29.
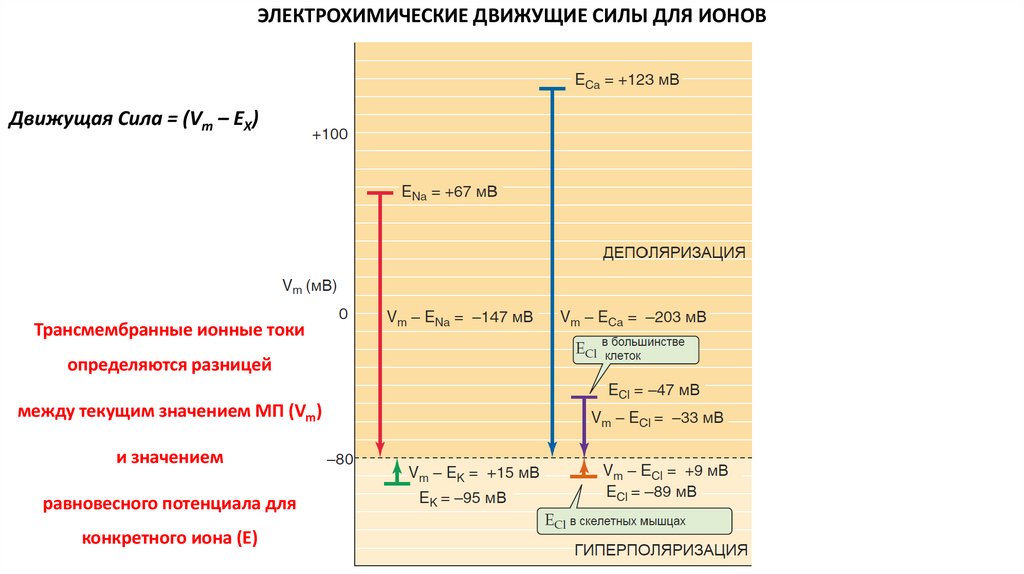
ЭЛЕКТРОХИМИЧЕСКИЕ ДВИЖУЩИЕ СИЛЫ ДЛЯ ИОНОВДвижущая Сила = (Vm – EX)
Трансмембранные ионные токи
определяются разницей
между текущим значением МП (Vm)
и значением
равновесного потенциала для
конкретного иона (E)
30.
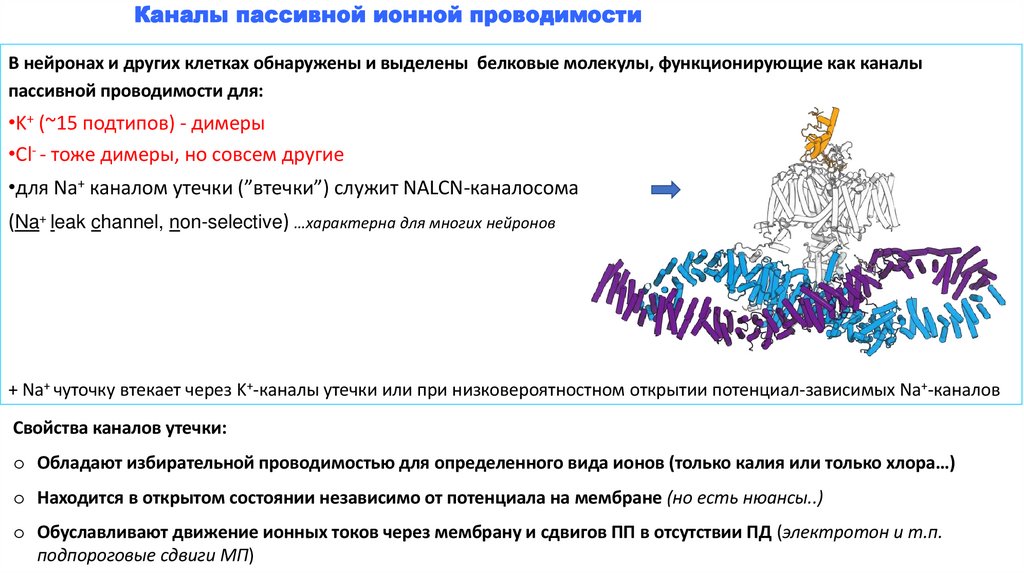
Каналы пассивной ионной проводимостиВ нейронах и других клетках обнаружены и выделены белковые молекулы, функционирующие как каналы
пассивной проводимости для:
•K+ (~15 подтипов) - димеры
•Cl- - тоже димеры, но совсем другие
•для Na+ каналом утечки (”втечки”) служит NALCN-каналосома
(Na+ leak channel, non-selective) …характерна для многих нейронов
+ Na+ чуточку втекает через K+-каналы утечки или при низковероятностном открытии потенциал-зависимых Na+-каналов
Свойства каналов утечки:
o Обладают избирательной проводимостью для определенного вида ионов (только калия или только хлора…)
o Находится в открытом состоянии независимо от потенциала на мембране (но есть нюансы..)
o Обуславливают движение ионных токов через мембрану и сдвигов ПП в отсутствии ПД (электротон и т.п.
подпороговые сдвиги МП)
31.
ИОННЫЕКАНАЛЫ
Функция: формирование в мембране гидрофильной поры
для прохождения ионов по электрохимическому градиенту
По сравнению с переносчиками:
•Меньшая специфичность
•Трансмембранный транспорт во много раз «мощнее»
•Практически нет насыщения
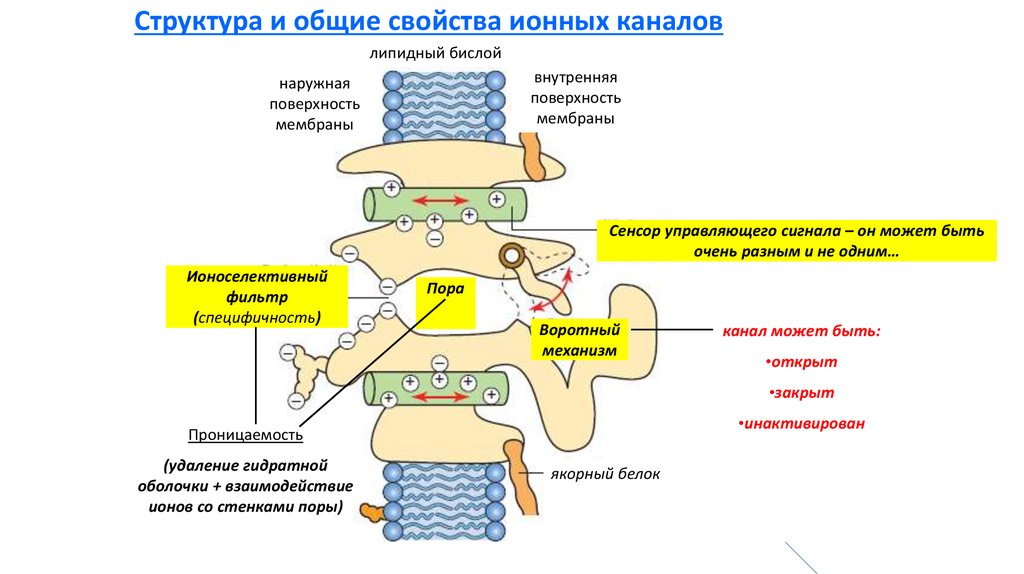
Структура и общие свойства ионных каналов
Несколько связанных между собой
белковых субъединиц
+ дополнительные регуляторные
субъединицы
Важны:
•Первичная структура (последовательность аминокислот в
каналообразующей субъединице)
◄ Это в
качестве
примера…
•Изменения конформации в разных частях интегрального белкового
комплекса
Разнообразные свойства различных
ионных каналов
(селективность, проводимость и др.)
32.
Структура и общие свойства ионных каналовлипидный бислой
внутренняя
поверхность
мембраны
наружная
поверхность
мембраны
Сенсор управляющего сигнала – он может быть
очень разным и не одним…
Ионоселективный
фильтр
(специфичность)
Пора
Воротный
механизм
канал может быть:
•открыт
•закрыт
•инактивирован
Проницаемость
(удаление гидратной
оболочки + взаимодействие
ионов со стенками поры)
якорный белок
33.
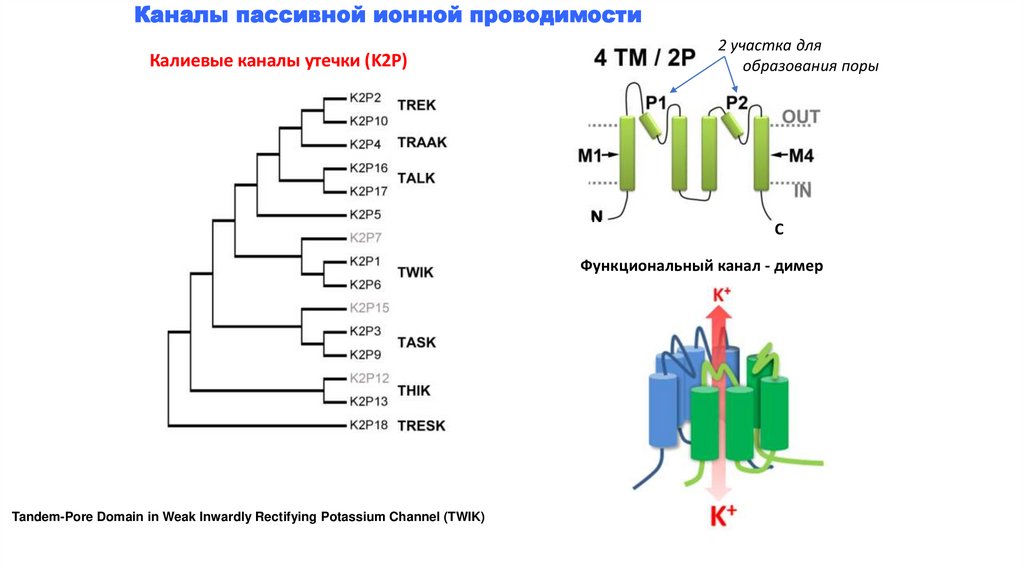
Каналы пассивной ионной проводимости2 участка для
образования поры
Калиевые каналы утечки (K2P)
N
C
Функциональный канал - димер
Tandem-Pore Domain in Weak Inwardly Rectifying Potassium Channel (TWIK)
34.
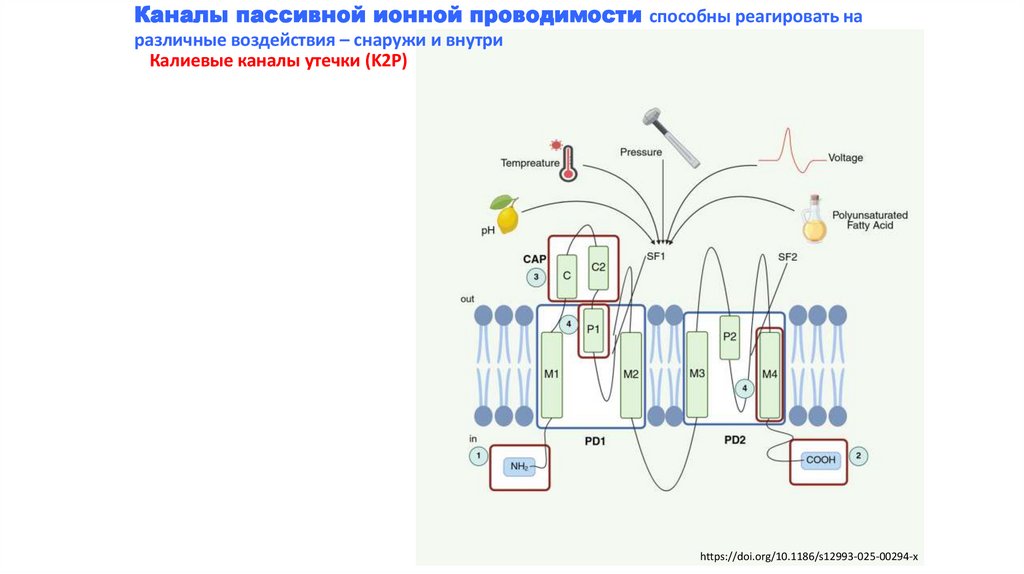
Каналы пассивной ионной проводимости способны реагировать наразличные воздействия – снаружи и внутри
Калиевые каналы утечки (K2P)
https://doi.org/10.1186/s12993-025-00294-x
35.
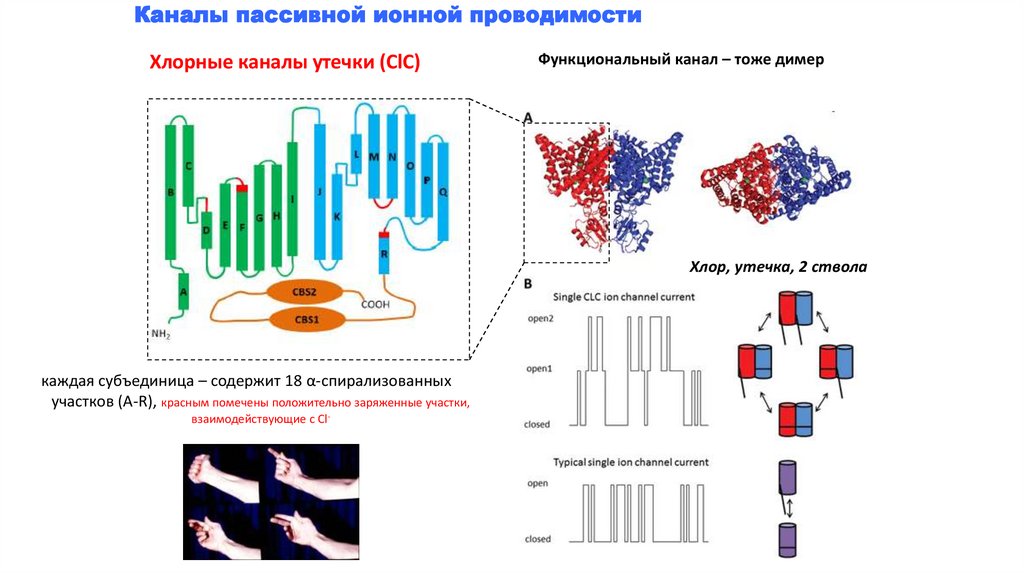
Каналы пассивной ионной проводимостиХлорные каналы утечки (СlC)
Функциональный канал – тоже димер
Хлор, утечка, 2 ствола
каждая субъединица – содержит 18 α-спирализованных
участков (А-R), красным помечены положительно заряженные участки,
взаимодействующие с Cl-
36.
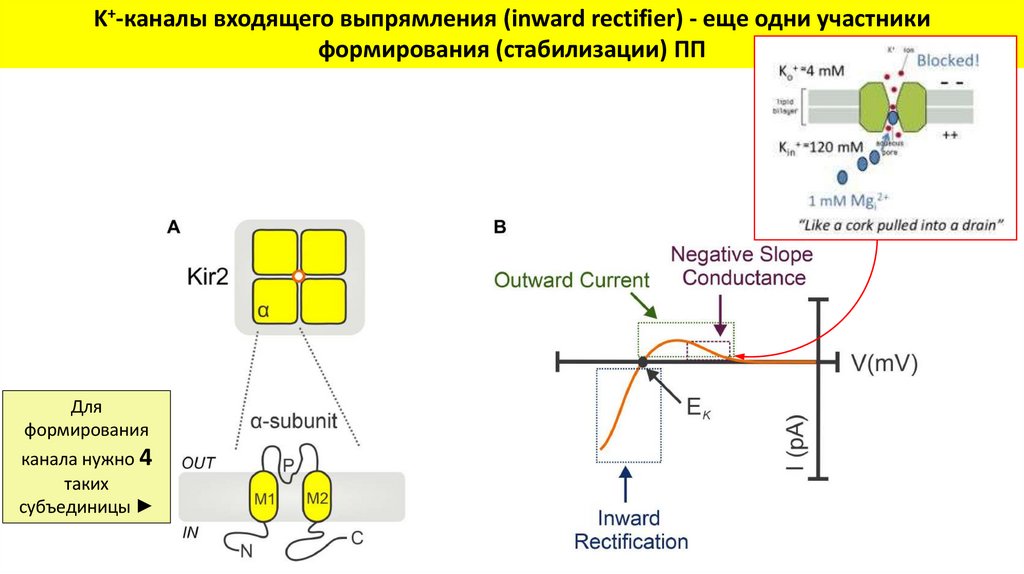
K+-каналы входящего выпрямления (inward rectifier) - еще одни участникиформирования (стабилизации) ПП
Для
формирования
канала нужно 4
таких
субъединицы
37.
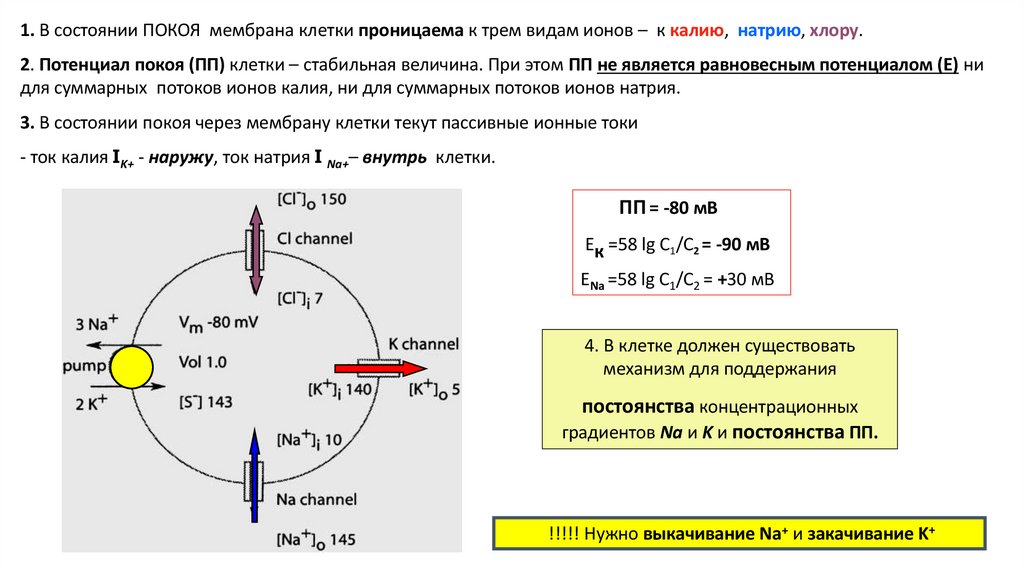
1. В состоянии ПОКОЯ мембрана клетки проницаема к трем видам ионов – к калию, натрию, хлору.2. Потенциал покоя (ПП) клетки – стабильная величина. При этом ПП не является равновесным потенциалом (Е) ни
для суммарных потоков ионов калия, ни для суммарных потоков ионов натрия.
3. В состоянии покоя через мембрану клетки текут пассивные ионные токи
- ток калия IK+ - наружу, ток натрия I Na+– внутрь клетки.
ПП = -80 мВ
Ек =58 lg C1/C2 = -90 мВ
ЕNa =58 lg C1/C2 = +30 мВ
4. В клетке должен существовать
механизм для поддержания
постоянства концентрационных
градиентов Na и K и постоянства ПП.
!!!!! Нужно выкачивание Na+ и закачивание K+
38.
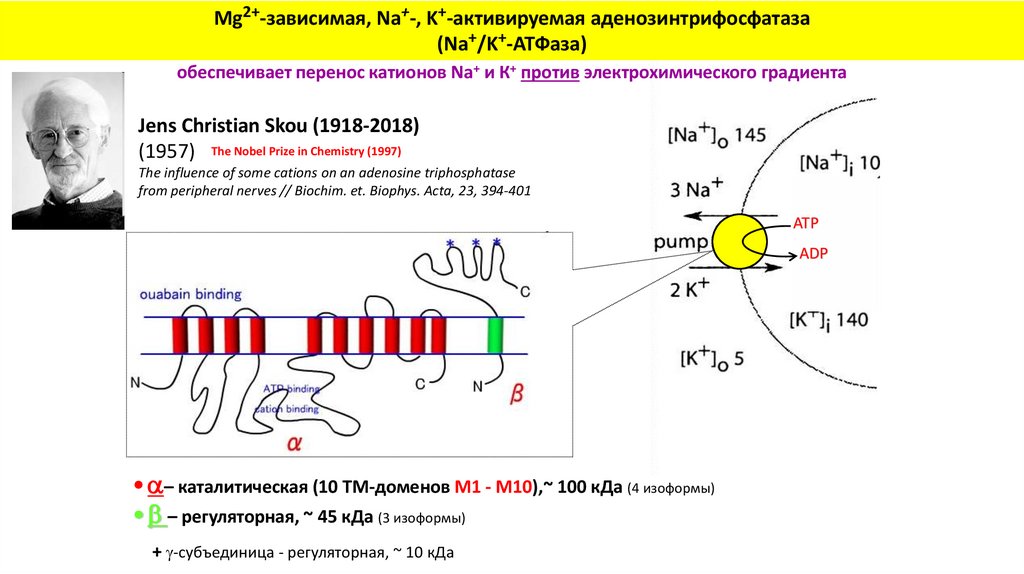
Mg2+-зависимая, Na+-, K+-активируемая аденозинтрифосфатаза(Na+/K+-АТФаза)
обеспечивает перенос катионов Na+ и К+ против электрохимического градиента
Jens Christian Skou (1918-2018)
(1957) The Nobel Prize in Chemistry (1997)
The influence of some cations on an adenosine triphosphatase
from peripheral nerves // Biochim. et. Biophys. Acta, 23, 394-401
ATP
ADP
• – каталитическая (10 ТМ-доменов M1 - M10),~ 100 кДа (4 изоформы)
• – регуляторная, ~ 45 кДа (3 изоформы)
+ -субъединица - регуляторная, ~ 10 кДа
39.
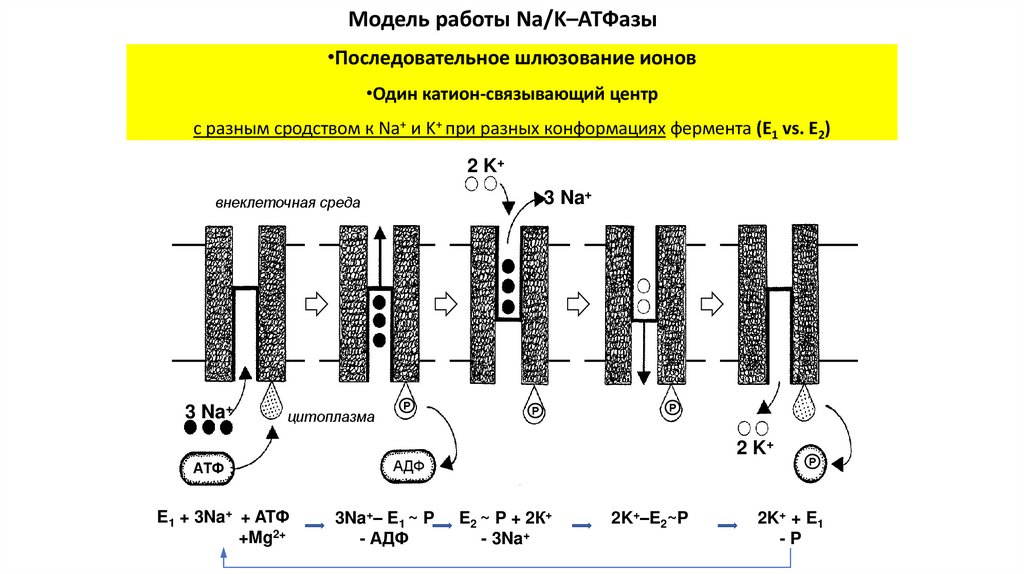
Модель работы Na/K–ATФазы•Последовательное шлюзование ионов
•Один катион-связывающий центр
с разным сродством к Na+ и K+ при разных конформациях фермента (E1 vs. E2)
2 K+
3 Na+
внеклеточная среда
3 Na+
цитоплазма
АТФ
E1 + 3Na+ + ATФ
+Mg2+
℗
℗
℗
2 K+
АДФ
3Na+– E1 ~ P
- АДФ
E2 ~ P + 2К+
- 3Na+
2K+–E2~P
℗
2K+ + E1
-P
40.
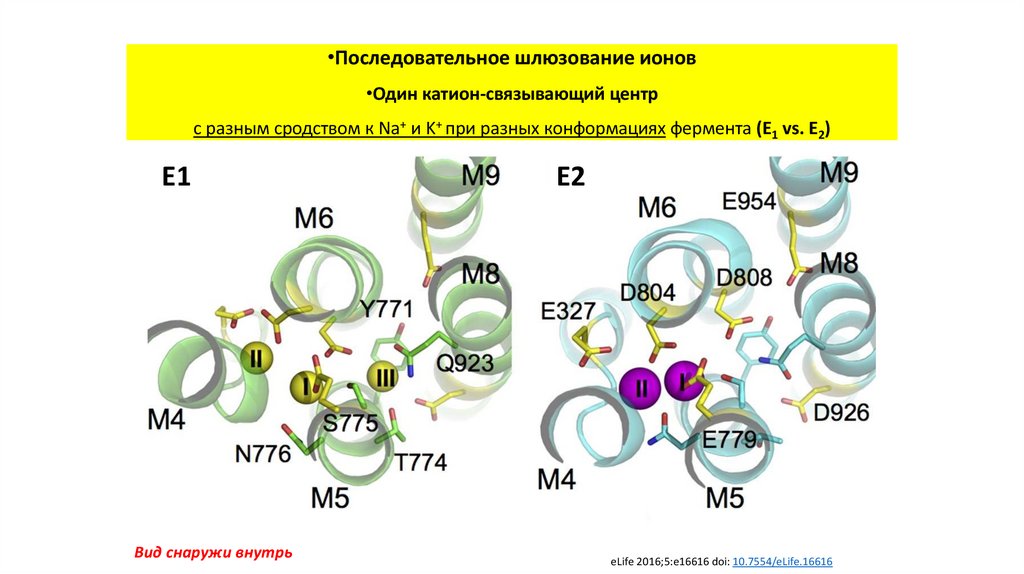
•Последовательное шлюзование ионов•Один катион-связывающий центр
с разным сродством к Na+ и K+ при разных конформациях фермента (E1 vs. E2)
E1
Вид снаружи внутрь
E2
eLife 2016;5:e16616 doi: 10.7554/eLife.16616
41.
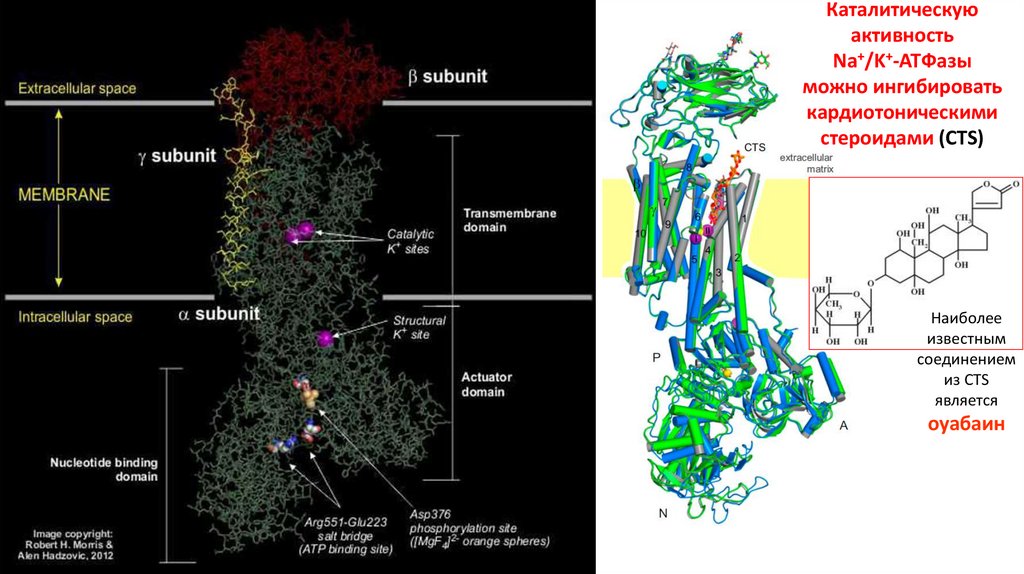
Каталитическуюактивность
Na+/K+-ATФазы
можно ингибировать
кардиотоническими
стероидами (CTS)
Наиболее
известным
соединением
из CTS
является
оуабаин
42.
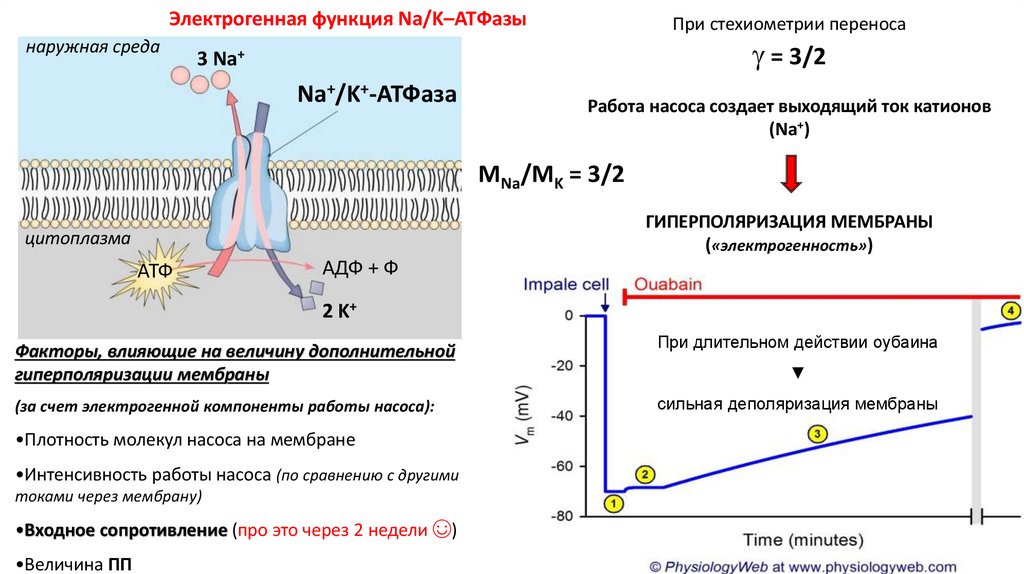
Электрогенная функция Na/K–ATФазынаружная среда
При стехиометрии переноса
= 3/2
3 Na+
Na+/K+-АТФаза
Работа насоса создает выходящий ток катионов
(Na+)
МNa/МK = 3/2
ГИПЕРПОЛЯРИЗАЦИЯ МЕМБРАНЫ
(«электрогенность»)
цитоплазма
АТФ
АДФ + Ф
2 K+
Факторы, влияющие на величину дополнительной
гиперполяризации мембраны
(за счет электрогенной компоненты работы насоса):
•Плотность молекул насоса на мембране
•Интенсивность работы насоса (по сравнению с другими
токами через мембрану)
•Входное сопротивление (про это через 2 недели ☺)
•Величина ПП
При длительном действии оубаина
▼
сильная деполяризация мембраны
43.
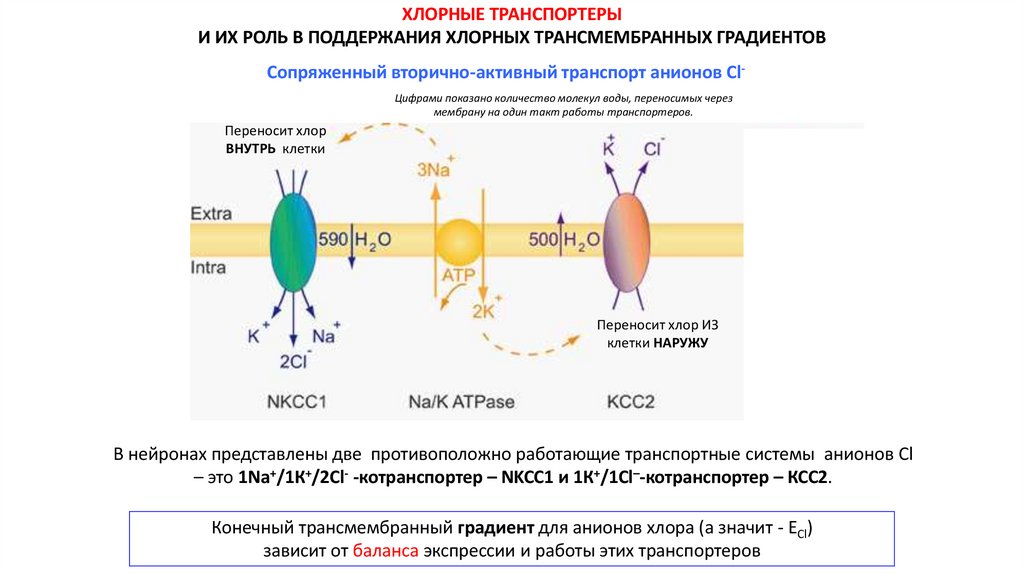
ХЛОРНЫЕ ТРАНСПОРТЕРЫИ ИХ РОЛЬ В ПОДДЕРЖАНИЯ ХЛОРНЫХ ТРАНСМЕМБРАННЫХ ГРАДИЕНТОВ
Сопряженный вторично-активный транспорт анионов ClЦифрами показано количество молекул воды, переносимых через
мембрану на один такт работы транспортеров.
Переносит хлор
ВНУТРЬ клетки
Переносит хлор ИЗ
клетки НАРУЖУ
В нейронах представлены две противоположно работающие транспортные системы анионов Cl
– это 1Na+/1К+/2Cl- -котранспортер – NKCC1 и 1К+/1Cl–-котранспортер – КСС2.
Конечный трансмембранный градиент для анионов хлора (а значит - ECl)
зависит от баланса экспрессии и работы этих транспортеров
44.
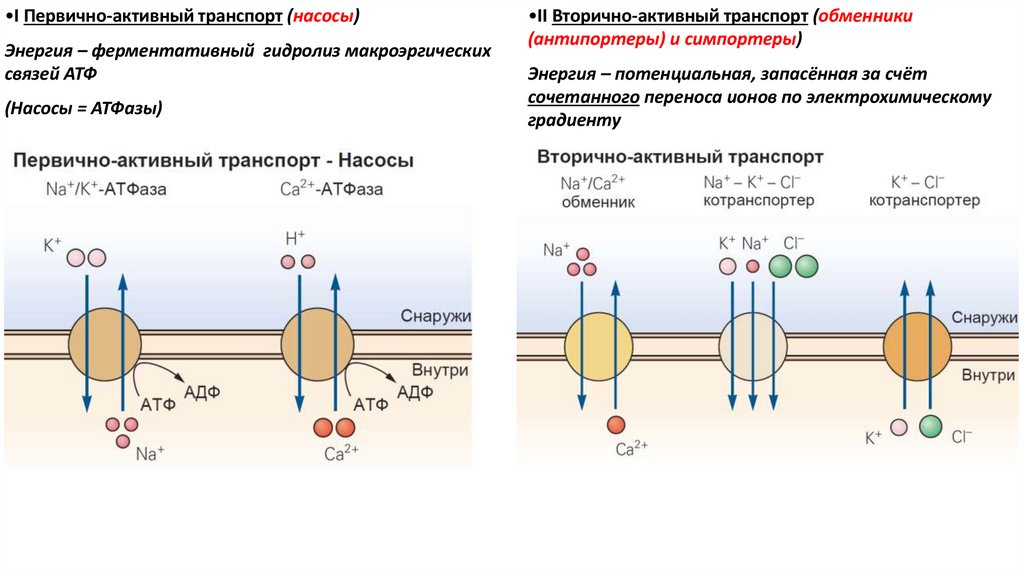
•I Первично-активный транспорт (насосы)Энергия – ферментативный гидролиз макроэргических
связей АТФ
(Насосы = АТФазы)
•II Вторично-активный транспорт (обменники
(антипортеры) и симпортеры)
Энергия – потенциальная, запасённая за счёт
сочетанного переноса ионов по электрохимическому
градиенту
45.
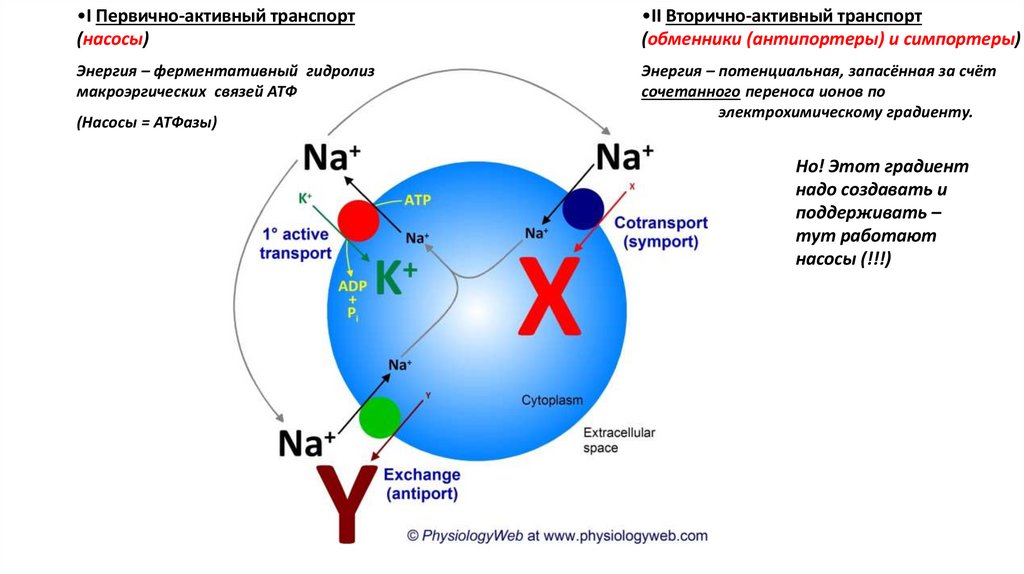
•I Первично-активный транспорт(насосы)
•II Вторично-активный транспорт
(обменники (антипортеры) и симпортеры)
Энергия – ферментативный гидролиз
макроэргических связей АТФ
Энергия – потенциальная, запасённая за счёт
сочетанного переноса ионов по
электрохимическому градиенту.
(Насосы = АТФазы)
Но! Этот градиент
надо создавать и
поддерживать –
тут работают
насосы (!!!)
46.
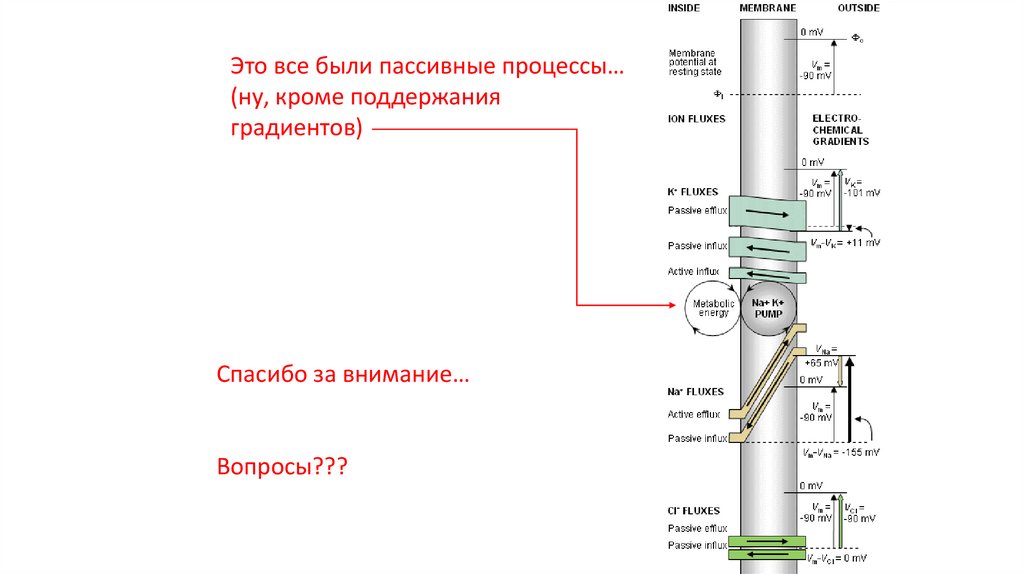
Это все были пассивные процессы…(ну, кроме поддержания
градиентов)
Спасибо за внимание…
Вопросы???