


















")















 biology
biologySimilar presentations:


")




")


Фізіологічні властивості серця. Будова серця
1.
ТЕМА ЛЕКЦІЇ:“ФІЗІОЛОГІЧНІ
ВЛАСТИВОСТІ СЕРЦЯ”
2. БУДОВА СЕРЦЯ
3. БУДОВА СЕРЦЯ
Серце більшою своєю частиною (2/3) лежить у лівійполовині грудної порожнини, меншою частиною (1/3)
у правій. Поздовжня вісь серця спрямована косо:
зверху вниз, справа наліво і ззаду наперед. Форма
серця непостійна і залежить від віку, конституції.
Розрізняють три основних положення серця: косе,
поперечне і вертикальне. Встановлена корелятивна
залежність ступеня нахилу серця від конституції. В
людей з широкою грудною кліткою (брахіморфна
будова тіла) серце розміщено здебільшого поперечно, в
людей з вузькою і довгою грудною кліткою
(доліхоморфна будова тіла) воно розташоване більш
вертикально.
4. БУДОВА СЕРЦЯ
Міокард, або серцевий м'яз, складається із серцевої м'язовоїтканини і прошарків пухкої сполучної тканини з судинами та
нервами.
Серцева
м'язова
тканина
за
будовою
є
поперечнопосмугованою. Поперечна смугастість має ту ж
природу, що і в скелетних м'язах, тобто зумовлена оптичною
неоднорідністю міофібрил, які побудовані з двох типів
міофіламентів. Серцевий м'яз побудований з волокон, які
анастомозують між собою, утворюючи сітку. Між волокнами
розташована пухка сполучна канина, багата судинами та нервами.
Усі м'язові волокна серцевого м'яза утворені окремими одно- або
двоядерними м’язовими клітинами, які розташовані ланцюжком і
мають у розрізі прямокутну форму. Ці клітини називають
кардіоміоцитами. Вони бувають двох видів: скоротливі, або
типові серцеві міоцити і провідні, або атипові, серцеві міоцити,
що належать до так званої провідної системи серця.
5. Структурно-функціональні особливості серця
Стінка серця складається з трьохшарів: ендокарда, міокарда і епікарда.
Міокард утворюється з окремих
м'язових волокон, які складаються з
послідовно з'єднаних (кінець в кінець)
клітин- кардіоміоцитів, що мають
спільну мембрану, це так звані
нексуси.
Нексуси забезпечують функціональну
однорідність (функціональний
синцитій). Нексуси надзвичайно
чутливі до нестачі кисню, впливу
катехоламінів, а також до значних
фізичних навантажень. Це може
викликати порушення проведення
збудження в міокарді.
6. Скоротливі кардіоміоцити
У скоротливих міоцитах багато саркоплазми і відносно маломіофібрил. У саркоплазмі скоротливих міоцитів є значна
кількість мітохондрій. Саркоплазматична сітка не так сильно
розвинена, як у скелетних м'язах, і не утворює великих
термінальних цистерн. У клітинах серцевого м'яза Т-трубочки
заходять всередину на рівні Z-пластинок, тому кількість їх
відповідає числу саркомерів. Т-трубочки у два рази ширші, ніж
у скелетних м'язах і, крім того, відрізняються тим, що
вистелені базальною мембраною, яка лежить назовні від
сарколеми. Тут також відсутня типова картина тріад, тому що
цистерни саркоплазматичної сітки, які контактують з Ттрубочками малі і не утворюють повних кілець навколо
міофібрил.
7. Атипові кардіоміоцити
Серед атипових кардіоміоцитів за морфологічними та функціональнимиособливостями можна визначити три типи клітин. Клітини першого типу
мають назву пейсмейкерних клітин (Р-клітин), або водіїв ритму. Пейсмейкерні
клітини локалізуються у центральній частині синоатріального вузла.
Саркоплазматична сітка розвинена слабо, Т-система відсутня.
Клітини другого типу – це перехідні клітини, функціональне значення яких
полягає у передачі збудження від Р-клітин до клітин провідних шляхів і
скоротливих елементів міокарду. Локалізуються ці клітини на периферії
синоатріального вузла і становлять більшу частину атріовентрикулярного
вузла. Морфологічно це тонкі витягнуті клітини, менші за діаметром, ніж
типові кардіоміоцити. Міофібрил у них дещо більше, ніж у Р-клітинах, але
менше, ніж у скоротливих кардіоміоцитів і розташування їх менш
впорядковане.
Клітини третього типу - це клітини пучка провідної системи та його ніжок (так
звані волокна Пуркіньє). Вони передають збудження від перехідних клітин до
скоротливих кардіоміоцитів шлуночків. Міофібрил у них мало, вони
розташовуються на периферії волокна, орієнтовані у різних напрямках.
.
8. Будова міокарда передсердь
Міокард передсердь має двашари: циркулярний і
поздовжній.
М'язові шари передсердя.
1 - м'язи, які оточують гирла
лівих легеневих вен; 2 - м'язи, які
оточують гирла правих легеневих
вен; 3 - праві легеневі вени; 4 верхня порожниста вена; 5 - м'язи
гирла верхньої порожнистої вени;
6 - м'язи правого передсердя; 7 нижня порожниста вена; 8 - гирло
венозного синуса; 9 - м'язи лівого
передсердя; 10 - ліві легеневі
вени.
9. Будова міокарда передсердь
На відміну від шлуночкових кардіоміоцитів, якімають циліндричну форму, передсердні міоцити
часто мають відростки, у них менше мітохондрій,
міофібрил, та не розвинена саркоплазматична
сітка. Особливістю цих міоцитів є також відносно
добре виражена гранулярна ендоплазматична сітка
і комплекс Гольджі, які беруть участь у синтезі
специфічних передсердних гранул діаметром 300400
нм.
Гранули
містять
передсердний
натрійуретичний фактор, який за своїм впливом є
антагоністом
ренін-ангіотензин-альдостеронової
системи.
10.
Будова міокарда шлуночківВ міокарді шлукочків виділяють три шари.
Зовнішній і внутрішній шари мають
спіралеподібну форму і є спільними для
обох шлуночків. Середній шар – це шар
циркулярних волокон, який йде окремо в
кожному шлуночку.
Шари міокарда шлуночків серця.
1 – волокна поверхневого шару, які йдуть
від лівого фіброзного кільця до правого
шлуночка; 2 - поверхневий шар міокарда в
стінці правого шлуночка; 3 - завиток
серця; 4 – глибокий шар міокарда в стінці
лівого шлуночка; 5 - середній шар
міокарда в стінці лівого шлуночка; 6 поверхневий шар міокарда в стінці лівого
шлуночка.
11. Віртуальна модель міокарда шлуночків
• Для розуміння гвинтового напрямкуповздовжніх субендо-і
субепікардіальних волокон, потрібно
уявити рух «по спіралі» пальців
складених рук. Рух вгору по спіралі в
бік пальців правої руки назвемо
правонаправленою спіраллю (R), якщо
ж повернути складені долоні лівої
руки, то це - лівонаправлена спіраль
(L). Цей рух притаманний даному
відтинку волокна, і не можливо по
ходу волокна в одному місці змінити
напрямок спіралі з право на
лівонаправлену. Це виключає ефекти їх
накладання і перехрещування.
12. Віртуальна модель міокарда шлуночків
Моделювання напрямку волокон міокарда лівогошлуночка показує, що гвинтовий правонаправлений
напрямок субендокардіальних волокон поступово
переходить у лівонаправлену спіраль
субепікардіальних волокон.
Рис. А. Флюоресцентні фарби забарвлюють
різноспрямовані вихрові сили: внутрішній вихровий
потік опускається (червона фарба) а зовнішній піднімається (зелена фарба).
Рис В,С. Поздовжній і поперечний зріз у вигляді двох
різноспрямованих спіралей: червоний колір –
низхідний потік; синій - висхідний.
Рис D. Субендокардіальна частина має геометричну
конфігурацію, по якій рух відбувається в напрямку
правонаправленої спіралі, а субепікардіальні
волокна обертаються в напрямку лівонаправленої.
Примітки:1 - субендокардіальні волокна;
2 - сосочкові м'язи,
3 - завиток верхівки
4 - периферичні волокна волокна,
5 - субепікардіальні волокна.
13. Провідна система серця
До складу провіденої системи входять: синусовий вузол (синуснопередсердний, синоатріальний), який знаходиться біля місцявпадіння порожнистих вен у праве передсердя. Від синусового вузла
до вушка лівого передсердя йде міжпередсердний пучок Бахмана. А
до другого вузла провідникової системи - передсердно-шлуночкового
(атріовентрикулярного) – йдуть міжвузлові провідні тракти (пучки
Бахмана, Венкебаха та Тореля). Від атріовентрикулярного вузла йде
міжшлуночковою перегородкою пучок Гіса (передсердношлуночковий пучок), який ділиться на дві ніжки праву та ліву. Ліва
ніжка в свою чергу ділиться на передню та задню гілки. Права ніжка
та гілки лівої ніжки переходять у волокна Пуркін'є.
Крім основних елементів провідної системи є додаткові її елементи:
пучок Кента, пучок Джеймса і пучок Махейма. Ці пучки можуть
проводити збудження із передсердь до шлуночків. Пучок Кента може
проводити збудження від передсердь, в обхід атріовентрикулярного
вузла, до правого шлуночка. Пучок Джеймса може імпульси з
передсердь проводити до пучка Гіса в обхід атріовентрикулярного
вузла. Пучок Махейма може імпульси від атріовентрикулярного
вузла, обминаючи пучок Гіса і нижче лежачі відділи, нести до лівого
шлуночка.
14.
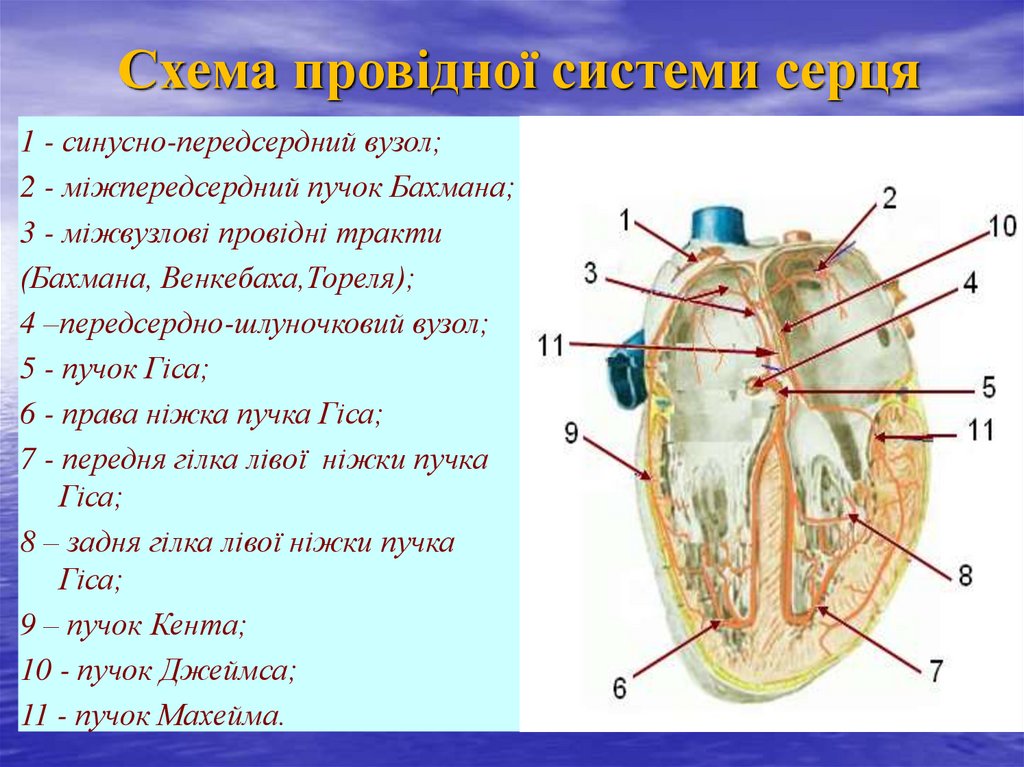
Схема провідної системи серця1 - синусно-передсердний вузол;
2 - міжпередсердний пучок Бахмана;
3 - міжвузлові провідні тракти
(Бахмана, Венкебаха,Тореля);
4 –передсердно-шлуночковий вузол;
5 - пучок Гіса;
6 - права ніжка пучка Гіса;
7 - передня гілка лівої ніжки пучка
Гіса;
8 – задня гілка лівої ніжки пучка
Гіса;
9 – пучок Кента;
10 - пучок Джеймса;
11 - пучок Махейма.
15.
Основні типи іонних каналів в кардіоміоцитах:1) Швидкі натрієві канали, які знаходяться у клітинах міокарду
передсердь і шлуночків, у волоконах Пуркиньє та А-В вузла.
2) Кальцієві канали:
а) L-типу (від англійського lolng-lasting – довготривалий), які активуються
при заряді мембрани біля -20 мВ;
б) Т-типу (від англійського transient – тимчасовий), які активуються при
заряді мембрани біля -50 мВ;.
3) Калієві канали:
а) сповільненого випрямлення, які впливають на перебіг фази плато
кардіоміоцитів;
б) аномального випрямлення, через які калій виходить із клітини у фазу
швидкої реполяризації скоротливого міокарду передсердь і шлуночків та
волокон Пуркиньє.
4) Ліганд-залежні канали:
а) ацетилхолін-чутливі калієві канали зустрічаються в клітинах
синоатріального і атріовентрикулярного вузлів, клітинах міокарду передсердь;
б) АТФ-чутливі калієві канали властиві клітинам скоротливого міокарду
передсердь і шлуночків;
в) кальцій-активовані неспецифічні канали, зустрічаються в клітинах
скоротливого міокарду шлуночків і у волокнах Пуркиньє.
16. Фізіологічні властивості серця
• Автоматизм• Провідність
• Збудливість
• Рефрактерність
• Скоротливість
17.
Електрофізіологічні властивостіатипових клітин
В клітинах провідної системи відсутній стабільний
потенціал спокою, замість нього присутня фаза
повільної спонтанної діастолічної деполяризації, яка
обумовлена входом через мембрану кардіоміоцита
іонів Са2+, що веде до нагромадження в клітині
позитивно заряджених іонів. При досягненні
мембранного потенціалу рівня критичної точки
деполяризації (біля -60 мВ), спонтанно виникає
потенціал дії, що поширюється провідною
системою, а звідти на скоротливий міокард.
18.
• Автоматизм серця – це здатність клітин провідноїсистеми серця самостійно (автономно) виробляти
біоелектричні імпульси, які викликають його збудження.
• Структури провідної системи мають різний ступінь
автоматизму. Встановлено так званий ґрадієнт
автоматії. Він проявляється в зниженні здатності до
автоматизму різних структур провідної системи в міру її
віддалення від синусно-передсердного вузла. Так, якщо в
синусно-передсердному вузлі кількість потенціалів дії в
середньому складає 60-90 імп/хв, а в клітинах пучка Гіса 30-40 імп/хв, то в волокнах Пуркін'є - менше 20 імп/хв.
Градієнт автоматії обумовлений різною спонтанною
проникністю мембрани клітин провідної системи до іонів
Са2+. Виходячи з того, що синусно-передсердний вузол
нав'язує свій ритм нижче лежачим відділам провідної
системи його називають водієм ритму першого порядку
або пейсмекером першого порядку. Водієм ритму другого
порядку, є атріо-вентрикулярний вузол. Водієм ритму
третього порядку є пучок Гіса і його розгалуження.
19.
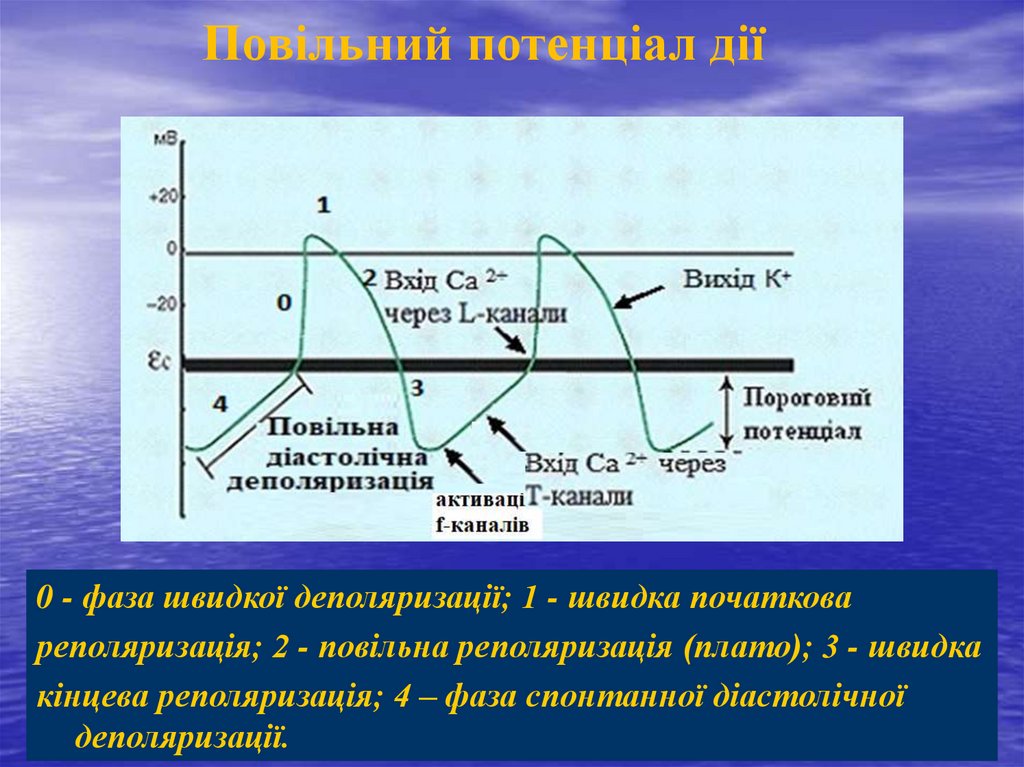
Повільний потенціал дії0 - фаза швидкої деполяризації; 1 - швидка початкова
реполяризація; 2 - повільна реполяризація (плато); 3 - швидка
кінцева реполяризація; 4 – фаза спонтанної діастолічної
деполяризації.
20.
У минулому столітті існувало 3 основних теорії автоматії серця.Прохаска і Мюллер висунули нейрогенну теорію, вважаючи причиною
його ритмічних скорочень нервові імпульси. Гаскелл і Енгельман
запропонували міогенну теорію, згідно з якою імпульси збудження
виникають у самому серцевому м'язі. Існувала теорія гормону серця,
який виробляється в ньому і ініціює його скорочення. В кінці 19
століття в різних ділянках міокарда передсердь і шлуночків були
виявлені скупчення своєрідних за будовою, м'язових клітин, які назвали
атиповими. Ці клітини більші в діаметрі, ніж скоротливі, в них менше
скорочувальних елементів та більше гранул глікогену. В останні роки
встановлено, що ці скупчення утворені Р-клітинами (клітинами
Пуркіньє) або пейсмекерами. Крім того, у цих скупченнях є також
перехідні клітини. Вони займають проміжне положення між
скоротливими і Р-клітинами та служать для передачі збудження,
утворюючи провідну систему серця.
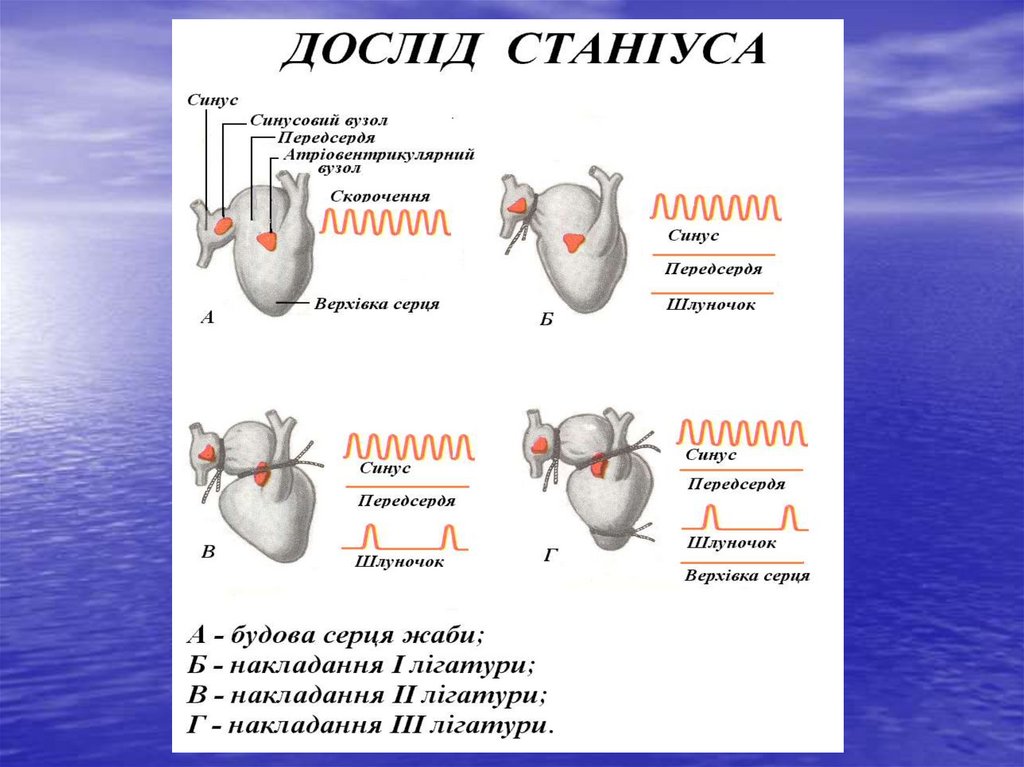
Роль різних відділів провідної системи в автоматії серця вперше була
встановлена Станніусом і Гаскеллом.
21.
22. Засвоєння ритму
У звичайних умовах автоматіявсіх ділянок провідної системи
пригнічується
синуснопередсердним
вузлом,
який
нав'язує їм свій ритм. Тому всі
частини провідної системи хоча і
мають власний ритм, починають
працювати в єдиному ритмі.
Явище, при якому структури з
сповільненим ритмом генерації
потенціалів дії засвоюють більш
частий ритм інших ділянок
провідної системи називається
засвоєнням ритму.
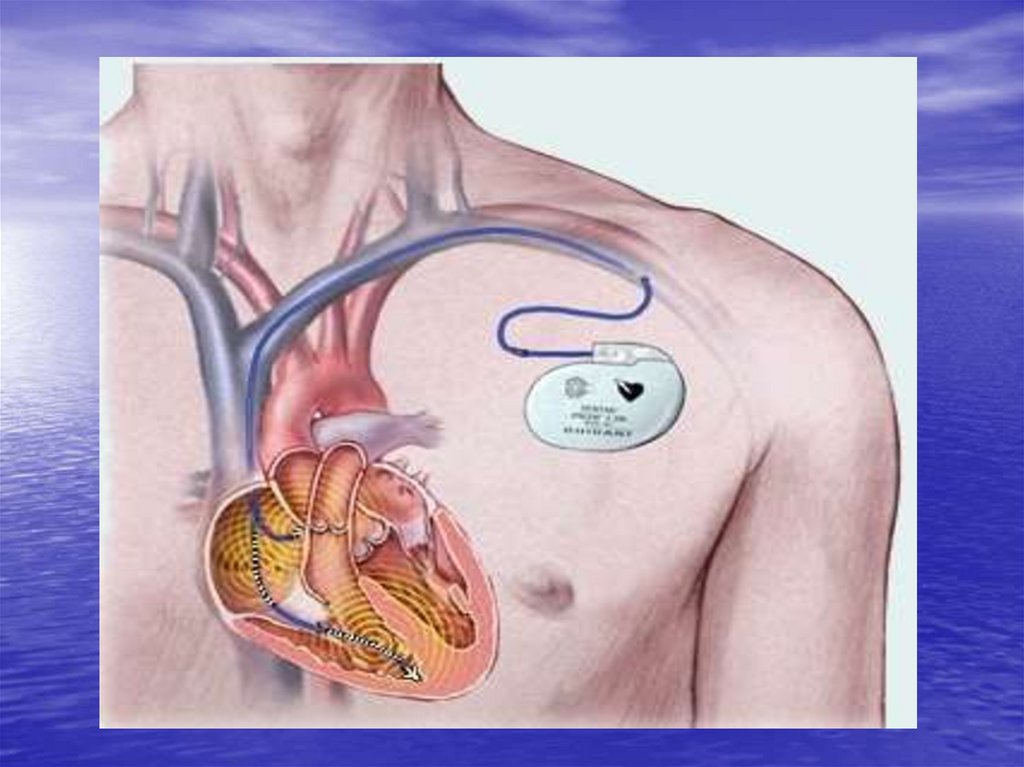
23. Клініко-фізіологічні аспекти встановлення штучного водія ритму (кардіостимулятора)
Кардіостимулятор - це складний електронний прилад, який виконує дві функції: 1)аналізує роботу серця, тобто його ритм і стан провідності; 2) при необхідності, він
посилає серцю регулярні електричні імпульси для корекції порушення власного ритму
серця.
Існує три типи кардіостимуляторів:
Однокамерний - від нього відходить один електрод, який імплантується в передсердя
або шлуночки серця.
Двохкамерний - від нього відходить два електроди - один до передсердям, а інший - до
шлуночка.
«Бівентрикулярной - від нього відходить три електроди - один до передсердя, і по
одному до кожного шлуночка.
Пацієнту під місцевим знеболенням роблять розріз, паралельний ключиці. Далі вводять
електроди кардіостимулятора в певні ділянки серця. Коли всі електроди
кардіостимулятора будуть з'єднані з серцем, їх під’єднують до блоку живлення апарата
Сам кардіостимулятор, точніше блок живлення, розміщують під ключицею в підшкірножировій клітковині. Кардіостимулятор починає штучно генерувати імпульси. Коли
серцевий м'яз пацієнта раптово починає самостійно генерувати імпульси,
кардіостимулятор відключається. Як тільки серце зупиняється, включається апарат.
24.
25.
Електрофізіологічні властивостіклітин скоротливого міокарда
• Рівень потенціалу спокою в скоротливих
кардіоміоцитів знаходиться в межах -90 – -95 мВ і
є стабільним. Потенціал спокою клітин
скоротливого міокарда створюється дифузією
іонів К+ із клітини та входом іонів Сl- у
кардіоміоцит, проте на відміну від фазних
поперечно-посмугованих м'язів, хлорна
проникність мембрани порівняно з калієвою дуже
мала і відіграє незначну роль у формуванні
потенціалу спокою скоротливих кардіоміоцитів.
26.
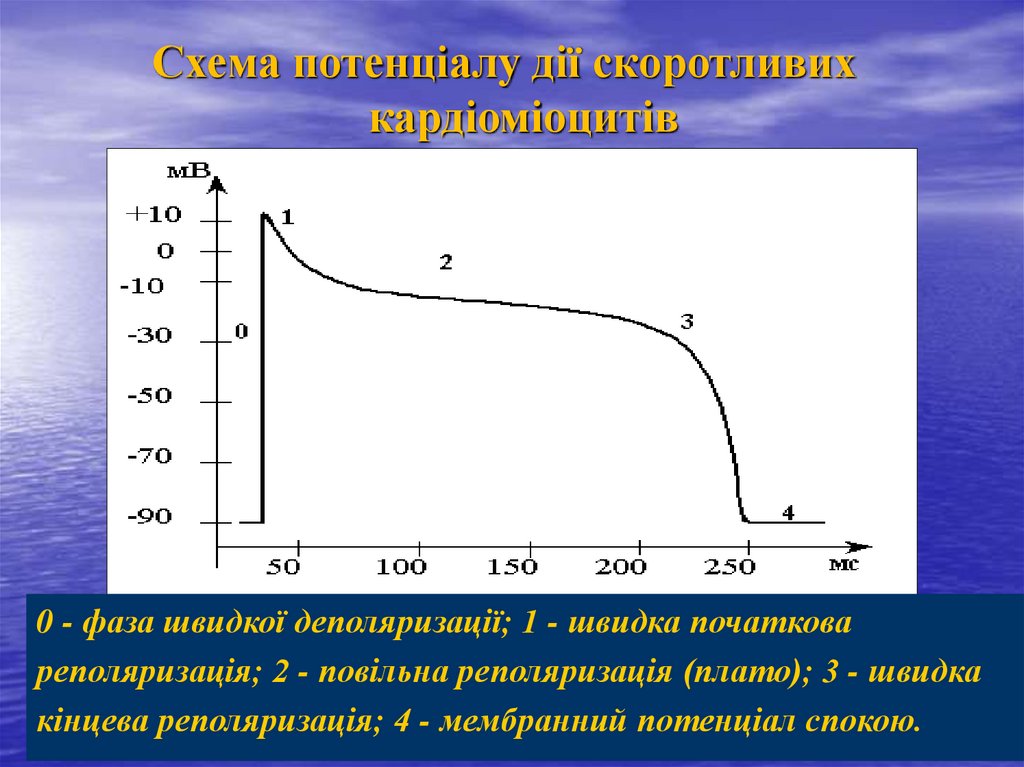
Схема потенціалу дії скоротливихкардіоміоцитів
0 - фаза швидкої деполяризації; 1 - швидка початкова
реполяризація; 2 - повільна реполяризація (плато); 3 - швидка
кінцева реполяризація; 4 - мембранний потенціал спокою.
27.
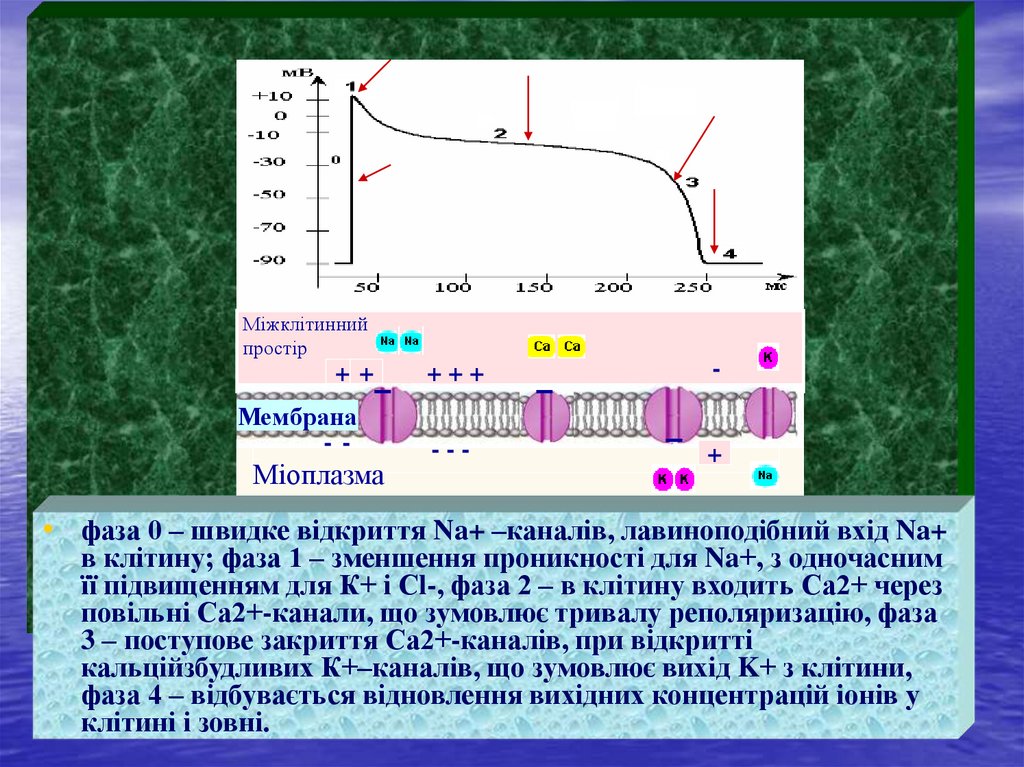
Міжклітиннийпростір
++
+++
-
Мембрана
--
---
+
Міоплазма
• фаза 0 – швидке відкриття Na+ –каналів, лавиноподібний вхід Na+
в клітину; фаза 1 – зменшення проникності для Na+, з одночасним
її підвищенням для К+ і Сl-, фаза 2 – в клітину входить Са2+ через
повільні Са2+-канали, що зумовлює тривалу реполяризацію, фаза
3 – поступове закриття Са2+-каналів, при відкритті
кальційзбудливих К+–каналів, що зумовлює вихід K+ з клітини,
фаза 4 – відбувається відновлення вихідних концентрацій іонів у
клітині і зовні.
28.
Електрофізіологічні властивостіатипових клітин
• В
клітинах
провідної
системи
відсутній
стабільний потенціал спокою, замість нього
присутня фаза повільної спонтанної діастолічної
деполяризації, яка обумовлена входом через
мембрану кардіоміоцита іонів Са2+, що веде до
нагромадження в клітині позитивно заряджених
іонів. При досягненні мембранного потенціалу
рівня критичної точки деполяризації (біля -60
мВ), спонтанно виникає потенціал дії, що
поширюється провідною системою, а звідти на
скоротливий міокард.
29.
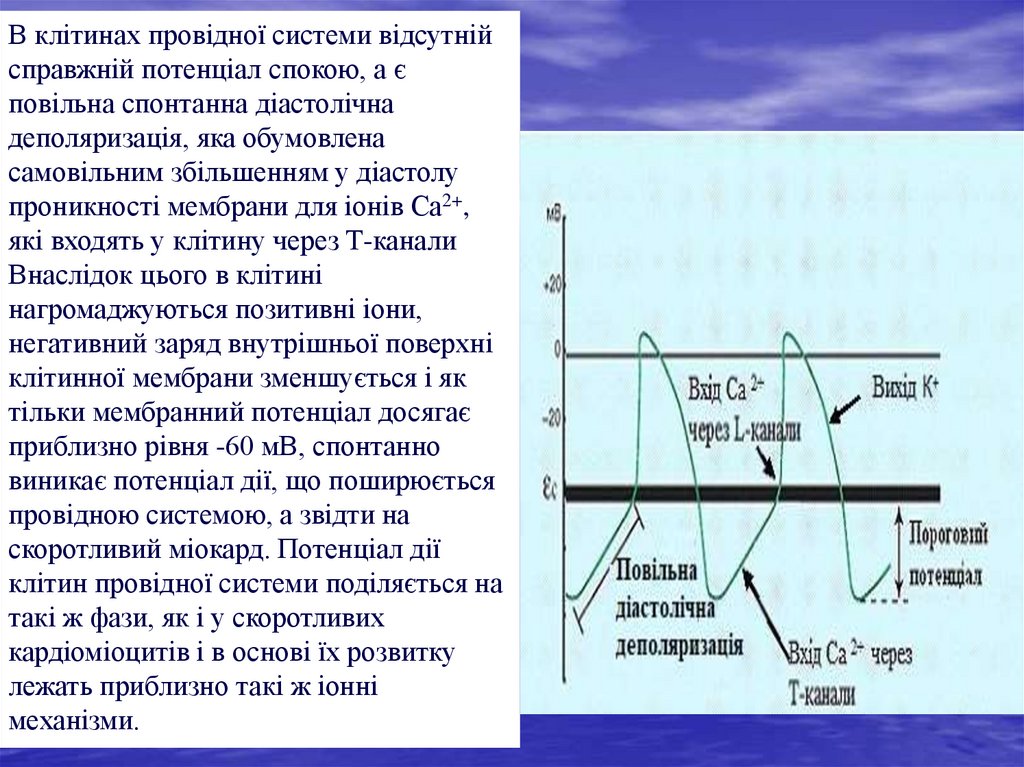
В клітинах провідної системи відсутнійсправжній потенціал спокою, а є
повільна спонтанна діастолічна
деполяризація, яка обумовлена
самовільним збільшенням у діастолу
проникності мембрани для іонів Са2+,
які входять у клітину через Т-канали
Внаслідок цього в клітині
нагромаджуються позитивні іони,
негативний заряд внутрішньої поверхні
клітинної мембрани зменшується і як
тільки мембранний потенціал досягає
приблизно рівня -60 мВ, спонтанно
виникає потенціал дії, що поширюється
провідною системою, а звідти на
скоротливий міокард. Потенціал дії
клітин провідної системи поділяється на
такі ж фази, як і у скоротливих
кардіоміоцитів і в основі їх розвитку
лежать приблизно такі ж іонні
механізми.
30. ПОТЕНЦІАЛИ ДІЇ РІЗНИХ ВІДДІЛІВ СЕРЦЯ
31. Поширення збудження в передсердях і А-В вузлі
• Поширення збудження в передсердяхЗбудження, яке виникло в сино-атріальному вузлі,
проводиться передсердями зі швидкістю 0,8–1,0 м/с.
Деполяризація охоплює раніше праве передсердя, а потім
– ліве Час охоплення збудженням обох передсердь - 0,1 с.
• Проведення збудження в передсердно-шлуночковому
вузлі
При передачі збудження із передсердь на шлуночки
спостерігається його затримка в атріо-вентрикулярному
вузлі. Вона пов'язана як з особливостями геометричної
структури вузла, так і з специфікою розвитку в ньому
електричних потенціалів, що пояснюється невеликою
щільністю Nа+ –каналів. Ця затримка має значення для
послідовного збудження і скорочення передсердь, а потім
шлуночків. Швидкість проведення збудження через атріовентрикулярний вузол становить близько 0,02 м/с.
32. Поширення збудження в шлуночках
• Швидкість проведення збудження пучком Гіса іволокнами Пуркіньє становить 1–1,5 м/с. Процес
деполяризації шлуночків починається від
середньої третини міжшлуночкової перегородки і
поширюється на верхівку і бокові стінки правого і
лівого шлуночка. Останніми деполяризуються
базальні відділи шлуночків і верхня третина
міжшлуночкової перегородки.
• Наступна затримка проведення збудження – у місці
контакту волокон Пуркіньє із скоротливими
міоцитами. Вона є наслідком сумації потенціалів
дії, що сприяє синхронізації процесу збудження
міокарду. Швидкість проведення збудження
шлуночками становить в середньому 0,3–0,9 м/с.
33.
Проведення збудження у серціМіжпередсердний
пучок Бахмана
С-В вузол
Пучки
Бахмана,
Вакенбаха,
Тореля
ЛП
Ліва ніжка
пучка Гіса
ПП
А-В вузол
Пучок
Гіса
Задня гілка
лівої ніжки
пучка Гіса
Права ніжка
пучка Гіса
Передня гілка
лівої ніжки
пучка Гіса
Перегородка
34.
• Збудливість – це здатність серця збуджуватися(або переходити в стан фізіологічноії активності).
Збудливість характерна клітинам провідної
системи серця та скоротливого міокарда.
• Зміна збудливості серця при збудженні
• Збудливість серцевого м'яза під час збудження
змінюється. Якщо співставити потенціал дії із
збудливістю, то виходить що під час 0, 1 і 2 фаз
клітина повністю незбудлива або рефрактерна. Це
так званий абсолютний рефрактерний період,
коли клітина не здатна відповісти на дію
подразника будь-якої сили і обумовлена
інактивацією Na+–каналів. Під час 3 фази має
місце відносний рефрактерний період. У цей
період надпорогове подразнення може викликати
збудження. Тобто в цей період має місце
відновлення збудливості.
35.
Співідношення між потенціалом дії, скороченнямта збудливістю скоротливого міокарду
I — потенціал дії скоротливого
кардіоміоцита (0, 1, 2, 3, 4 —
фази потенціалу дії);
II – зміни збудливості
кардіоміоцита в процесі
збудження;
а — вихідна збудливість;
б — період абсолютної
рефрактерності;
в — період відносної
рефрактерності;
г — період підвищеної
збудливості(фаза екзальтації).
36. Відмінності між збудливістю серця та скелетного м'яза
Відмінності між збудливістюсерця та скелетного м'яза
• Клітини скоротливого міокарда відрізняються від клітин
скелетних м'язів параметрами збудливості. Суттєвою
відмінністю між серцевим і скелетним м'язом є форма
потенціалу дії. Для серцевих міоцитів характерна коротка фаза
деполяризації і досить тривала фаза реполяризації. У міокарда
більш високий поріг подразливості, більш тривалий і
рефрактерний період.
Довготривалість періоду рефрактерності має надзвичайне
значення для повноцінного функціонування серця. За таких
умов, серце не може реагувати на подразнення високої
частоти, на відміну від скелетного м'яза.
37. Механізм скорочення серцевого м’яза
• Серію послідовних явищ у клітиніміокарда, що починається з
пускового механізму скорочення –
потенціалу дії мембрани з
наступними внутрішьоклітинними
процесами, які завершуються
вкороченням міофібрилів називають
спряженням збудження і скорочення.
• Структурною основою спряження
процесів збудження і скорочення
кардіоміоцитів є Т–система, яка
складається з поперечних і
повздовжніх Т-трубочок і цистерн
саркоплазматичного ретикулуму в
яких знаходиться Са2+.
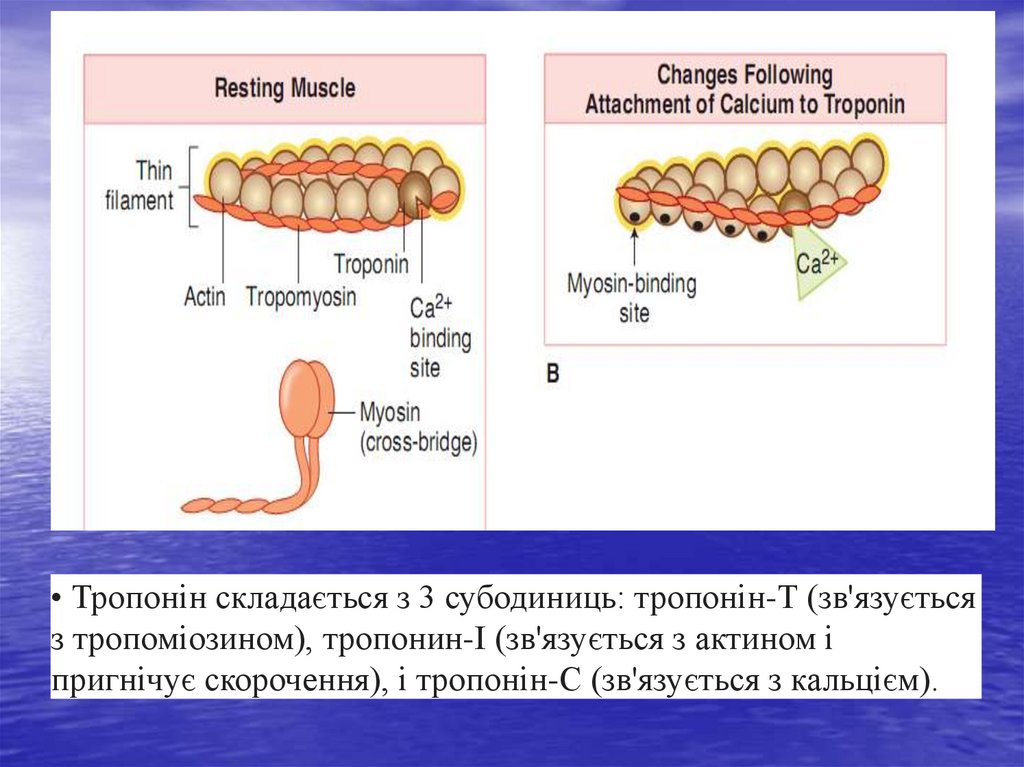
38. Структура актинових і міозинових ниток саркомера кардіоміоцита
Тонкі нитки саркомера складаються здвох скручених у спіраль ланцюгів
молекул актину, тісно пов'язаних з
регуляторними білками - тропоміозином і
тропоніном. Актин здатний утворювати
з'єднання з міозином в присутності АТФ
та іонів магнію, які активують АТФ-азу
міозину. Регуляція такого з'єднання
забезпечується головним чином
тропоніном С, який володіє високою
спорідненістю до іонів Са2+. Коли м'язове
волокно знаходиться в стані спокою і
тропонін С позбавлений іонів Са2+, весь
тропоніновий комплекс набуває такої
конформаційної структури, яка
перешкоджає взаємодії актину і міозину, і
скорочення м'язового волокна не
відбувається.
39.
• Тропонін складається з 3 субодиниць: тропонін-Т (зв'язуєтьсяз тропоміозином), тропонин-I (зв'язується з актином і
пригнічує скорочення), і тропонін-С (зв'язується з кальцієм).
40. Механізм скорочення серцевого м’яза
•Під впливом потенціалу дії,Са2+ з міжклітинного простору,
а також з цистерн
саркоплазматичного ретикулуму
поступає в міоплазму, де під
його впливом конформується
білок тропонін, який відсовує
тропоміозин від активних
центрів актину, внаслідок цього
між актином і міозином
утворюються мостики. При
цьому йде розщеплення АТФ,
енергія якого використовується
для ковзання актинових ниток.
Чим більше іонів кальцію
зв’язалось із тропоніном, тим
більше утворюється
актоміозинових мостиків і тим
більша сила скорочення м’яза.
41. Механізм розслаблення серцевого м’яза
•Розслаблення кардіоміоцита наступаєв результаті реполяризації мембрани.
Воно ґрунтується на тому, що під дією
реполяризації відбувається видалення
Са2+ від скоротливих білків з
наступним його захопленням помпами
цистерн саркоплазматичного
ретикулуму. Також Са2+ виводиться у
міжклітинну рідину за рахунок роботи
помп клітинних мембран. Основний
процес, який визначає розслаблення
кардіоміоцитів, – це видалення іонів
кальцію з саркоплазми, в результаті
чого концентрація Са2+ в ній
зменшується. При цьому комплекси
Са2+ з тропоніном С розпадаються,
тропоміозин зміщується по
відношенню до актинових філаментів і
закриває їх активні центри –
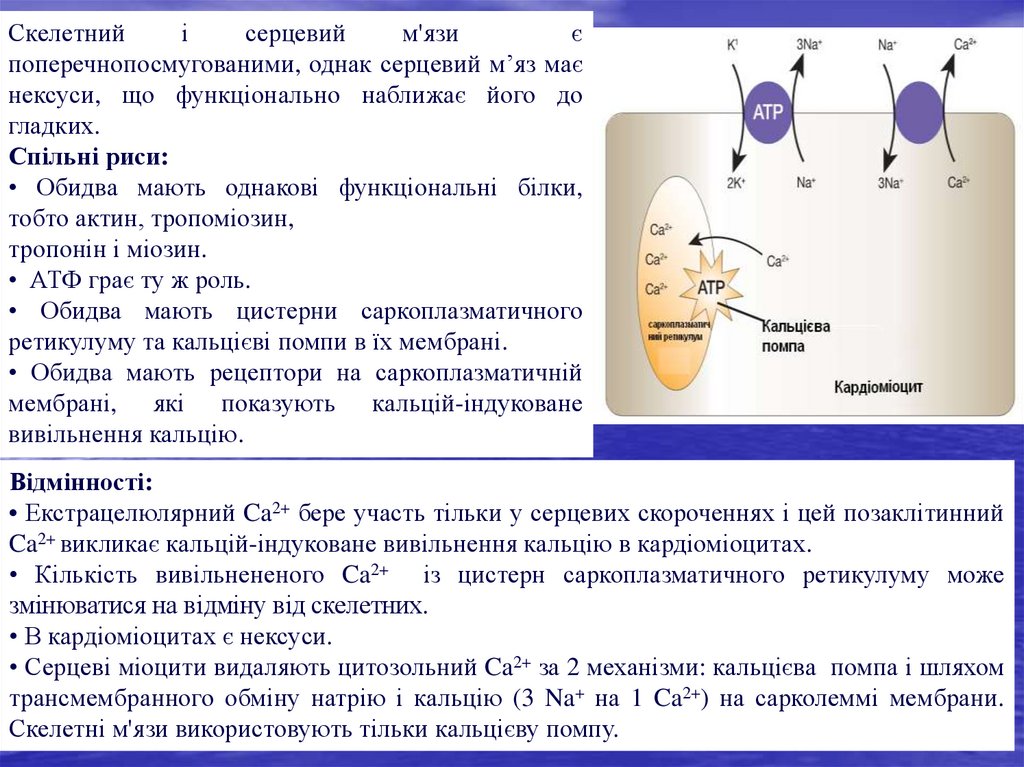
скорочення припиняється.
42.
Скелетнийі
серцевий
м'язи
є
поперечнопосмугованими, однак серцевий м’яз має
нексуси, що функціонально наближає його до
гладких.
Спільні риси:
• Обидва мають однакові функціональні білки,
тобто актин, тропоміозин,
тропонін і міозин.
• АТФ грає ту ж роль.
• Обидва мають цистерни саркоплазматичного
ретикулуму та кальцієві помпи в їх мембрані.
• Обидва мають рецептори на саркоплазматичній
мембрані, які показують кальцій-індуковане
вивільнення кальцію.
Відмінності:
• Екстрацелюлярний Ca2+ бере участь тільки у серцевих скороченнях і цей позаклітинний
Ca2+ викликає кальцій-індуковане вивільнення кальцію в кардіоміоцитах.
• Кількість вивільнененого Ca2+ із цистерн саркоплазматичного ретикулуму може
змінюватися на відміну від скелетних.
• В кардіоміоцитах є нексуси.
• Серцеві міоцити видаляють цитозольний Ca2+ за 2 механізми: кальцієва помпа і шляхом
трансмембранного обміну натрію і кальцію (3 Na+ на 1 Ca2+) на сарколеммі мембрани.
Скелетні м'язи використовують тільки кальцієву помпу.
43. Відмінності скоротливої функції міокарда і скелетного м'яза.
Відмінності скоротливої функціїміокарда і скелетного м'яза.
На відміну від скелетного м'яза в міокарді не виявлено
залежності між силою подразнення і величиною реакції на
допорогове подразнення серце зовсім не відповідає, але як
тільки сила подразника досягає порогового рівня, виникає
максимальне скорочення. Подальше наростання сили
подразника не змінює величини скорочення. Таким чином,
порогове подразнення є одночасно і максимальним. Ця
особливість скорочення серцевого м'яза отримала назву закону
"все або нічого". Його відкрив югославський фізіолог Боудічі.
•Скелетний м'яз відповідає градуально, тобто, чим більша
сила подразнення, тим більша сила скорочення. Оскільки
період рефрактерності надзвичайно тривалий у міокарді, то це
попереджує виникнення стійкого скорочення, а отже зупинки
серця в систолі, тоді, як часті повторні збудження скелетного
м'яза ведуть до тетанусу.