





 biology
biologySimilar presentations:

")


")





Анатомия и физиология человека. Потенциал действия. Лекция 2
1.
АНАТОМИЯ И ФИЗИОЛОГИЯ ЧЕЛОВЕКАЛЕКЦИЯ 2
•Потенциал действия
Гайдуков Александр Евгеньевич
МФТИ 2025
2.
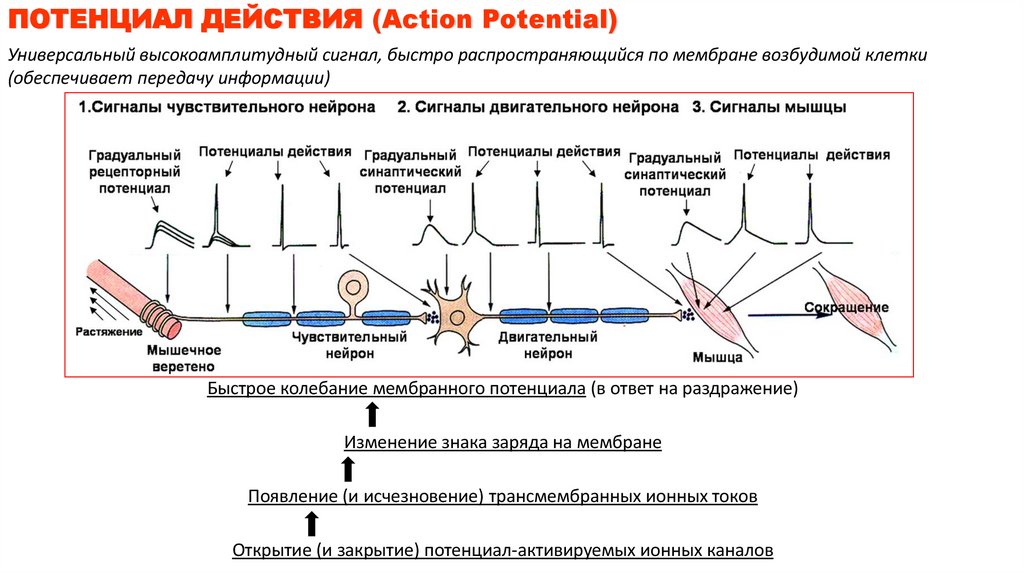
ПОТЕНЦИАЛ ДЕЙСТВИЯ (Action Potential)Универсальный высокоамплитудный сигнал, быстро распространяющийся по мембране возбудимой клетки
(обеспечивает передачу информации)
Быстрое колебание мембранного потенциала (в ответ на раздражение)
Изменение знака заряда на мембране
Появление (и исчезновение) трансмембранных ионных токов
Открытие (и закрытие) потенциал-активируемых ионных каналов
3.
МП(мВ)
0
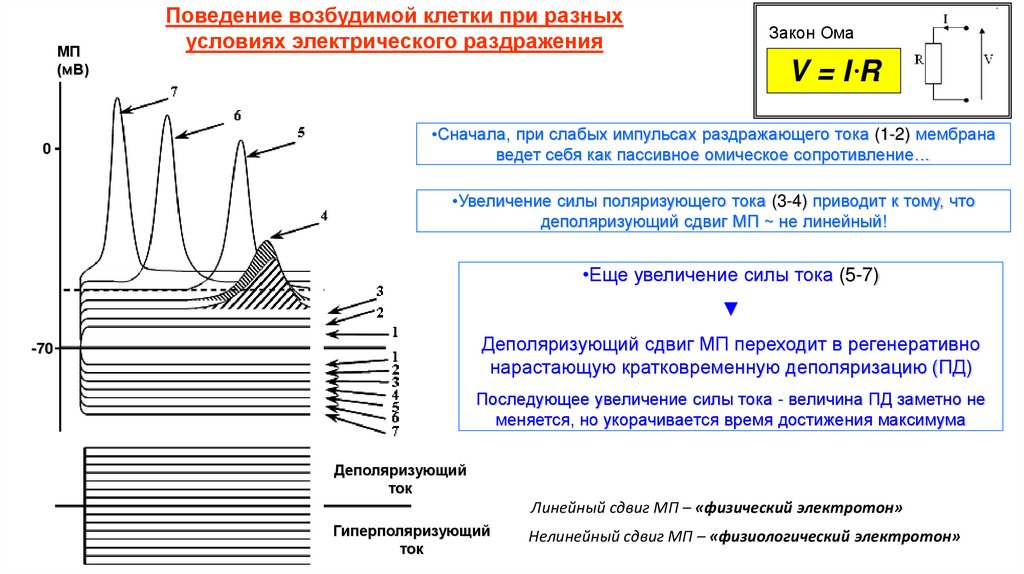
Поведение возбудимой клетки при разных
условиях электрического раздражения
Закон Ома
V=IR
•Сначала, при слабых импульсах раздражающего тока (1-2) мембрана
ведет себя как пассивное омическое сопротивление…
•Увеличение силы поляризующего тока (3-4) приводит к тому, что
деполяризующий сдвиг МП ~ не линейный!
•Еще увеличение силы тока (5-7)
▼
Деполяризующий сдвиг МП переходит в регенеративно
нарастающую кратковременную деполяризацию (ПД)
-70
Последующее увеличение силы тока - величина ПД заметно не
меняется, но укорачивается время достижения максимума
Деполяризующий
ток
Линейный сдвиг МП – «физический электротон»
Гиперполяризующий
ток
Нелинейный сдвиг МП – «физиологический электротон»
4.
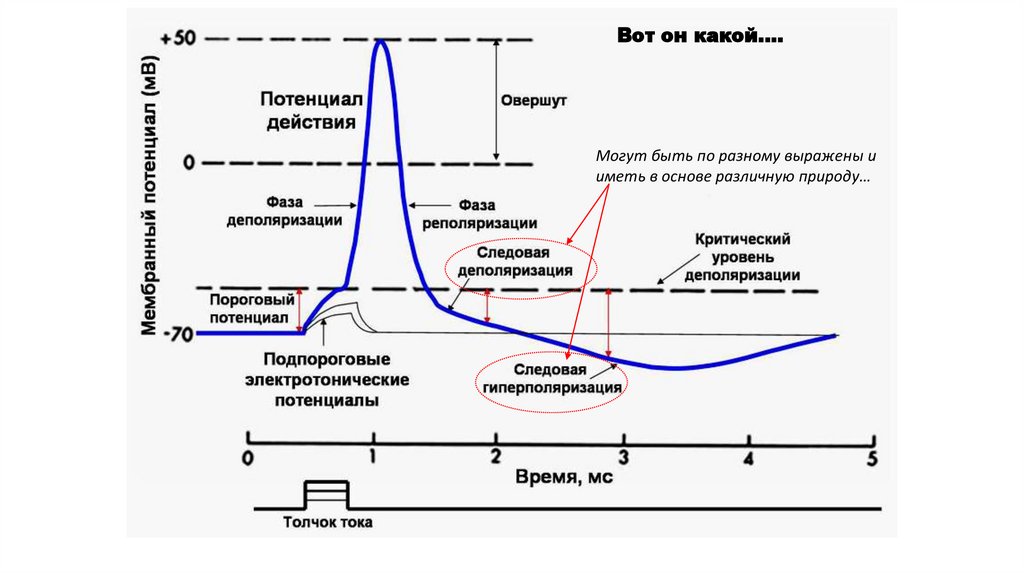
Вот он какой….Могут быть по разному выражены и
иметь в основе различную природу…
5.
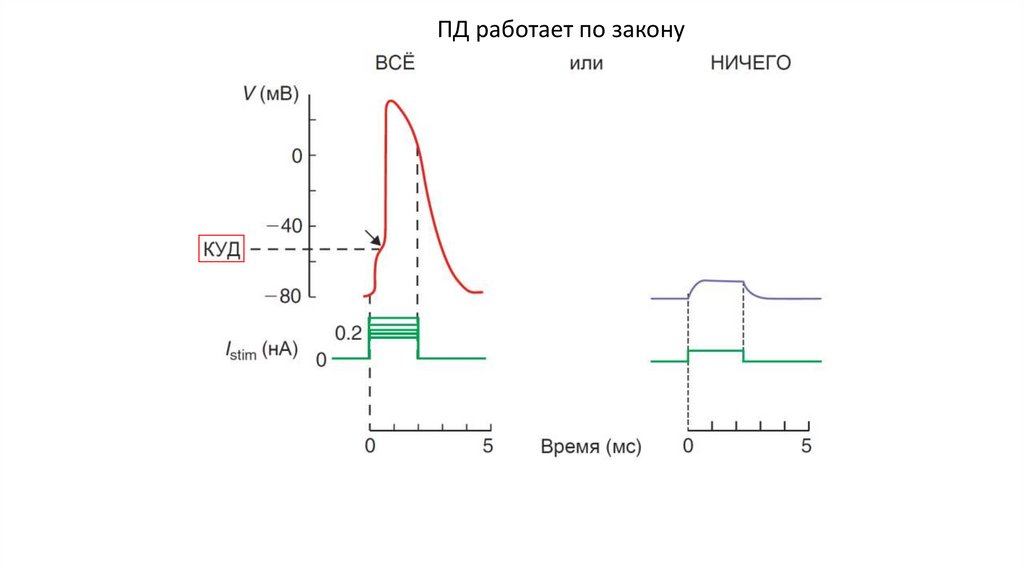
ПД работает по закону6.
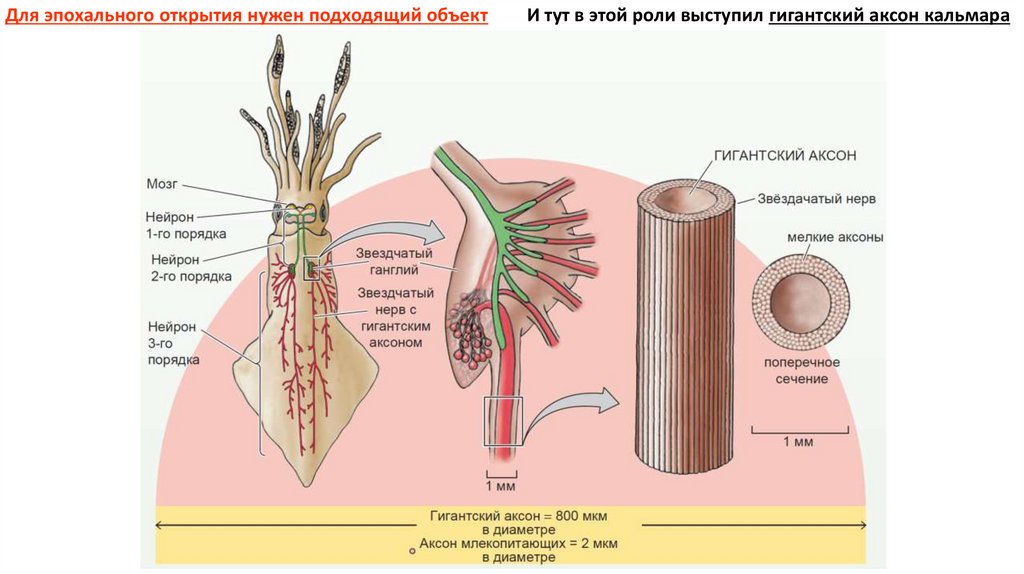
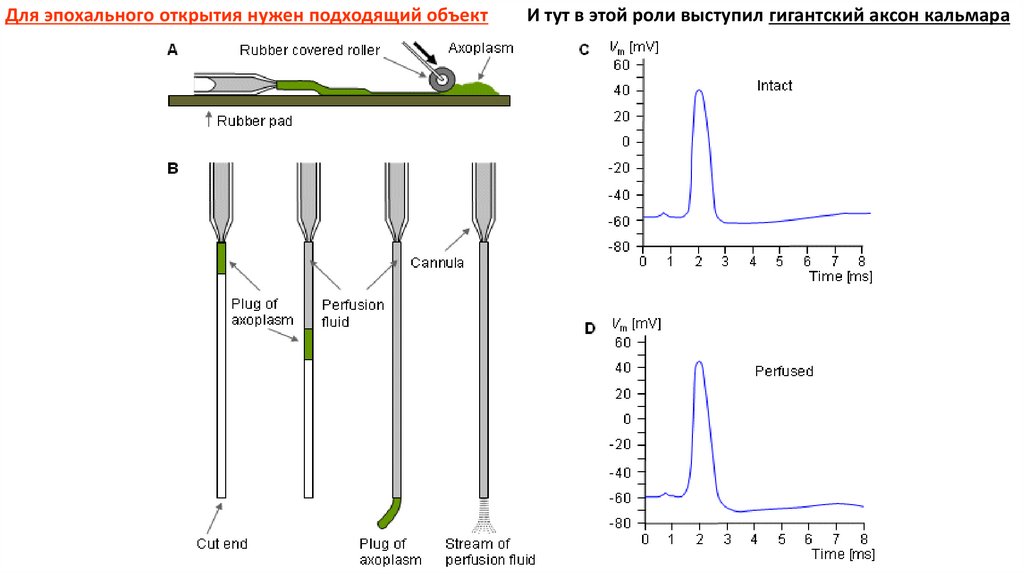
Для эпохального открытия нужен подходящий объектИ тут в этой роли выступил гигантский аксон кальмара
7.
Для эпохального открытия нужен подходящий объектИ тут в этой роли выступил гигантский аксон кальмара
8.
• 1939 - Ken Cole & Howard Curtis –– проводимость мембраны увеличивается (а сопротивление – снижается!) при развитии ПД• 1949 - Alan Hodgkin & Bernard Katz – показали ключевую роль ионов Na+ при развитии ПД
◄Изменение (уменьшение) содержания Na+ снаружи
Надо было изучить токи, текущие через мембрану, но…
Проблема:
протекание тока через мембрану ► изменение МП за счет
перераспределения зарядов, а изменение МП ► величина
протекающих токов
??? Что делать…
9.
Алан Ходжкин (справа)и
Эндрю Хаксли (слева)
RIP Huxley (†30.05.2012)
10.
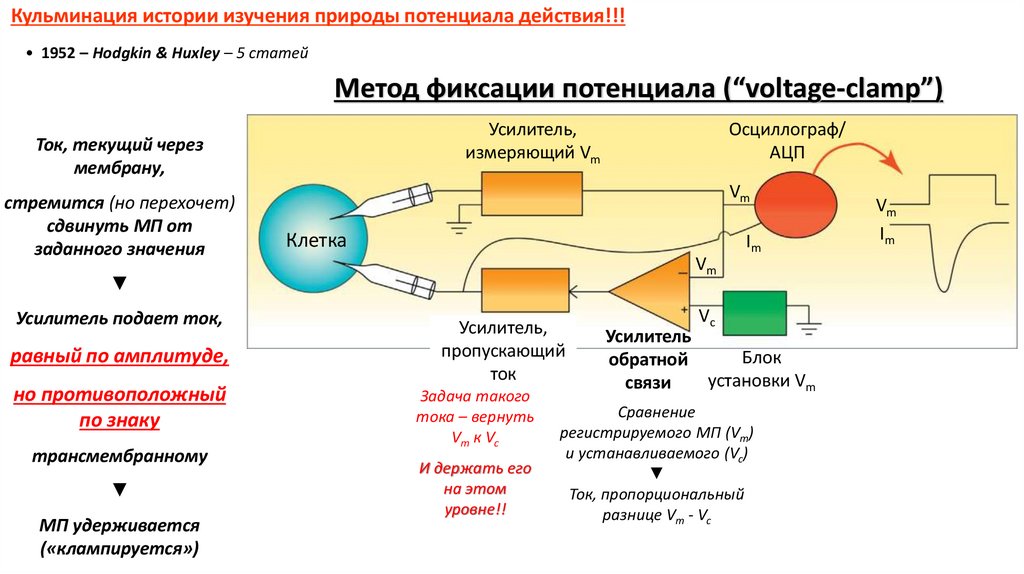
Кульминация истории изучения природы потенциала действия!!!• 1952 – Hodgkin & Huxley – 5 статей
Метод фиксации потенциала (“voltage-clamp”)
Усилитель,
измеряющий Vm
Ток, текущий через
мембрану,
стремится (но перехочет)
сдвинуть МП от
заданного значения
Vm
Клетка
Im
Vm
▼
Усилитель подает ток,
равный по амплитуде,
но противоположный
по знаку
трансмембранному
▼
МП удерживается
(«клампируется»)
Осциллограф/
АЦП
Усилитель,
пропускающий
ток
Задача такого
тока – вернуть
Vm к Vc
И держать его
на этом
уровне!!
Vс
Усилитель
Блок
обратной
установки Vm
связи
Сравнение
регистрируемого МП (Vm)
и устанавливаемого (Vc)
▼
Ток, пропорциональный
разнице Vm - Vc
Vm
Im
11.
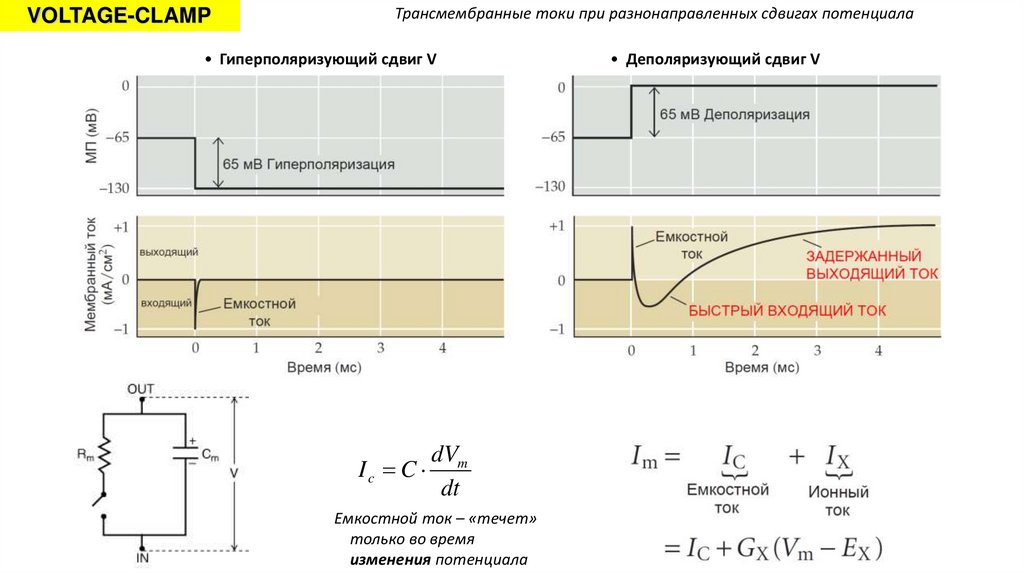
VOLTAGE-CLAMPТрансмембранные токи при разнонаправленных сдвигах потенциала
• Гиперполяризующий сдвиг V
Ic C
dVm
dt
Емкостной ток – «течет»
только во время
изменения потенциала
• Деполяризующий сдвиг V
12.
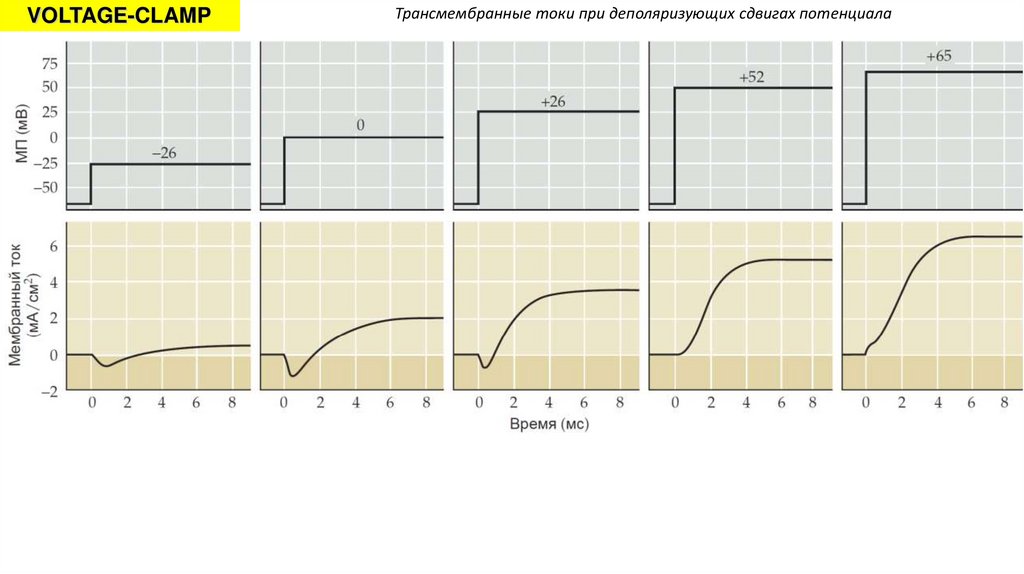
VOLTAGE-CLAMPТрансмембранные токи при деполяризующих сдвигах потенциала
13.
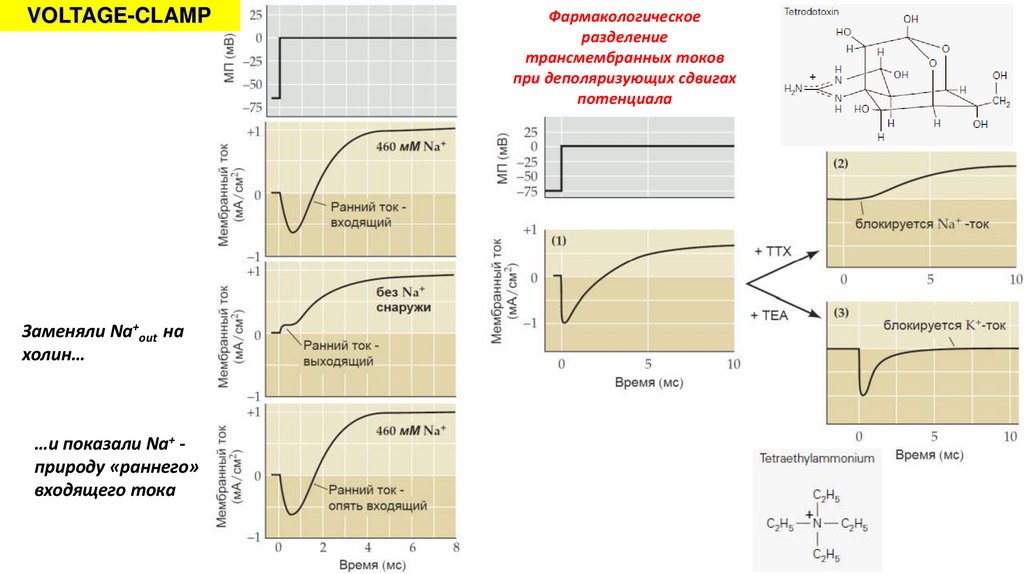
VOLTAGE-CLAMPЗаменяли Na+out на
холин…
…и показали Na+ природу «раннего»
входящего тока
Фармакологическое
разделение
трансмембранных токов
при деполяризующих сдвигах
потенциала
14.
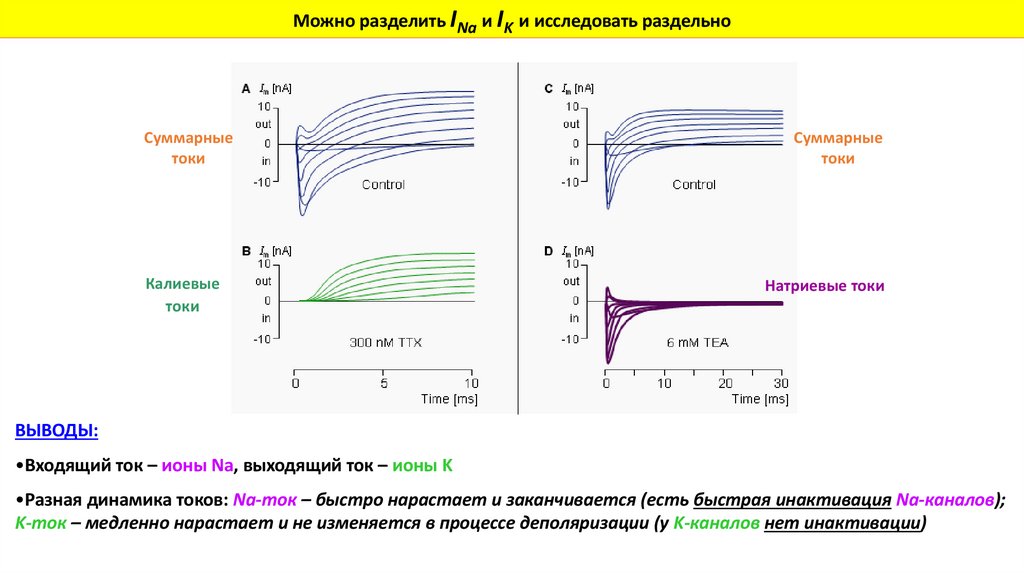
Можно разделить INa и IK и исследовать раздельноСуммарные
токи
Калиевые
токи
Суммарные
токи
Натриевые токи
ВЫВОДЫ:
•Входящий ток – ионы Na, выходящий ток – ионы K
•Разная динамика токов: Na-ток – быстро нарастает и заканчивается (есть быстрая инактивация Na-каналов);
K-ток – медленно нарастает и не изменяется в процессе деполяризации (у K-каналов нет инактивации)
15.
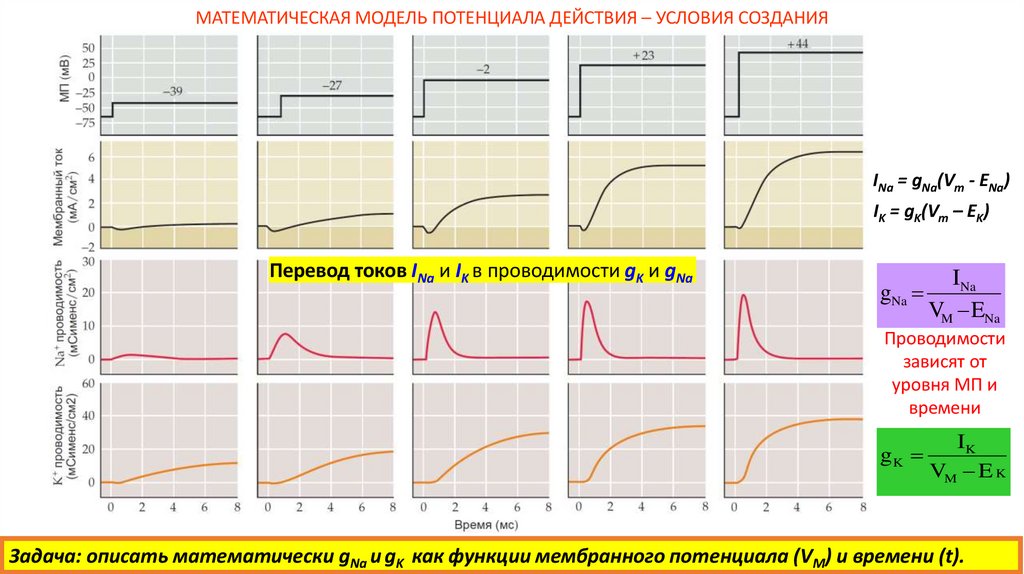
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ ПОТЕНЦИАЛА ДЕЙСТВИЯ – УСЛОВИЯ СОЗДАНИЯПеревод токов INa и IK в проводимости gK и gNa
INa = gNa(Vm - ENa)
IK = gK(Vm – EK)
Перевод токов INa и IK в проводимости gK и gNa
gNa
INa
VM ENa
Проводимости
зависят от
уровня МП и
времени
gK
IK
VM E K
Задача: описать математически gNa и gK как функции мембранного потенциала (VM) и времени (t).
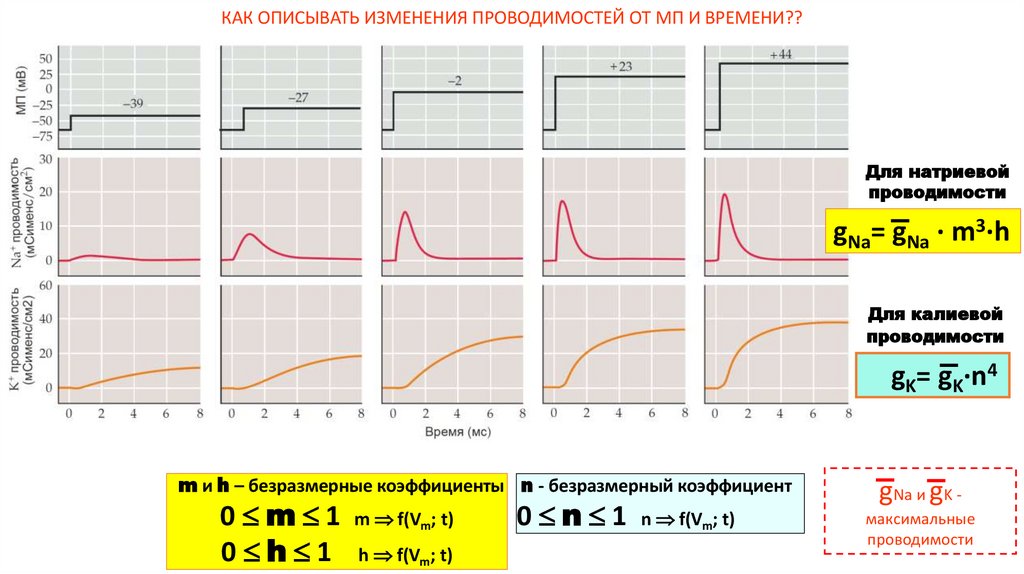
16.
КАК ОПИСЫВАТЬ ИЗМЕНЕНИЯ ПРОВОДИМОСТЕЙ ОТ МП И ВРЕМЕНИ??Для натриевой
проводимости
gNa= gNa · m3·h
Для калиевой
проводимости
gK= gK·n4
m и h – безразмерные коэффициенты n - безразмерный коэффициент
0 m 1 m f(Vm; t)
0 h 1 h f(Vm; t)
0 n 1 n f(Vm; t)
gNa и gK максимальные
проводимости
17.
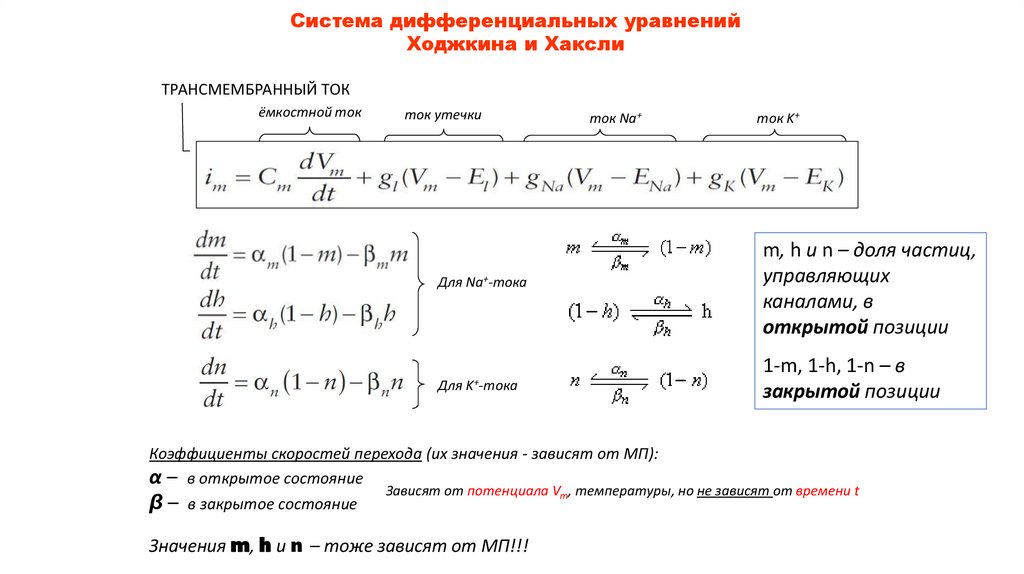
Система дифференциальных уравненийХоджкина и Хаксли
ТРАНСМЕМБРАННЫЙ ТОК
ёмкостной ток
ток утечки
ток Na+
Для Na+-тока
m, h и n – доля частиц,
управляющих
каналами, в
открытой позиции
Для K+-тока
1-m, 1-h, 1-n – в
закрытой позиции
Коэффициенты скоростей перехода (их значения - зависят от МП):
α – в открытое состояние
β – в закрытое состояние
ток K+
Зависят от потенциала Vm, температуры, но не зависят от времени t
Значения m, h и n – тоже зависят от МП!!!
18.
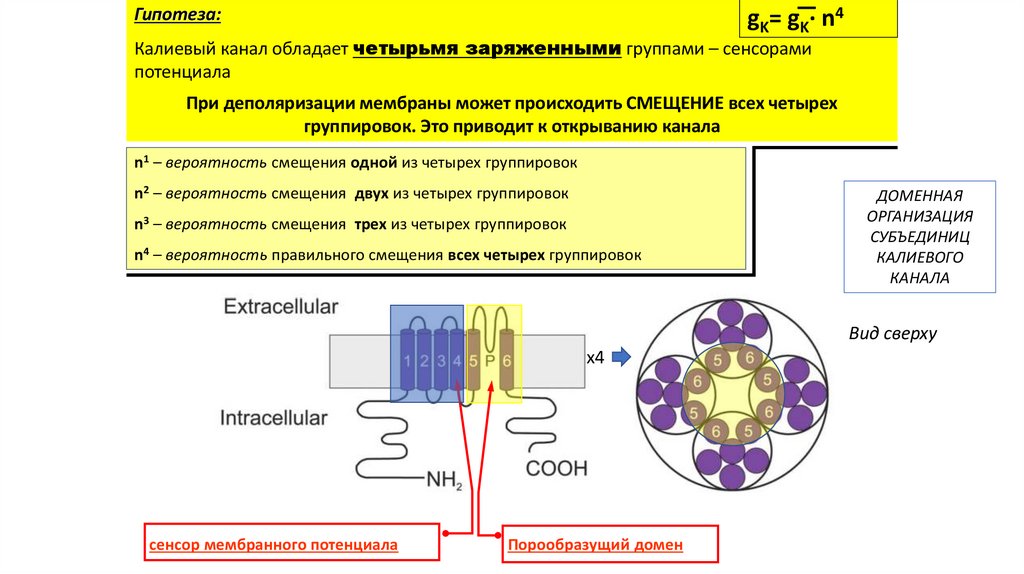
Гипотеза:gK= gK· n4
Калиевый канал обладает четырьмя заряженными группами – сенсорами
потенциала
При деполяризации мембраны может происходить СМЕЩЕНИЕ всех четырех
группировок. Это приводит к открыванию канала
n1 – вероятность смещения одной из четырех группировок
n2 – вероятность смещения двух из четырех группировок
n3 – вероятность смещения трех из четырех группировок
n4 – вероятноcть правильного смещения всех четырех группировок
19.
gK= gK· n4Гипотеза:
Калиевый канал обладает четырьмя заряженными группами – сенсорами
потенциала
При деполяризации мембраны может происходить СМЕЩЕНИЕ всех четырех
группировок. Это приводит к открыванию канала
n1 – вероятность смещения одной из четырех группировок
n2 – вероятность смещения двух из четырех группировок
n3 – вероятность смещения трех из четырех группировок
n4 – вероятноcть правильного смещения всех четырех группировок
ДОМЕННАЯ
ОРГАНИЗАЦИЯ
СУБЪЕДИНИЦ
КАЛИЕВОГО
КАНАЛА
Вид сверху
x4
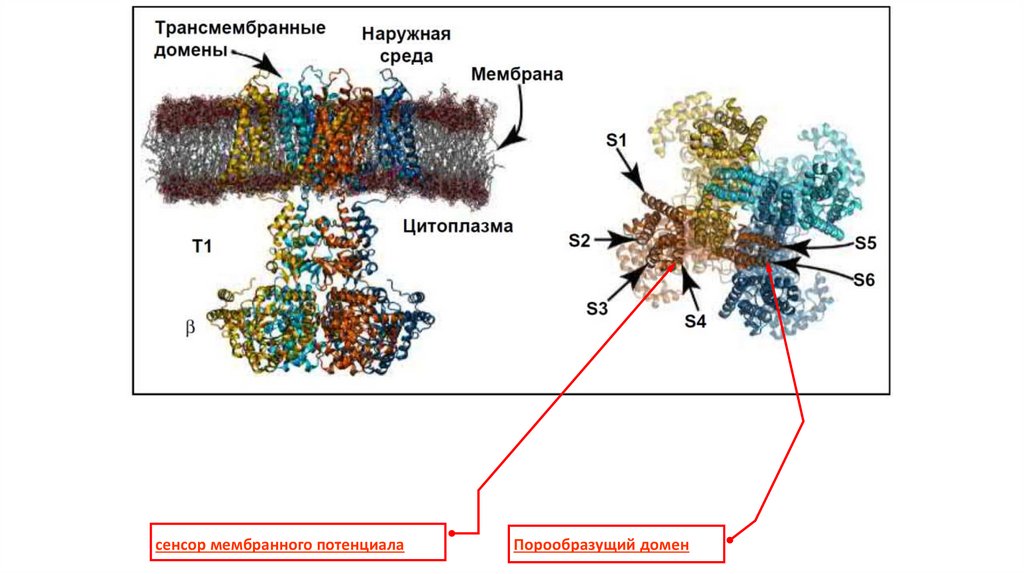
сенсор мембранного потенциала
Порообразущий домен
20.
сенсор мембранного потенциалаПорообразущий домен
21.
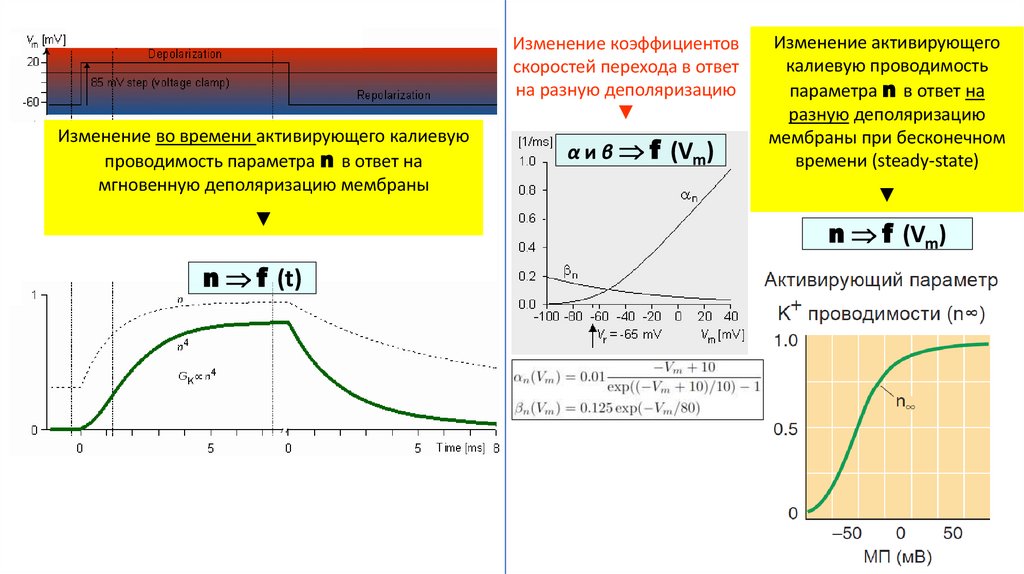
Изменение коэффициентовскоростей перехода в ответ
на разную деполяризацию
▼
Изменение во времени активирующего калиевую
проводимость параметра n в ответ на
мгновенную деполяризацию мембраны
▼
n f (t)
α и β f (Vm)
Изменение активирующего
калиевую проводимость
параметра n в ответ на
разную деполяризацию
мембраны при бесконечном
времени (steady-state)
▼
n f (Vm)
22.
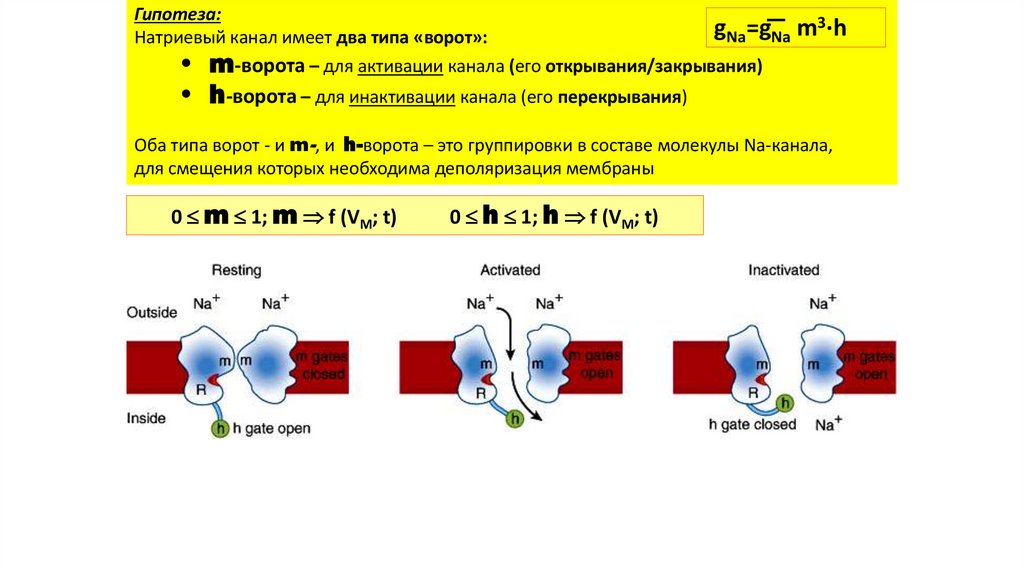
Гипотеза:Натриевый канал имеет два типа «ворот»:
gNa=gNa m3·h
• m-ворота – для активации канала (его открывания/закрывания)
• h-ворота – для инактивации канала (его перекрывания)
Оба типа ворот - и m-, и h-ворота – это группировки в составе молекулы Na-канала,
для смещения которых необходима деполяризация мембраны
0 m 1; m f (VM; t)
0 h 1; h f (VM; t)
23.
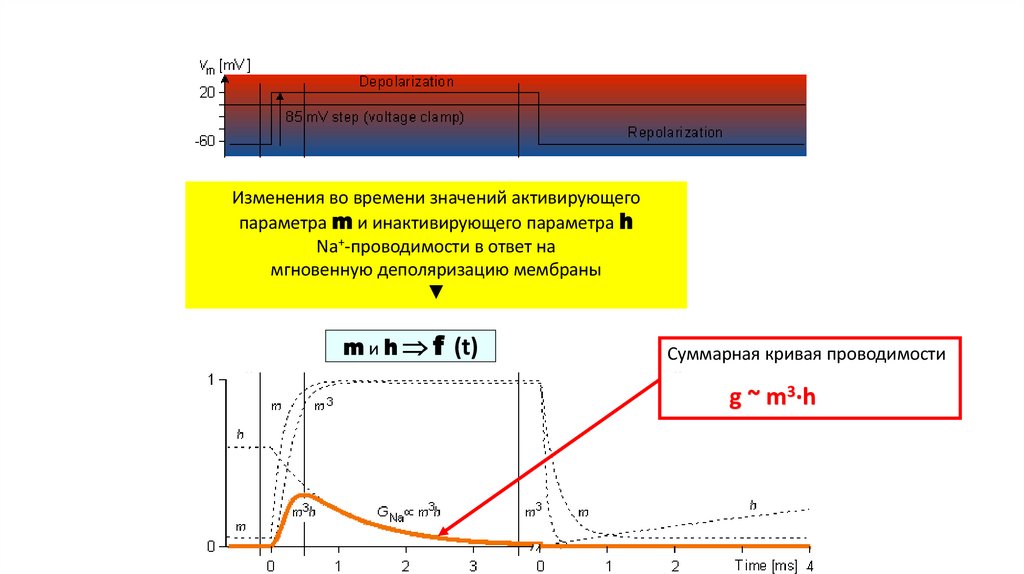
Изменения во времени значений активирующегопараметра m и инактивирующего параметра h
Na+-проводимости в ответ на
мгновенную деполяризацию мембраны
▼
m и h f (t)
Суммарная кривая проводимости
g ~ m3·h
24.
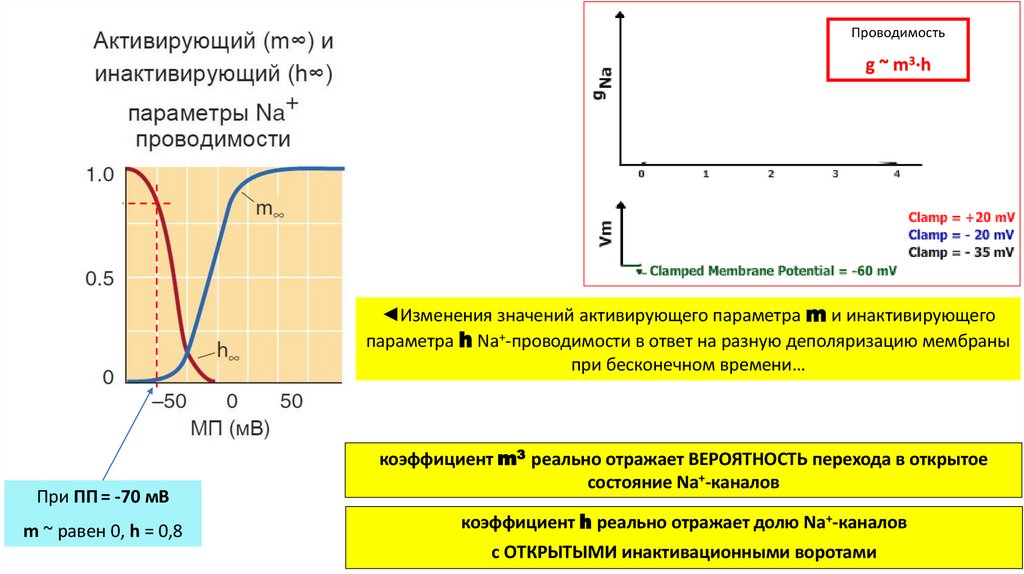
Проводимостьg ~ m3·h
◄Изменения значений активирующего параметра m и инактивирующего
параметра h Na+-проводимости в ответ на разную деполяризацию мембраны
при бесконечном времени…
При ПП = -70 мВ
m ~ равен 0, h = 0,8
коэффициент m3 реально отражает ВЕРОЯТНОСТЬ перехода в открытое
состояние Nа+-каналов
коэффициент h реально отражает долю Na+-каналов
с ОТКРЫТЫМИ инактивационными воротами
25.
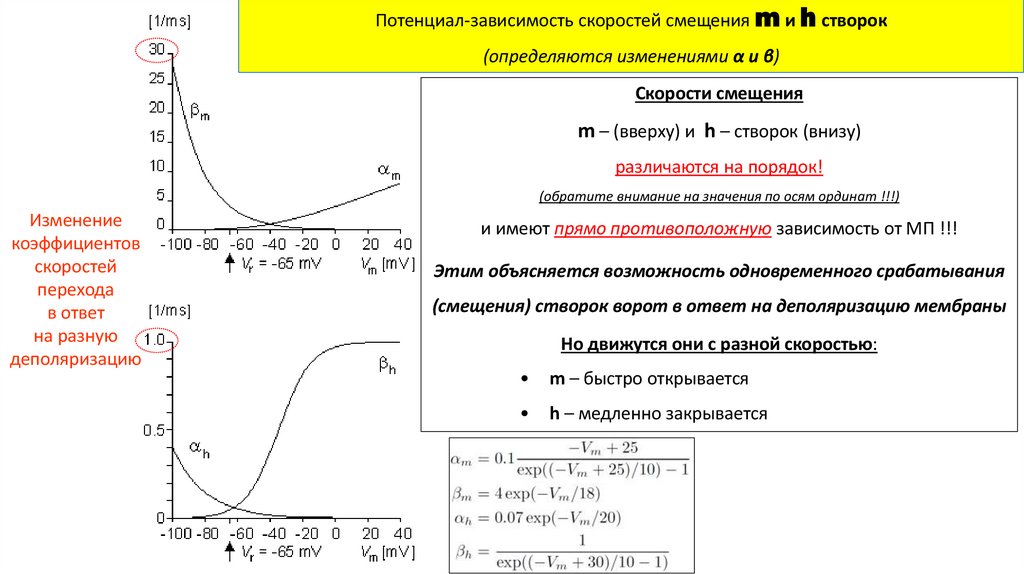
Потенциал-зависимость скоростей смещения m и h створок(определяются изменениями α и β)
Скорости смещения
m – (вверху) и h – створок (внизу)
различаются на порядок!
(обратите внимание на значения по осям ординат !!!)
Изменение
коэффициентов
скоростей
перехода
в ответ
на разную
деполяризацию
и имеют прямо противоположную зависимость от МП !!!
Этим объясняется возможность одновременного срабатывания
(смещения) створок ворот в ответ на деполяризацию мембраны
Но движутся они с разной скоростью:
m – быстро открывается
h – медленно закрывается
26.
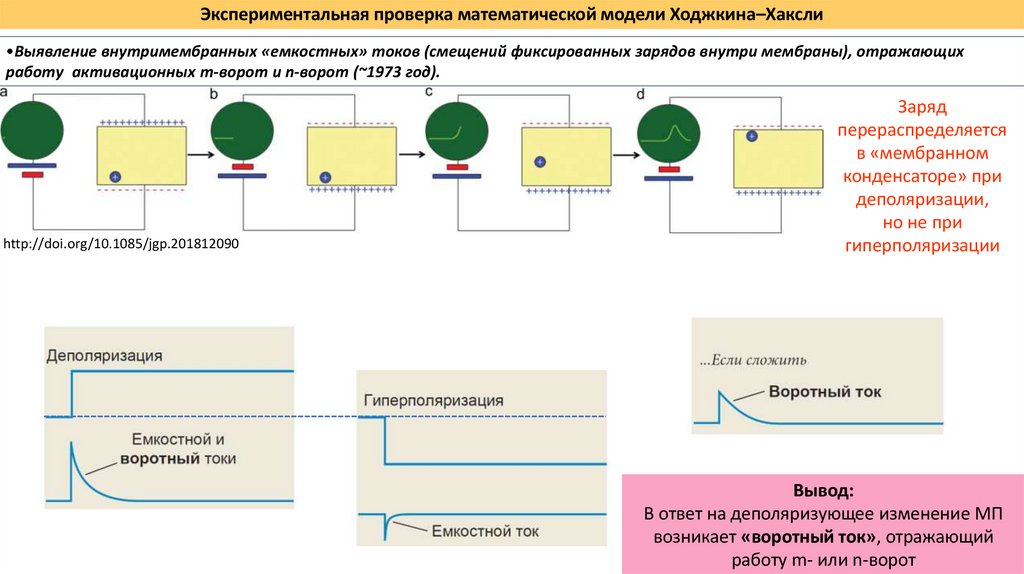
Экспериментальная проверка математической модели Ходжкина–Хаксли•Выявление внутримембранных «емкостных» токов (смещений фиксированных зарядов внутри мембраны), отражающих
работу активационных m-ворот и n-ворот (~1973 год).
http://doi.org/10.1085/jgp.201812090
Заряд
перераспределяется
в «мембранном
конденсаторе» при
деполяризации,
но не при
гиперполяризации
Вывод:
В ответ на деполяризующее изменение МП
возникает «воротный ток», отражающий
работу m- или n-ворот
27.
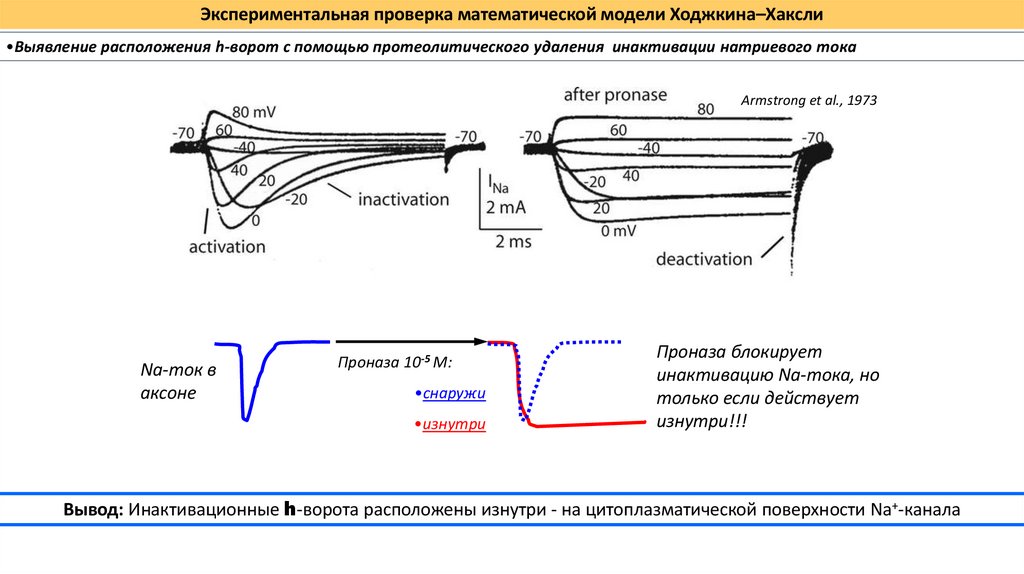
Экспериментальная проверка математической модели Ходжкина–Хаксли•Выявление расположения h-ворот с помощью протеолитического удаления инактивации натриевого тока
Armstrong et al., 1973
Na-ток в
аксоне
Проназа 10-5 М:
•снаружи
•изнутри
Проназа блокирует
инактивацию Na-тока, но
только если действует
изнутри!!!
Вывод: Инактивационные h-ворота расположены изнутри - на цитоплазматической поверхности Na+-канала
28.
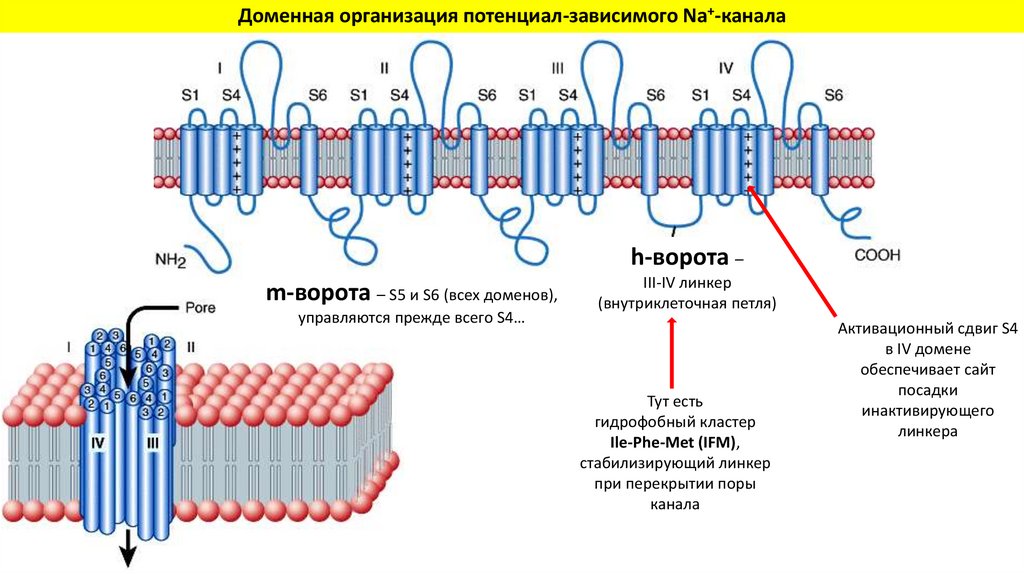
Доменная организация потенциал-зависимого Na+-каналаh-ворота –
m-ворота – S5 и S6 (всех доменов),
управляются прежде всего S4…
III-IV линкер
(внутриклеточная петля)
Тут есть
гидрофобный кластер
Ile-Phe-Met (IFM),
стабилизирующий линкер
при перекрытии поры
канала
Активационный сдвиг S4
в IV домене
обеспечивает сайт
посадки
инактивирующего
линкера
29.
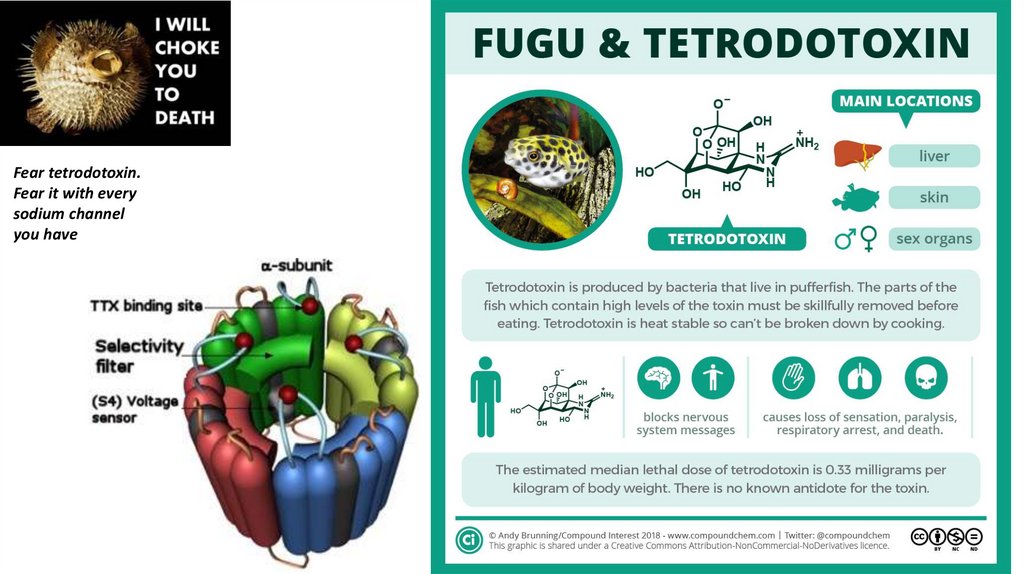
Fear tetrodotoxin.Fear it with every
sodium channel
you have
30.
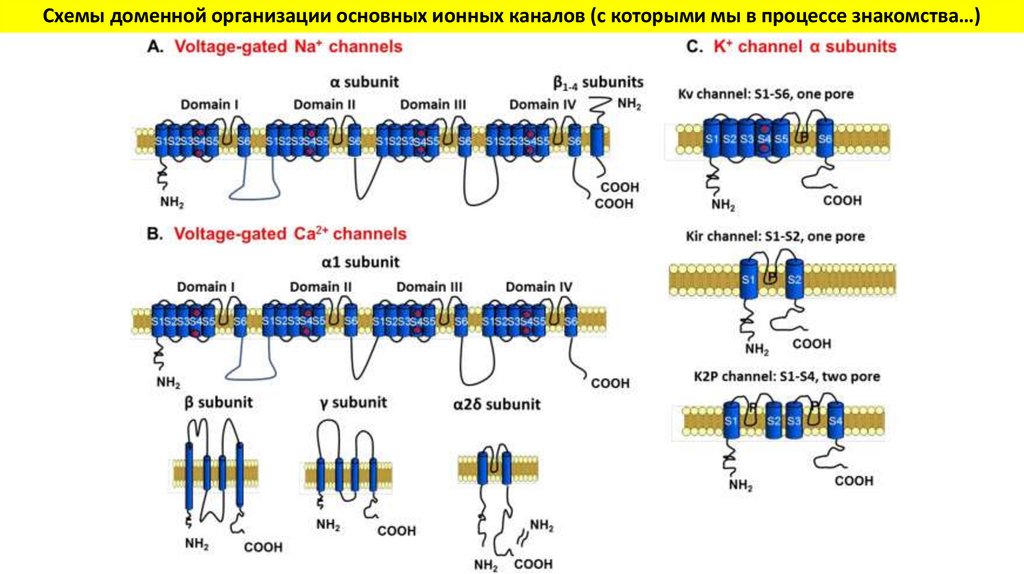
Схемы доменной организации основных ионных каналов (с которыми мы в процессе знакомства…)31.
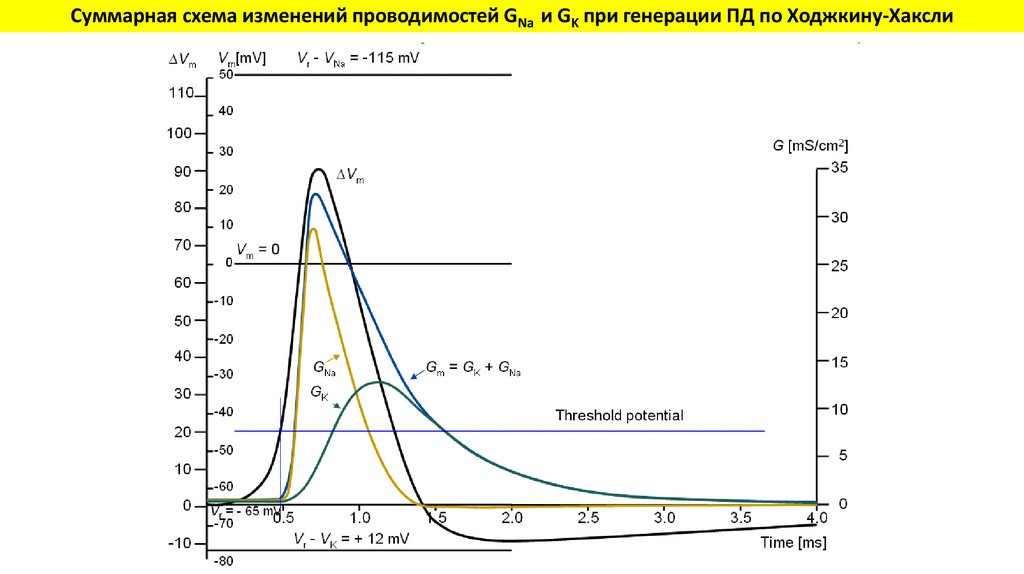
Суммарная схема изменений проводимостей GNa и GK при генерации ПД по Ходжкину-Хаксли32.
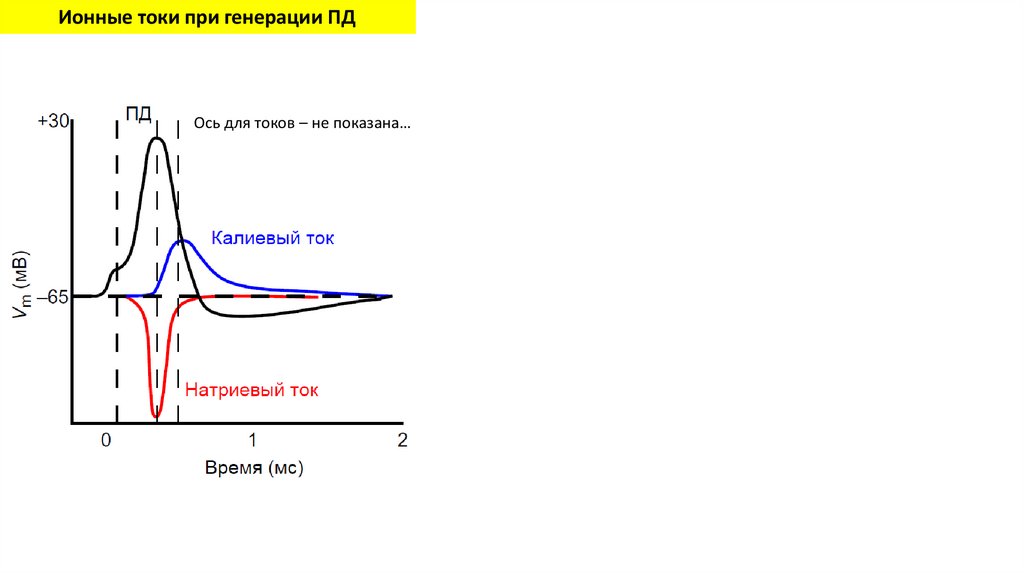
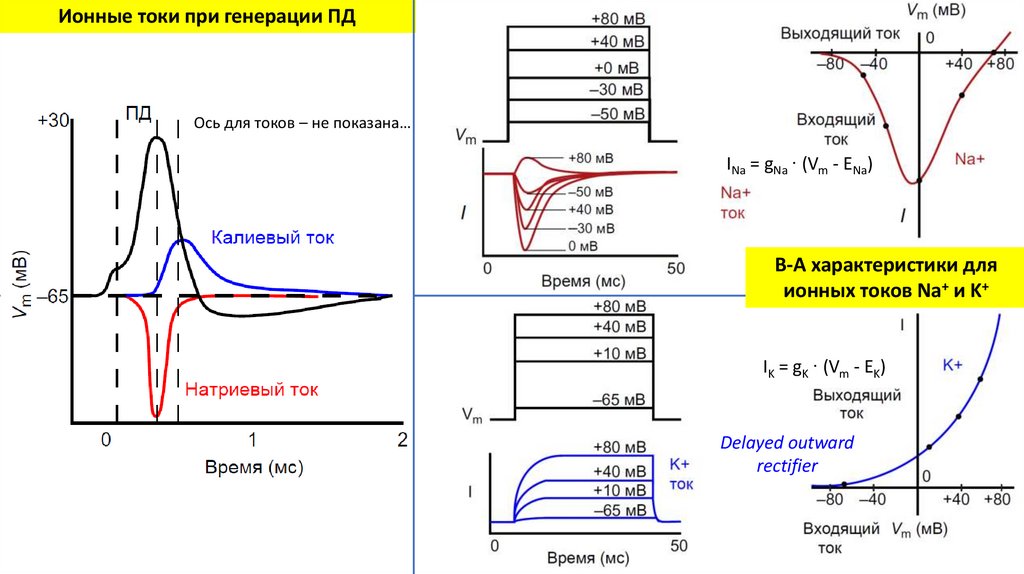
Ионные токи при генерации ПДОсь для токов – не показана…
33.
Ионные токи при генерации ПДОсь для токов – не показана…
INa = gNa ∙ (Vm - ENa)
В-А характеристики для
ионных токов Na+ и K+
IK = gK ∙ (Vm - EK)
Delayed outward
rectifier
34.
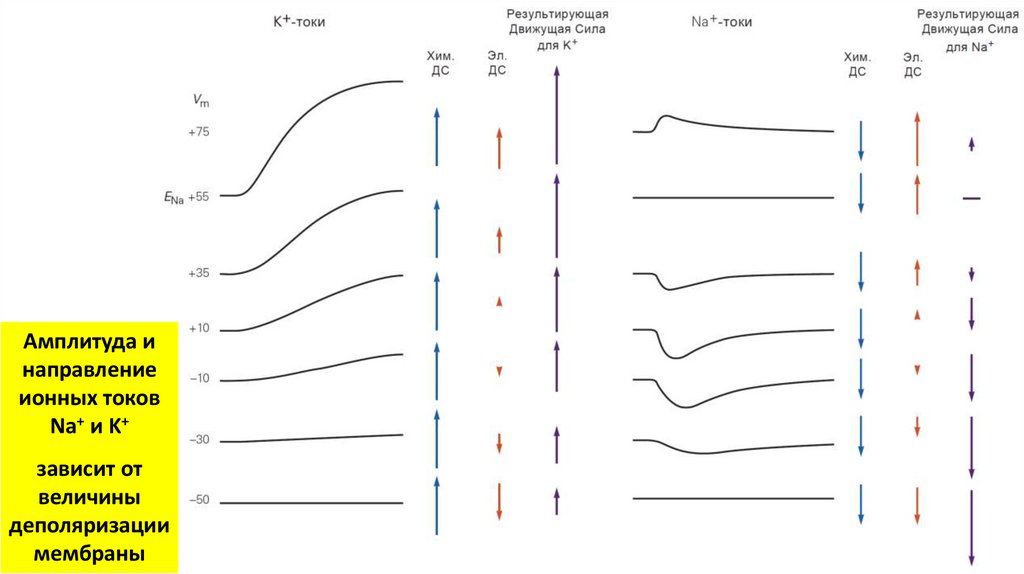
Амплитуда инаправление
ионных токов
Na+ и K+
зависит от
величины
деполяризации
мембраны
35.
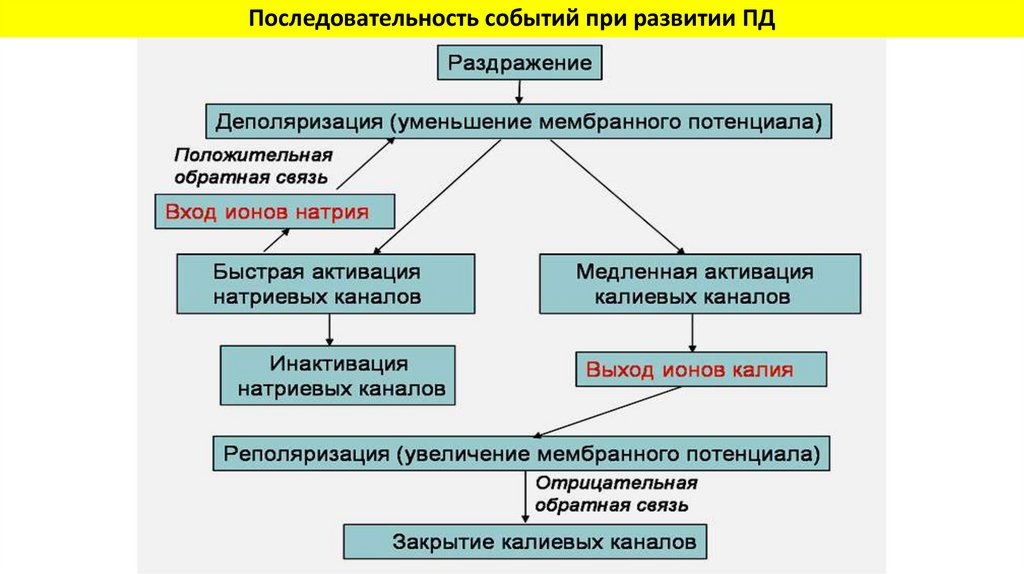
Последовательность событий при развитии ПД36.
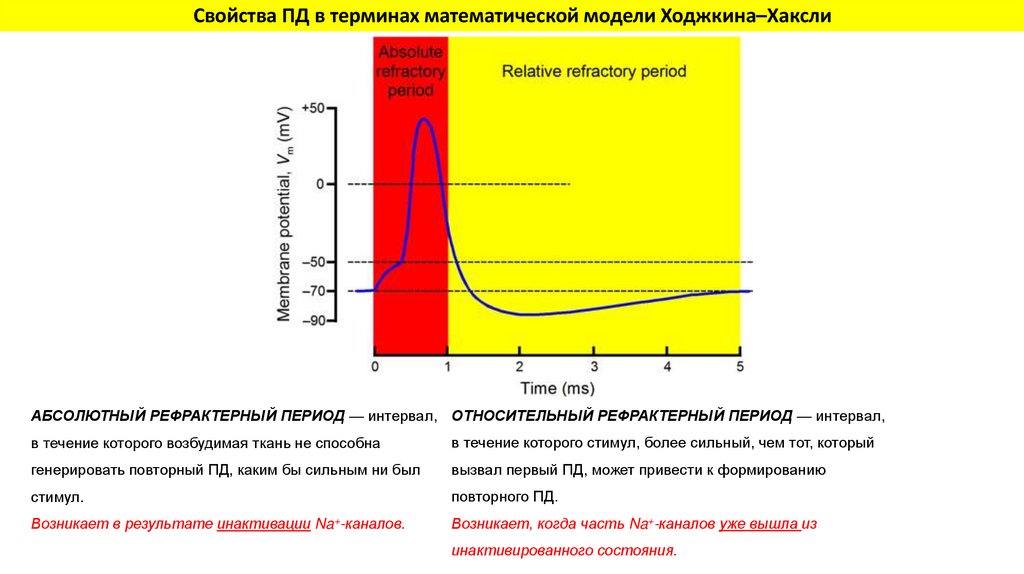
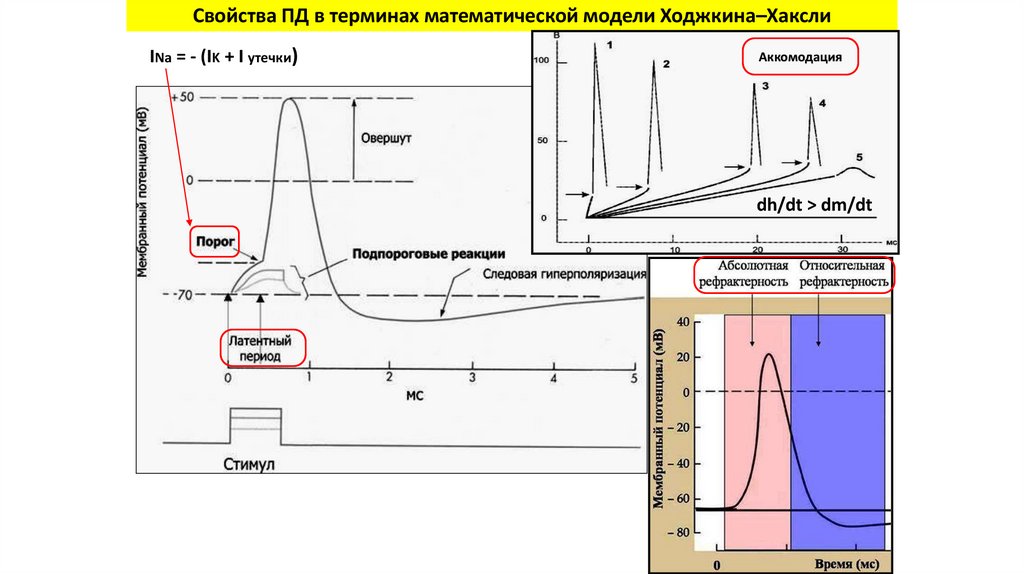
Свойства ПД в терминах математической модели Ходжкина–ХакслиАБСОЛЮТНЫЙ РЕФРАКТЕРНЫЙ ПЕРИОД — интервал, ОТНОСИТЕЛЬНЫЙ РЕФРАКТЕРНЫЙ ПЕРИОД — интервал,
в течение которого возбудимая ткань не способна
в течение которого стимул, более сильный, чем тот, который
генерировать повторный ПД, каким бы сильным ни был
вызвал первый ПД, может привести к формированию
стимул.
повторного ПД.
Возникает в результате инактивации Na+-каналов.
Возникает, когда часть Na+-каналов уже вышла из
инактивированного состояния.
37.
Свойства ПД в терминах математической модели Ходжкина–ХакслиINa = - (IK + I утечки)
Аккомодация
dh/dt > dm/dt
38.
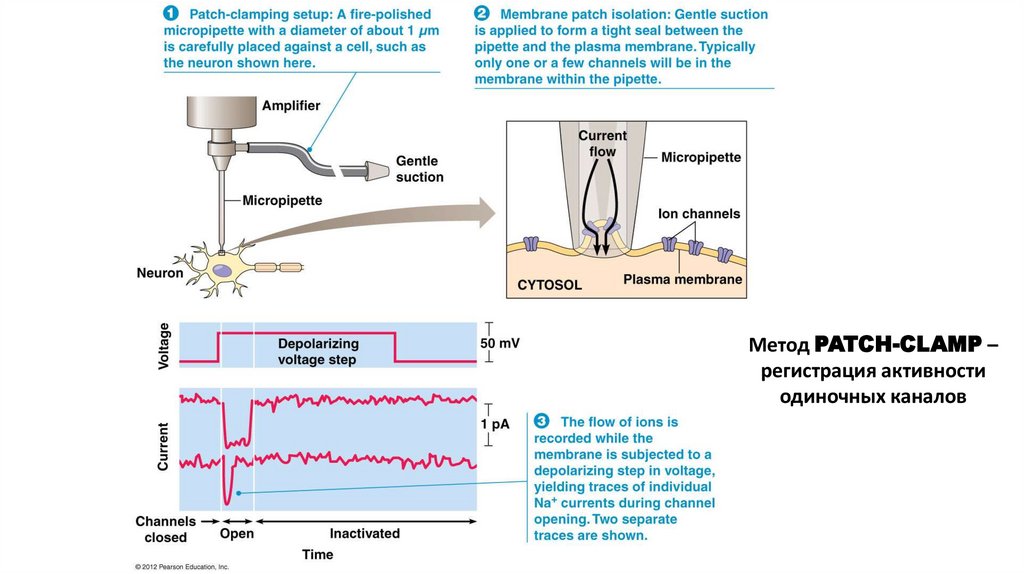
Метод PATCH-CLAMP –регистрация активности
одиночных каналов
39.
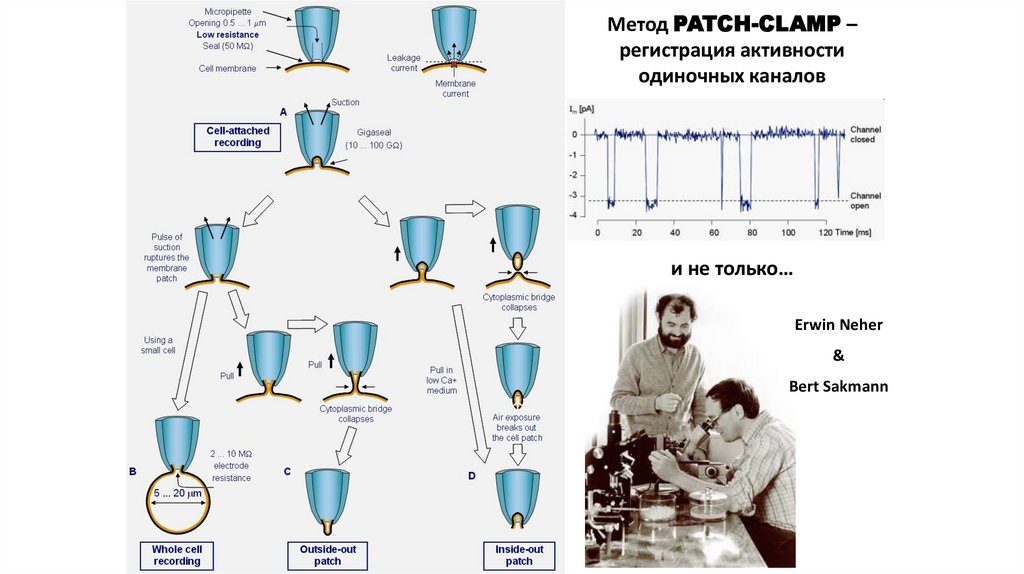
Метод PATCH-CLAMP –регистрация активности
одиночных каналов
и не только…
Erwin Neher
&
Bert Sakmann
40.
Какой-точувак…
▼
Erwin
Neher
▼
FENS Forum – July 2018 , Berlin, Germany
41.
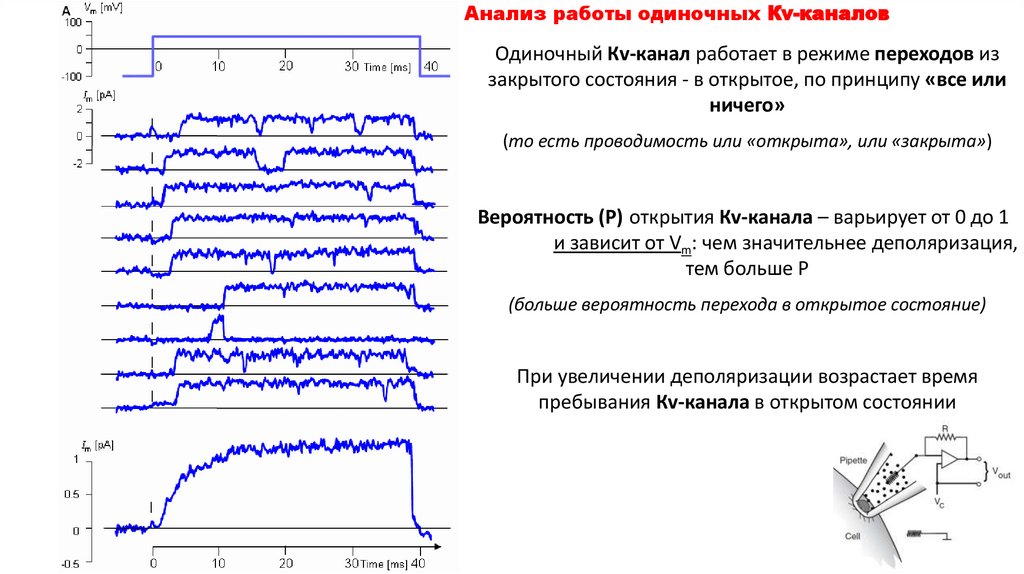
Анализ работы одиночных Кv-каналовОдиночный Кv-канал работает в режиме переходов из
закрытого состояния - в открытое, по принципу «все или
ничего»
(то есть проводимость или «открыта», или «закрыта»)
Вероятность (Р) открытия Кv-канала – варьирует от 0 до 1
и зависит от Vm: чем значительнее деполяризация,
тем больше Р
(больше вероятность перехода в открытое состояние)
При увеличении деполяризации возрастает время
пребывания Кv-канала в открытом состоянии
42.
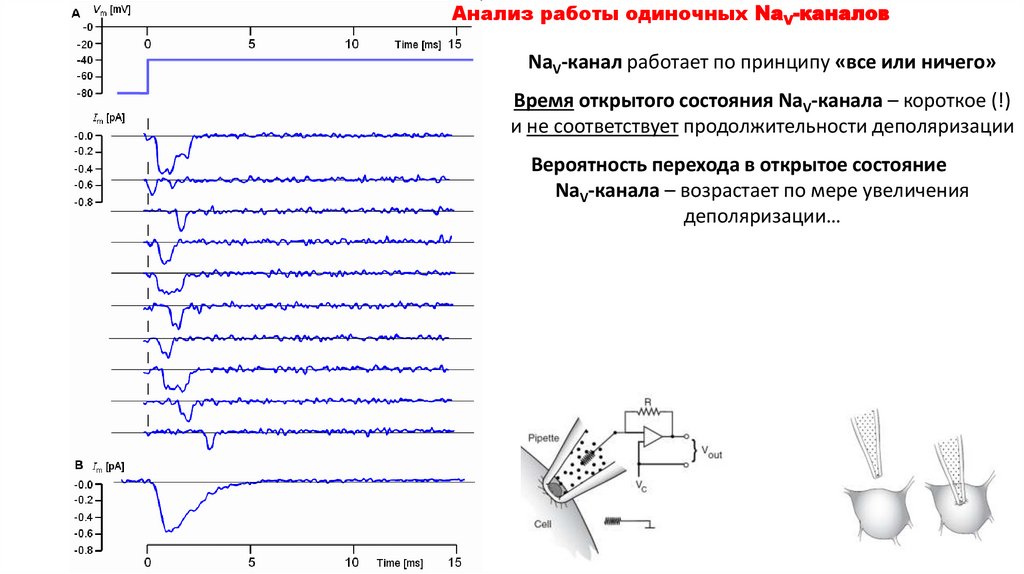
Анализ работы одиночных NaV-каналовNaV-канал работает по принципу «все или ничего»
Время открытого состояния NaV-канала – короткое (!)
и не соответствует продолжительности деполяризации
Вероятность перехода в открытое состояние
NaV-канала – возрастает по мере увеличения
деполяризации…
43.
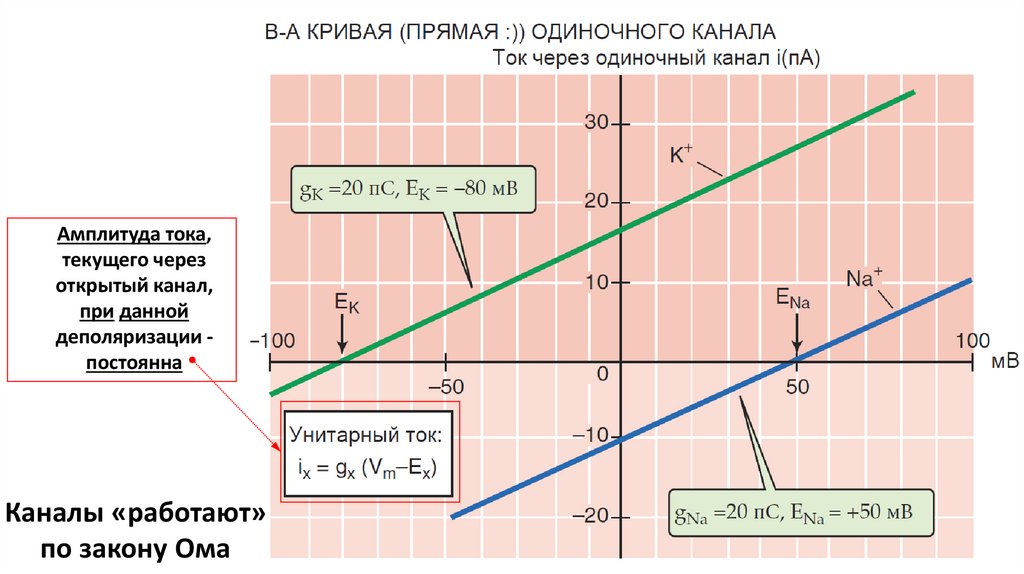
Амплитуда тока,текущего через
открытый канал,
при данной
деполяризации постоянна
Каналы «работают»
по закону Ома
44.
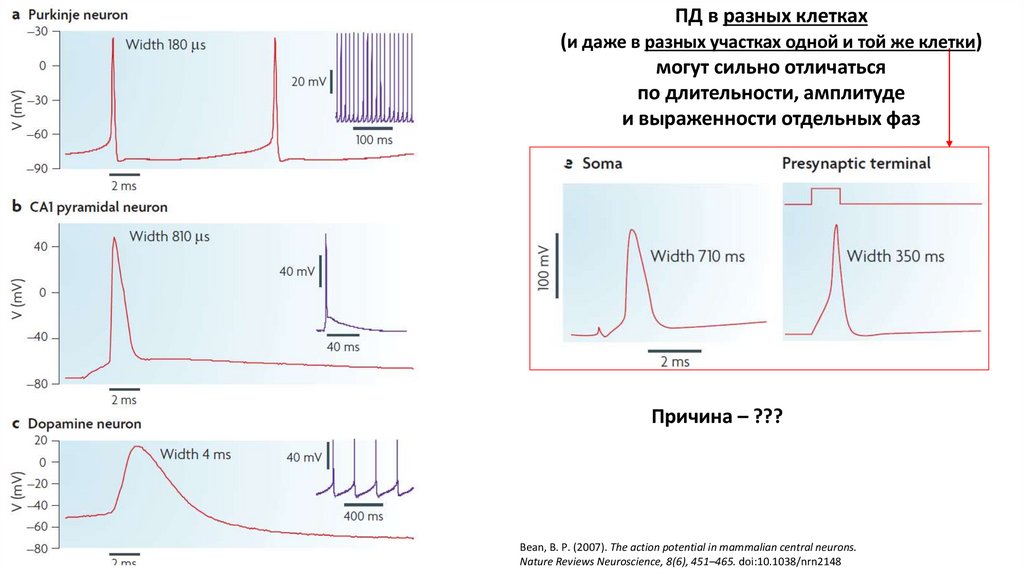
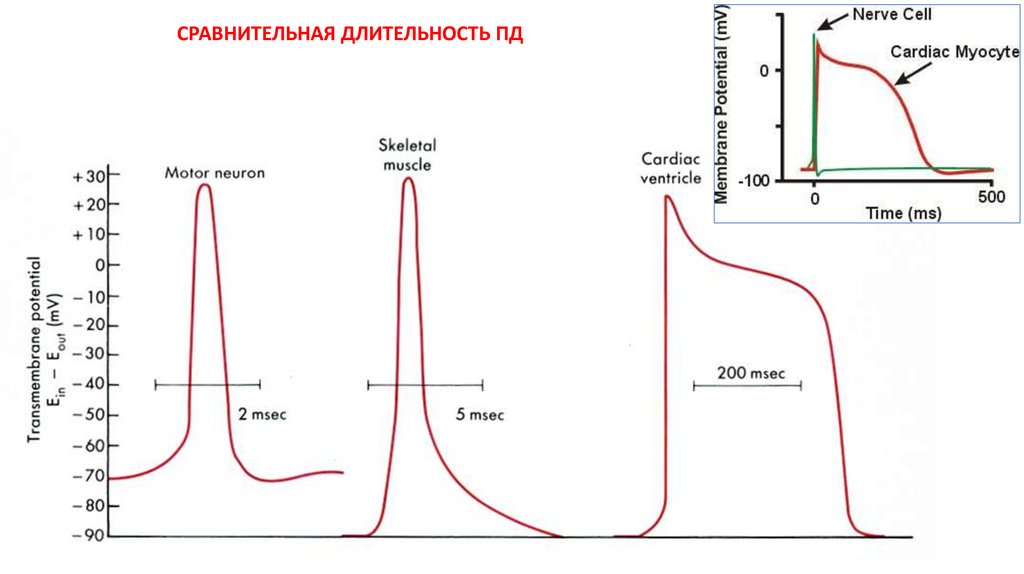
ПД в разных клетках(и даже в разных участках одной и той же клетки)
могут сильно отличаться
по длительности, амплитуде
и выраженности отдельных фаз
Причина – ???
Bean, B. P. (2007). The action potential in mammalian central neurons.
Nature Reviews Neuroscience, 8(6), 451–465. doi:10.1038/nrn2148
45.
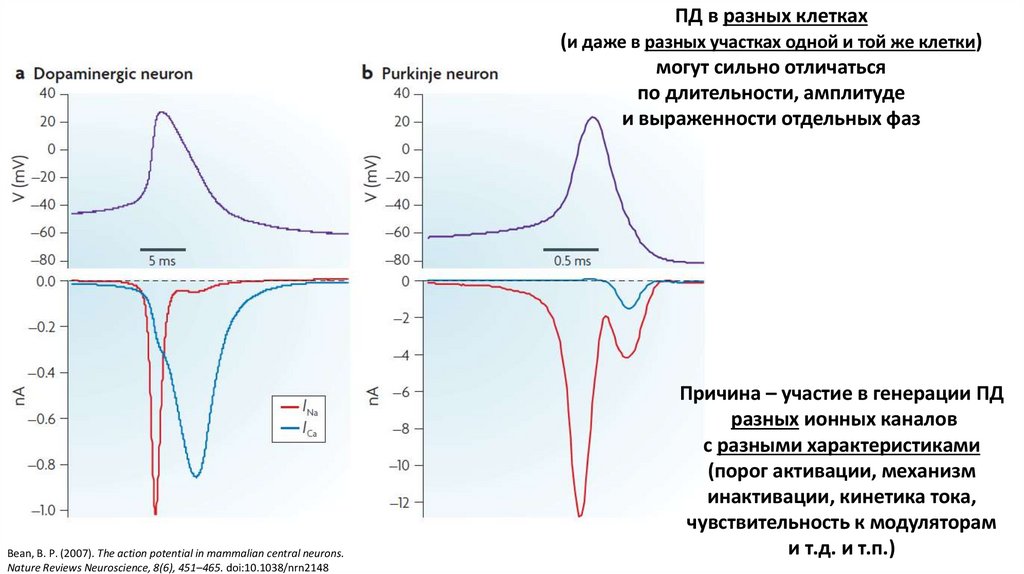
ПД в разных клетках(и даже в разных участках одной и той же клетки)
могут сильно отличаться
по длительности, амплитуде
и выраженности отдельных фаз
Bean, B. P. (2007). The action potential in mammalian central neurons.
Nature Reviews Neuroscience, 8(6), 451–465. doi:10.1038/nrn2148
Причина – участие в генерации ПД
разных ионных каналов
с разными характеристиками
(порог активации, механизм
инактивации, кинетика тока,
чувствительность к модуляторам
и т.д. и т.п.)
46.
СРАВНИТЕЛЬНАЯ ДЛИТЕЛЬНОСТЬ ПД47.
МЕХАНИЗМЫ ПРОВЕДЕНИЯ ПД – НА СЛЕДУЮЩЕЙ ЛЕКЦИИЗАКЛЮЧЕНИЕ
•Интегральные натриевый и
калиевый токи, регистрируемые
при генерации ПД, отражают
потенциал-зависимость и
временной ход натриевой и
калиевой проводимостей (g)
мембраны.
•Изменения gNa и gK отражают
особенности перехода и
пребывания в открытом состоянии
отдельных натриевых и калиевых
каналов, а также инактивированного состояния
отдельных натриевых каналов.
48.
Спасибо за внимание…Вопросы???
Alan Lloyd
Hodgkin
Andrew Fielding
Huxley
The Nobel Prize
in Physiology or Medicine 1963
"for their discoveries concerning
the ionic mechanisms involved in
excitation and inhibition in the
peripheral and central portions of
the nerve cell membrane"
Электрофизиолог
и пытаются
отобрать у
кальмара
гигантский
аксон…