


















 biology
biologySimilar presentations:




")





Физиология мышечного сокращения
1.
Мы учим и лечимс 1888 года
Физиология мышечного сокращения
Самойлова Анна
Викторовна
Доцент кафедры
нормальной физиологии,
канд. биол. наук
2.
Сократительные клеткиСпособность к сокращению (сократимость) — это
свойство, присущее прежде всего мышечным клеткам
• Мышечные волокна
• Перициты
• Миоэпителиальные клетки экзокринных желез
• Миофибробласты
• Эндотелиальные клетки
• Наружные волосковые клетки внутреннего уха
3.
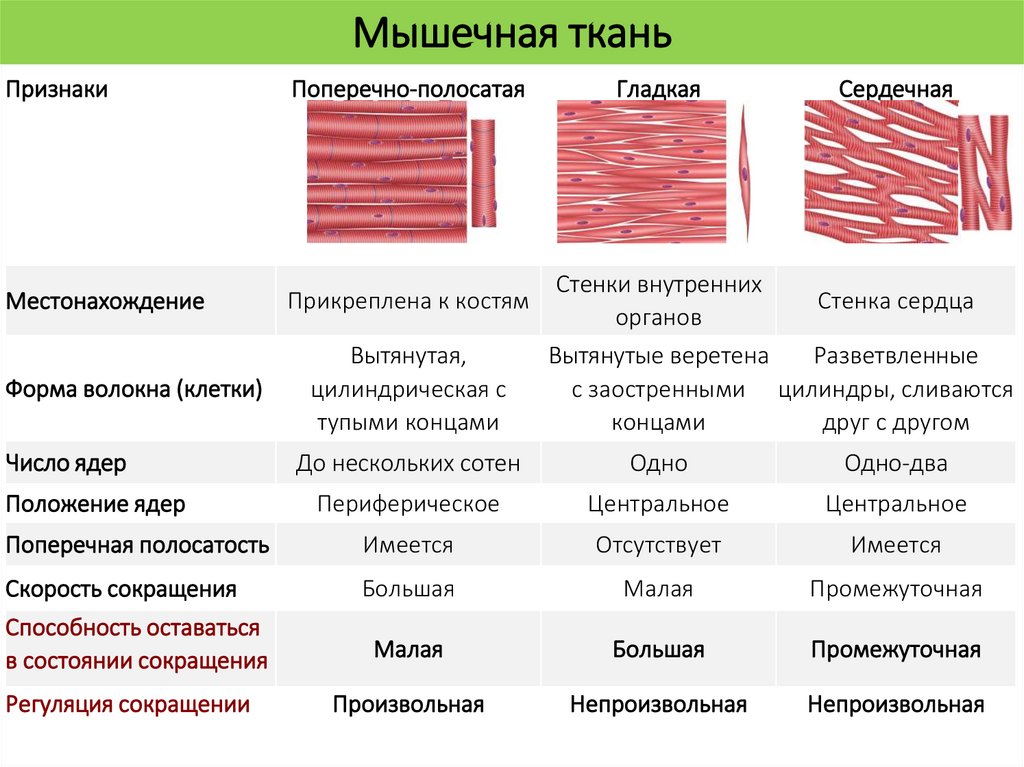
Мышечная тканьПризнаки
Поперечно-полосатая
Местонахождение
Прикреплена к костям
Гладкая
Сердечная
Стенки внутренних
Стенка сердца
органов
Вытянутые веретена
Разветвленные
с заостренными цилиндры, сливаются
концами
друг с другом
Форма волокна (клетки)
Вытянутая,
цилиндрическая с
тупыми концами
Число ядер
До нескольких сотен
Одно
Одно-два
Периферическое
Центральное
Центральное
Поперечная полосатость
Имеется
Отсутствует
Имеется
Скорость сокращения
Большая
Малая
Промежуточная
Способность оставаться
в состоянии сокращения
Малая
Большая
Промежуточная
Регуляция сокращении
Произвольная
Непроизвольная
Непроизвольная
Положение ядер
4.
Свойства скелетных мышц:Физиологические:
1. Возбудимость (ниже
чем у нерва); ПП = -90
мВ
2. Проводимость (низкая
(10–13 м/с);
3. Рефрактерность
(длиннее чем у
нервного волокна);
4. Лабильность (низкая).
Специфические
(физические):
1. Сократимость
(способность
укорачиваться или
развивать напряжение).
2. Эластичность
(способность развивать
напряжение при
растягивании).
5.
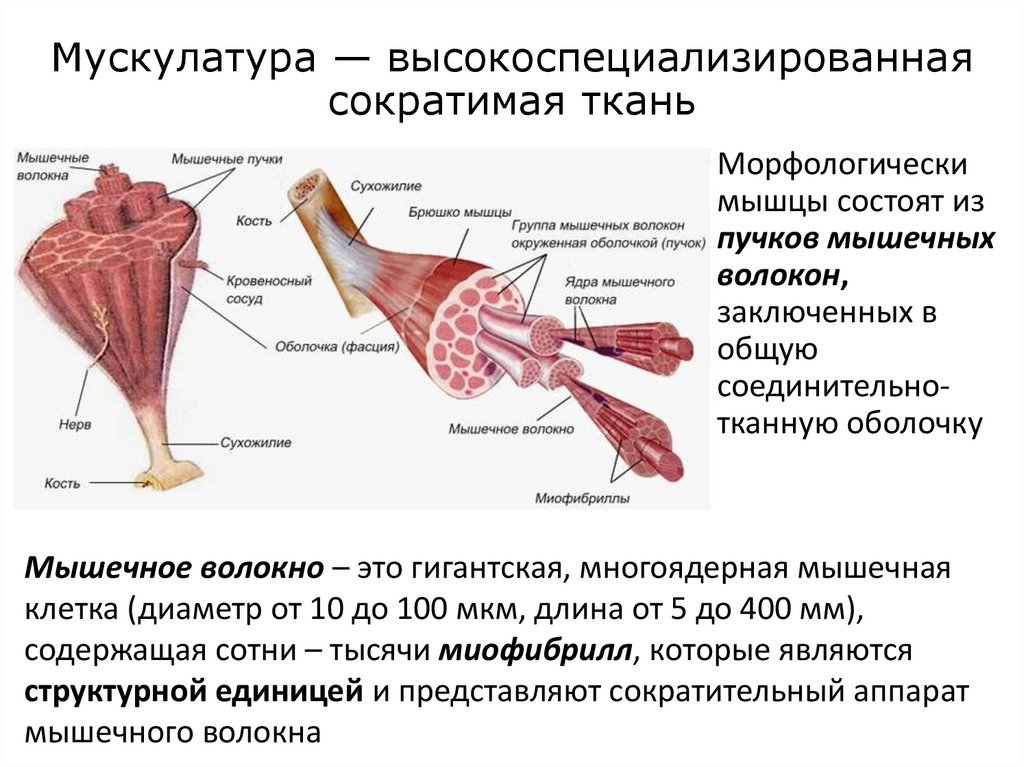
Мускулатура — высокоспециализированнаясократимая ткань
Морфологически
мышцы состоят из
пучков мышечных
волокон,
заключенных в
общую
соединительнотканную оболочку
Мышечное волокно – это гигантская, многоядерная мышечная
клетка (диаметр от 10 до 100 мкм, длина от 5 до 400 мм),
содержащая сотни – тысячи миофибрилл, которые являются
структурной единицей и представляют сократительный аппарат
мышечного волокна
6.
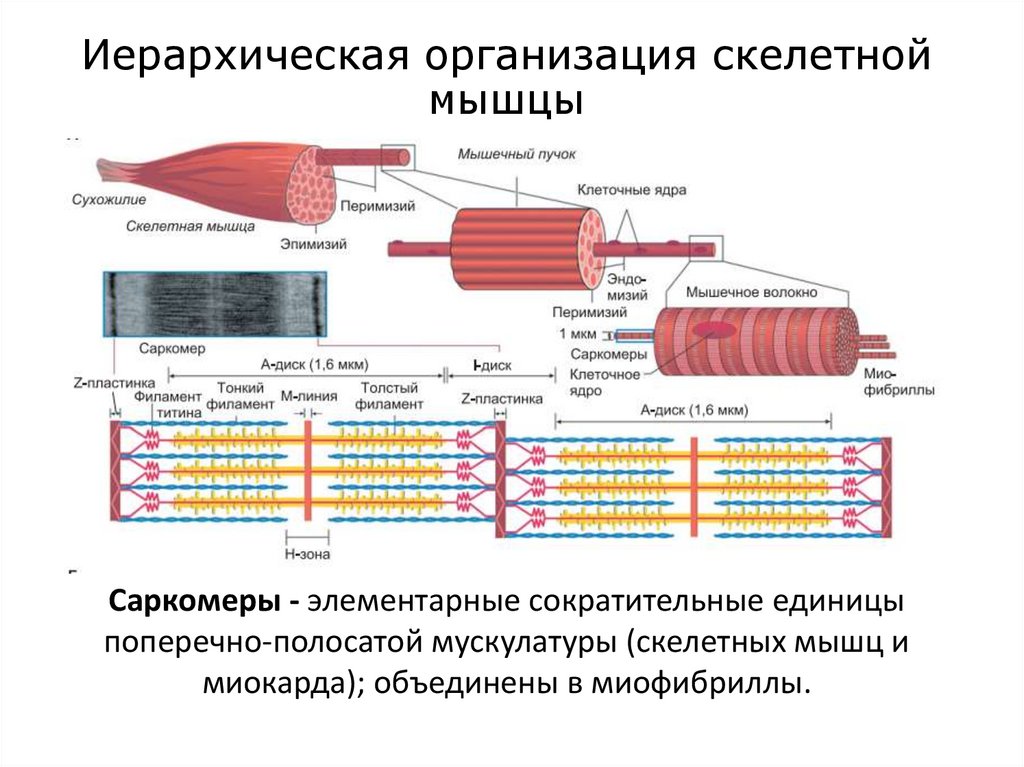
Иерархическая организация скелетноймышцы
Саркомеры - элементарные сократительные единицы
поперечно-полосатой мускулатуры (скелетных мышц и
миокарда); объединены в миофибриллы.
7.
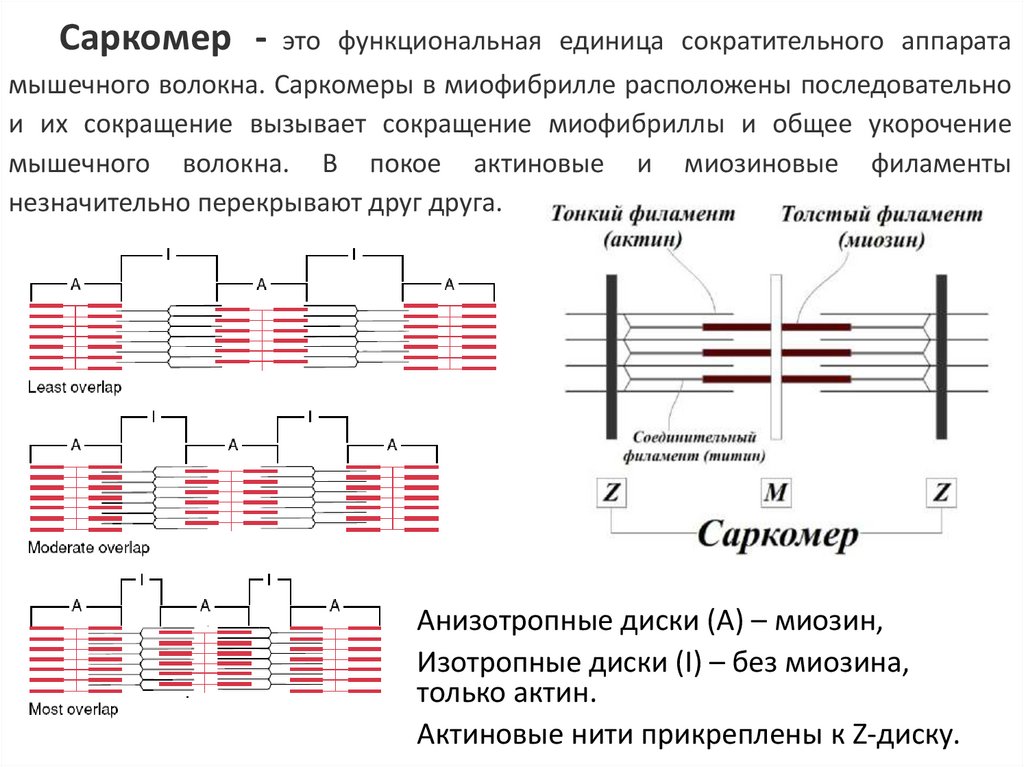
Саркомер - это функциональная единица сократительного аппаратамышечного волокна. Саркомеры в миофибрилле расположены последовательно
и их сокращение вызывает сокращение миофибриллы и общее укорочение
мышечного волокна. В покое актиновые и миозиновые филаменты
незначительно перекрывают друг друга.
Анизотропные диски (А) – миозин,
Изотропные диски (I) – без миозина,
только актин.
Актиновые нити прикреплены к Z-диску.
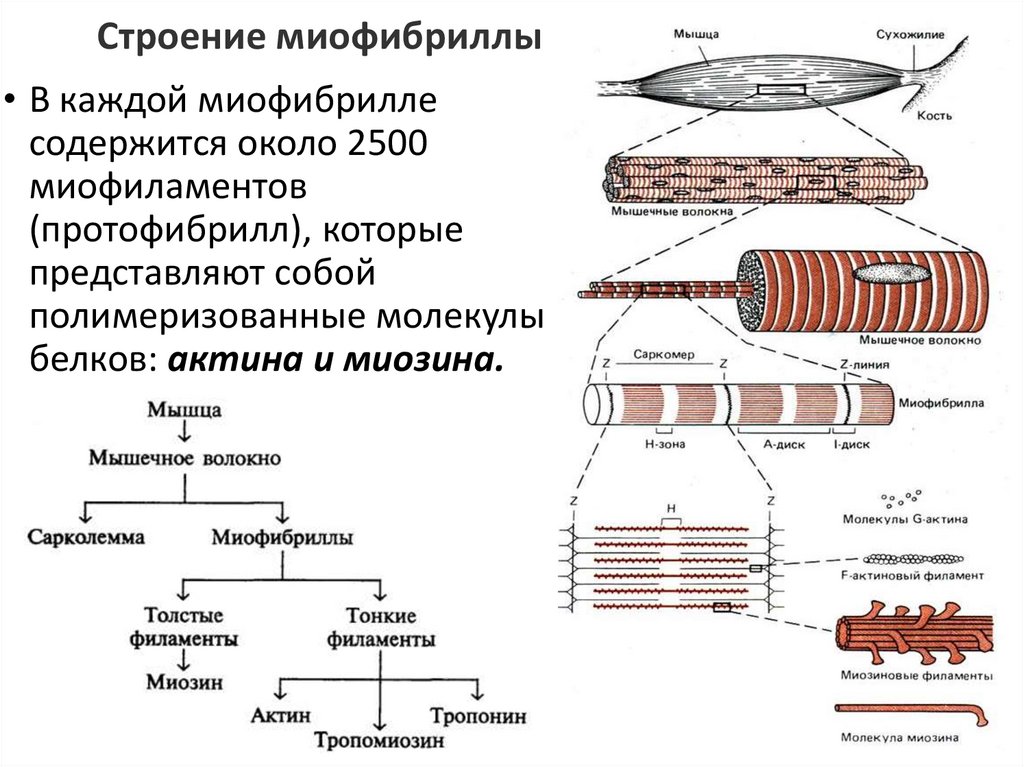
8.
Строение миофибриллы• В каждой миофибрилле
содержится около 2500
миофиламентов
(протофибрилл), которые
представляют собой
полимеризованные молекулы
белков: актина и миозина.
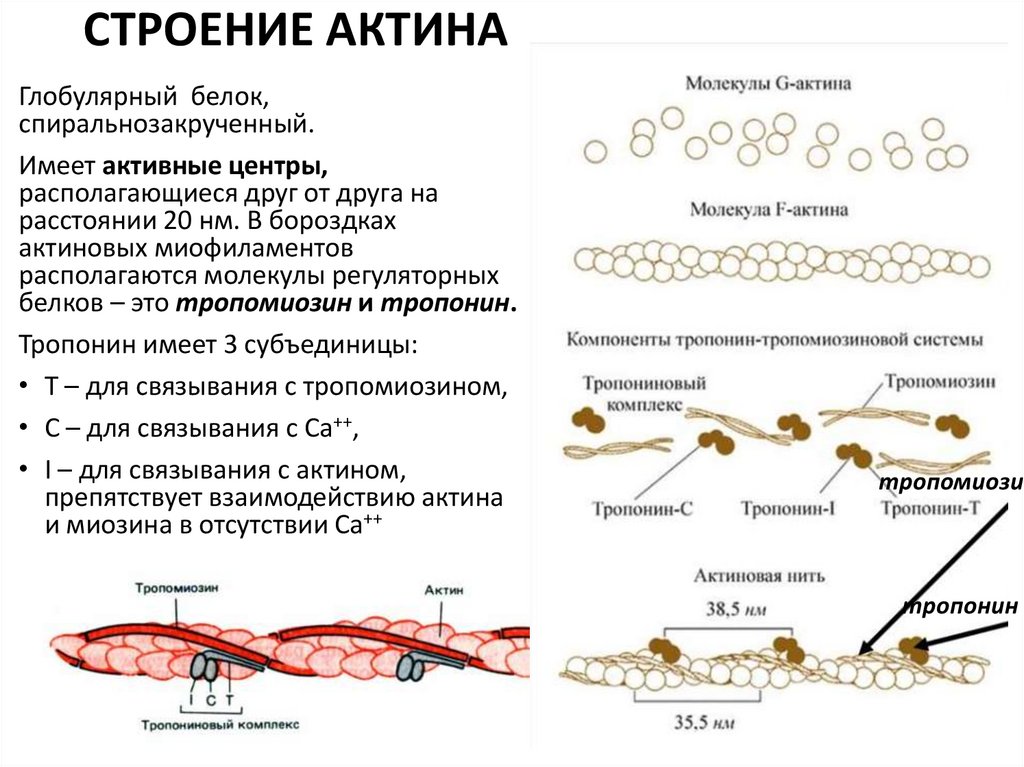
9.
СТРОЕНИЕ АКТИНАГлобулярный белок,
спиральнозакрученный.
Имеет активные центры,
располагающиеся друг от друга на
расстоянии 20 нм. В бороздках
актиновых миофиламентов
располагаются молекулы регуляторных
белков – это тропомиозин и тропонин.
Тропонин имеет 3 субъединицы:
• Т – для связывания с тропомиозином,
• С – для связывания с Са++,
• I – для связывания с актином,
препятствует взаимодействию актина
и миозина в отсутствии Са++
тропомиозин
тропонин
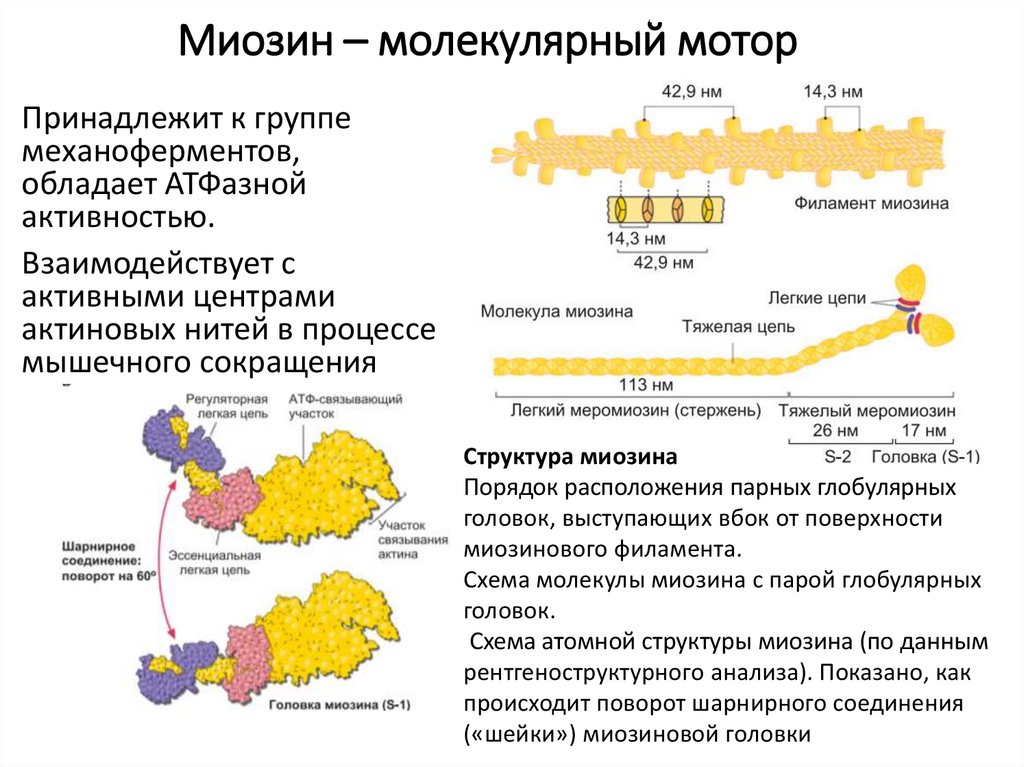
10.
Миозин – молекулярный моторПринадлежит к группе
механоферментов,
обладает АТФазной
активностью.
Взаимодействует с
активными центрами
актиновых нитей в процессе
мышечного сокращения
Структура миозина
Порядок расположения парных глобулярных
головок, выступающих вбок от поверхности
миозинового филамента.
Схема молекулы миозина с парой глобулярных
головок.
Схема атомной структуры миозина (по данным
рентгеноструктурного анализа). Показано, как
происходит поворот шарнирного соединения
(«шейки») миозиновой головки
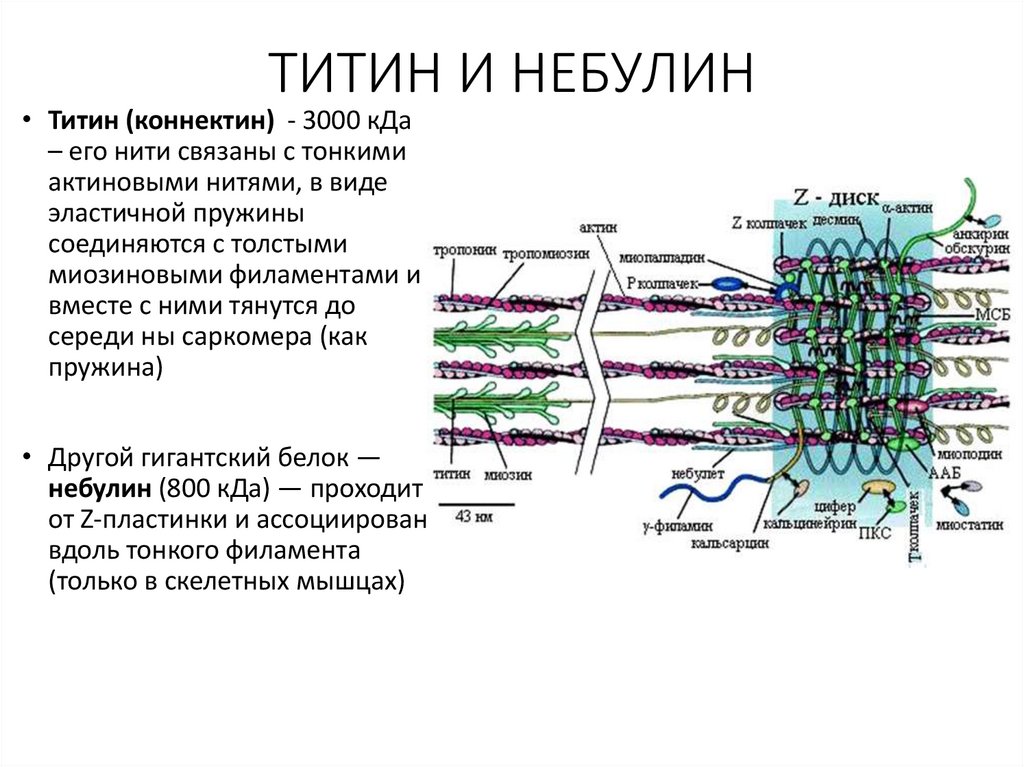
11.
ТИТИН И НЕБУЛИН• Титин (коннектин) - 3000 кДа
– его нити связаны с тонкими
актиновыми нитями, в виде
эластичной пружины
соединяются с толстыми
миозиновыми филаментами и
вместе с ними тянутся до
середи ны саркомера (как
пружина)
• Другой гигантский белок —
небулин (800 кДа) — проходит
от Z-пластинки и ассоциирован
вдоль тонкого филамента
(только в скелетных мышцах)
12.
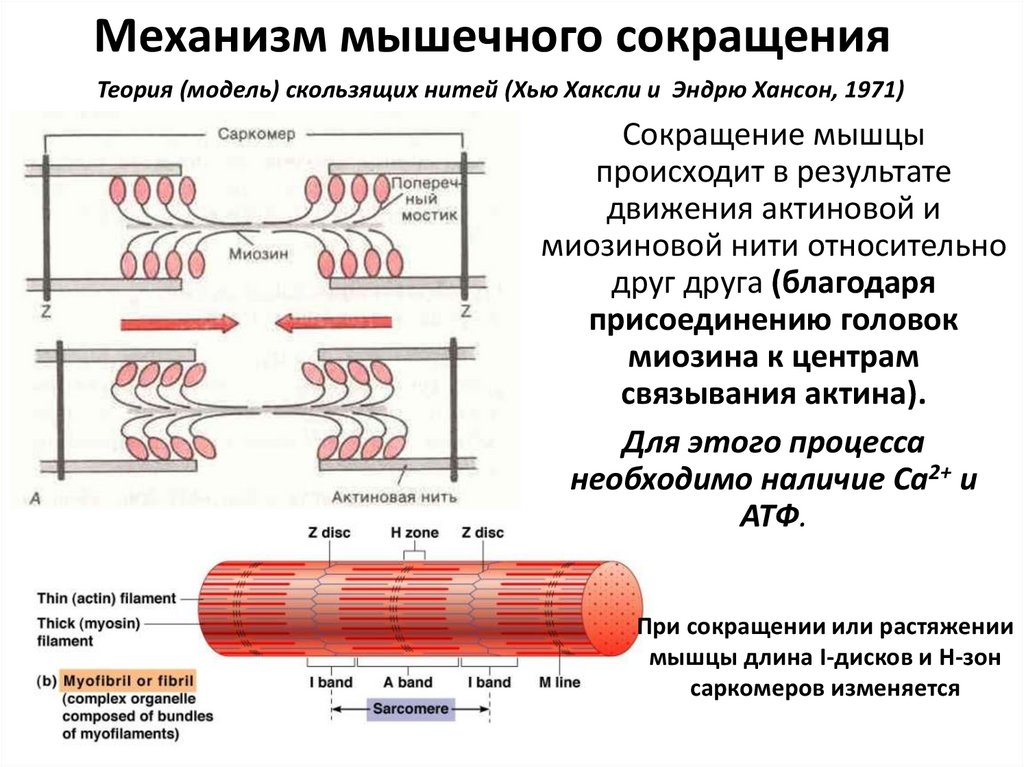
Механизм мышечного сокращенияТеория (модель) скользящих нитей (Хью Хаксли и Эндрю Хансон, 1971)
Сокращение мышцы
происходит в результате
движения актиновой и
миозиновой нити относительно
друг друга (благодаря
присоединению головок
миозина к центрам
связывания актина).
Для этого процесса
необходимо наличие Са2+ и
АТФ.
При сокращении или растяжении
мышцы длина I-дисков и Н-зон
саркомеров изменяется
13.
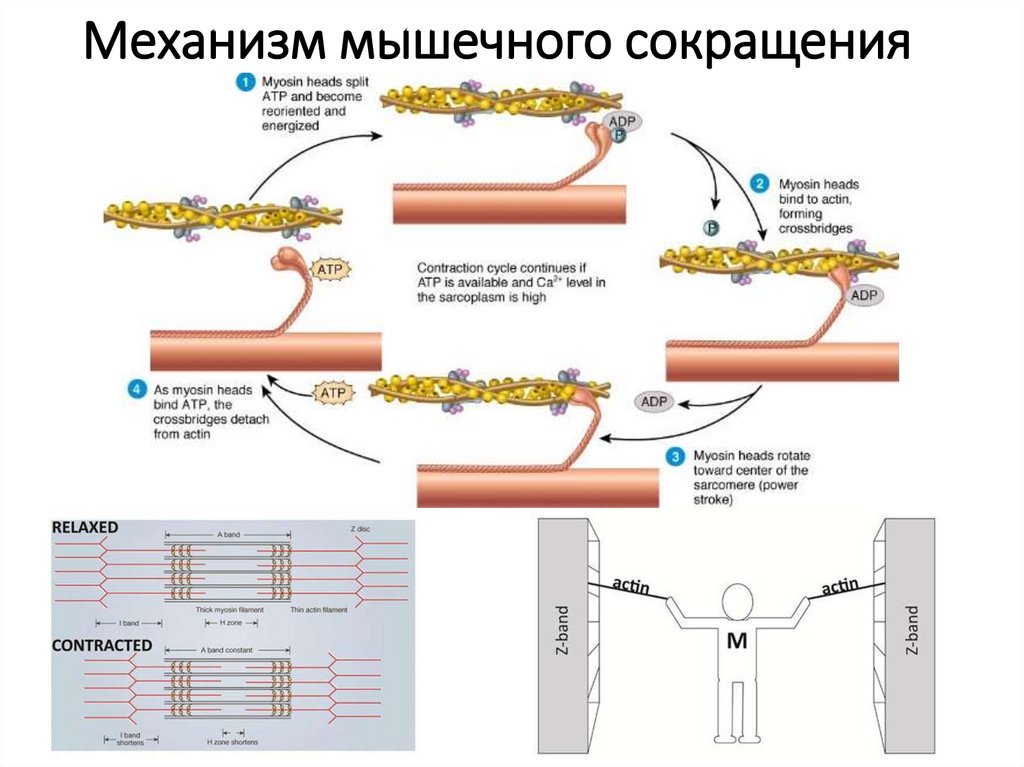
Механизм мышечного сокращения14.
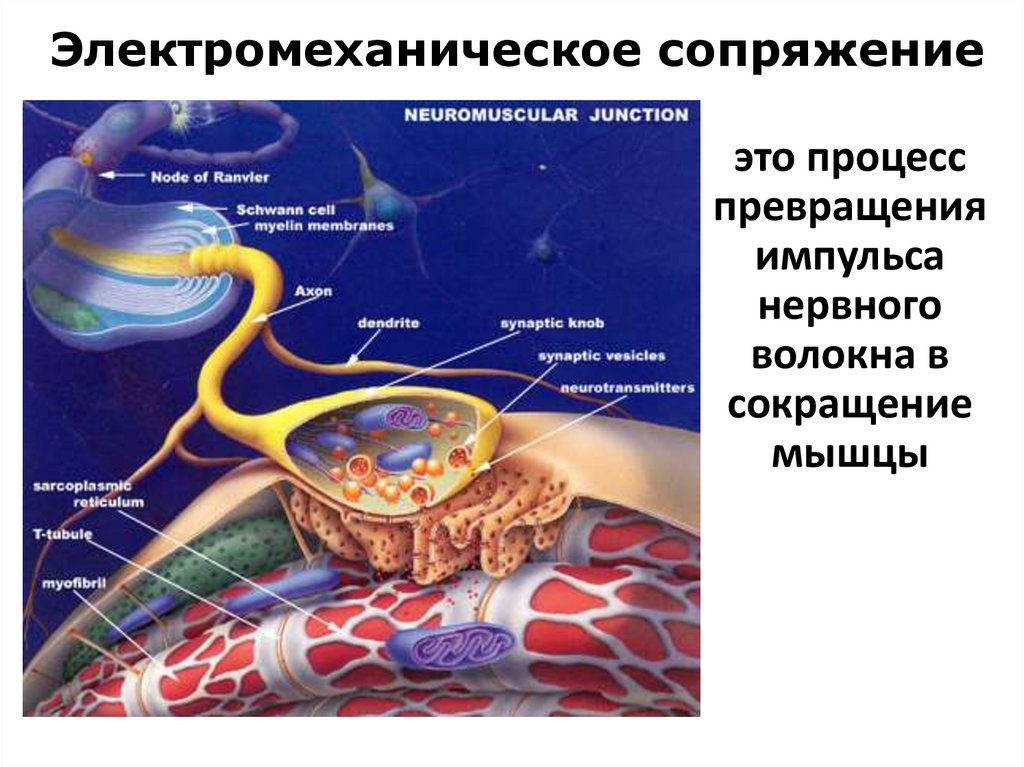
Электромеханическое сопряжениеэто процесс
превращения
импульса
нервного
волокна в
сокращение
мышцы
15.
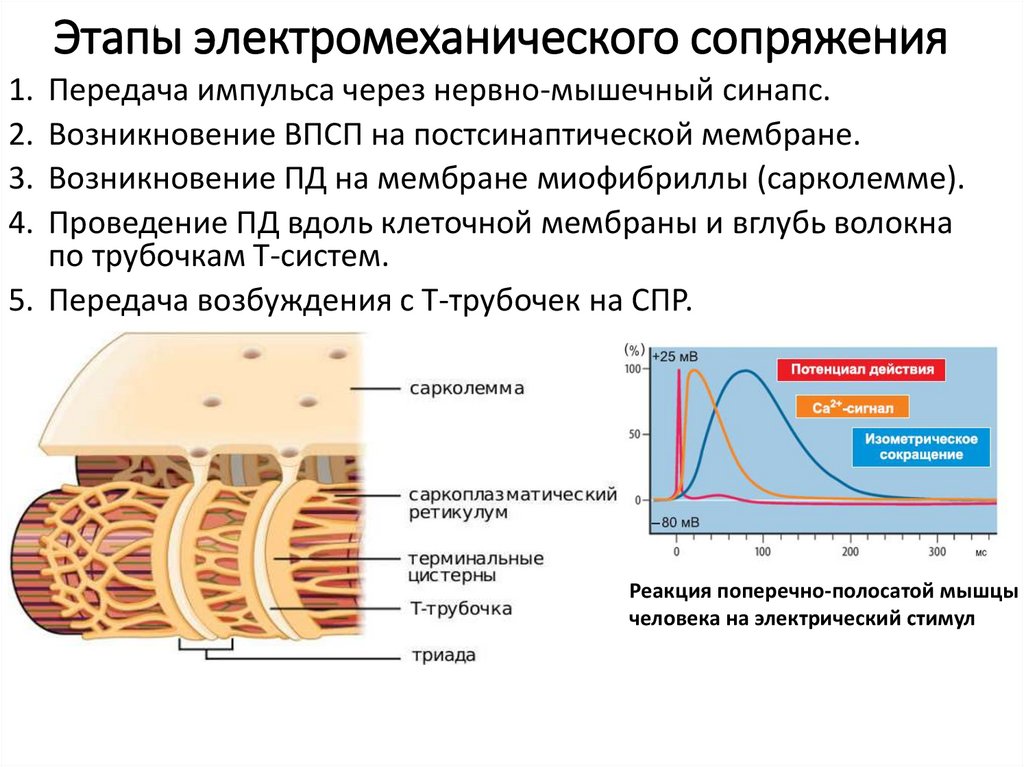
Этапы электромеханического сопряжения1. Передача импульса через нервно-мышечный синапс.
2. Возникновение ВПСП на постсинаптической мембране.
3. Возникновение ПД на мембране миофибриллы (сарколемме).
4. Проведение ПД вдоль клеточной мембраны и вглубь волокна
по трубочкам Т-систем.
5. Передача возбуждения с Т-трубочек на СПР.
Реакция поперечно-полосатой мышцы
человека на электрический стимул
16.
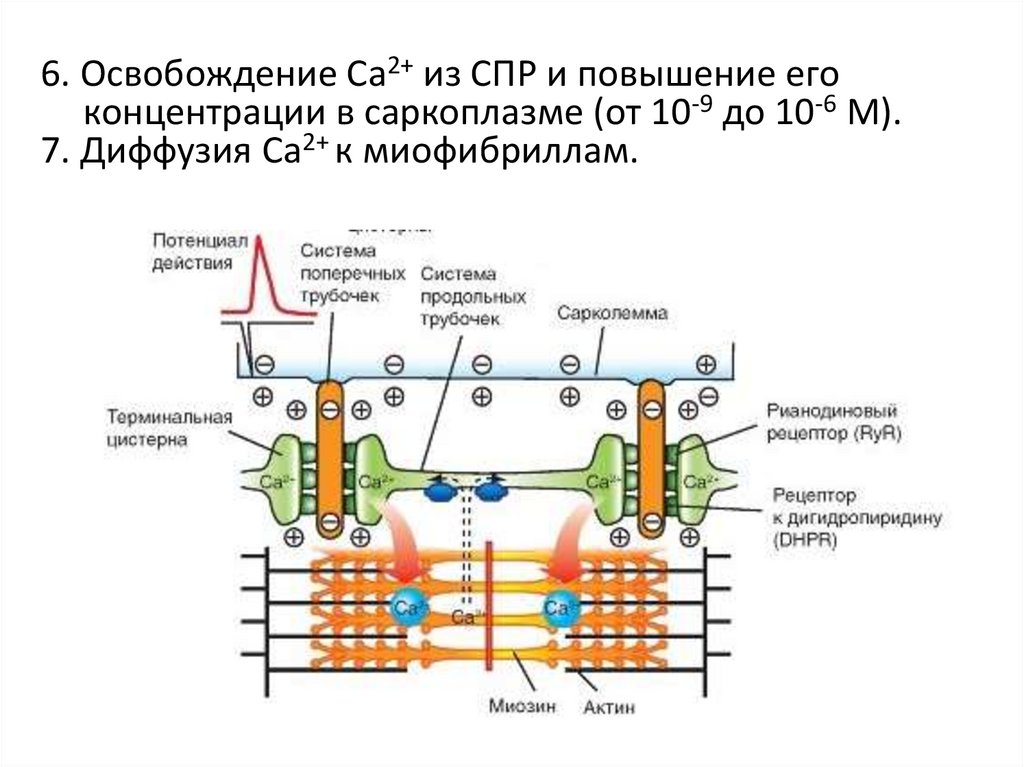
6. Освобождение Са2+ из СПР и повышение егоконцентрации в саркоплазме (от 10-9 до 10-6 М).
7. Диффузия Са2+ к миофибриллам.
17.
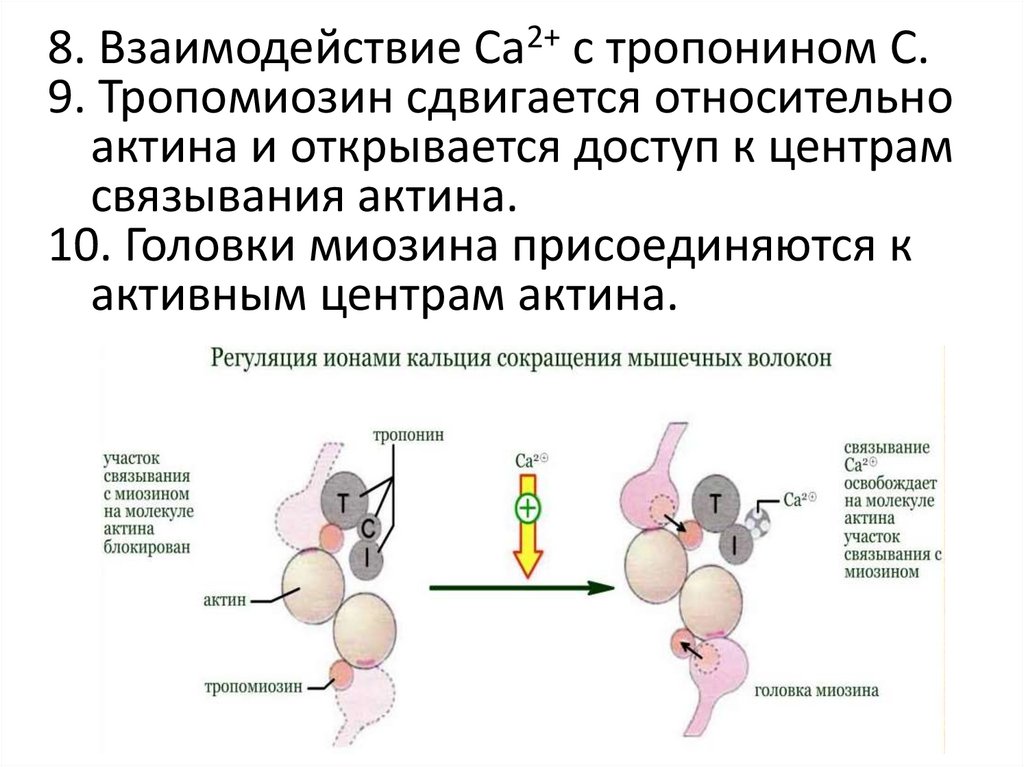
8. Взаимодействие Са2+ с тропонином С.9. Тропомиозин сдвигается относительно
актина и открывается доступ к центрам
связывания актина.
10. Головки миозина присоединяются к
активным центрам актина.
18.
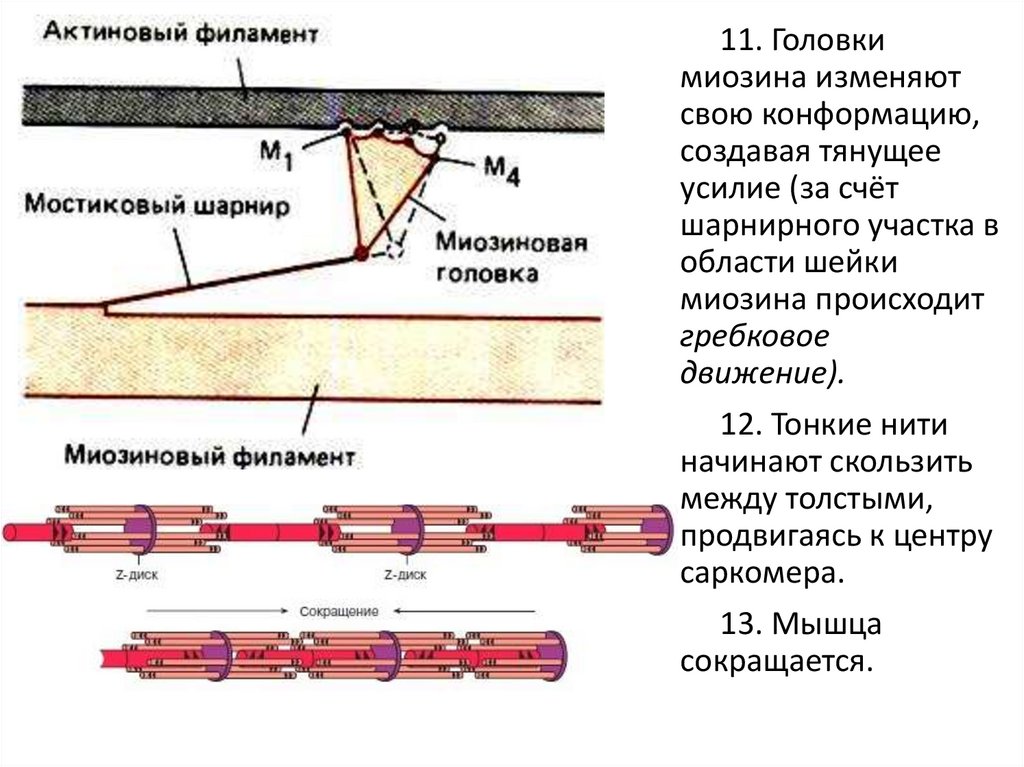
11. Головкимиозина изменяют
свою конформацию,
создавая тянущее
усилие (за счёт
шарнирного участка в
области шейки
миозина происходит
гребковое
движение).
12. Тонкие нити
начинают скользить
между толстыми,
продвигаясь к центру
сaркомера.
13. Мышца
сокращается.
19.
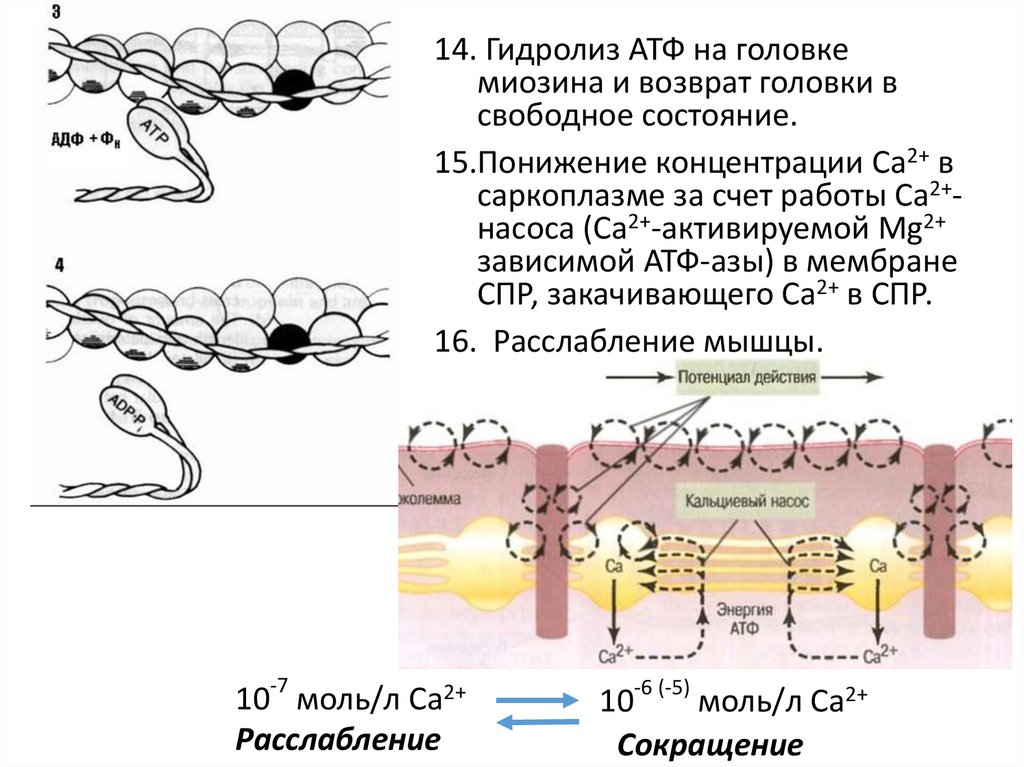
14. Гидролиз АТФ на головкемиозина и возврат головки в
свободное состояние.
15.Понижение концентрации Са2+ в
саркоплазме за счет работы Са2+насоса (Са2+-активируемой Mg2+
зависимой АТФ-азы) в мембране
СПР, закачивающего Са2+ в СПР.
16. Расслабление мышцы.
-7
10 моль/л Са2+
Расслабление
10-6 (-5) моль/л Са2+
Сокращение
20.
Энергозатраты:Энергия АТФ расходуется на:
1. Образование поперечных мостиков,
осуществляющих продольное скольжение
актиновых филаментов (основная часть энергии
гидролиза АТФ);
2. Ca2+-насос: выкачивание Ca2+ из саркоплазмы в
саркоплазматический ретикулум после окончания
сокращения;
3. Na+/K+-насос: перемещение ионов натрия и калия
через мембрану МВ для обеспечения
соответствующего ионного состава вне- и
внутриклеточной среды.
21.
Каким образом Са2+ выделяется из СПР?Существует несколько гипотез высвобождения Са2+
из СПР:
1. Волна деполяризации спускается по Т-трубочкам
и переходит на СПР благодаря близости её
мембраны. Вследствие деполяризации последней
происходит
открывание
Са2+-каналов
и
освобождение Са2+ из СПР (Зима В.Л. и McDonald
P.L.)
В настоящее время эта гипотеза является несостоятельной.
22.
2. Во внешнюю мембрану миоцита встроеныпотенциалзависимые и/или рецепторуправляемые
Са2+-каналы. Небольшое повышение
концентрации Са2+ в миоплазме при его входе из
внеклеточного пула индуцирует освобождение
Са2+ из СПР (Bianchi S.P. и Shanes А.М.)
Данная модель характерна для ГМК и кардиомиоцитов.
23.
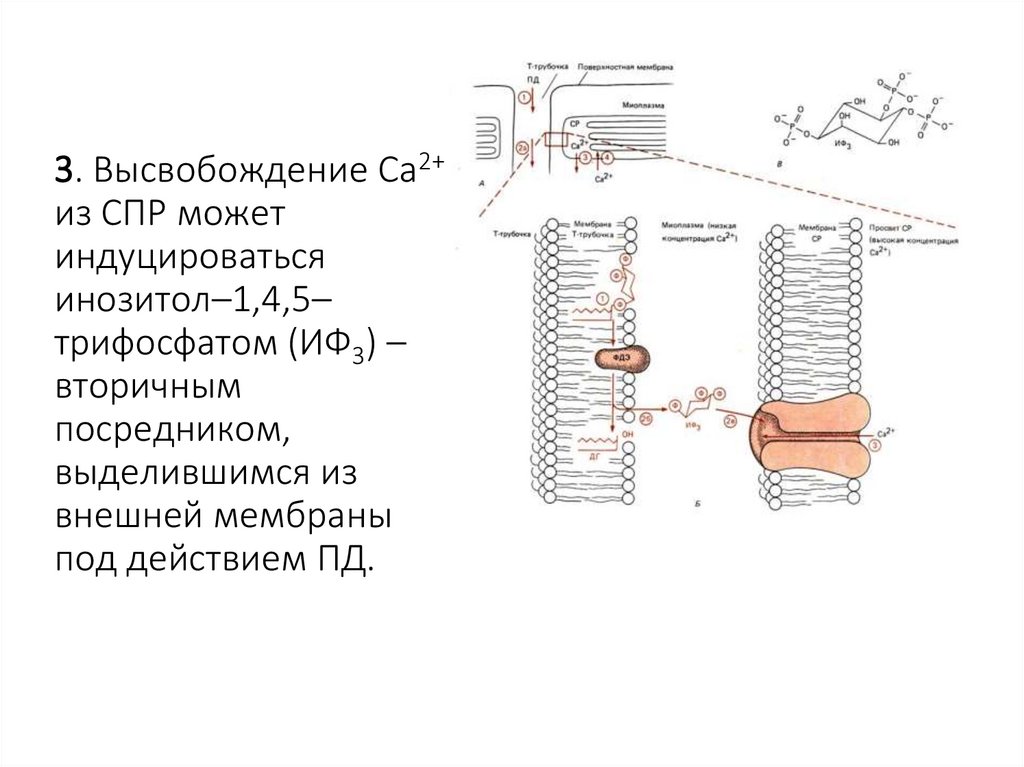
3. Высвобождение Са2+из СПР может
индуцироваться
инозитол–1,4,5–
трифосфатом (ИФ3) –
вторичным
посредником,
выделившимся из
внешней мембраны
под действием ПД.
24.
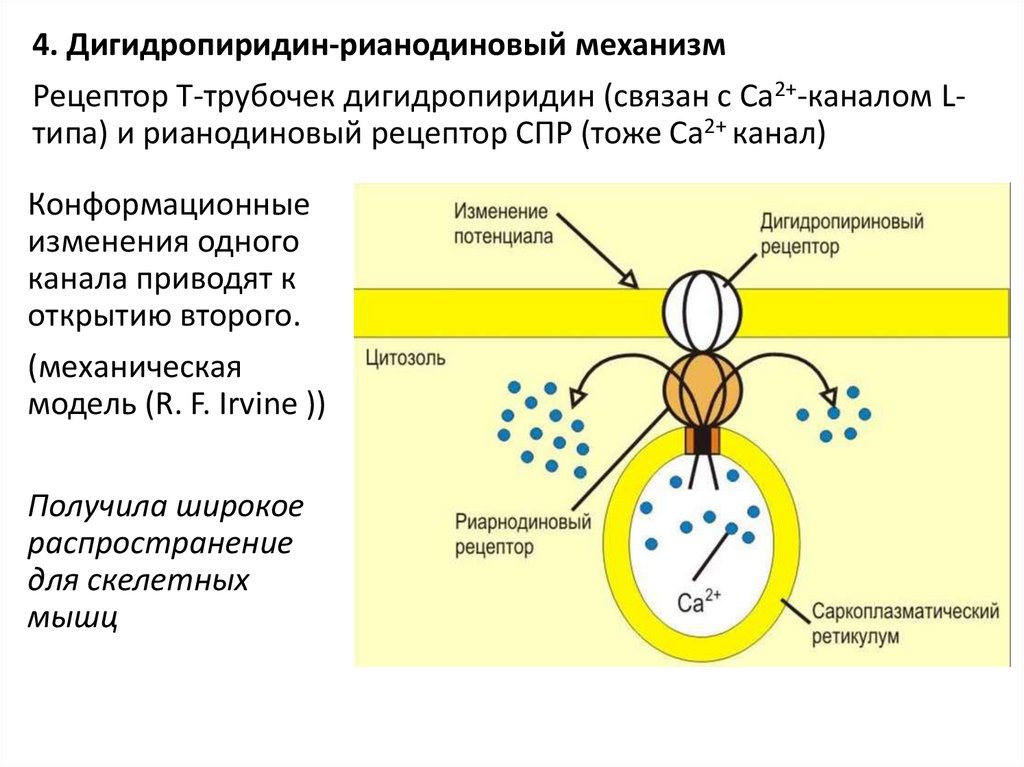
4. Дигидропиридин-рианодиновый механизмРецептор Т-трубочек дигидропиридин (связан с Са2+-каналом Lтипа) и рианодиновый рецептор СПР (тоже Са2+ канал)
Конформационные
изменения одного
канала приводят к
открытию второго.
(механическая
модель (R. F. Irvine ))
Получила широкое
распространение
для скелетных
мышц
25.
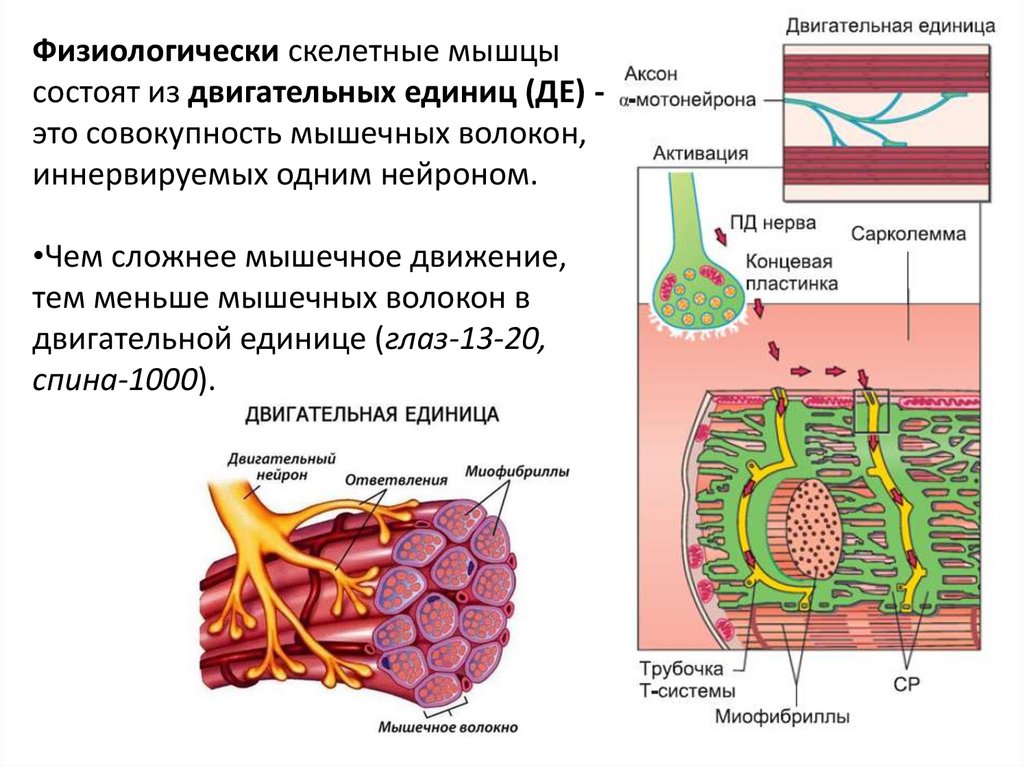
Физиологически скелетные мышцысостоят из двигательных единиц (ДЕ) это совокупность мышечных волокон,
иннервируемых одним нейроном.
•Чем сложнее мышечное движение,
тем меньше мышечных волокон в
двигательной единице (глаз-13-20,
спина-1000).
26.
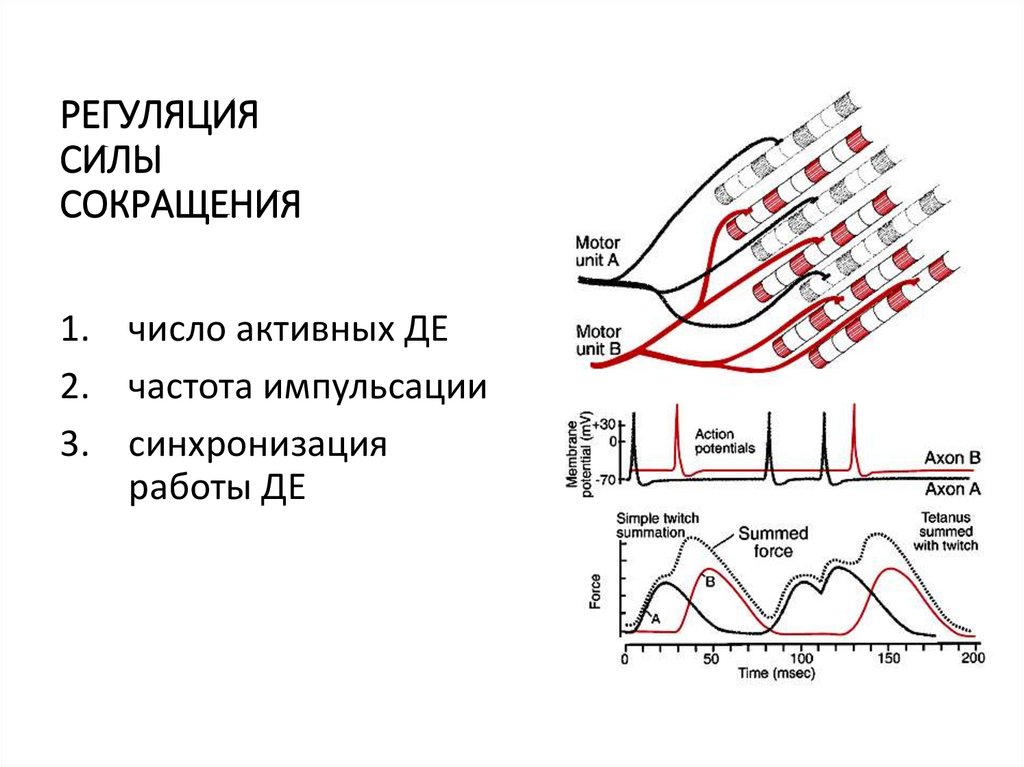
РЕГУЛЯЦИЯСИЛЫ
СОКРАЩЕНИЯ
1. число активных ДЕ
2. частота импульсации
3. синхронизация
работы ДЕ
27.
Режимы мышечныхсокращений.
Суммация мышечных
сокращений
28.
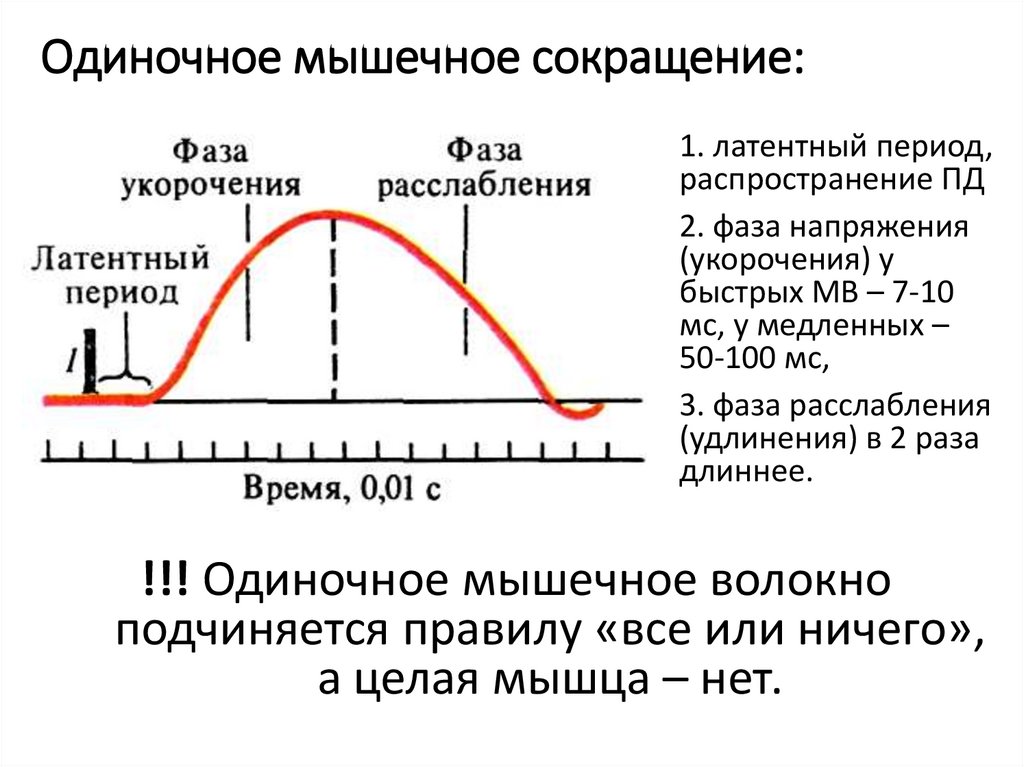
Одиночное мышечное сокращение:1.
2.
3.
1. латентный период,
распространение ПД
2. фаза напряжения
(укорочения) у
быстрых МВ – 7-10
мс, у медленных –
50-100 мс,
3. фаза расслабления
(удлинения) в 2 раза
длиннее.
!!! Одиночное мышечное волокно
подчиняется правилу «все или ничего»,
а целая мышца – нет.
29.
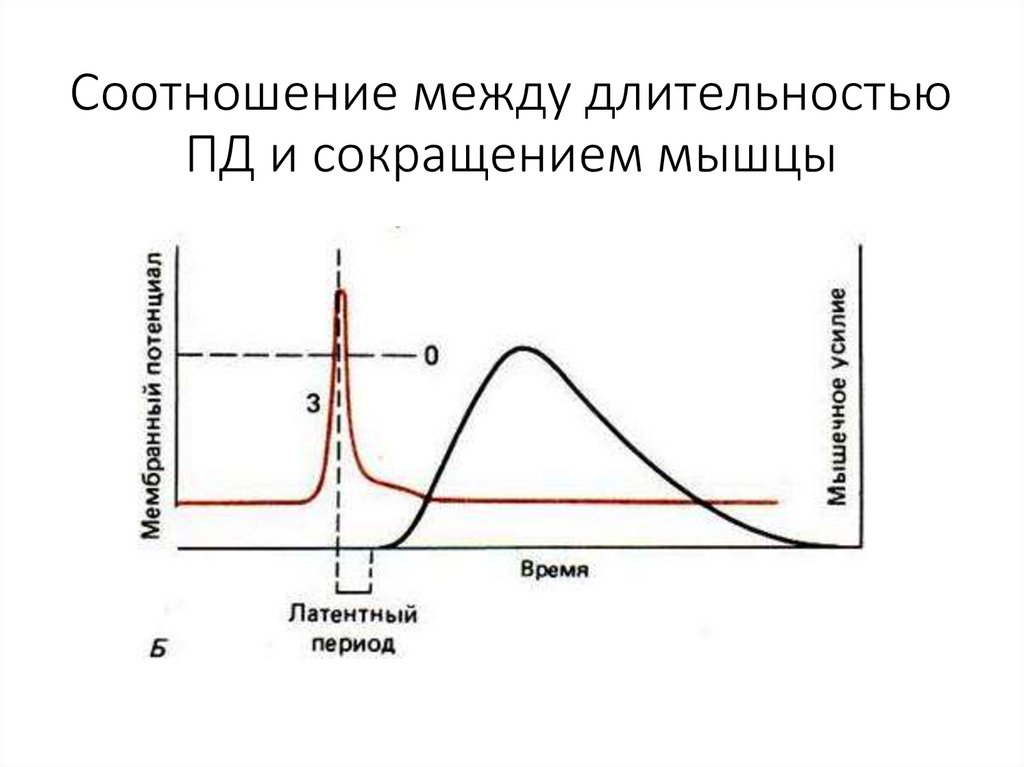
Соотношение между длительностьюПД и сокращением мышцы
30.
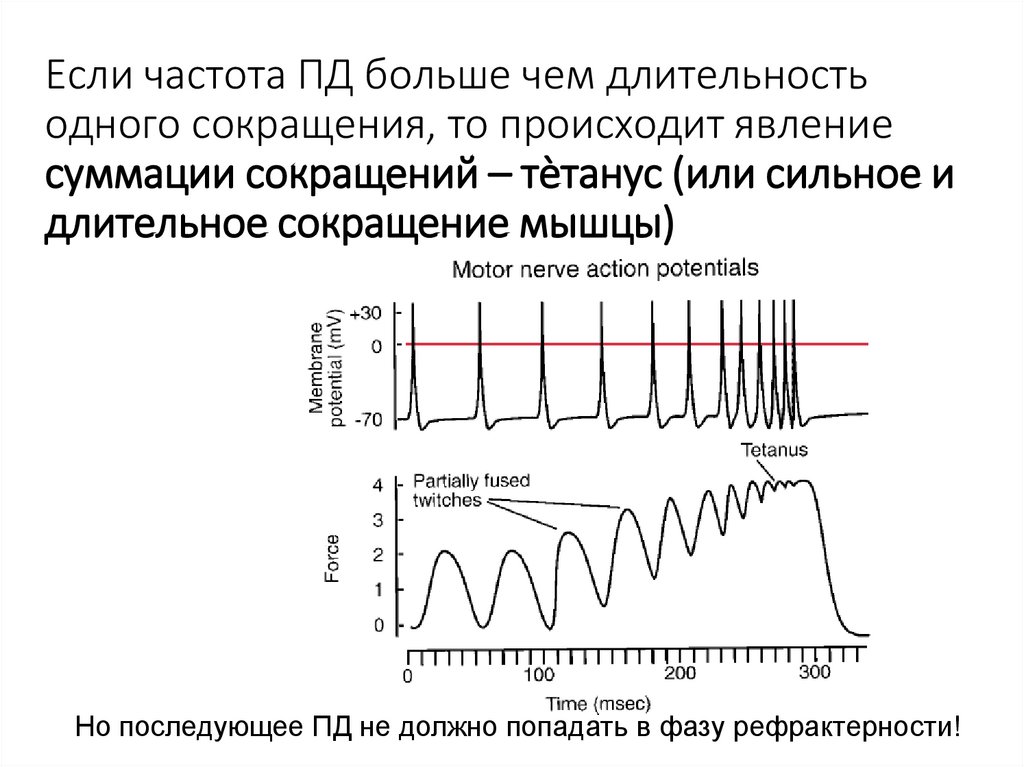
Если частота ПД больше чем длительностьодного сокращения, то происходит явление
суммации сокращений – тѐтанус (или сильное и
длительное сокращение мышцы)
Но последующее ПД не должно попадать в фазу рефрактерности!
31.
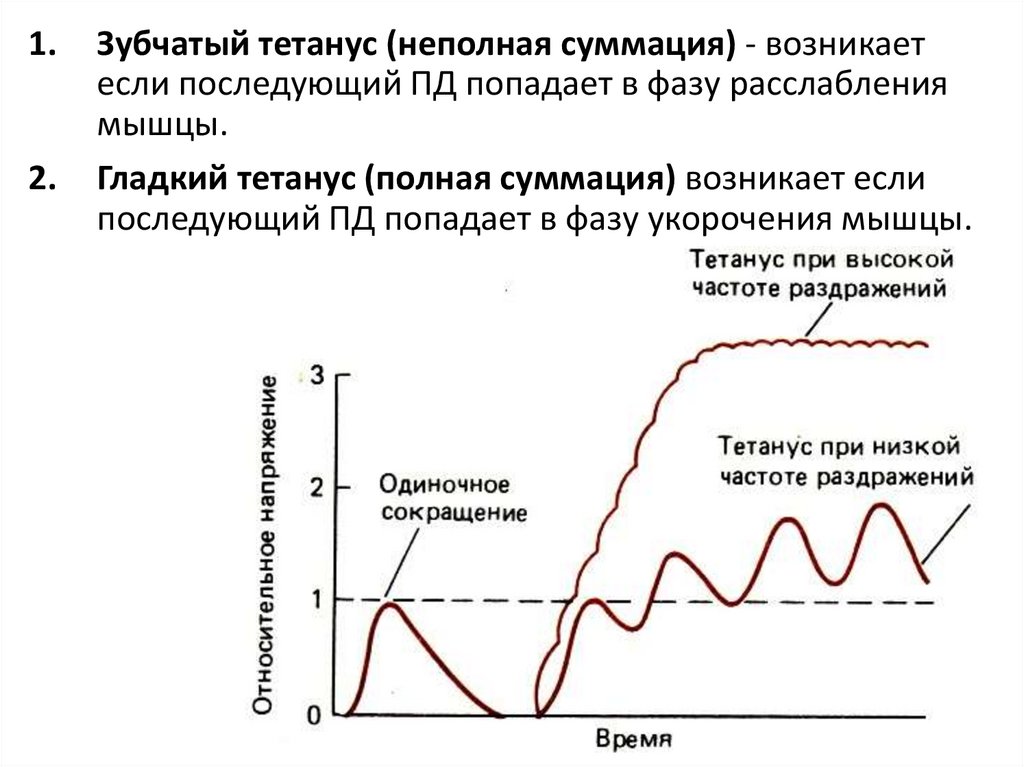
1.2.
Зубчатый тетанус (неполная суммация) - возникает
если последующий ПД попадает в фазу расслабления
мышцы.
Гладкий тетанус (полная суммация) возникает если
последующий ПД попадает в фазу укорочения мышцы.
32.
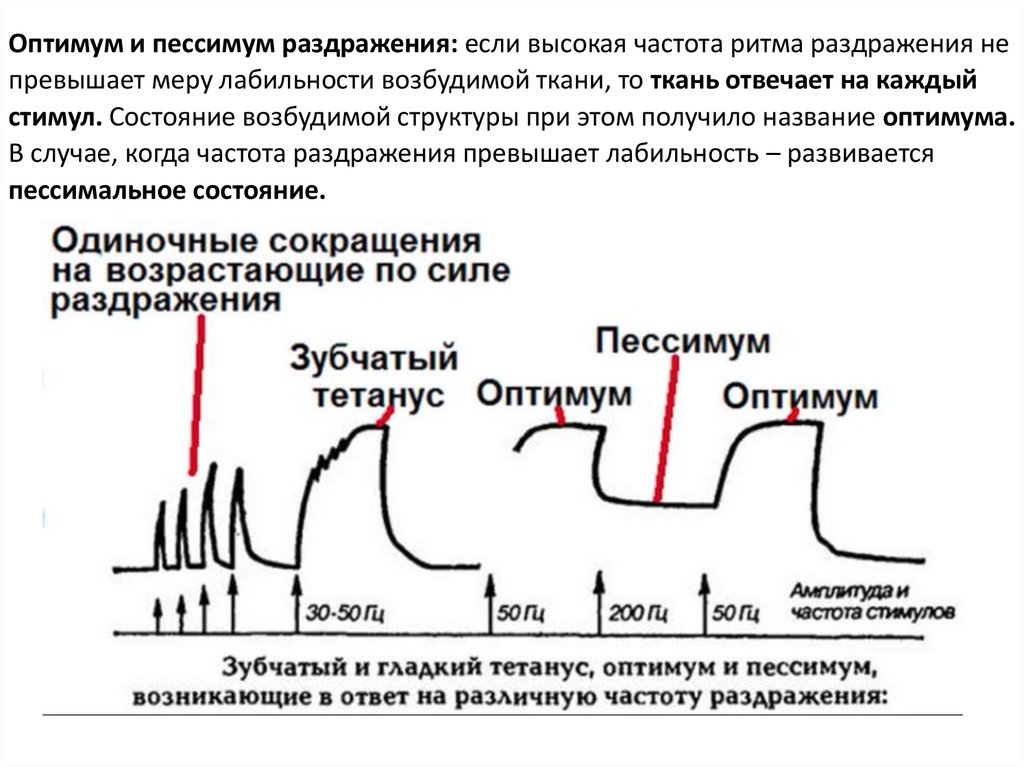
Оптимум и пессимум раздражения: если высокая частота ритма раздражения непревышает меру лабильности возбудимой ткани, то ткань отвечает на каждый
стимул. Состояние возбудимой структуры при этом получило название оптимума.
В случае, когда частота раздражения превышает лабильность – развивается
пессимальное состояние.
33.
Механизм оптимума – повышениевозбудимости (стимул попадает в
фазу экзальтации)
Механизм пессимума – снижение
возбудимости (стимул попадает в
фазу абсолютной рефрактерности!)
34.
Теории возникновения тетануса1. Гельмгольц – тетанус – это механическое
сложение одиночных мышечных сокращений,
2. Введенский – тетанус наступает если каждый
последующий импульс раздражения попадает в
фазу экзальтации предыдущего ПД,
3. Бабский – тетанус возникает в результате
суммации концентраций Са2+ , выделяющегося
из СПР после предыдущего и последующего
раздражений.
35.
КОНТРАКТУРА- обратимое нераспространяющееся стационарное
сокращение
Типы контрактур:
1. Посттетаническая (высокий остаточный Са2+)
2. Кофеиновая – кофеин проникая внутрь клетки
приводит к открытию Са2+-каналов в СПР.
3. Калиевая – увеличение К+ во внеклеточной среде
приводит к стойкой деполяризации и высвобождению
Са2+ из СПР
Главной причиной развития контрактур является
истощение запасов АТФ в мышцах.
36.
Основные физиологическиесвойства скелетных мышц
37.
По характеру сокращения:1. Фазные - осуществляют энергичные сокращения
(произвольная мускулатура человека). Могут
возбуждаться на одиночный импульс
раздражения.
2. Тонические - специализированы на поддержание
тонусa (позные мышцы, наружные ушные и
наружные глазные мышцы). Сокращаются очень
медленно слабо и долго.
38.
По скорости сокращения(определяется типом миозина)
1. Быстрые - быстрый миозин (высокая активность АТФ-азы).
• Аксоны имеют мало терминалей и мало иннервируемых мышечных волокон.
• В миофибриллах хорошо развит сарко-плазматический ретикулум (СПР) и Тсистема.
• Активен Са++насос.
• Сокращаются быстро, сильно и сразу утомляются.
2. Медленные - медленный миозин (меньшая активность АТФ-азы).
• Аксоны имеют много терминалей и иннервируют много миофибрилл (от сотен до
2000).
• Хуже развит СПР и Т-система. Много миоглобина, поэтому они имеют красный цвет.
• Имеют много митохондрий.
• Сокращаются медленно, сильно и долго без утомления, а после утомления быстро
восстанавливаются.
• Преобладают в мышцах, поддерживающих позу человека, четырёхглавой мышце
бедра, икроножной и дыхательных мышцах.
39.
По выполняемой работеКонцентрический тип: когда внешняя нагрузка
меньше, чем развиваемое мышцей напряжение
(мышца укорачивается и совершает движение положительная работа).
Эксцентрический тип: когда внешняя нагрузка
больше, чем развиваемое мышцей напряжение
(мышца растягивается и совершает
отрицательную работу).
40.
По типу окисления:1. Гликолитические (белые): не имеют миоглобина, мало митохондрий,
АТФ образуется в основном по механизму гликолиза и быстро
расщепляется, т.к. высокоактивна АТФ-аза.
2. Окислительные (красные): занимают промежуточное положение
между фазными быстрыми ДЕ гликолитического типа и фазными
медленными ДЕ. В них содержится достаточно много миоглобина,
имеется много митохондрий, синтезируется достаточное количество
АТФ по механизму окислительного фосфорилирования. Однако
активность АТФ-азы у них низкая. Они сокращаются быстро, сильно и
достаточно долго. Такие НМЕ преобладают в мышцах длительно
летающих птиц.
41.
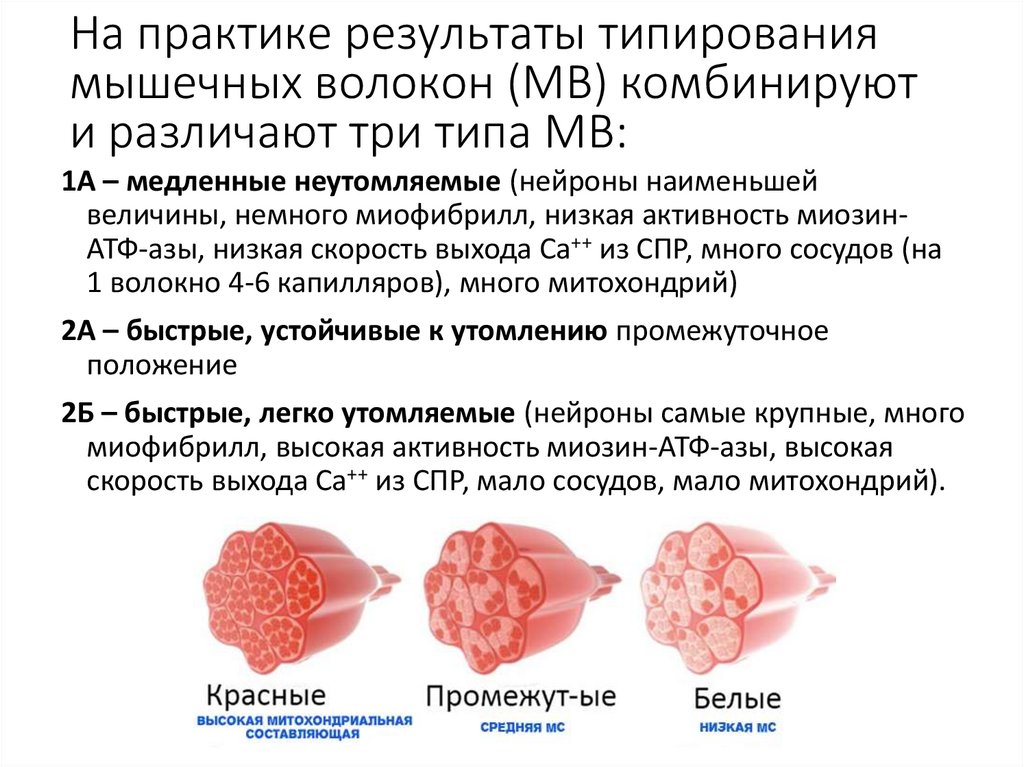
На практике результаты типированиямышечных волокон (МВ) комбинируют
и различают три типа МВ:
1А – медленные неутомляемые (нейроны наименьшей
величины, немного миофибрилл, низкая активность миозинАТФ-азы, низкая скорость выхода Са++ из СПР, много сосудов (на
1 волокно 4-6 капилляров), много митохондрий)
2А – быстрые, устойчивые к утомлению промежуточное
положение
2Б – быстрые, легко утомляемые (нейроны самые крупные, много
миофибрилл, высокая активность миозин-АТФ-азы, высокая
скорость выхода Са++ из СПР, мало сосудов, мало митохондрий).
42.
ВИДЫ СОКРАЩЕНИЯ:1. изометрическое (длина мышцы не
меняется, изменяется лишь напряжение
(при фиксированных концах мышцы).
2. изотоническое (длина мышцы меняется
при неизмененном напряжении)
3. ауксотоническое (меняется длина и
напряжение мышцы)
43.
Физиологические особенностигладких мышц
1) нестабильный мембранный потенциал, который
поддерживает мышцы в состоянии постоянного частичного
сокращения – тонуса;
2) самопроизвольная автоматическая активность;
3) сокращение в ответ на растяжение;
4) высокая чувствительность к химическим веществам.
5) для сокращения ГМК Са2+ используют не только из СПР, но
и из внеклеточной среды.
6) фазы укорочения и расслабления ГМК длятся в среднем от
1 до 3 сек, что в десятки раз продолжительнее сокращения
скелетной мышцы.
44.
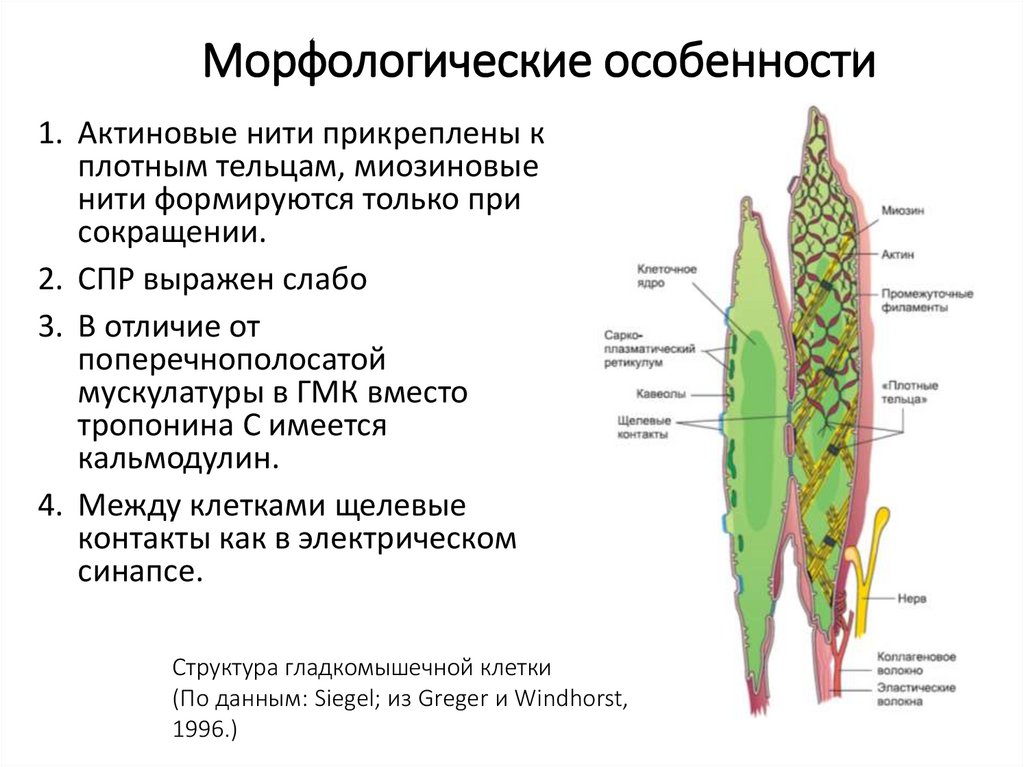
Морфологические особенности1. Актиновые нити прикреплены к
плотным тельцам, миозиновые
нити формируются только при
сокращении.
2. СПР выражен слабо
3. В отличие от
поперечнополосатой
мускулатуры в ГМК вместо
тропонина С имеется
кальмодулин.
4. Между клетками щелевые
контакты как в электрическом
синапсе.
Структура гладкомышечной клетки
(По данным: Siegel; из Greger и Windhorst,
1996.)
45.
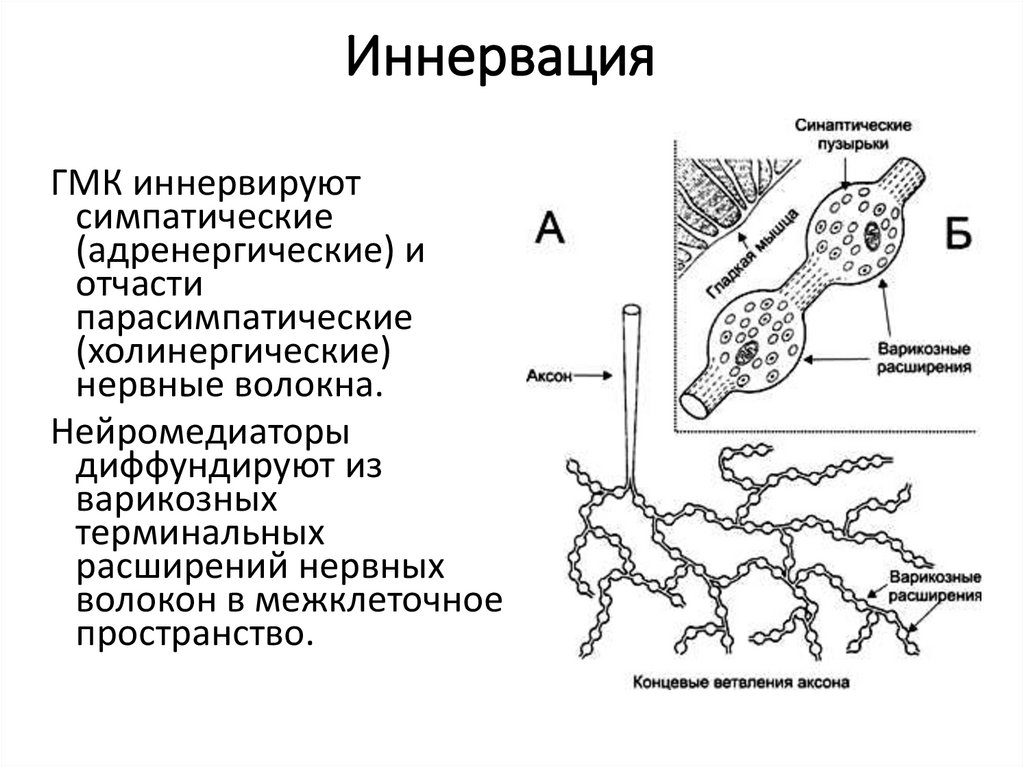
ИннервацияГМК иннервируют
симпатические
(адренергические) и
отчасти
парасимпатические
(холинергические)
нервные волокна.
Нейромедиаторы
диффундируют из
варикозных
терминальных
расширений нервных
волокон в межклеточное
пространство.
46.
Гуморальная регуляцияВ мембрану разных ГМК встроены рецепторы
ацетилхолина, гистамина, атриопептина, ангиотензина,
адренорецепторы и множество других.
Сокращение ГМК вызывают адреналин, норадреналин,
ангиотензин, вазопрессин, действуя на свой рецептор и
активируя G-белок, который в свою очередь активирует
фосфолипазу С.
Расслабление ГМК вызывают атриопептин, брадикинин,
гистамин, VIP, которые действуют на G-белок,
активирующий аденилатциклазу.
Характер ответа определяют рецепторы.
47.
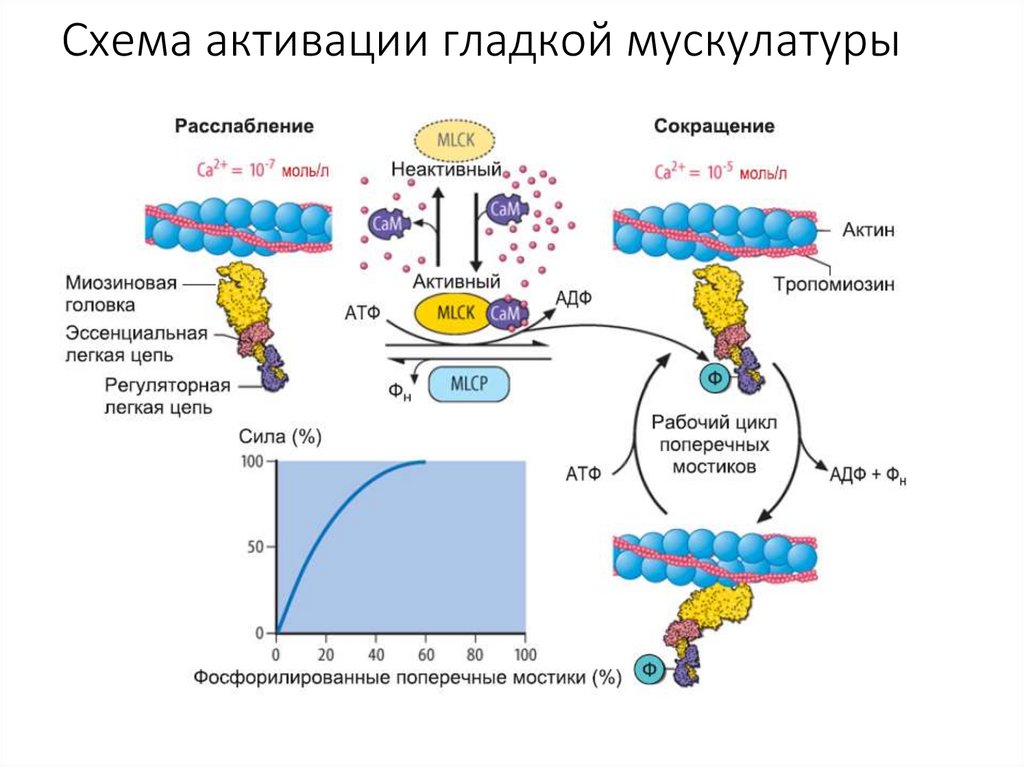
Схема активации гладкой мускулатуры48.
Холиномиметики усиливаютсокращение
1. оказывают на мышцу тот же эффект, что и ацетилхолин.
Различие заключается в том, что эти вещества не
разрушаются ацетилхолинэстеразой или разрушаются
более медленно, в течение многих минут и даже часов
(метахолин, карбахол и никотин).
2. инактивируют ацетилхолинэстеразу таким образом, что
она теряет способность гидролизовать ацетилхолин,
выделяющийся в концевой двигательной пластинке
(неостигмин, физостигмин и
диизопропилфлуорофосфат).