


























 chemistry
chemistrySimilar presentations:






")



Структура и функции нуклеиновых кислот. Лекция №15
1.
Лекция № 15Структура и функции
нуклеиновых кислот
2.
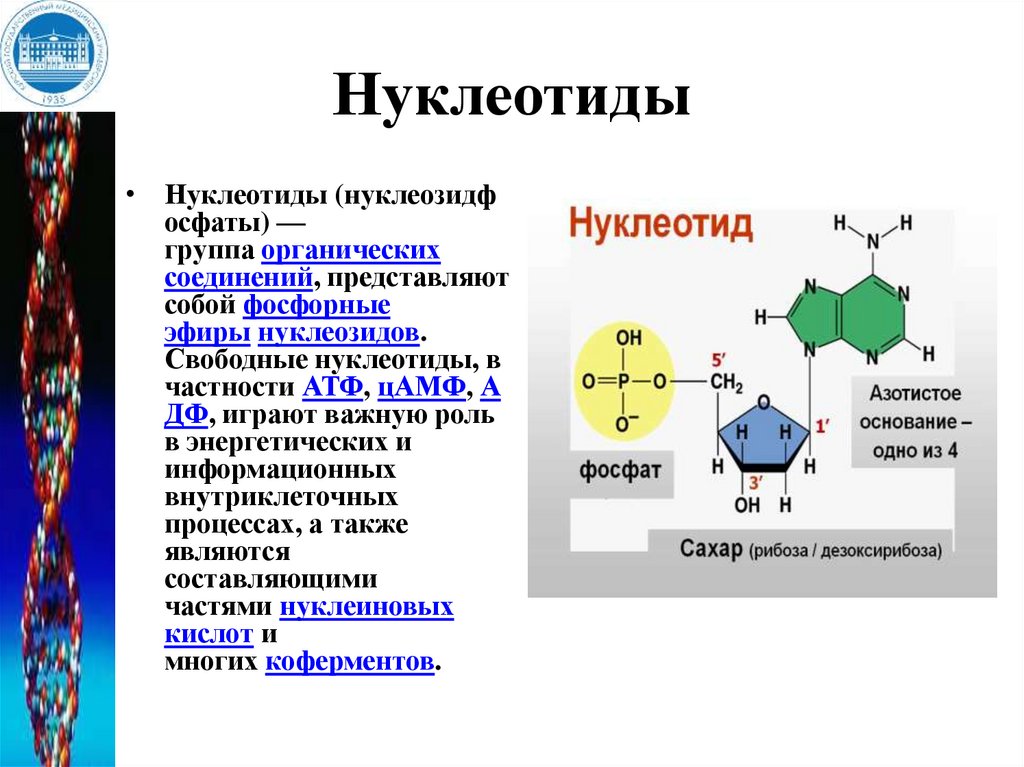
Нуклеотиды• Нуклеотиды (нуклеозидф
осфаты) —
группа органических
соединений, представляют
собой фосфорные
эфиры нуклеозидов.
Свободные нуклеотиды, в
частности АТФ, цАМФ, А
ДФ, играют важную роль
в энергетических и
информационных
внутриклеточных
процессах, а также
являются
составляющими
частями нуклеиновых
кислот и
многих коферментов.
3.
Общая характеристикаазотистых оснований:
1. по строению:
• пуриновые аденин (А) и
гуанин (Г),
• пиримидиновые цитозин (Ц) и
урацил (У) в РНК
и цитозин (Ц) и
тимин (Т) – в
ДНК.
2. по
распространению и
степени
встречаемости:
• мажорные
(основные,
преобладающие)
• минорные (редко
встречающиеся).
4.
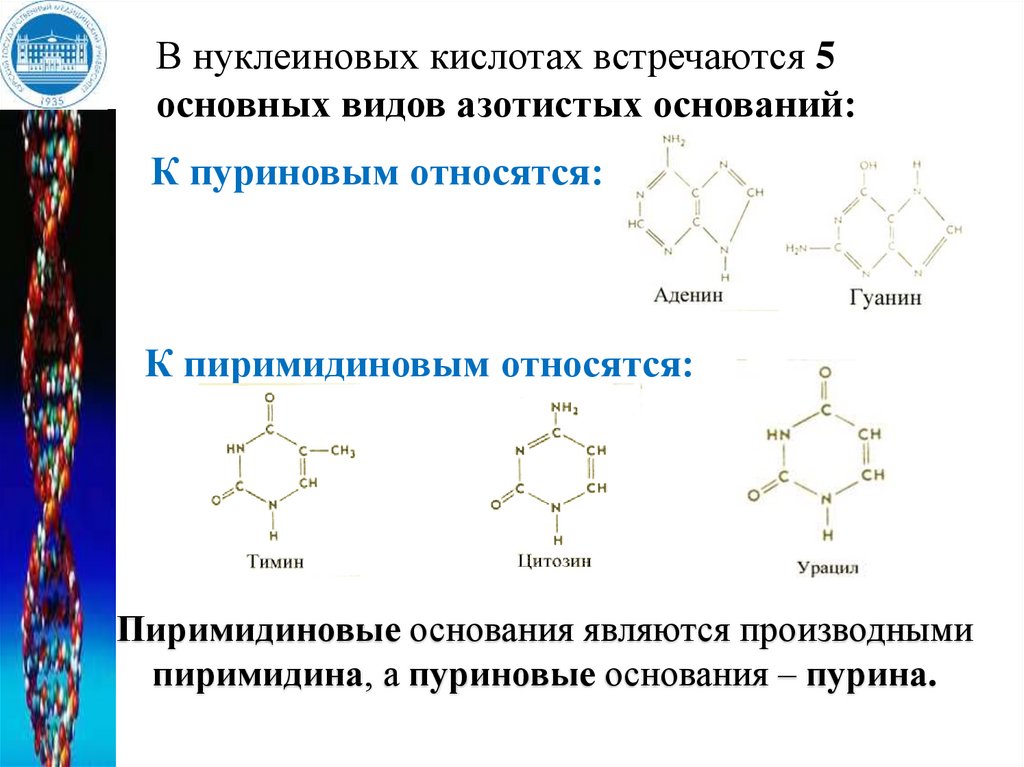
В нуклеиновых кислотах встречаются 5основных видов азотистых оснований:
К пуриновым относятся:
К пиримидиновым относятся:
Пиримидиновые основания являются производными
пиримидина, а пуриновые основания – пурина.
5.
Биологическая роль нуклеотидов:Мономер ДНК и РНК.
Форма запасания энергии в клетке (например, АТФ, ГТФ).
Источник фосфатной группы (например, при
фосфорилировании глюкозы).
4. Коферментная функция (НАД, НАДФ, ФАД).
5. Могут служить активаторами ферментов (это, в основном
характерно для цАМФ и цГМФ).
6. Для активирования различных субстратов, в результате чего
соединения с мононуклеотидом они приобретают
макроэргическую связь и могут вступать в химические
реакции:
а) для активирования глюкозы – необходим УТФ, получается УДФглюкоза;
б) для холина и других азотистых соединений, а также
глицеролсодержащих соединений – ЦТФ, получается ЦДФ-холин и
др.;
в) для аминокислот – АТФ, получается аминоацил-аденилат.
7. Нуклеотиды могут выполнять регуляторную функцию
(например, гуанозинтетрафосфат участвует в матричных
синтезах).
1.
2.
3.
6.
РНК и ДНК• В клетке имеются два типа нуклеиновых кислот:
рибонуклеиновая кислота (РНК) и
дезоксирибонуклеиновая кислота (ДНК).
• ДНК и РНК – полимерные цепочки, состоящие из
отдельных звеньев – нуклеотидов.
www.themegallery.com
7.
8.
9.
Синтез пиримидиновых нуклеотидов• Синтез пиримидиновых оснований
происходит во всех клетках организма. В
реакциях синтеза участвует
аспарагиновая кислота, глутамин, СО2,
затрачивается 2 молекулы АТФ. В
отличие от разветвленного синтеза
пуринов этот синтез происходит линейно,
т.е. пиримидиновые
нуклеотиды образуются последовательн
о, друг за другом.
• Условно можно выделить 3 общих этапа
синтеза и реакции синтеза УТФ и ЦТФ:
10.
Этапы синтеза пиримидиновыхнуклеотидов
1. Образование карбамоилфосфата
• Образование карбамоилфосфата в
отличие от синтеза мочевины
происходит в цитозоле большинства
клеток организма.
2. Образование пиримидинового кольца
• Формирование пиримидинового
кольца происходит после
присоединения аспартата и реакций
дегидратации и окисления. Первым
пиримидиновым основанием
является оротовая кислота.
11.
Синтез пиримидиновых нуклеотидов3. Синтез
оротидинмонофосфата и
уридинмонофосфорной
кислоты
• В реакции
с фосфорибозилдифосфатом
(ФРДФ) к оротовой кислоте
присоединяется рибозо-5фосфат и образуется
оротидилмонофосфат,
при декарбоксилировании
превращающийся в
уридинмонофосфат (УМФ).
• Источником
фосфорибозилдифосфата
является первая из двух
реакций синтеза
фосфорибозиламина при
образовании пуринов.
12.
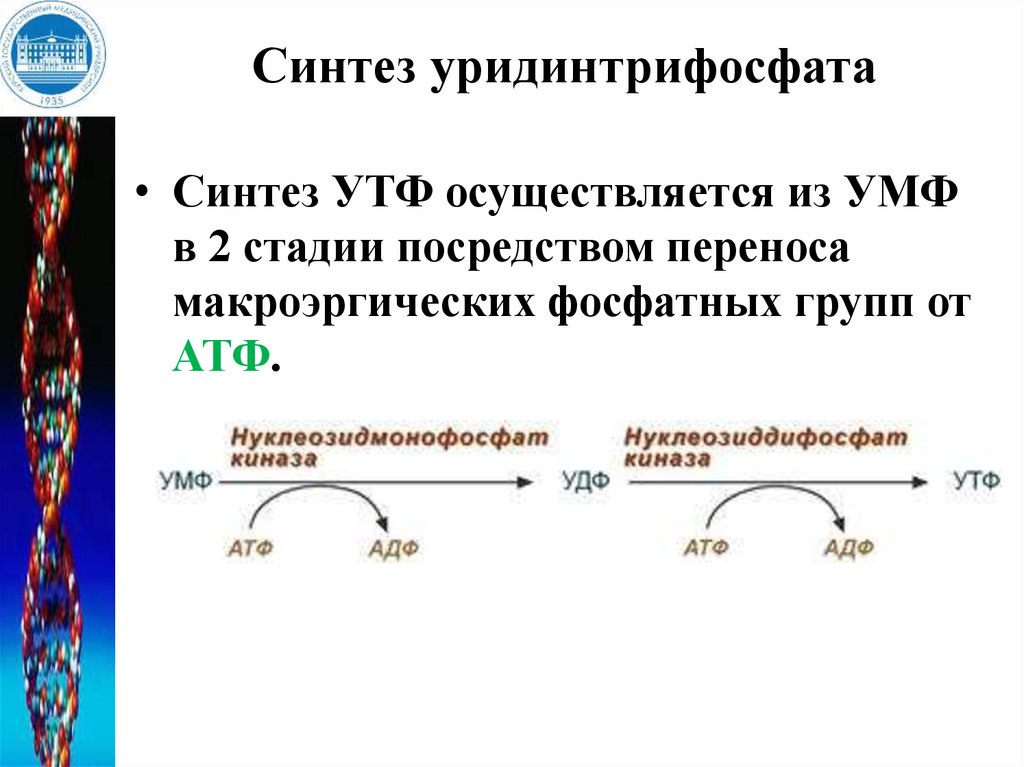
Синтез уридинтрифосфата• Синтез УТФ осуществляется из УМФ
в 2 стадии посредством переноса
макроэргических фосфатных групп от
АТФ.
13.
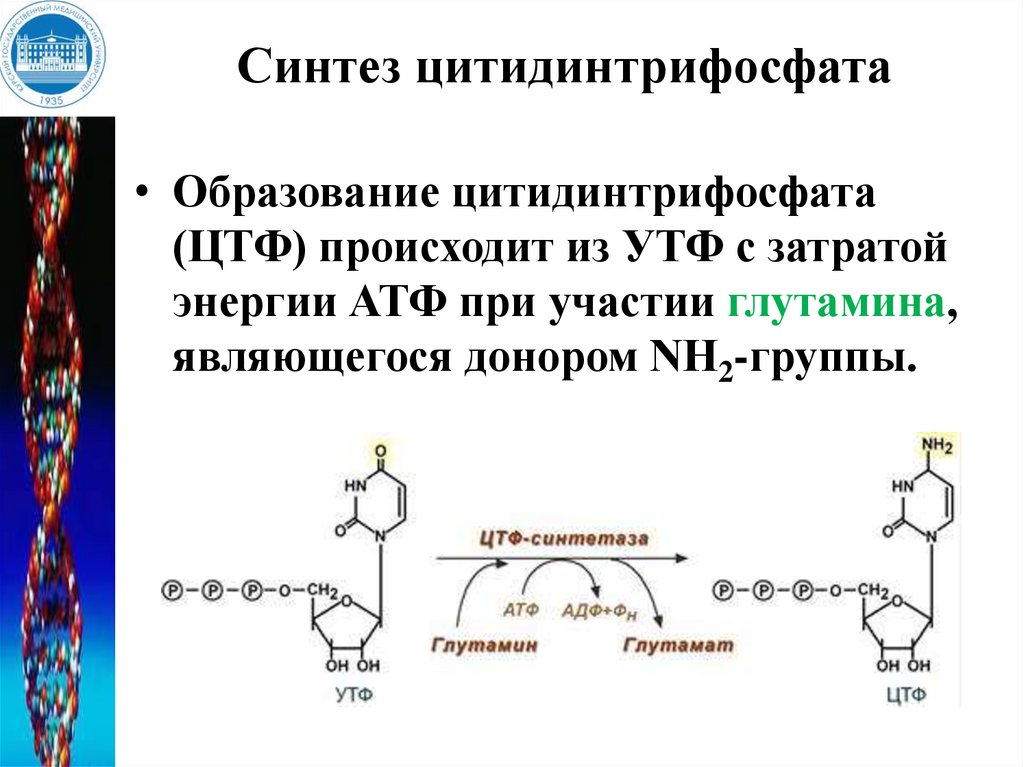
Синтез цитидинтрифосфата• Образование цитидинтрифосфата
(ЦТФ) происходит из УТФ с затратой
энергии АТФ при участии глутамина,
являющегося донором NH2-группы.
14.
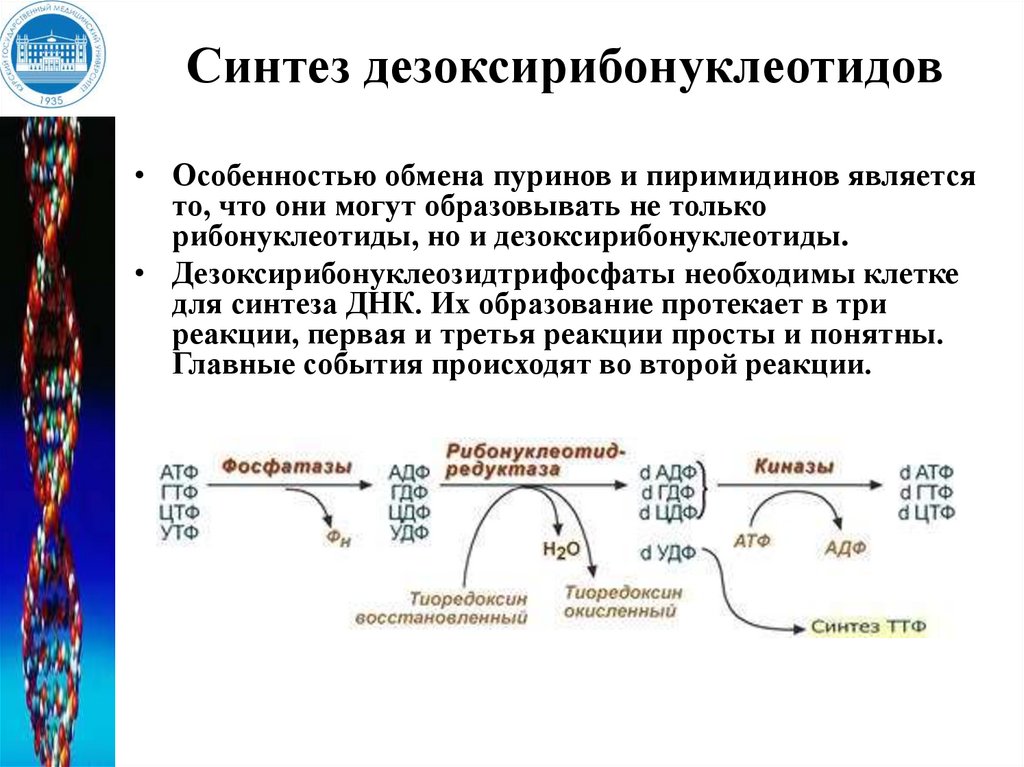
Синтез дезоксирибонуклеотидов• Особенностью обмена пуринов и пиримидинов является
то, что они могут образовывать не только
рибонуклеотиды, но и дезоксирибонуклеотиды.
• Дезоксирибонуклеозидтрифосфаты необходимы клетке
для синтеза ДНК. Их образование протекает в три
реакции, первая и третья реакции просты и понятны.
Главные события происходят во второй реакции.
15.
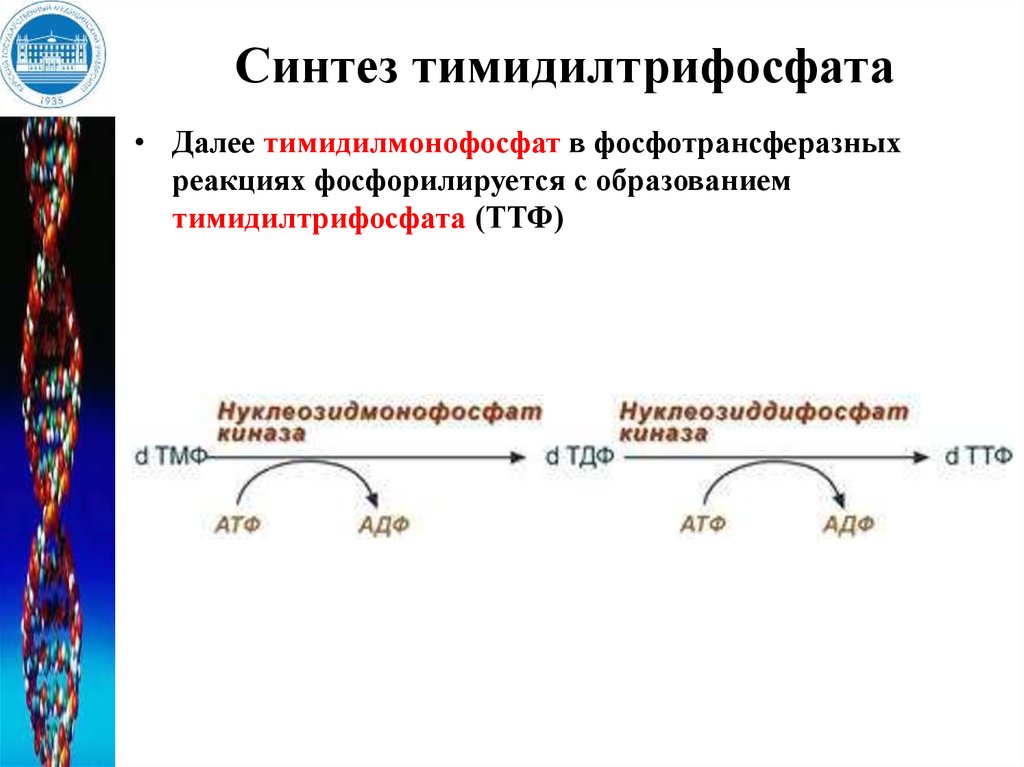
Синтез тимидилтрифосфата• Далее тимидилмонофосфат в фосфотрансферазных
реакциях фосфорилируется с образованием
тимидилтрифосфата (ТТФ)
16.
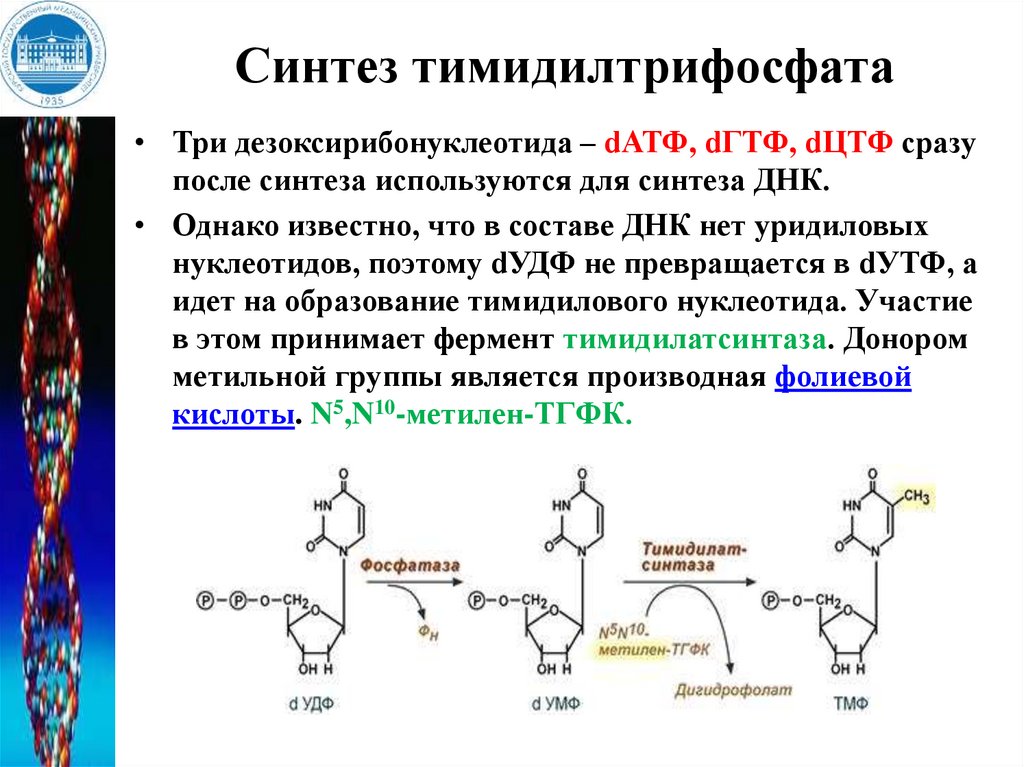
Синтез тимидилтрифосфата• Три дезоксирибонуклеотида – dАТФ, dГТФ, dЦТФ сразу
после синтеза используются для синтеза ДНК.
• Однако известно, что в составе ДНК нет уридиловых
нуклеотидов, поэтому dУДФ не превращается в dУТФ, а
идет на образование тимидилового нуклеотида. Участие
в этом принимает фермент тимидилатсинтаза. Донором
метильной группы является производная фолиевой
кислоты. N5,N10-метилен-ТГФК.
17.
Механизм противомикробногодействия сульфаниламидов
• Целая группа антибактериальных препаратов способна
подавлять синтез ТТФ в микробных клетках
благодаря структурной схожести пара-аминобензойной
кислоты (компонента ТГФК) и вещества сульфаниламида.
• При синтезе фолиевой кислоты (витамин В9) микробной
клеткой сульфаниламид конкурирует за активные центры
фермента с пара-аминобензойной кислотой, что нарушает
синтез фолатов бактерией. В результате снижается
количество ТГФК в клетке, снижается образование
тимидилтрифосфата, тормозится синтез ДНК и
размножение микроорганизмов.
• При терапии побочные
эффекты сульфаниламидов
также связаны с замедлением
синтеза ДНК.
18.
Ингибиторы синтезатимидилового нуклеотида
• Исключительная роль тимидилатсинтазы и фолиевой
кислоты в синтезе ТМФ обусловила поиск и использование
ингибиторов этой реакции для прекращения синтеза
тимидилового нуклеотида и, как следствие, блока синтеза
ДНК, что нашло применение в противоопухолевой терапии
("бестиминовая смерть").
• Фторурацил – противоопухолевое (цитостатическое)
средство, механизм его действия определяется
превращением в 5-фтор-производные dУМФ и dУТФ,
конкурентно ингибирующих фермент тимидилатсинтазу,
что ведет к снижению репликации ДНК из-за недостатка
ТМФ. Метотрексат, аминоптерин, триметоприм являются
ингибиторами дигидрофолатредуктазы, что препятствует
ресинтезу ТГФК и лишает реакцию кофермента.
19.
Распад пиримидиновых нуклеотидовРаспад пиримидиновых нуклеотидов происходит параллельно, с
использованием одинаковых реакций и ферментов. Можно
отметить несколько специфичных ферментов:
1. Фермент 5'-нуклеотидаза отщепляет 5'-фосфатную группу от
ЦМФ, УМФ и ТМФ .
2. Дезаминаза проводит окислительное дезаминирование
цитидина.
3. Нуклеозид-фосфорилаза удаляет рибозу от уридина и
тимидина.
4. Дигидроурацил-дегидрогеназа – восстановление урацила и
тимина.
5. Дигидропиримидиназа осуществляет гидролитическое
расщепление пиримидинового кольца.
6. Далее происходит гидролитическое отщепление аммиака и
углекислого газа.
7. После окончательного разрушения пиримидинового кольца
появившиеся β-аминокислоты направляются в
реакции трансаминирования, после чего соответствующие
кетокислоты изомеризуются и далее сгорают в ЦТК.
20.
Нарушения обмена пиримидинов• β-Аминоизобутиратацидурия
• Так как продукты катаболизма пиримидинов
хорошо растворимы, то они хорошо
выводятся с мочой и клинические симптомы
патологий слабо выражены. Примером
подобного нарушения является βаминоизобутиратацидурия, т.е. накопление βаминоизомасляной кислоты, конечного
продукта распада тиминового кольца, при
которой имеется дефект соответствующей
трансаминазы. Заболевание протекает
бессимптомно и встречается, в основном, у
японцев и китайцев (около 25%).
21.
ОротатацидурияНаследственная -
связана с одновременным
дефектом оротатфосфорибозилтрансферазы и оротидилдекарбокс
илазы (ферменты синтеза УМФ),
так как эти ферменты являются
двумя активными центрами
одного белка. Заболевание
проявляется на втором полугодии
жизни лейкопенией и гипохромно
й мегалобластической анемией,
появляется сонливость, вялость, о
тставание умственного и
физического развития. Диагноз
ставится при обнаружении в
охлажденной моче оранжевых
игольчатых кристаллов оротовой
кислоты (болезнь оранжевых
пеленок).
Приобретенная - при
дефекте каких-либо ферментов
синтеза мочевины, кроме
карбамоилфосфатсинтетазы. При
этом карбамоилфосфат
митохондрий (в норме
используемый для образования
мочевины) выходит из них и
используется для избыточного
синтеза оротовой кислоты.
Заболевание обычно
сопровождается гипераммониеми
ей, при лечении
подагры аллопуринолом,
который может превращаться в
оксипуринолмононуклеотид,
являющийся
ингибитором оротатдекарбоксила
зы, что опять же ведет к
накоплению оротата.
22.
Синтез пуриновых нуклеотидов1 этап Синтез 5'-фосфорибозиламина
Первая реакция синтеза пуринов заключается в
активации углерода в положении С1 рибозо-5-фосфата,
это достигается синтезом 5-фосфорибозил-1дифосфата (ФРДФ). Фосфорибозил-дифосфат
является тем якорем, на основе которого синтезируется
сложный пуриновый цикл.
Вторая реакция – это перенос NH2-группы глутамина
на активированный атом С1 с образованием 5'фосфорибозиламина. Указанная NH2-группа
фосфорибозиламина уже принадлежит будущему
пуриновому кольцу и ее азот будет атомом номер 9.
23.
Синтез пуриновых нуклеотидов• 2 этап Синтез
инозинмонофосфата
• 5-фосфорибозиламин вовлекается
в девять реакций, и в результате
образуется первый пуриновый
нуклеотид –
инозинмонофосфорная
кислота (ИМФ). В этих реакциях
источниками атомов пуринового
кольца
являются глицин, аспартат,
• еще одна
молекула глутамина, углекислый
газ и
производные тетрагидрофолиево
й кислоты (ТГФК). В целом на
синтез пуринового кольца
затрачивается энергия 6 молекул
АТФ.
24.
Синтез пуриновых нуклеотидов3 этап Синтез аденозинмонофосфата
и гуанозинмонофосфата
Гуанозинмонофосфат (ГМФ)
образуется в двух реакциях –
сначала ИМФ окисляется ИМФдегидрогеназой до
ксантозилмонофосфата, источником
кислорода является вода,
акцептором водорода – НАД. После
этого работает ГМФ-синтетаза, она
использует универсальный
клеточный донор NH2-групп –
глутамин, источником энергии для
реакции служит АТФ.
Аденозинмонофосфат (АМФ) также
образуется в двух реакциях, но в
качестве донора NH2-группы
выступает аспарагиновая кислота. В
первой, аденилосукцинатсинтетазной, реакции на
присоединение аспартата
используется энергия распада ГТФ,
во второй
реакции аденилосукцинатлиаза производит удаление части
аспарагиновой кислоты в виде
фумарата.
25.
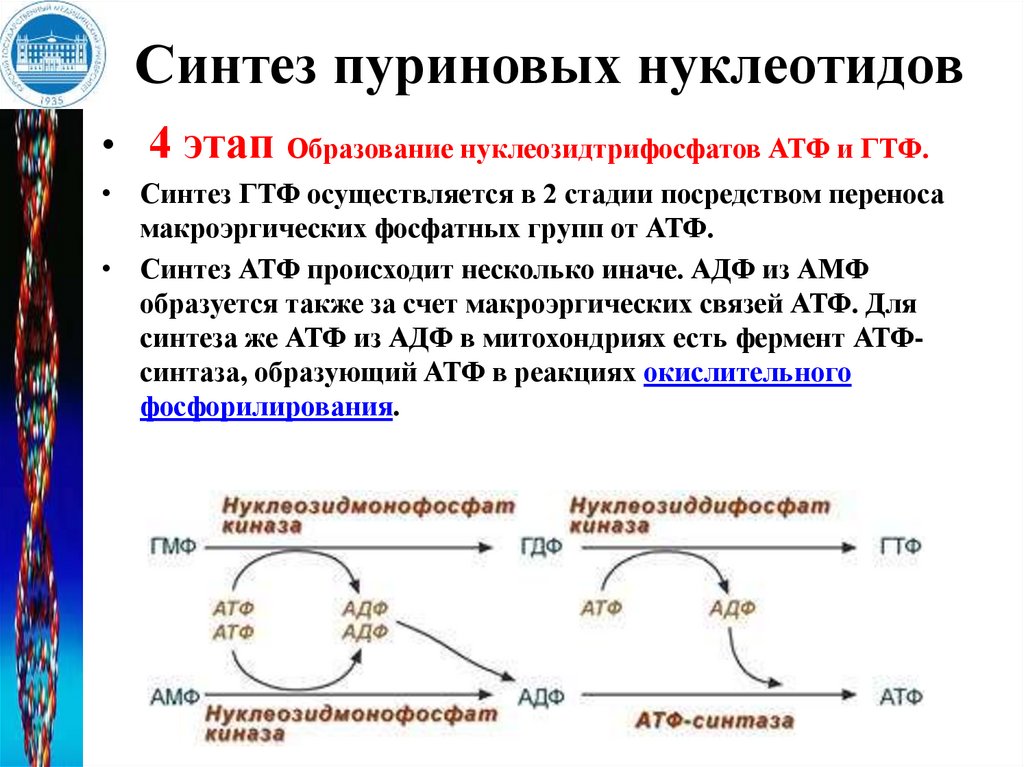
Синтез пуриновых нуклеотидов• 4 этап Образование нуклеозидтрифосфатов АТФ и ГТФ.
• Синтез ГТФ осуществляется в 2 стадии посредством переноса
макроэргических фосфатных групп от АТФ.
• Синтез АТФ происходит несколько иначе. АДФ из АМФ
образуется также за счет макроэргических связей АТФ. Для
синтеза же АТФ из АДФ в митохондриях есть фермент АТФсинтаза, образующий АТФ в реакциях окислительного
фосфорилирования.
26.
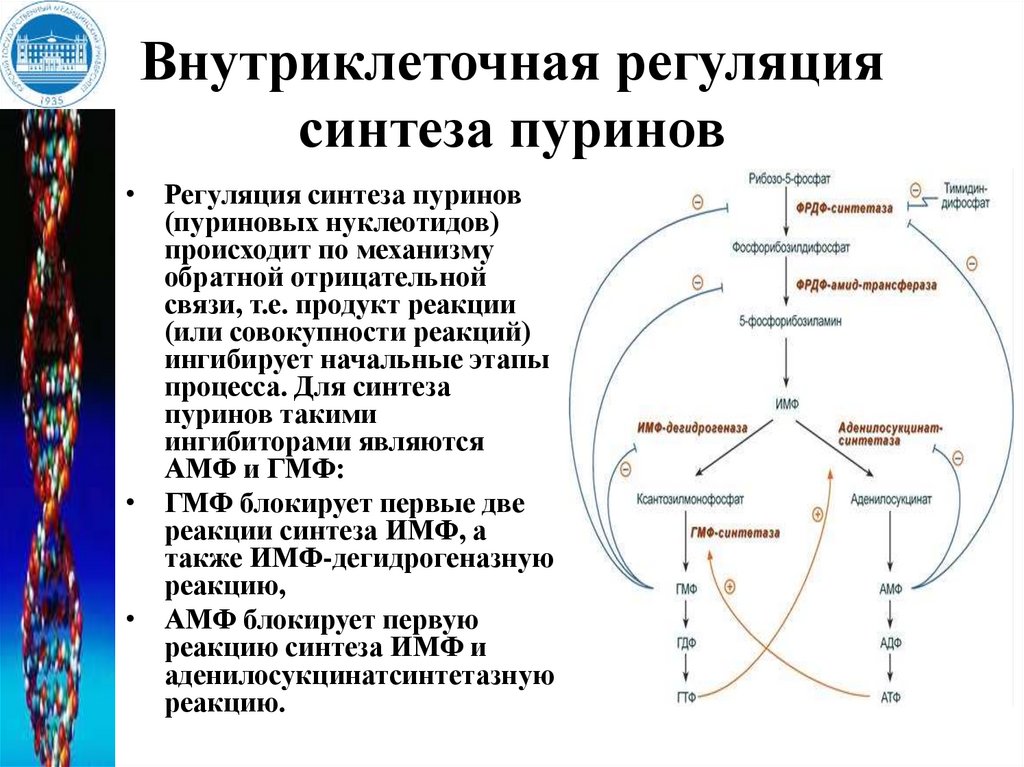
Внутриклеточная регуляциясинтеза пуринов
• Регуляция синтеза пуринов
(пуриновых нуклеотидов)
происходит по механизму
обратной отрицательной
связи, т.е. продукт реакции
(или совокупности реакций)
ингибирует начальные этапы
процесса. Для синтеза
пуринов такими
ингибиторами являются
АМФ и ГМФ:
• ГМФ блокирует первые две
реакции синтеза ИМФ, а
также ИМФ-дегидрогеназную
реакцию,
• АМФ блокирует первую
реакцию синтеза ИМФ и
аденилосукцинатсинтетазную
реакцию.
27.
Внутриклеточная регуляциясинтеза пуринов
Кроме этого,
имеется положительная
перекрестная регуляция со
стороны АТФ и ГТФ, а именно –
как участник реакций каждый из
них оказывает стимулирующее
влияние на синтез другого
нуклеотида. Это заключается в
том, что АТФ, принимая участие в
ГМФ-синтетазной реакции,
облегчает синтез ГМФ. В свою
очередь, ГТФ является донором
энергии для синтеза АМФ,
участвуя в аденилосукцинатсинтетазной реакции.
Пиримидиновый
нуклеотид тимидиндифосфат (ТДФ
) также блокирует фермент ФРДФсинтетазу, что снижает синтез
ФРДФ, который используется при
синтезе не только пуринов, но и
пиримидинов.
28.
Лекарственная регуляция• При разработке новых
противоопухолевых средств были
предложены ингибиторы ферментов:
• ФРДФ-амидтрансферазы –
азосерин, диазонорлейцин,
• ИМФ-дегидрогеназы – микофеноловая
кислота, 6-меркаптопурин,
• аденилосукцинат-лиазы – 6меркаптопурин.
29.
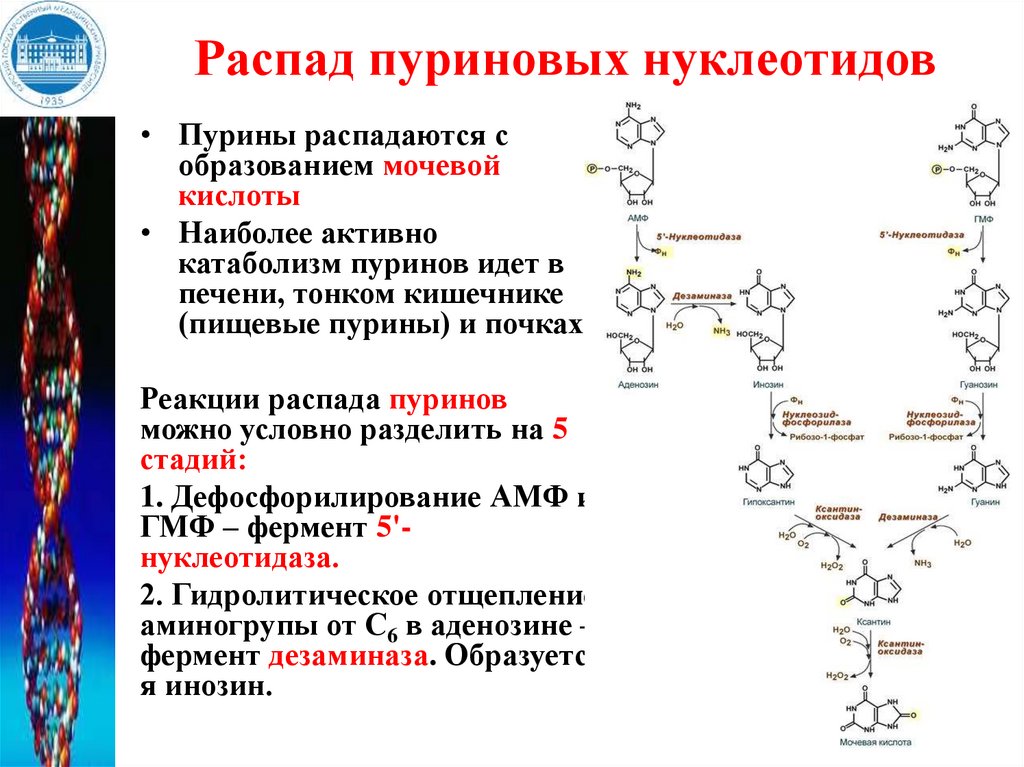
Распад пуриновых нуклеотидов• Пурины распадаются с
образованием мочевой
кислоты
• Наиболее активно
катаболизм пуринов идет в
печени, тонком кишечнике
(пищевые пурины) и почках.
Реакции распада пуринов
можно условно разделить на 5
стадий:
1. Дефосфорилирование АМФ и
ГМФ – фермент 5'нуклеотидаза.
2. Гидролитическое отщепление
аминогрупы от С6 в аденозине –
фермент дезаминаза. Образуетс
я инозин.
30.
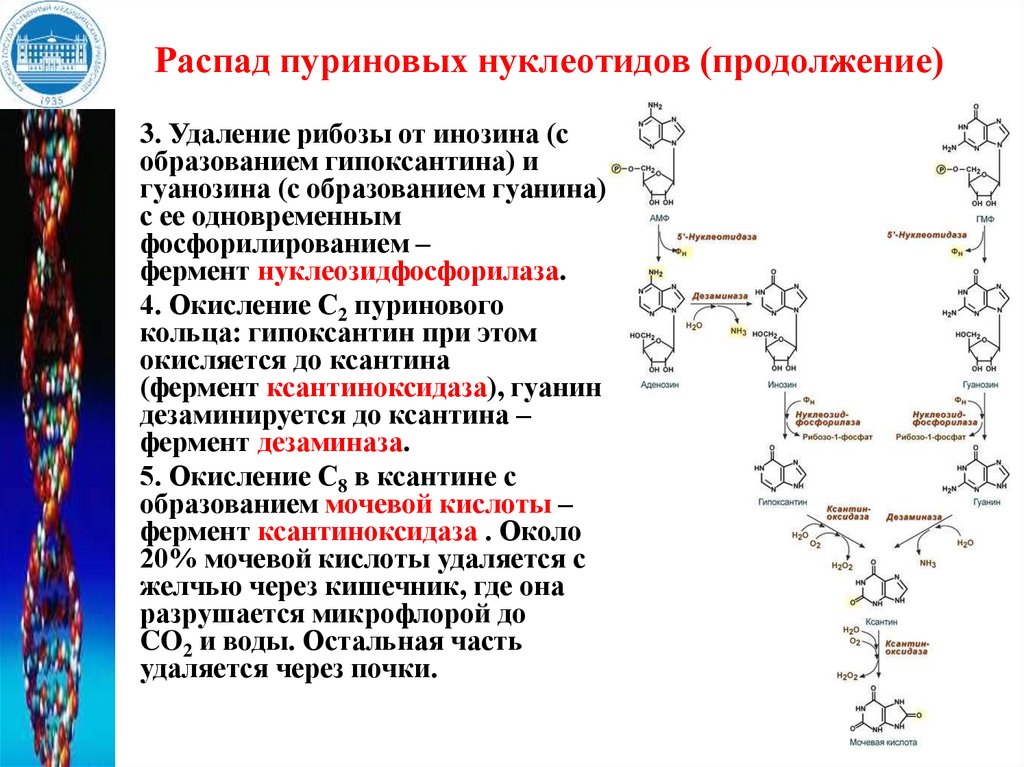
Распад пуриновых нуклеотидов (продолжение)3. Удаление рибозы от инозина (с
образованием гипоксантина) и
гуанозина (с образованием гуанина)
с ее одновременным
фосфорилированием –
фермент нуклеозидфосфорилаза.
4. Окисление С2 пуринового
кольца: гипоксантин при этом
окисляется до ксантина
(фермент ксантиноксидаза), гуанин
дезаминируется до ксантина –
фермент дезаминаза.
5. Окисление С8 в ксантине с
образованием мочевой кислоты –
фермент ксантиноксидаза . Около
20% мочевой кислоты удаляется с
желчью через кишечник, где она
разрушается микрофлорой до
CO2 и воды. Остальная часть
удаляется через почки.
31.
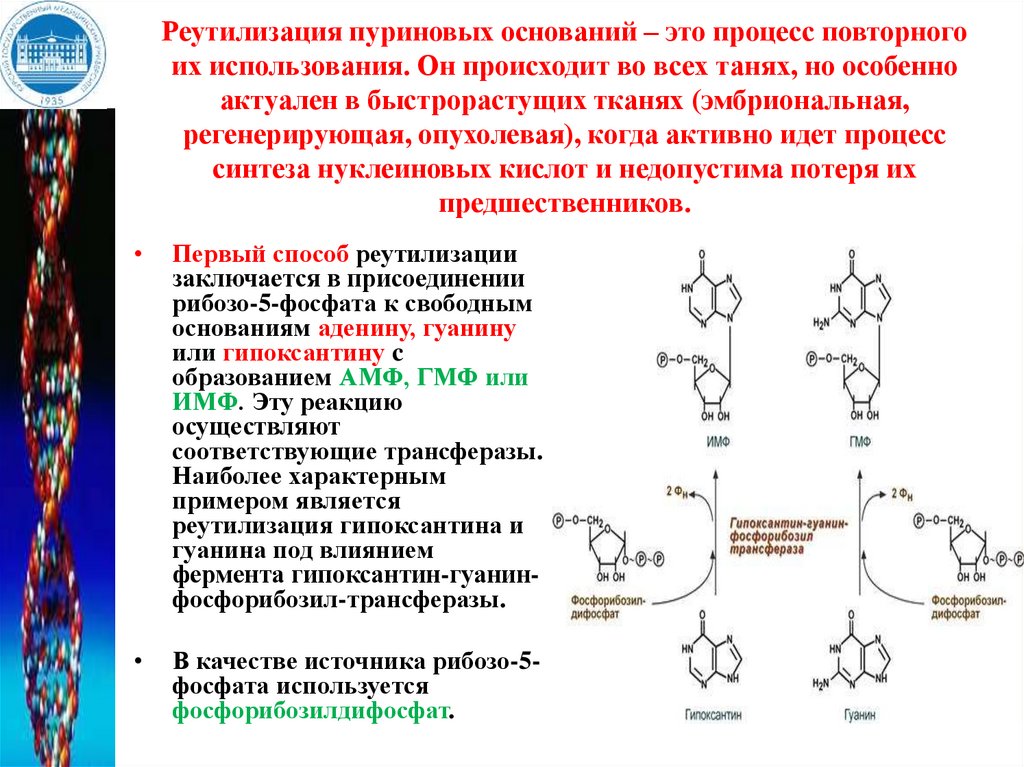
Реутилизация пуриновых оснований – это процесс повторногоих использования. Он происходит во всех танях, но особенно
актуален в быстрорастущих тканях (эмбриональная,
регенерирующая, опухолевая), когда активно идет процесс
синтеза нуклеиновых кислот и недопустима потеря их
предшественников.
Первый способ реутилизации
заключается в присоединении
рибозо-5-фосфата к свободным
основаниям аденину, гуанину
или гипоксантину c
образованием АМФ, ГМФ или
ИМФ. Эту реакцию
осуществляют
соответствующие трансферазы.
Наиболее характерным
примером является
реутилизация гипоксантина и
гуанина под влиянием
фермента гипоксантин-гуанинфосфорибозил-трансферазы.
В качестве источника рибозo-5фосфата используется
фосфорибозилдифосфат.
32.
Реутилизация пуриновых оснований – это процесс повторногоих использования. Он происходит во всех танях, но особенно
актуален в быстрорастущих тканях (эмбриональная,
регенерирующая, опухолевая), когда активно идет процесс
синтеза нуклеиновых кислот и недопустима потеря их
предшественников.
• Второй способ реутилизируются
пуриновые рибонуклеозиды
и дезоксирибонуклеозиды.
Для этого существуют
ферменты с групповой
специфичностью аденозинк
иназа (для аденозина,
гуанозина, инозина и их
дезоксиформ)
и дезоксицитидинкиназа,
которая фосфорилирует
дезоксигуанозин и
дезоксиаденозин с
образованием dГМФ и
dАМФ, и дезоксицитидин до
dЦМФ.
33.
Накопление мочевой кислоты• Самым частым нарушением обмена пуринов
является повышенное образование мочевой кислоты с
развитием гиперурикемии. Особенностью является то, что
растворимость солей мочевой кислоты (уратов) в плазме
крови невелика и при превышении порога растворимости в
плазме (около 0,7 ммоль/л) они кристаллизуются в
периферических зонах с пониженной температурой.
• В зависимости от длительности и
тяжести гиперурикемия проявляется:
• Появление тофусов (греч. tophus – пористый камень, туф) –
отложение уратных кристаллов в коже и подкожных слоях, в
мелких суставах ног и рук, в сухожилиях, хрящах, костях и
мышцах.
• Нефропатии в результате кристаллизации мочевой кислоты
с поражением почечных канальцев и мочекаменная болезнь.
• Подагра – поражение мелких суставов.
• Для диагностики нарушений используют определение
концентрации мочевой кислоты в крови и моче.
34.
Подагра (греч. poclos – нога, agra –захват, дословно – "нога в капкане")
• В крови мочевая кислота находится в форме ее солей – уратов
натрия. Из-за низкой растворимости ураты способны оседать в
зонах с пониженной температурой, например, в мелких суставах
стоп и пальцев ног. Накапливающиеся в межклеточном
веществе ураты некоторое время фагоцитируются, но фагоциты
не способны разрушить пуриновое кольцо. В результате это
приводит к гибели самих фагоцитов, к выходу лизосомальных
ферментов, активации свободнорадикального окисления и
развитию острой воспалительной реакции –
развивается подагрический артрит. В 50-75% случаев первым
признаком заболевания является мучительная ночная боль в
больших пальцах ног.
35.
Подагра (греч. poclos – нога, agra –захват, дословно – "нога в капкане")
Длительное время подагру считали "болезнью гурманов",
однако затем внимание исследователей переместилось к
наследственному изменению активности ферментов
метаболизма пуринов:
увеличение активности ФРДФ-синтетазы – приводит к
избыточному синтезу пуринов,
уменьшение активности гипоксантин-гуанин-фосфорибозилтрансферазы – из-за этого ФРДФ не используется
для реутилизации пуриновых оснований, а участвует в первой
реакции их синтеза. В результате возрастает количество
разрушающихся пуринов и одновременно повышается их
образование.
Оба ферментативных нарушения рецессивны и сцеплены с Xхромосомой. Подагрой страдает 0,3-1,7% взрослого населения
земного шара, соотношение заболевших мужчин и женщин
составляет 20 : 1.
36.
Лечение подагры• Диета – снижение поступления предшественников мочевой
кислоты с пищей и уменьшение ее образования в организме.
Для этого из рациона исключаются продукты, содержащих
много пуриновых оснований – пиво, кофе, чай, шоколад,
мясные продукты, печень, красное вино. Предпочтение
отдается вегетарианской диете с количеством чистой воды не
менее 2 л в сутки.
• К лекарственным средствам лечения подагры
относят аллопуринол, по структуре схожий с гипоксантином.
Ксантиноксидаза окисляет аллопуринол в аллоксантин, и
последний остается прочно связанным с активным центром
фермента и ингибирует его. Фермент осуществляет, образно
говоря, самоубийственный катализ. Как следствие, ксантин
не превращается в мочевую кислоту, и поскольку
гипоксантин и ксантин лучше растворимы в воде, то они
более легко выводятся из организма с мочой.
37.
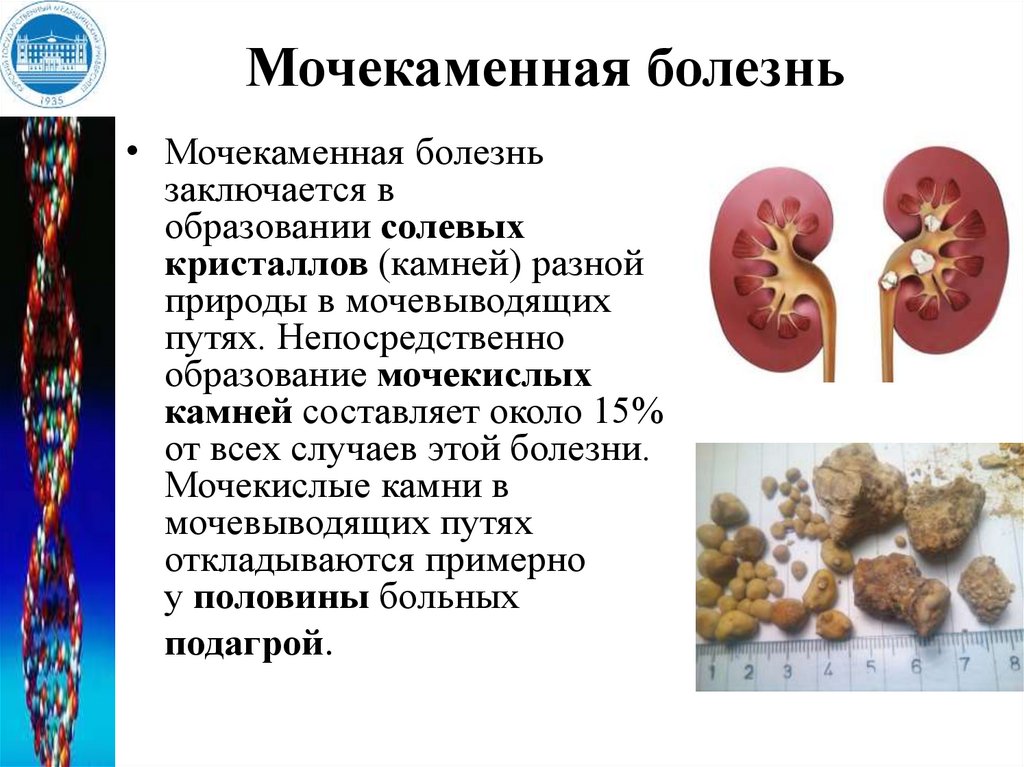
Мочекаменная болезнь• Мочекаменная болезнь
заключается в
образовании солевых
кристаллов (камней) разной
природы в мочевыводящих
путях. Непосредственно
образование мочекислых
камней составляет около 15%
от всех случаев этой болезни.
Мочекислые камни в
мочевыводящих путях
откладываются примерно
у половины больных
подагрой.
38.
Мочекаменная болезнь• Наиболее часто такие камни представлены в
дистальных канальцах и собирательных трубочках.
Причиной отложения кристаллов мочевой
кислоты является гиперурикемия и повышенное
выведение уратов натрия с мочой. Главным
провоцирующим фактором кристаллизации
является увеличение кислотности мочи. При
понижении рН мочи ниже 5,75 ураты (енольная
форма) переходят в менее
растворимую кетоформу и кристаллизуются в
почечных канальцах.
39.
Основы лечениямочекаменной болезни
• Так же, как при подагре, лечение
сводится к
• беспуриновой диеты с большим
количеством чистой воды
• использованию аллопуринола.
• В дополнение
рекомендуется растительная диета,
приводящая к защелачиванию мочи,
что увеличивает в моче долю более
растворимой в воде енольной
формы мочевой кислоты. Вместе с
этим, уже имеющиеся кристаллы
мочевой кислоты (а также кристаллы
оксалатов) способны растворяться при
подщелачивании мочи.
40.
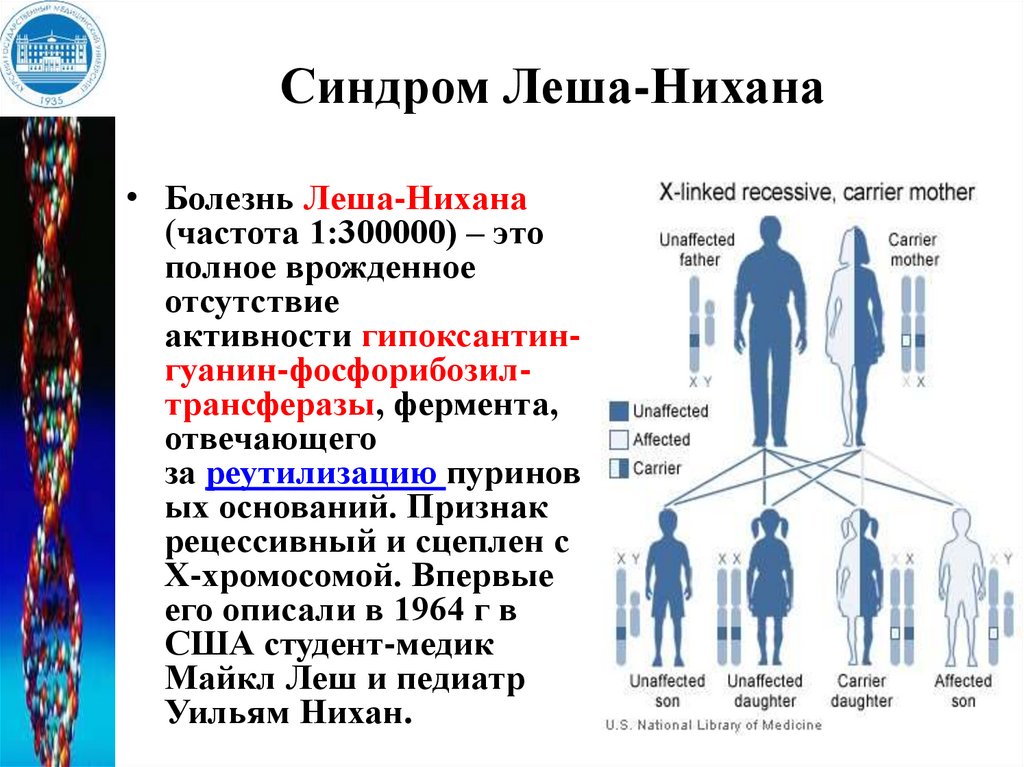
Синдром Леша-Нихана• Болезнь Леша-Нихана
(частота 1:300000) – это
полное врожденное
отсутствие
активности гипоксантингуанин-фосфорибозилтрансферазы, фермента,
отвечающего
за реутилизацию пуринов
ых оснований. Признак
рецессивный и сцеплен с
Х-хромосомой. Впервые
его описали в 1964 г в
США студент-медик
Майкл Леш и педиатр
Уильям Нихан.
41.
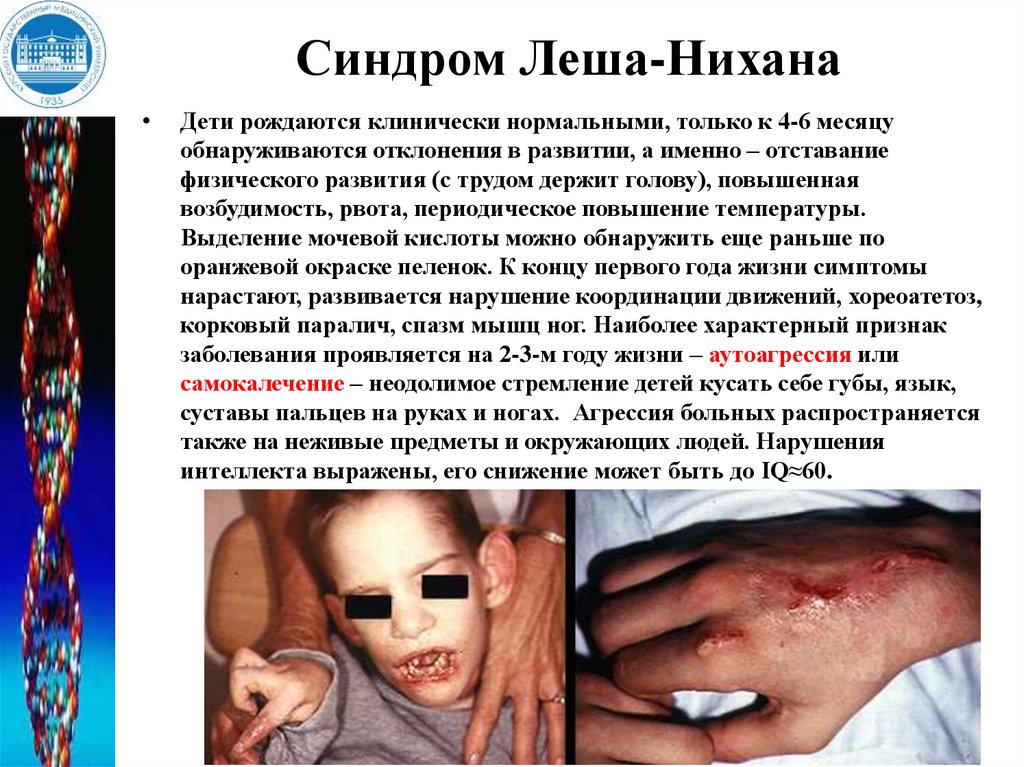
Синдром Леша-НиханаДети рождаются клинически нормальными, только к 4-6 месяцу
обнаруживаются отклонения в развитии, а именно – отставание
физического развития (с трудом держит голову), повышенная
возбудимость, рвота, периодическое повышение температуры.
Выделение мочевой кислоты можно обнаружить еще раньше по
оранжевой окраске пеленок. К концу первого года жизни симптомы
нарастают, развивается нарушение координации движений, хореоатетоз,
корковый паралич, спазм мышц ног. Наиболее характерный признак
заболевания проявляется на 2-3-м году жизни – аутоагрессия или
самокалечение – неодолимое стремление детей кусать себе губы, язык,
суставы пальцев на руках и ногах. Агрессия больных распространяется
также на неживые предметы и окружающих людей. Нарушения
интеллекта выражены, его снижение может быть до IQ≈60.
42.
Синдром Леша-Нихана• Лечение основано на
беспуриновой диете с
большим количеством
чистой воды и с
использованием
аллопуринола.
Производится
контроль деятельности
пациента с помощью
физических
ограничений и
психологической
терапии.