























")

 обнаружил, что особые длинные тяжи у кальмаров и каракатиц являются не кровеносными сосудами, как считалось")



")

























 biology
biologySimilar presentations:








")

Основы электрофизиологии. Общая характеристика возбудимых тканей. Потенциал покоя. Потенциал действия
1.
Лекция №2Основы электрофизиологии
Общая характеристика
возбудимых тканей.
Потенциал покоя.
Потенциал действия.
2.
2Основные проявления
жизнедеятельности
Физиологический покой
Физиологическая активность
Раздражение
Возбуждение
Торможение
3. 3
Общая характеристика возбудимых тканей.Все клетки нашего организма обладают свойством
раздражимости.
Раздражимость – это способность клеток переходить из
состояния
физиологического
покоя
в
состояние
функциональной активности при действии внешних или
внутренних стимулов.
Три вида тканей: нервная, мышечная и секреторная
обладают особой формой раздражимости – возбудимостью.
Возбудимость
–
способность
возбуждением на внешние стимулы.
ткани
реагировать
Возбуждение – реакция на раздражение путем изменения
мембранного потенциала.
4. 4 Раздражение и виды раздражителей.
• Раздражение – это процесс воздействия на живойобъект внешних по отношению к нему факторов.
• Раздражители – факторы внешней среды, вызывающие
переход биосистемы в активное состояние.
Раздражители разделяют по следующим факторам:
природе раздражителей;
биологическому значению;
количественному признаку.
5. 5 По природе раздражителя:
• 1. физические ( температура, звук, свет, электрический ток и т.д.).• 2. химические (соли, кислоты, щелочи и т.д.).
• 3. физико-химические (изменение осмоса, парциального давления
газов, рН).
• 4. биологические (вирусы, бактерии, токсины антитела и т.д.).
6. 6 По биологическому значению:
1. адекватные – раздражитель к которому в естественных условиях убиосистем есть специализированные воспринимающие структуры
(рецепторы).
Свет – для фоторецепторов;
Упругие механические колебания среды – для рецепторов слуха.
2. неадекватные – раздражители не являющиеся в естественных
условиях средством возбуждения для данных биоструктур.
7. 7 По количественному признаку или порогу раздражения:
В зависимости от силы действия раздражителиделятся на:
1. пороговые – минимальная сила раздражителя,
способная вызвать возбуждение.
подпороговые – сила раздражения ниже
порога возбуждения.
2. максимальные – минимальная сила раздражителя,
вызывающая максимальный ответ ткани.
Раздражители, сила которых меньше или больше
максимальной называются, соответственно:
субмаксимальными и супермаксимальными.
8. 8 Все возбудимые ткани обладают общими свойствами:
Возбудимостью
Проводимостью
Лабильностью
Аккомодацией
9. 9
Возбудимостьколичественно
характеризуется
порогом возбуждения, который определяется
минимальной
силой
раздражения,
способной
вызывать возбуждение.
Проводимость
–
способность
проводить
возбуждение.
Оценивается
по
скорости
распространения
возбуждения (м/с).
Лабильность – функциональная подвижность ткани.
Оценивается по максимальному числу импульсов,
которое возбудимая ткань способна воспроизвести в
соответствии с частотой раздражения.
Нерв – до 1000 П Д/с, мышца – 500 ПД/с, синапс –
50 ПД/с.
Аккомодация – повышение порога возбуждения к
медленно
нарастающему
или
постоянно
действующему раздражителю.
10. 10 Законы раздражения:
• Закон силы• Закон времени
• Закон крутизны нарастания
раздражителя
11. 11 Закон силы
Чемсильней
раздражение,
тем
до
определенного предела сильнее ответная
реакция биосистемы.
Закон
постулирован
для
целостного
биологического объекта.
Например, нервного ствола, состоящего из
отдельных аксонов; скелетной мышцы и т.д.
Связано это с тем, что каждое отдельное волокно
отвечает на раздражитель по типу «все или ничего»,
но порог возбуждения у них отличается.
Поэтому суммарная активность объекта будет
находится в градуальной зависимости от силы
раздражителя.
12. 12 Закон времени
Чем длительнее раздражение, тем сильней доопределенных пределов ответная реакция живой
ткани.
В определенном диапозоне эта зависимость имеет
характер гиперболы. На ней выделяют реобазу,
полезное время и хроноксию.
Реобаза - соответствует порогу возбуждения.
Полезное время – минимальное время, в течение
которого раздражитель, равный одной реабазе,
должен действовать на ткань, чтобы вызвать
возбуждение.
Хроноксия – время, в течение которого должен
действовать раздражитель, равный двум реабазам,
чтобы вызвать возбуждение.
13. 13 ЗАКОН «СИЛА - ДЛИТЕЛЬНОСТЬ»
Этот закон былэкспериментал
ьно
установллен и
сформулирован
независимо
друг от друга
тремя
учеными:
Лапик,
Гоорвиг, Вейс.
14. 14 Закон крутизны нарастания раздражителя или градиента:
Чемвыше
крутизна
нарастания
раздражителя во времени, тем больше до
известного
предела
величина
функционального ответа.
В основе этого закона лежат физикохимические и функциональные изменения,
вызываемые в раздражаемом объекте.
Возбуждение развивается в том случае, если
активационные процессы достигают
пороговой критической величины раньше
инактивационных.
15. 15
ЗАКОН ГРАДИЕНТА РАЗДРАЖЕНИЯ(АККОМОДАЦИЯ)
16.
История открытиябиоэлектри чества.
17.
18. 18 Первый опыт Гальвани
При подвешивании нервно - мышечного препаратас помощью медного крючка, проходящего через
спинной мозг препарата, имело место сокращение
мышц лапки каждый раз, когда эта лапка
соприкасалась с железным поручнем балкона.
19.
19ВОЛЬТОВ
СТОЛБ
Однако
ему
возразил
его
знаменитый соотечественник физик А. Вольта, который считал,
что в «балконном» опыте мышца
является лишь чувствительным
«электрометром»
электричества,
порождаемого
контактной
разностью потенциалов разных
металлов, используемых в опытах
Гальвани.
Вольтов столб, состоящий из металлических
дисков, разделенных кружками мокрой
ткани.
20. 20 Второй опыт Гальвани
Позднее,защищая
свою
точку зрения от
возражений
оппонентов,
Гальвани
воспроизводит
различные
модификации
опытов,
в
которых
сокращение
мышцы
вызывалось
путем
набрасывания
нерва с помощью
стеклянной
палочки
на
поврежденный и
неповрежденный
участок мышцы.
21. 21 Опыт Маттеучи
• Открытия Гальвани былиподтверждены в работах
Маттеучи (1837).
Однако,
Маттеучи
обнаружил
явление
вторичного
или
индуцированного
сокращения:
при
помещении нерва одного
нервно-мышечного
препарата на мышцу другого
препарата и раздражителя
нерва
этого
препарата,
Маттеучи
наблюдал
сокращение мышцы обоих
препаратов.
22. 22 Работы Э.Дюбуа-Реймона
( выполненны в 1840-1860 гг. )В
этих
работах
благодаря
высокочувствительному гальванометру и ряду
других технических новшеств удалось впервые
определить электрические процессы в мышце,
зарегистрировав
потенциал
наружной
и
внутренней поверхности мембраны клеток.
Он впервые установил, что наружная
мембрана
заряжена
положительно
по
отношению к внутренней, и эта разность
потенциалов
изменяется
при
сокращении
мышцы.
23.
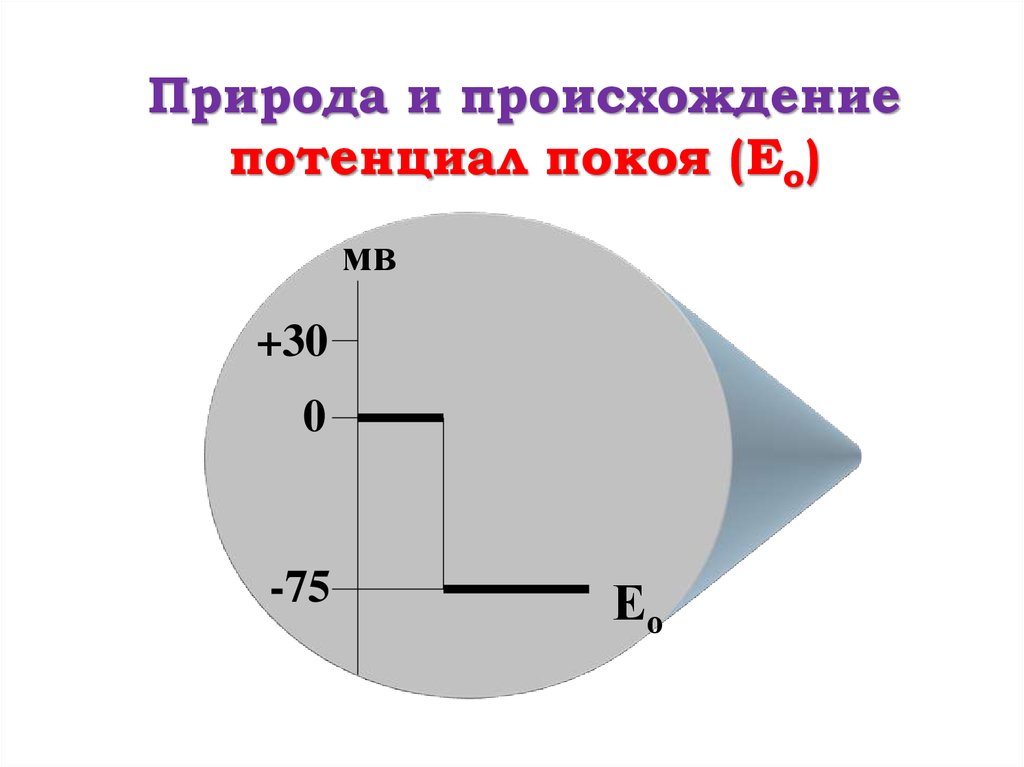
Природа и происхождениепотенциал покоя (Ео)
мв
+30
0
-75
Ео
24. 24
В 1896 г. В.Ю. Чаговец впервые высказалгипотезу о ионном механизме электрических
потенциалов в живых клетках. Он сделал попытку
применить теорию электролитической
диссоциации Аррениуса - для их объяснения.
В 1902 г. Бернштейном была развита
мембранно-ионная теория, согласно которой
клеточная поверхность представляет собой
полупроницаемую мембрану, которая в
состоянии физиологического покоя
проницаема для ионов калия и практически
непроницаема для остальных вне - и
внутриклеточных ионов.
25. 25 Диффузионный концентрационный потенциал
Представим сосуд, разделенный искусственной полупроницаемой мембраной.Стенки пор этой мембраны заряжены
электроотрицательно, поэтому они
пропускают только катионы и непроницаемы
для анионов. В обе половины сосуда налит
солевой раствор, содержащий ионы К+,
однако их концентрация в правой части
сосуда выше, чем в левой. Вследствие этого
концентрационного градиента ионы К+
начинают диффундировать из правой
половины сосуда в левую, принося туда свой
положительный заряд. Это приводит к тому,
что непроникающие анионы начинают
скапливаться у мембраны в правой половине
сосуда. Своим отрицательным зарядом они
электростатически будут удерживать К+ у
поверхности мембраны в левой половине
сосуда. В результате мембрана поляризуется,
и между двумя ее поверхностями создается
разность потенциалов, соответствующая
равновесному калиевому потенциалу (Ек).
26. 26 Двойной электрический слой (мембрана поляризована)
27. 27 Уравнение НЕРНСТА
[ K +o]RT
Ек =
ℓn
nF
[ K +i]
Ек – равновесный калиевый потенциал;
R - универсальная газовая постоянная;
T - температура среды;
n - валентность иона ( для К+ = 1);
F - число Фарадея;
К+ - концентрация калия с внешней (о) и внутренней
(i) стороны мембраны.
28. 28 Английский зоолог Д. Юнг (1936) обнаружил, что особые длинные тяжи у кальмаров и каракатиц являются не кровеносными сосудами, как считалось
28Английский зоолог
Д. Юнг (1936)
обнаружил, что особые
длинные тяжи у
кальмаров и
каракатиц являются
не кровеносными
сосудами, как
считалось ранее, а
необычайно толстыми
аксонами.
29. 29 Экспериментальная проверка гипотезы Бернштейна
В 1962 г. Ходжкин с сотрудниками подтвердил гипотезуБернштейна:
Из аксона кальмара выдавили аксоплазму и спавшуюся
оболочку заполнили искусственным солевым раствором.
Когда концентрация калия была близка к внутриклеточной, между внутренней и наружной мембраной установилась
разность потенциалов близкая к ПП (-50/-80 мВ), и волокно
проводило импульсы.
При снижении концентрации калия или увеличении –
мембранный потенциал менялся, или даже изменял свой знак
(потенциал становился положительным, если в наружном
растворе [ K +o] > [ K +i] ).
Это доказывало, что концентрационный градиент калия
является основным фактором, определяющим величину ПП
нервного волокна.
30. 30
Современные представления о природебиоэлектрических явлений в тканях базируются на
результатах работ Алана Ходжкина, Эндрью Хаксли,
Бернарда Катца.
Эти ученые в 40-50 годах нашего века
модифицировали и экспериментально обосновали
мембранно-ионную теорию Бернштейна.
В настоящее время их взгляды о природе
биоэлектрических явлений пользуются всеобщим
признанием.
Согласно их представлениям, наличие
электрических потенциалов в живых клетках
обусловлено различной концентрацией ионов Na+, K+,
Ca2+ и Cl- внутри и вне клетки, а также различной
проницаемостью для них клеточной мембраны.
31.
32. 32 Мембранно-ионная теория происхождения потенциала покоя (ПП)
Все клетки живого организма поляризованы. Между внутреннейсредой клетки и внешней средой существует разность
потенциалов.
Авторы современной теории: Ходжкин, Катц, Хаксли 1949-1952 гг.
Положения теории:
I. Ионы вне и внутри клетки распределены
неравномерно.
Катионов калия внутри клетки больше в 40-50 раз,
натрия меньше в 8-12 раз,
анионов хлора меньше в 30-40 раз.
II. Проницаемость мембраны для разных ионов
различна.
В состоянии покоя:
Р калия : Р натрия : Р хлора = 1 : 0,04 : 0, 45
33. 33 Схема регистрации мембранного потенциала клетки
34. 34
35. 35 Концентрации Na+, K+ и Cl– внутри и вне клетки и пути проникновения этих ионов через клеточную мембрану : через специфические ионные каналы и
35Концентрации Na+, K+ и Cl– внутри и вне клетки и пути
проникновения этих ионов через клеточную мембрану :
через специфические ионные каналы или с помощью
Na/K–насоса.
При данных концентрационных градиентах
равновесные потенциалы ENa, EK и EСl– равны
указанным, мембранный потенциал Em = –90 мВ
36. 36 Физиологический мембранный потенциал
в возбудимых клетках называетсяпотенциалом покоя (ПП)
37. 37
38. 38 Если подставить значения концентраций из таблицы, то:
ЕK+ = - 97 мВЕNa+ = + 66 мВ
39. 39 Уравнение ГОЛЬДМАНА
Уравнение ГОЛЬДМАНА39
RT
Ем
ℓn
=
nF
Pk [ Ko+] + PNa [ Nao+]+PCl [ Cli- ]
Pk[ Ki+]+ PNa [ Nai+] + PCl[ Clo–]
Ем – равновесный калиевый потенциал;
R - универсальная газовая постоянная;
T - температура среды;
n - валентность иона;
F - число Фарадея;
P – проницаемость иона для мембраны;
К+ - концентрация калия с внешней (о)
и внутренней
(i)
стороны мембраны.
Na+ - концентрация натрия с внешней (о) и внутренней (i)
стороны мембраны.
Cl- - концентрация хлора с внешней (о) и внутренней (i) стороны
мембраны.
40. 40 Потенциал покоя создается:
Неравновесным распределениемкалия, натрия, хлора и кальция
внутри клетки по отношению к
внешней среде.
Разной проницаемостью мембраны
для этих ионов.
Основным потенциалобразующим
ионом является катион калия.
Постоянство потенциала покоя
поддерживается работой ионных
насосов.
Электрогенность калий-натриевого
насоса вносит дополнительный вклад
в создание мембранного потенциала.
41.
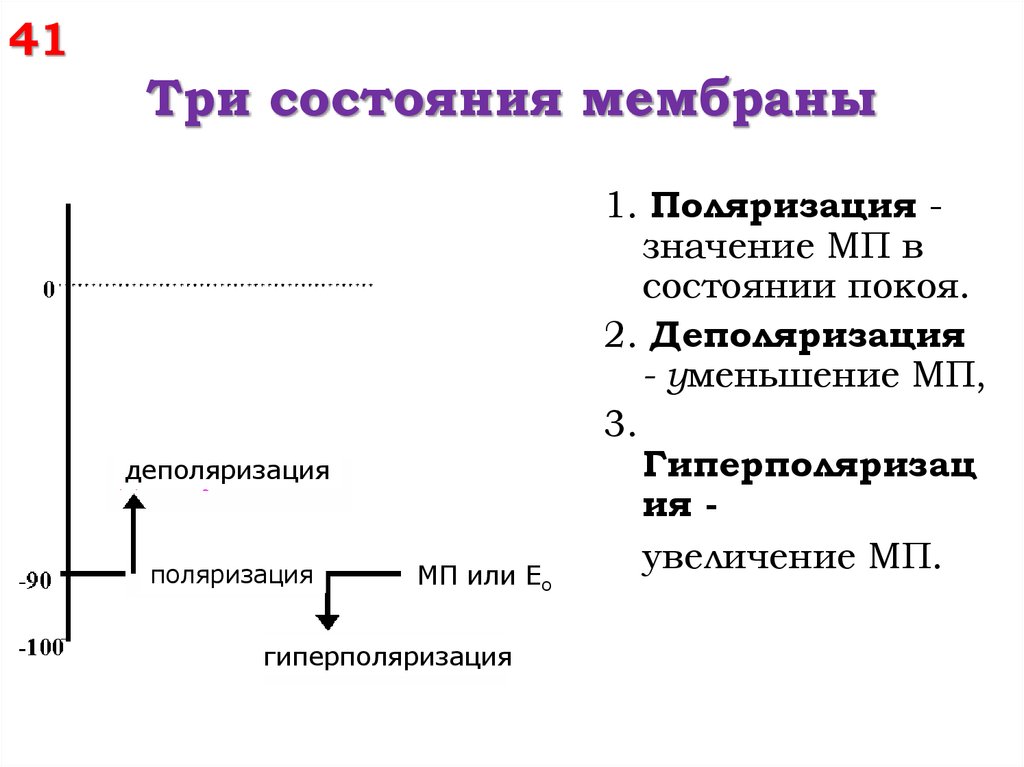
41Три состояния мембраны
деполяризация
поляризация
МП или Ео
гиперполяризация
1. Поляризация значение МП в
состоянии покоя.
2. Деполяризация
- уменьшение МП,
3.
Гиперполяризац
ия увеличение МП.
42. 42 Два варианта ответной реакции возбудимой структуры на действие раздражителя:
1. локальный ответ (ЛО)- надействие подпорогового
раздражителя,
2. потенциал действия (ПД,
импульс) - на действие порогового и
надпорогового раздражителя
43. Потенциал действия
44.
44Мембранно-ионная теория
происхождения
потенциала действия (ПД)
ПД – это быстрое изменение мембранного
потенциала, которые возникают в месте
нанесения раздражения и распространяются
на всю мембрану клетки.
В основе возбуждения лежит
изменение проницаемости мембраны для
натрия и калия, связанное с активацией
потенциалзависимых натриевых и калиевых
каналов.
45. 45
46. 46 Натриевая гипотеза происхождения ПД
При действии на клетку раздражителя проницаемостьмембраны для ионов Na+ резко повышается за счет
активации (открывания) натриевых каналов.
При этом ионы Na+ по концентрационному градиенту
интенсивно перемещаются из вне - во внутриклеточное
пространство. Вхождению ионов Na+ в клетку способствует и
электростатический
градиент.
В
итоге
проницаемость
мембраны для Na+ становится в 20 раз больше
проницаемости для ионов К+.
Поскольку поток Na+ в клетку начинает превышать калиевый
ток из клетки, то происходит постепенное снижение потенциала
покоя, приводящее к реверсии - изменению знака
мембранного потенциала. При этом внутренняя поверхность
мембраны становится положительной по отношению к ее
внешней поверхности. Указанные изменения мембранного
потенциала соответствуют восходящей фазе потенциала
действия - фазе деполяризации.
47. 47 Экспериментальное доказательство натриевой природы деполяризации
В 1939 г. Ходжкин и Хаксли установили, что мембранныйпотенциал во время развития ПД не просто уменьшается до нуля, а
меняет свой знак на противоположный. Эти данные противоречили
гипотезе Бернштейна о том, что во время ПД мембрана становиться
проницаемой для всех ионов.
Важным аргументом в пользу натриевой природы деполяризации
ПД стал факт тесной зависимости амплитуды ПД от [ Nao+]. В опытах с
перфузией
аксонов
кальмара
изнутри
солевыми
растворами
установлено:
При замене аксоплазмы солевым раствором, в котором К+ частично
заменен на Na+, - амплитуда ПД резко снижается.
При полной замене К+ на Na+ волокно
способность генерировать ПД.
полностью утрачивает
При замене внеклеточного Na+ на холин хлорид нервное волокно
также полностью утрачивает способность генерировать ПД.
Все это позволило сделать вывод о зависимости величины ПД
от градиента натрия.
48. 48
Мембранахарактеризуется
повышенной
проницаемостью для ионов Na+ лишь очень короткое
время 0.2 - 0.5 мс.
После этого проницаемость мембраны для ионов Na+
вновь понижается, а для К+ возрастает. В результате поток
Na+ внутрь клетки резко ослабляется, а ток К+ из клетки
усиливается.
В течение потенциала действия в клетку поступает
значительное количество Na+, а ионы К+ покидают клетку.
Восстановление
клеточного
ионного
баланса
осуществляется благодаря работе Na+/К+-АТФ-азы.
Итогом этих процессов и является реполяризация
мембраны: внутреннее содержимое клетки вновь
приобретает отрицательный заряд по отношению к
внешней поверхности мембраны.
49.
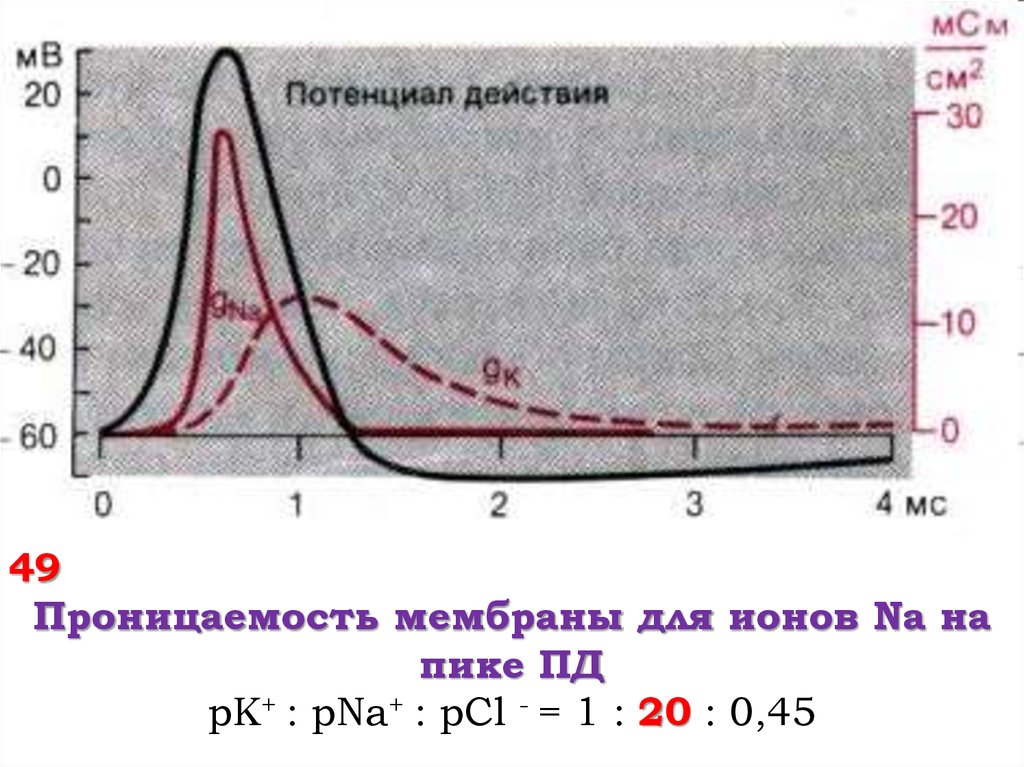
49Проницаемость мембраны для ионов Na на
пике ПД
pK+ : pNa+ : pCl - = 1 : 20 : 0,45
50. 50
51. 51 ЦИКЛ ХОДЖКИНА-ХАКСЛИ
51ЦИКЛ
Раздражитель
ХОДЖКИНА-ХАКСЛИ
деполяризация мембраны
возрастание
входящего
Na+ - тока
повышение Na+
проницаемости
52. 52
СОСТОЯНИЕ НАТРИЕВЫХ КАНАЛОВNa+
m
h
СОСТОЯНИЕ
ПОТЕНЦИАЛА
ПОКОЯ
m
m
h
h
СОСТОЯНИЕ
ДЕПОЛЯРИЗАЦИИ
МЕМБРАНЫ
Р Е А К Т И В А Ц И Я Na-канала
РЕПОЛЯРИЗАЦИЯ
МЕМБРАНЫ
53. 53
54.
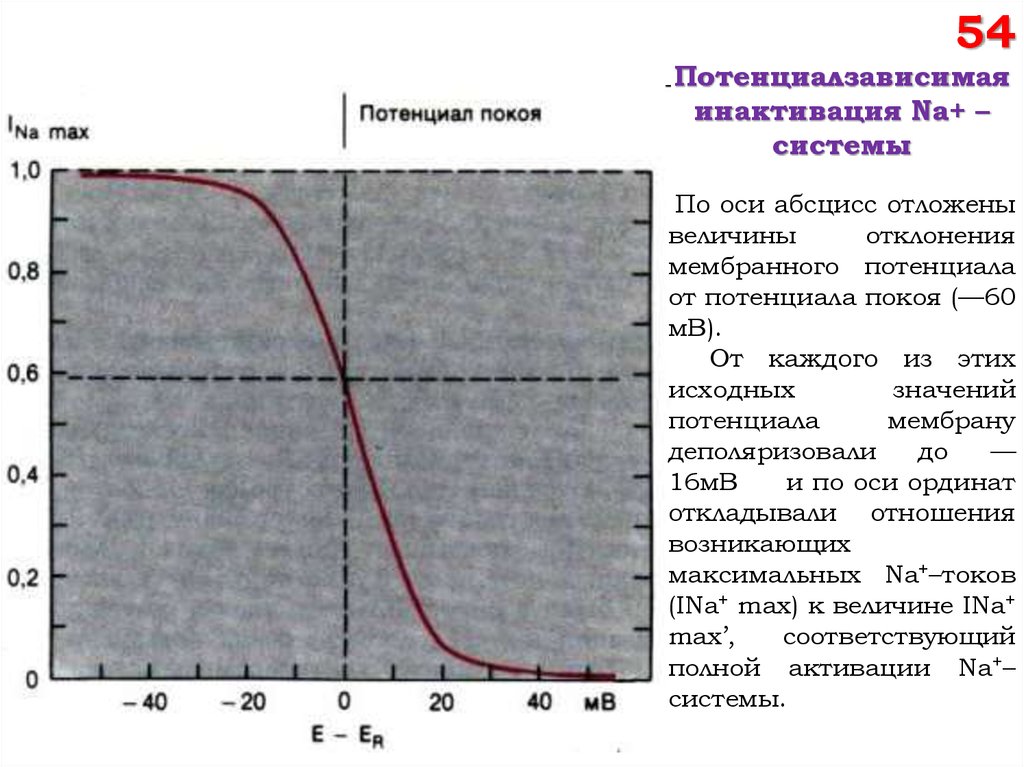
54Потенциалзависимая
инактивация Na+ –
системы
По оси абсцисс отложены
величины
отклонения
мембранного потенциала
от потенциала покоя (—60
мВ).
От каждого из этих
исходных
значений
потенциала
мембрану
деполяризовали
до
—
16мВ
и по оси ординат
откладывали отношения
возникающих
максимальных Na+–токов
(INa+ max) к величине INa+
max’,
соответствующий
полной активации Na+–
системы.
55. 55
Nа+ - потенциалозависимые каналы, вмембране нервного и скелетного мышечного
волокна, характеризуются, как время - так
и потенциалозависимостью.
Активируются на уровне ~ -60 мВ,
полностью
инактивируются
уже
при
деполяризации мембраны до ~ -40 мВ.
Их работа подчиняется закону цикла
Ходжкина
Хаксли
и
обеспечивает
кратковременный, но мощный быстрый
входящий Nа- ток.
56. 56
СОСТОЯНИЕ КАЛИЕВЫХ КАНАЛОВК+
m
К+
К+
m
m
СОСТОЯНИЕ
ПОТЕНЦИАЛА
ПОКОЯ
СПАЙК
СОСТОЯНИЕ
РЕПОЛЯРИЗАЦИИ
МЕМБРАНЫ
Блокада калиевых каналов тетраэтиламмонием
резко удлиняет процесс реполяризации
57. 57
58. 58
59. 59 Потенциал действия:
1.2.
3.
4.
5.
Возникает в ответ на пороговые и сверхпороговые
раздражители.
Обусловлен увеличением проницаемости мембраны
для натрия и калия. При этом
деполяризация
определяется натриевым током, реполяризация калиевым.
Вслед за основными фазами потенциала действия
возникают
следовые
потенциалы.
Следовой
отрицательный
связан
с
повышенной
проводимостью
мембраны
для
натрия,
положительный - для калия.
Потенциал
действия
распространяется
и
подчиняется правилу “Все или ничего”.
При возбуждении активируется калий-натриевый
насос.
60. ТРИ СОСТОЯНИЯ МЕМБРАНЫ
0Деполяризация
-90
-100
Поляризация
МПП или Е0
Гиперполяризация