




























 medicine
medicineSimilar presentations:










Системы антибиотикорезистентности. Транспозоны и интегроны
1.
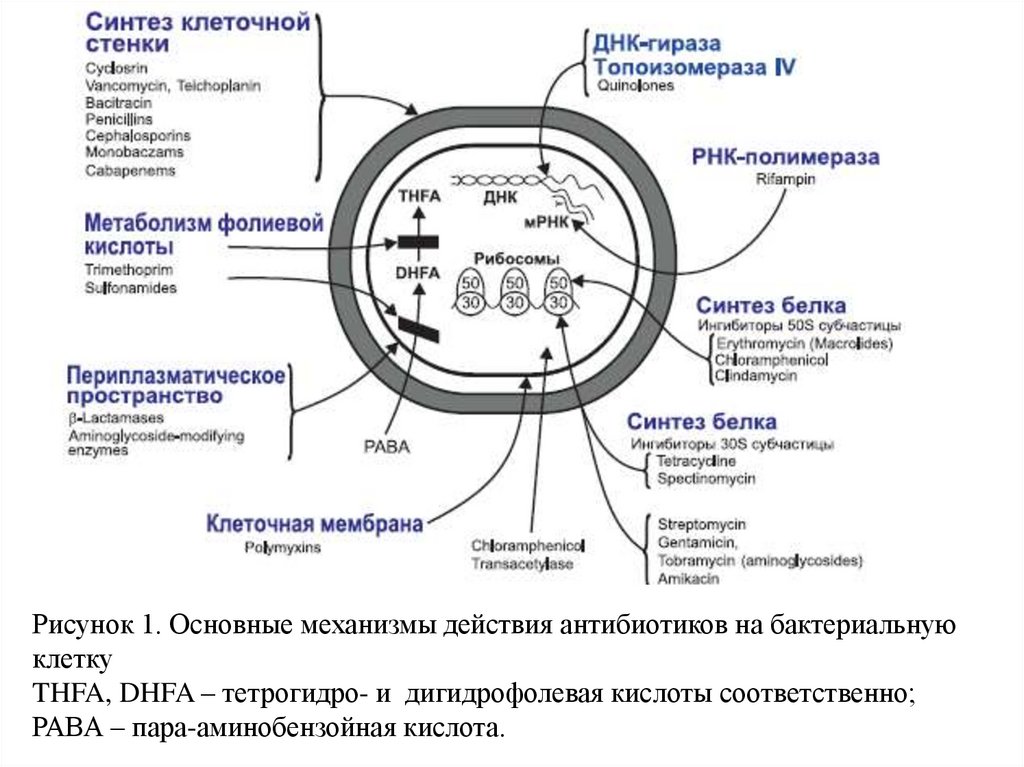
По механизму действия на бактериальную клеткувсе известные антибиотики можно разделить на 4
группы:
1. Синтез клеточной стенки
2. Изменение проницаемости мембраны
3. Ингибирование нуклеотидного метаболизма
4. Нарушение синтеза белка
2.
Рисунок 1. Основные механизмы действия антибиотиков на бактериальнуюклетку
THFA, DHFA – тетрогидро- и дигидрофолевая кислоты соответственно;
РАВА – пара-аминобензойная кислота.
3.
Системы резистентности включают:1. Детоксикация или инактивация антибиотика;
2. Уменьшение проникновения или усиление выведения
(эффлюкс);
3. Обход, устранение или сокращение стадии,
чувствительной для действия антибиотика;
4. Изменение мишени действия антибиотика;
5. Суперпродукция соединения, на которое действует
антибиотик;
6. Замена стадии метаболизма.
4.
Устойчивость к антибиотикам,ингибирующим
синтез
клеточной стенки
5.
Антибиотики группы пенициллинаЯвляются первыми антибиотиками, внедренными в
клиническую практику. Впервые пенициллин был обнаружен
в 1929 г. А. Флемингом, но в чистом виде пенициллин был
получен лишь в 1940 г. английским патологом Х.У. Флори
(H.W. Florey) и английским биохимиком Э.Б.Чейном
(E.B.Chein).
В 1942 г. российский микробиолог 3. В. Ермольева
самостоятельно получила отечественный пенициллин.
6.
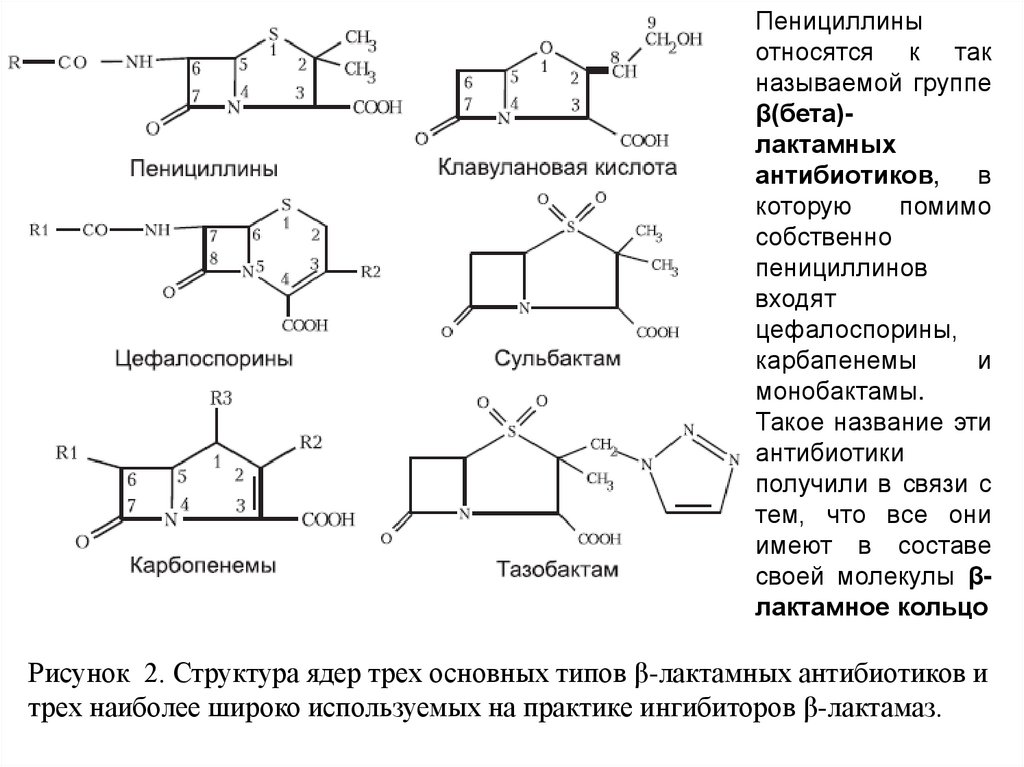
Пенициллиныотносятся к так
называемой группе
β(бета)лактамных
антибиотиков, в
которую
помимо
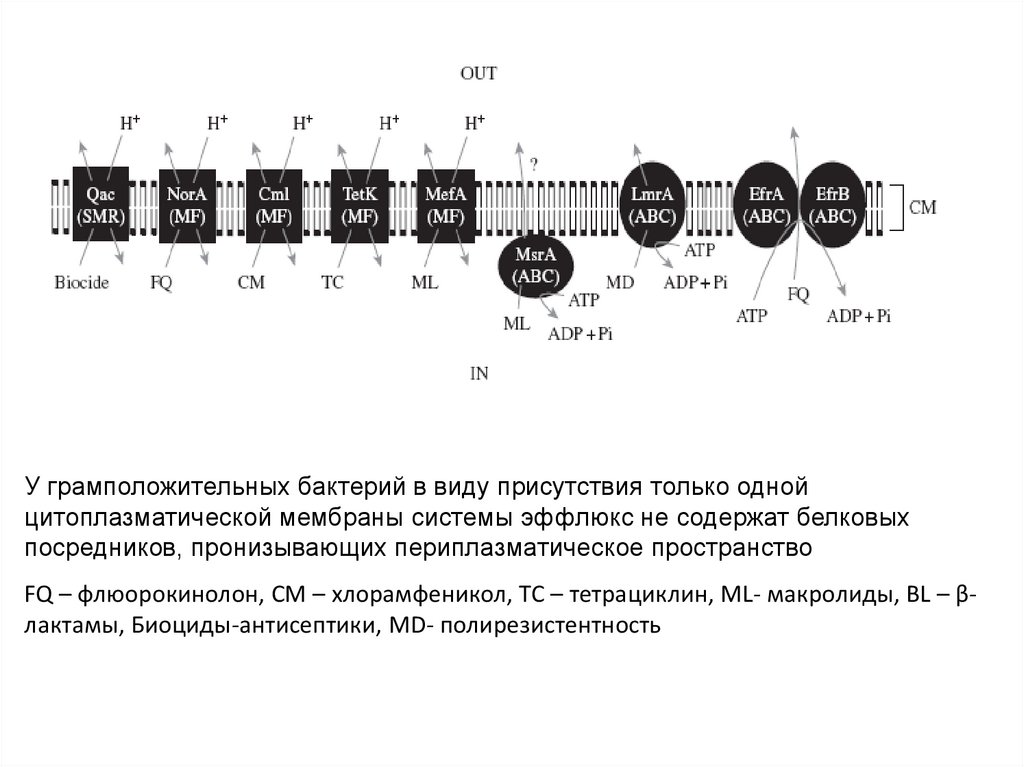
собственно
пенициллинов
входят
цефалоспорины,
карбапенемы
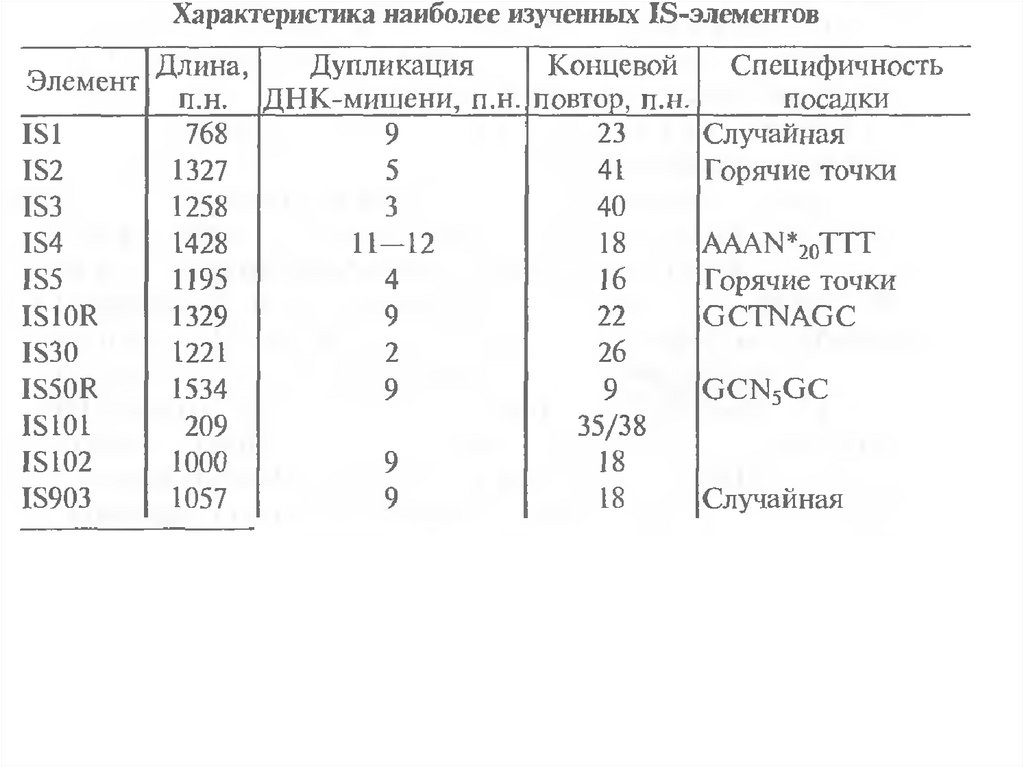
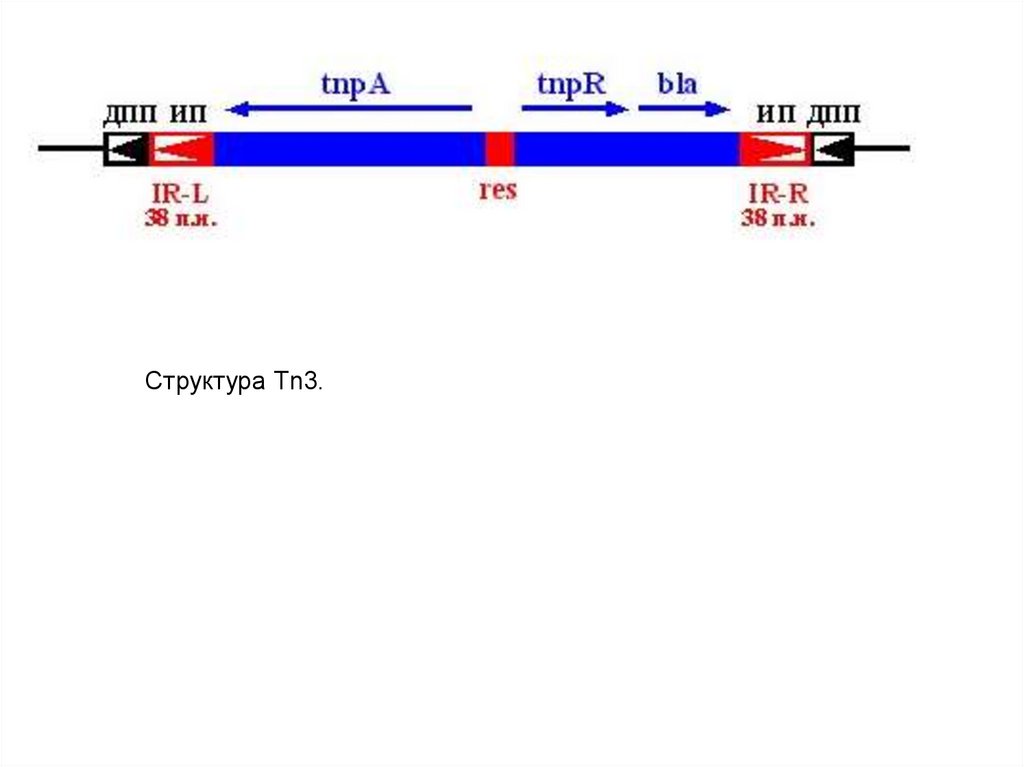
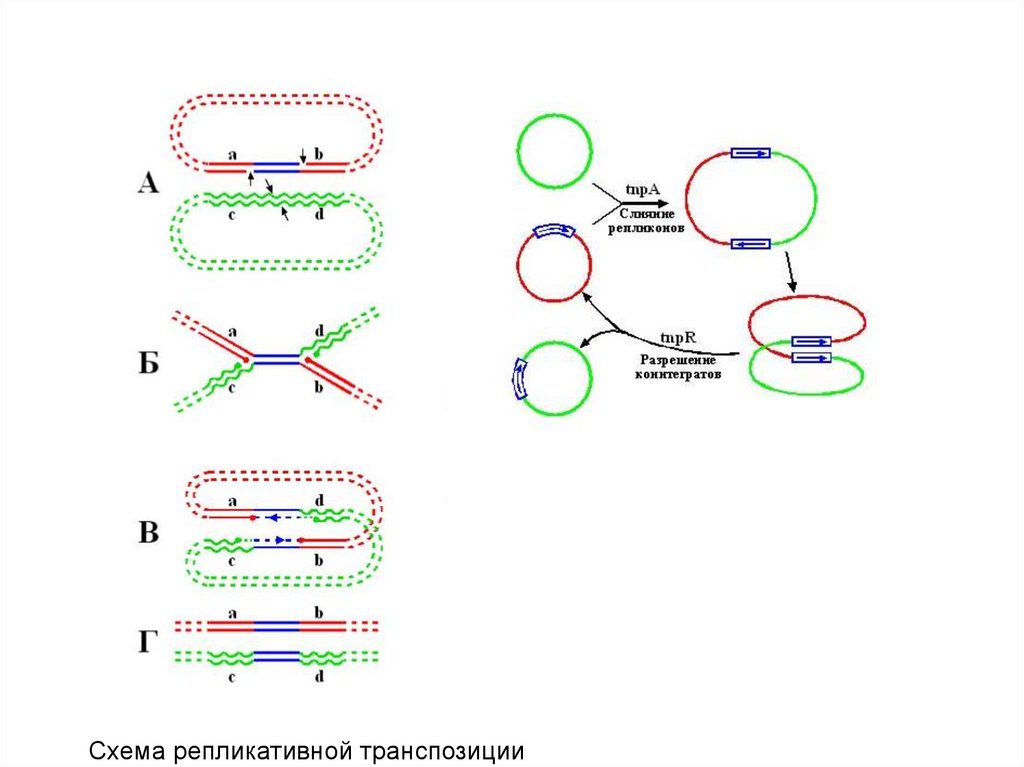
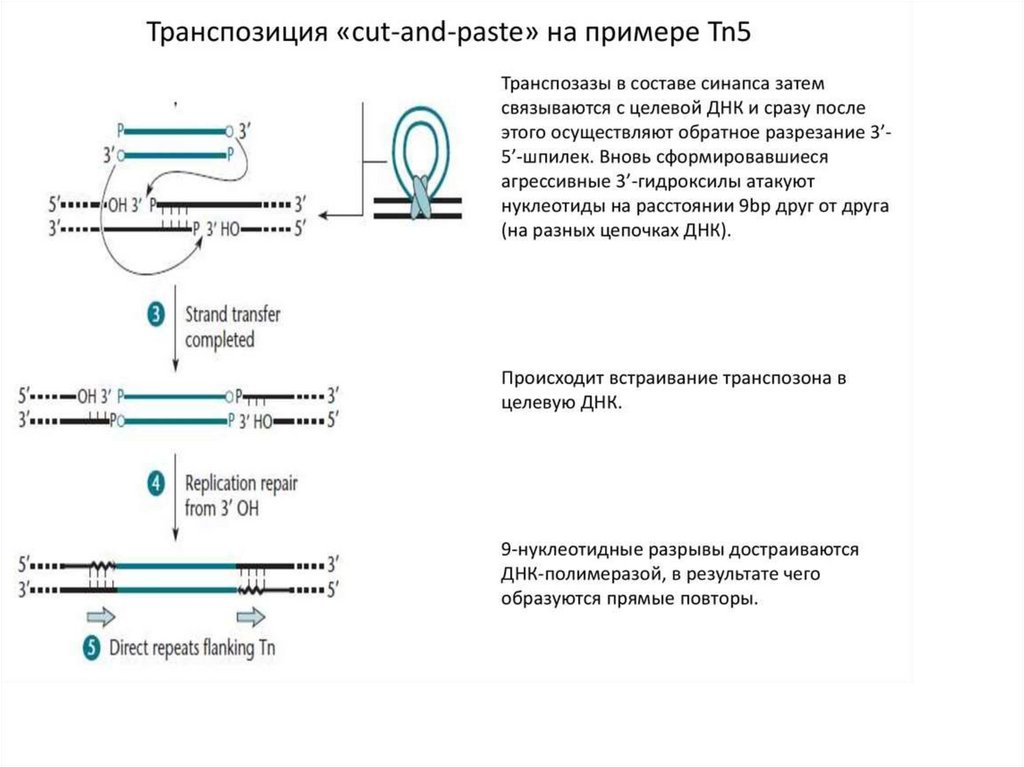
и
монобактамы.
Такое название эти
антибиотики
получили в связи с
тем, что все они
имеют в составе
своей молекулы βлактамное кольцо
Рисунок 2. Структура ядер трех основных типов β-лактамных антибиотиков и
трех наиболее широко используемых на практике ингибиторов β-лактамаз.
7.
В настоящее время выделяют более 400 β-лактамаз, отличающихся друг отдруга по способности гидролизовать отдельные группы β-лактамного кольца
антибиотиков, например, только пенициллинов или только цефалоспоринов,
или тех и других в равной степени.
У грамотрицательных микроорганизмов β-лактамазы локализованы в
периплазматическом
пространстве,
а
у
грамположительных
микроорганизмов β-лактамазы свободно диффундируют в окружающую
бактерии среду.
Пенициллины (и другие β-лактамные антибиотики), бета-лактамное кольцо
которых разрушается β-лактамазами (пенициллиназами) бактерий,
относятся к пенициллиназонеустойчивым антибиотикам.
В настоящее время в клинической практике широко используют
пенициллины (и другие β-лактамные антибиотики), содержащие в структуре
β-лактамного кольца специальные радикалы (химические группы),
защищающие антибиотики от расщепления β-лактамазами. Такие
антибиотики называют пенициллиназорезистентными. Кроме того, в
клиническую
практику
внедрены
комбинированные
препараты,
включающие β-лактамный антибиотик и специальный ингибитор βлактамаз.
8.
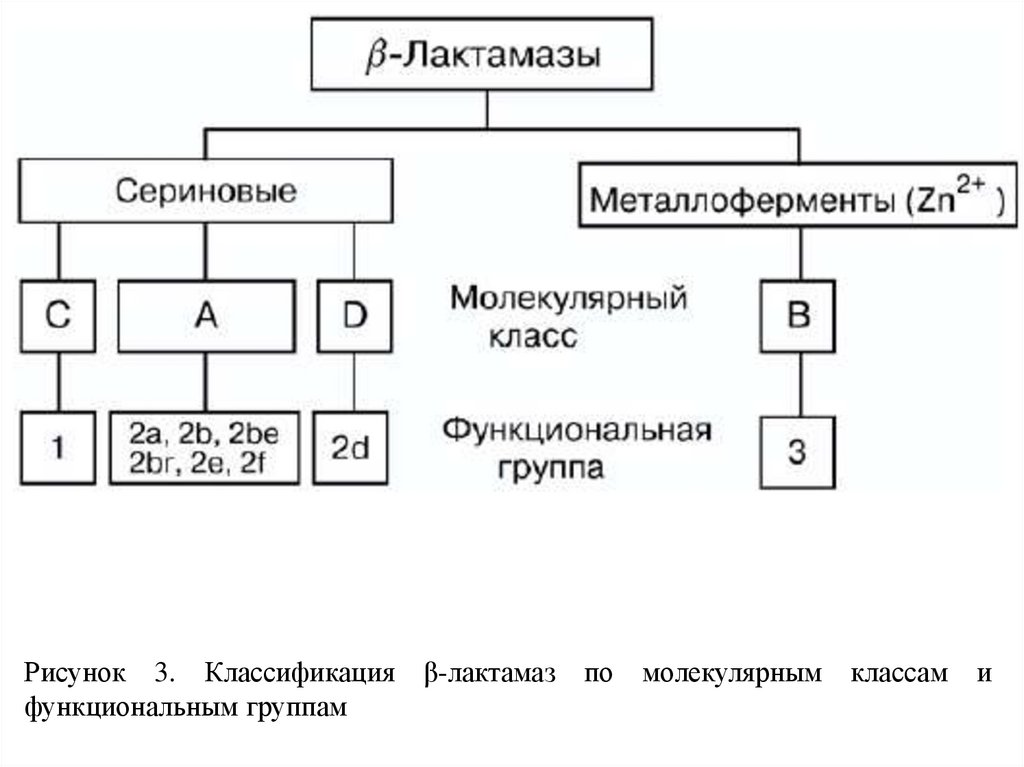
Рисунок 3. Классификацияфункциональным группам
β-лактамаз по молекулярным
классам
и
9.
В клетках прокариот существует большая группа белков иферментов,
называемая
«пенициллин-связывающими
белками» (ПСБ), которые участвуют в синтезе клеточной
стенки. В эту группу входят ферменты самых различных
классов, включая оксидоредуктазы и гидролазы. Основой
бактерицидного действия β-лактамов является подавление
роста клеточной стенки за счет ингибирования ПСБ.
В результате связывания пенициллинов с ПСБ, с одной
стороны, подавляется синтез пептидогликана (ригидного
слоя бактерий), а с другой - активизируются
аутолитические ферменты, приводящие к лизису клетки.
10.
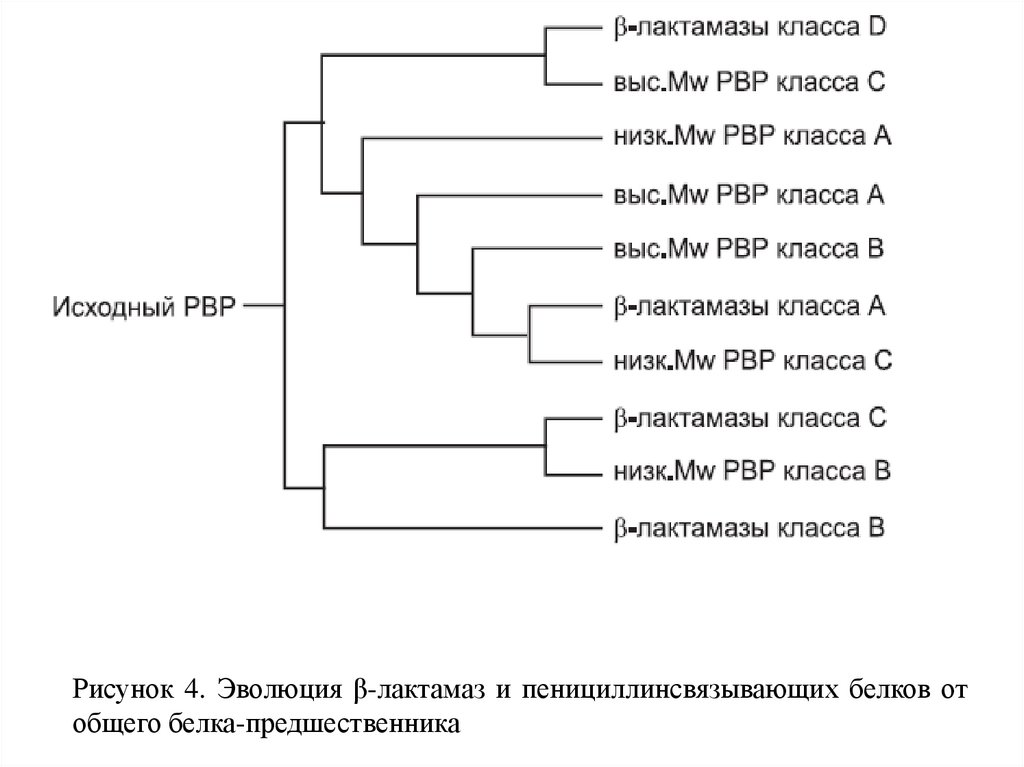
Рисунок 4. Эволюция β-лактамаз и пенициллинсвязывающих белков отобщего белка-предшественника
11.
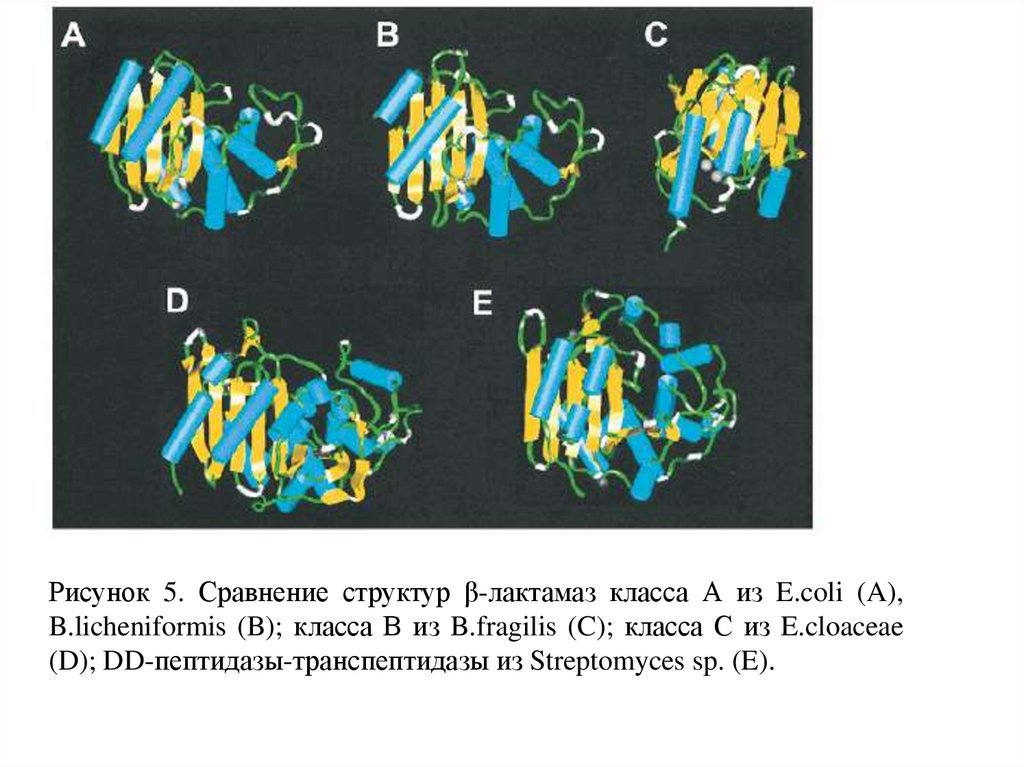
Рисунок 5. Сравнение структур β-лактамаз класса А из E.coli (A),B.licheniformis (B); класса В из B.fragilis (C); класса С из E.cloaceae
(D); DD-пептидазы-транспептидазы из Streptomyces sp. (E).
12.
Temoniera –первый
пациент, у
которого
была
обнаружена
ТЕМ-1
Рисунок 6. Положение и типы аминокислотных замен в β-лактамазе ТЕМ-1,
приводящих к образованию ферментов с «расширенным спектром действия»
13.
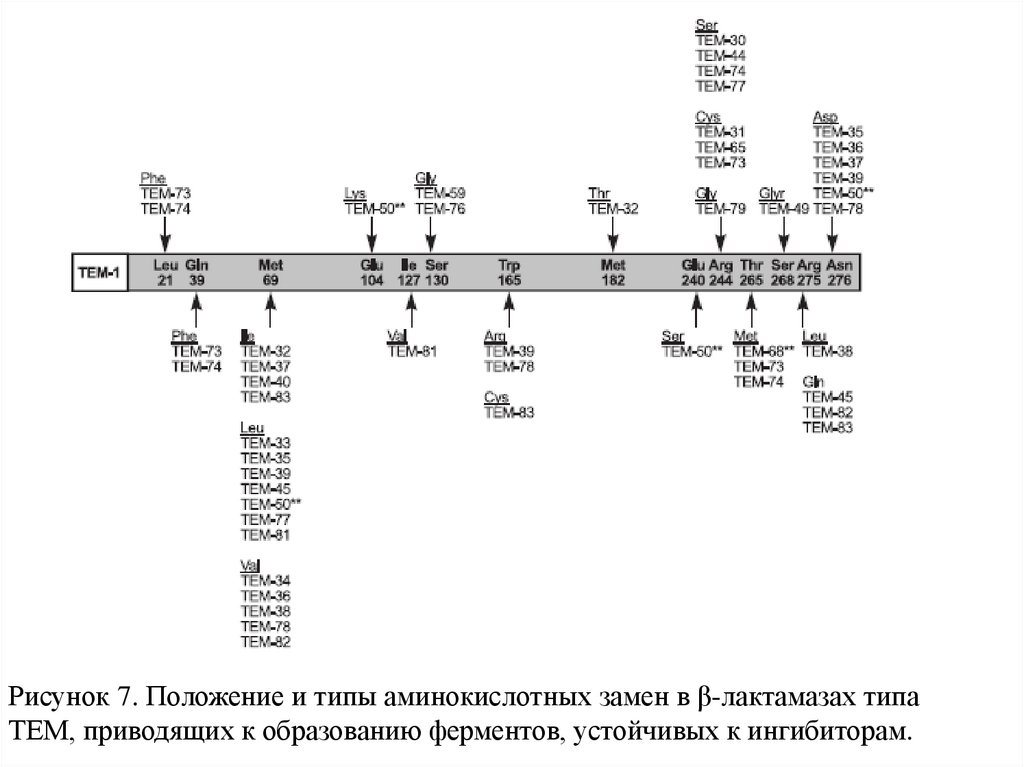
Рисунок 7. Положение и типы аминокислотных замен в β-лактамазах типаТЕМ, приводящих к образованию ферментов, устойчивых к ингибиторам.
14.
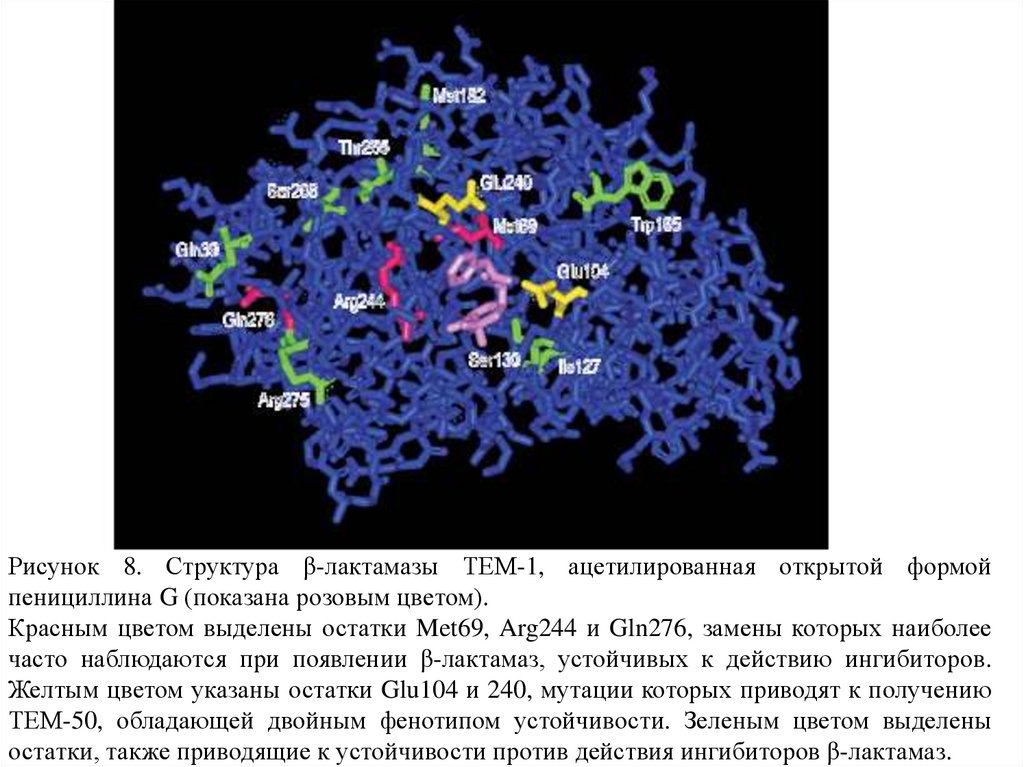
Рисунок 8. Структура β-лактамазы ТЕМ-1, ацетилированная открытой формойпенициллина G (показана розовым цветом).
Красным цветом выделены остатки Met69, Arg244 и Gln276, замены которых наиболее
часто наблюдаются при появлении β-лактамаз, устойчивых к действию ингибиторов.
Желтым цветом указаны остатки Glu104 и 240, мутации которых приводят к получению
ТЕМ-50, обладающей двойным фенотипом устойчивости. Зеленым цветом выделены
остатки, также приводящие к устойчивости против действия ингибиторов β-лактамаз.
15.
Характерной тенденцией последних 5–10 лет являются:1. Появление все новых и новых мутантных форм этих ферментов, устойчивых к
действию новых препаратов (включая цефалоспорины IV поколения и
карбапенемы). Около 50% новых ферментов были найдены в течение пяти
последних лет.
2. Комбинация в одном штамме сразу нескольких типов резистентности.
Например из более 800 штаммов из госпиталей и клиник Москвы, СанктПетербурга, Томска и Назрани, проверенных на резистентность к β-лактамам
более чем в 50% изолятов было обнаружено по две различных β-лактамазы (SHV
+ CTX-M или ТЕМ + СЕХ-М), а у 12% штаммов одновременно присутствовали
ТЕМ, SHV и СТХ-М.
3. Высокая скорость распространения новых мутантов во всем мире. Например,
β-лактамаза СТХ-М, которую еще недавно практически не находили в Российской
Федерации. Ферменты этой группы были обнаружены в 40% штаммов,
полученных из клиник Москвы и Санкт-Петербурга – наиболее часто
посещаемых городов России. Исследования показали, что устойчивость к
цефалоспоринам III-IV поколения (цефтазидиму) возникает уже через два-три
месяца с момента его применения в клинике. Поэтому во многих госпиталях
через каждые три месяца практикуют заменять использованный β-лактам на
новый. Появление мутантных β-лактамаз с несколькими фенотипами
устойчивости требует разработки методов экспресс-диагностики типов
резистентности с использованием методов генетической инженерии.
16.
Гликопептидные антибиотики (ванкомицин и тейкопланин) традиционноиспользуются при лечении энтерококковых инфекций.
Несмотря на то, что ванкомицин применяется в медицинской практике с
начала 50-х годов, первое сообщение об устойчивости энтерококков к этому
антибиотику появилось только в конце 80-х годов (Vancomycin-resistant
Enterococcus faecium – VRE).
17.
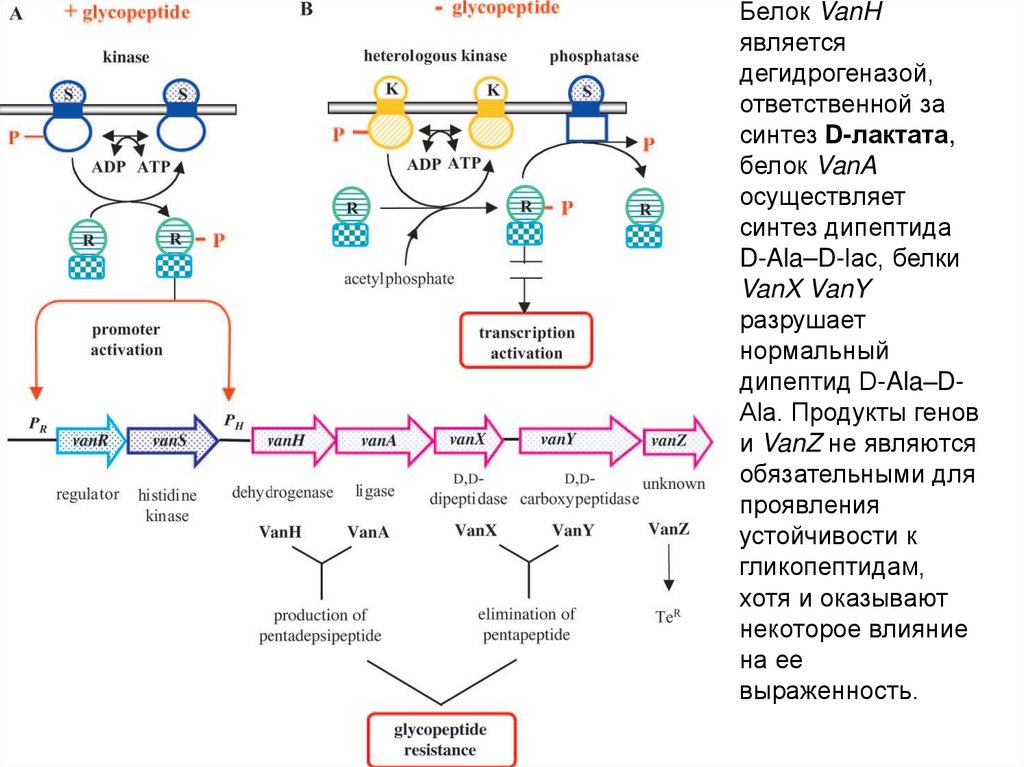
У энтерококков c фенотипом VanA синтез модифицированногопредшественника является результатом активности минимум 7
генов, входящих в состав оперона, локализованного на
транспозоне Tn1546. Гены, детерминирующие устойчивость к
ванкомицину
начинают
экспрессироваться
только
в
присутствии
антибиотика.
Это
обусловлено
функционированием так называемой двухкомпонентной
системы (широко распространенный механизм ответа
бактериальной клетки на изменения внешней среды).
18.
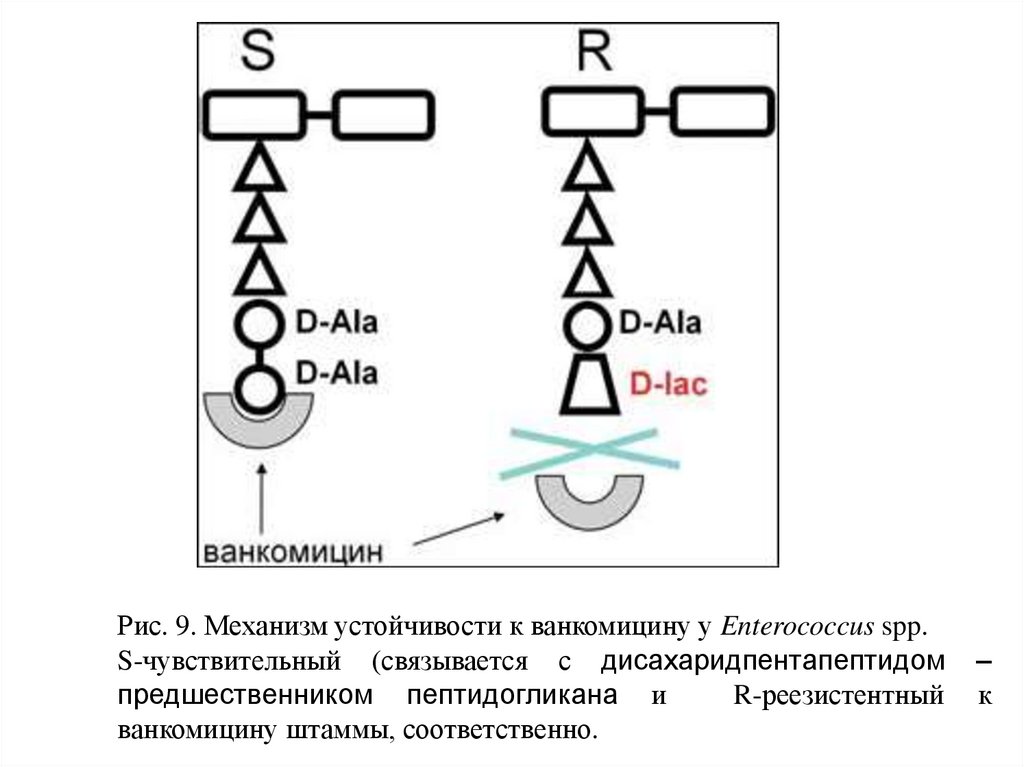
Рис. 9. Механизм устойчивости к ванкомицину у Enterococcus spp.S-чувствительный (связывается с дисахаридпентапептидом
предшественником пептидогликана и
R-реезистентный
ванкомицину штаммы, соответственно.
–
к
19.
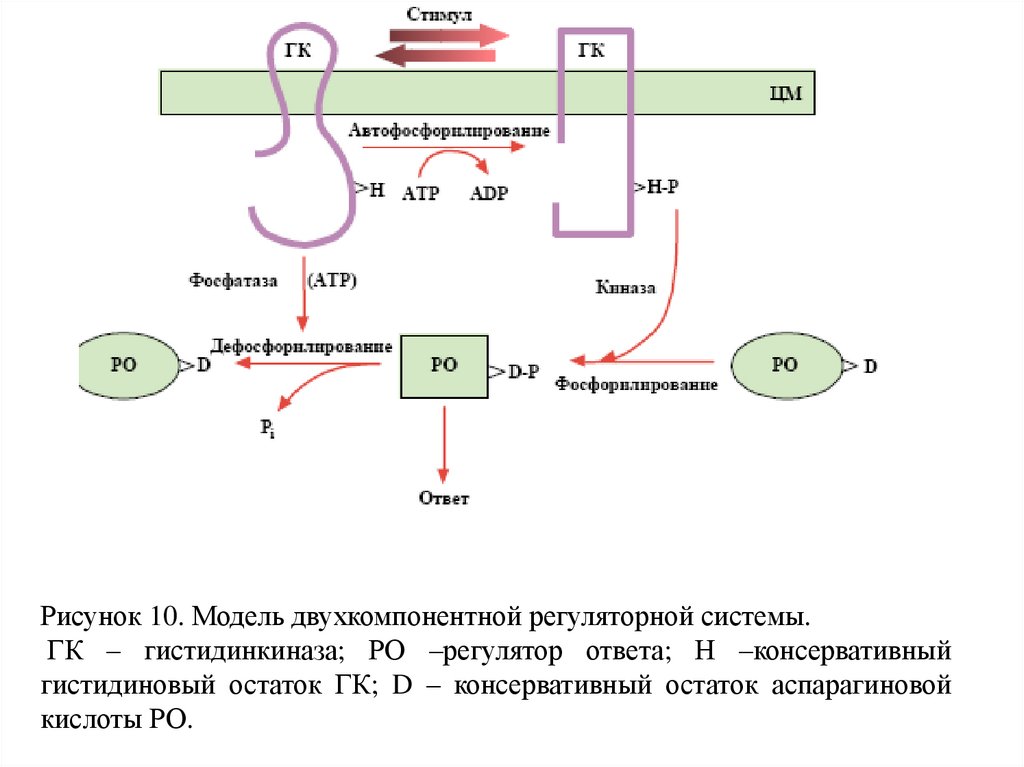
Рисунок 10. Модель двухкомпонентной регуляторной системы.ГК – гистидинкиназа; РО –регулятор ответа; Н –консервативный
гистидиновый остаток ГК; D – консервативный остаток аспарагиновой
кислоты РО.
20.
Белок VanHявляется
дегидрогеназой,
ответственной за
синтез D-лактата,
белок VanA
осуществляет
синтез дипептида
D-Ala–D-lac, белки
VanX VanY
разрушает
нормальный
дипептид D-Ala–DAla. Продукты генов
и VanZ не являются
обязательными для
проявления
устойчивости к
гликопептидам,
хотя и оказывают
некоторое влияние
на ее
выраженность.
21.
Структура оперона, определяющего фенотип VanB,принципиально сходна со структурой оперона VanA,
описанной выше. Выявлена почти 80%-ная гомология между
генами,
кодирующими
ферменты
синтеза
модифицированного мономера. Однако в структуре генов,
кодирующих регуляторные белки, найдены значительные
отличия – степень гомологии составляет 25–35%. Этим,
вероятно, можно объяснить тот факт, что у штаммов,
демонстрирующих фенотип VanB, тейкопланин не
индуцирует
продукцию
ферментов,
синтезирующих
модифицированный предшественник. В результате штаммы
VanB фенотипа при вариабельном уровне устойчивости к
ванкомицину сохраняют чувствительность к тейкопланину,
что является отличительным признаком указанного фенотипа.
22.
Среди антибиотиков нарушающихсинтез
белка
наиболее
часто
плазмидное происхождение имеют
гены, детерминирующие устойчивость
к аминогликозидным антибиотикам,
хлорамфениколу,
антибиотикам
группы МКЛС и тетрациклину.
23.
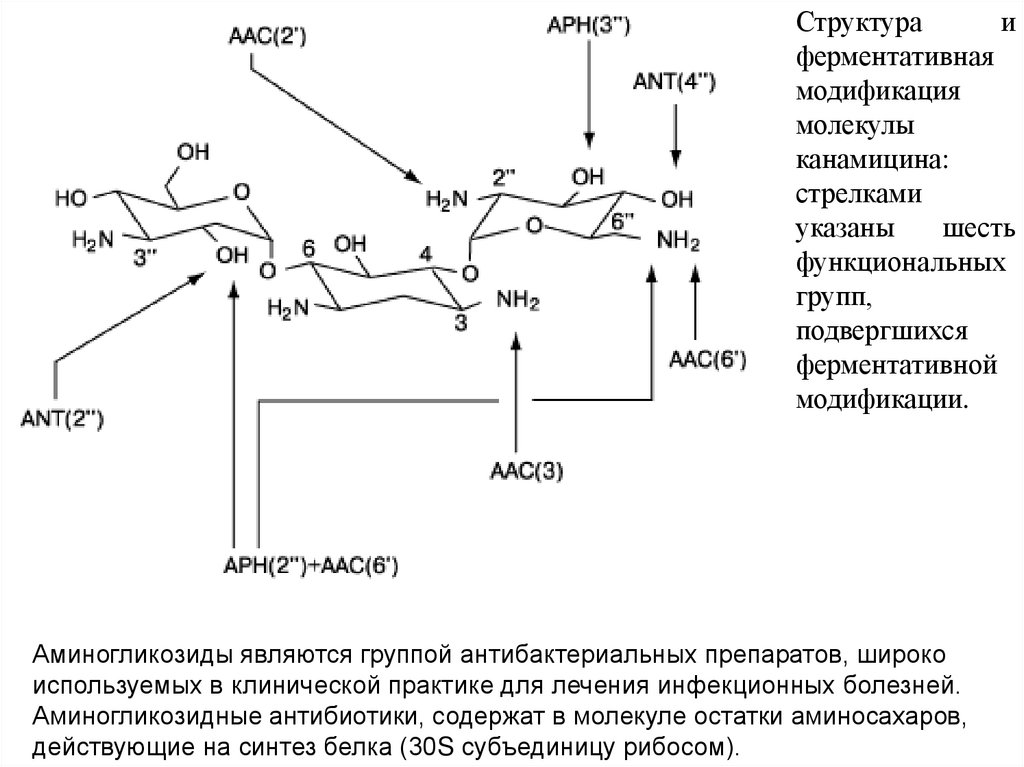
Структураи
ферментативная
модификация
молекулы
канамицина:
стрелками
указаны
шесть
функциональных
групп,
подвергшихся
ферментативной
модификации.
Аминогликозиды являются группой антибактериальных препаратов, широко
используемых в клинической практике для лечения инфекционных болезней.
Аминогликозидные антибиотики, содержат в молекуле остатки аминосахаров,
действующие на синтез белка (30S субъединицу рибосом).
24.
Ферментативная модификация молекулы антибиотика являетсянаиболее распространенным механизмом резистентности.
Различают три вида аминогликозидмодифицирующих ферментов
(АГМФ):
1. Фосфотрансферазы
2. Ацетилтрансферазы,
3. Аденилилтрансферазы
(нуклеотидилтрансферазы).
25.
Аминогликозидфосфотрансферазы (APH) – это ферменты, модифицирующиеаминогликозидные
антибиотики
путем
фосфорилирования
их
гидроксильных групп в присутствии АТФ в качестве кофактора. В
большей или меньшей степени изучен ряд APH, продуцируемых как
грамотрицательными, так и грамположительными микроорганизмами.
В зависимости от положения гидроксильной группы антибиотика,
модифицируемой ферментом, различают 5 типов фосфотрансфераз:
APH(3'); APH(3''); APH(6); АРН(4); АРН(2'') – в составе бифункционального
фермента ААС(6') + АРН(2'').
Наибольшее клиническое значение имеет фермент АРН(3'), распространенный
у грамотрицательных бактерий и обусловливающий резистентность к
аминогликозидам разных поколений. Данный фермент может быть представлен
7 изоферментами: APH(3')-I – APH(3')-VII, кодируемыми соответствующими
генами: aphA1 – aphA7.
Наиболее распространенным является фермент APH(3')-I, обусловливающий
резистентность к канамицину, неомицину, мономицину и ливидомицину. Этот
фермент обнаружен у представителей семейства Enterobacteriaceae,
Pseudomonas spp., Acinetobacter spp. Широкому распространению кодирующего
этот фермент гена способствовала его локализация в составе различных
транспозонов: Tn903, Tn6 (Tn 2680), Tn 1525, Tn 1699, Tn 2350, Tn 4350.
26.
Аминогликозиднуклеотидилтрансферазы (ANT), называемые такжеаминогликозидаденилилтрансферазами
(AAD),
осуществляют
модификацию молекулы аминогликозидных антибиотиков за счет
аденилирования гидроксильных групп их молекул в присутствии АТФ в
качестве ко-фактора. Известно 4 типа нуклеотидилтрансфераз: ANT(4'),
ANT(2''), ANT(3'') и ANT(6').
Аденилилтрансфераза ANT(4'), обозначаемая также как ANT(4')(4''),
обусловливает резистентность стафилококков и стрептококков к амикацину,
тобрамицину,
канамицину,
неомицину,
мономицину,
бутирозину,
ливидомицину и исепамицину. Ген, кодирующий этот фермент, обнаружен в
составе многокопийных и низкокопийных плазмид.
Аминогликозидацетилтрансферазы (ААС) ацетилируют аминогруппы
молекулы аминогликозидов в присутствии ацетил-КоA в качестве кофактора. Известны 4 типа этих ферментов: AAC(6'), AAC(2'), AAC(3) и
AAC(1).
При изучении структуры гена, кодирующего аденилилтрансферазу
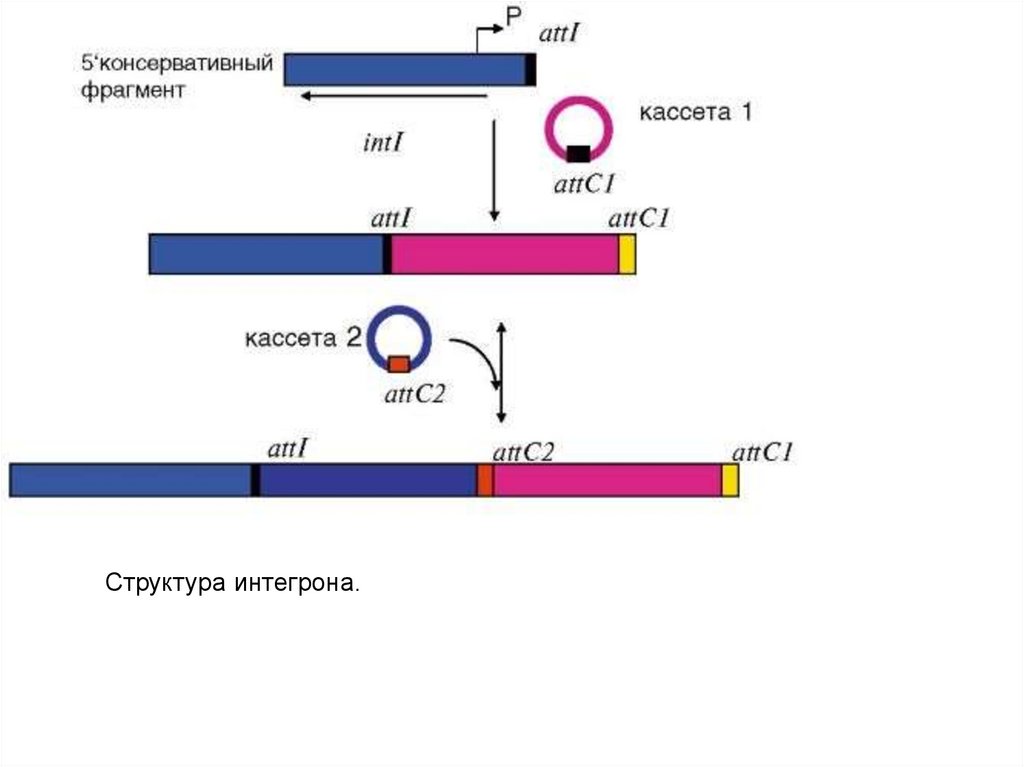
ANT(3'')(9), показано, что он входит в состав интегронов.
27.
Природная устойчивость ко всем аминогликозидным антибиотикам(перекрестная резистентность) характерна для облигатных анаэробов и
обусловлена отсутствием системы электронного транспорта, что не
позволяет антибиотику проникнуть внутрь бактериальной клетки.
Приобретенная резистентность к аминогликозидам, обусловленная
снижением электронного транспорта, часто наблюдается у P.aeruginosa,
реже – у представителей семейства Enterobacteriaceae. В отношении
таких штаммов аминогликозиды могут быть эффективны только при
использовании высоких доз. Кроме того, было показано, что низкие
уровни резистентности к аминогликозидам у микроорганизмов могут
быть обусловлены мутациями, изменяющими липополисахаридный
состав клеточной стенки бактерий. Такой тип резистентности, повидимому, обусловлен уменьшением числа потенциальных участков
связывания липополисахаридов с положительно заряженными
аминогруппами в составе аминогликозидов. К резистентности могут
также приводить мутации генов, кодирующих пориновые белки внешней
мембраны грамотрицательных бактерий. Однако этот механизм
встречается значительно реже, чем изменения липополисахаридов или
системы электронного транспорта.
28.
Стрептомици́н—
исторически
второй антибиотик (после пенициллина), он же первый из
группы аминогликозидов, а также первый, оказавшийся
эффективным против туберкулеза и чумы. Был открыт
американским микробиологом Зельманом Ваксманом. Образуется в
процессе жизнедеятельности лучистых грибов Streptomyces
globisporus streptomycini или других родственных микроорганизмов.
У стрептомициноустойчивых микроорганизмов рибосомы в
результате модификации белков 30S субъединицы теряют
способность связывать стрептомицин.. Имеются также сообщения о
резистентности к гентамицину, вызванной структурными
изменениями белка 50S.
Хромосомальная устойчивость к стрептомицину
мутациями гена rpsL, кодирующего белок 12S
обусловлена
29.
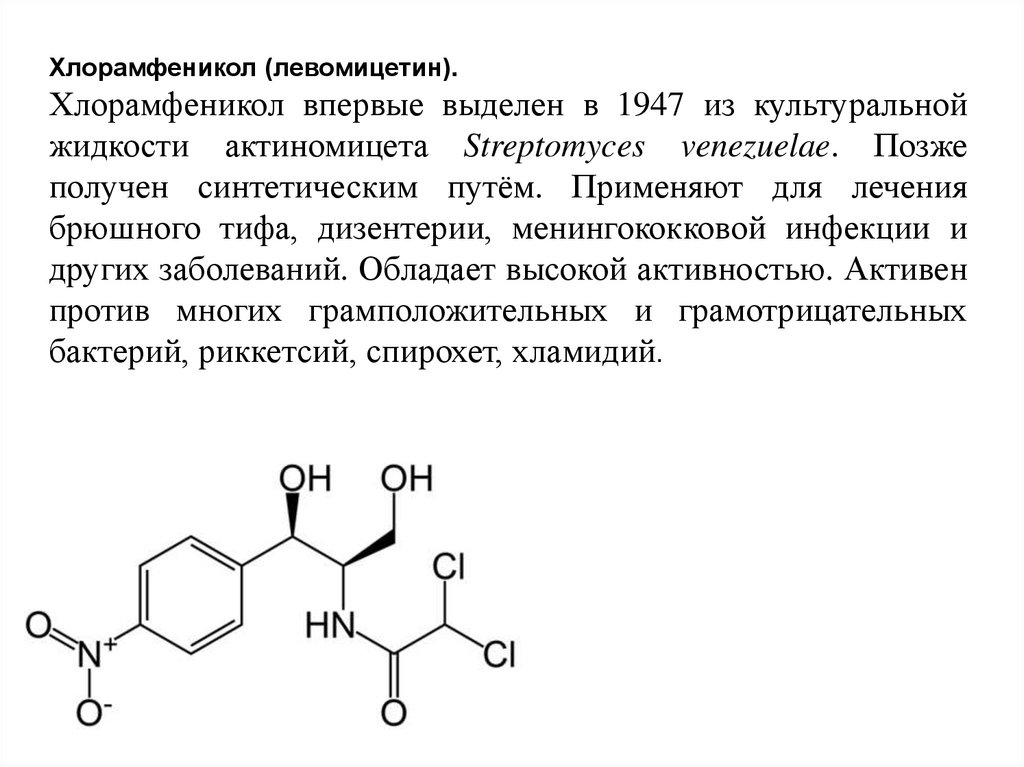
Хлорамфеникол (левомицетин).Хлорамфеникол впервые выделен в 1947 из культуральной
жидкости актиномицета Streptomyces venezuelae. Позже
получен синтетическим путём. Применяют для лечения
брюшного тифа, дизентерии, менингококковой инфекции и
других заболеваний. Обладает высокой активностью. Активен
против многих грамположительных и грамотрицательных
бактерий, риккетсий, спирохет, хламидий.
30.
Наиболеечасто
встречающимся
механизмом
устойчивости к хлорамфениколу является инактивация
антибиотика
посредством
его
модификации
(ацетилирования) с помощью ацетилтрансфераз (САТ),
кодируемых генами плазмид и транспозонов. Фермент
обеспечивает преобразование хлорамфеникола сначала в
3-ацетоксихлорамфеникола,
а
затем
в
1,3-диацетоксихлорамфеникол, который является не
активным, Существует несколько типов САТ. Достаточно
распространенным
является
тип
I,
имеющий
транспозонную природу (локализован в транспозоне
Tn9).
31.
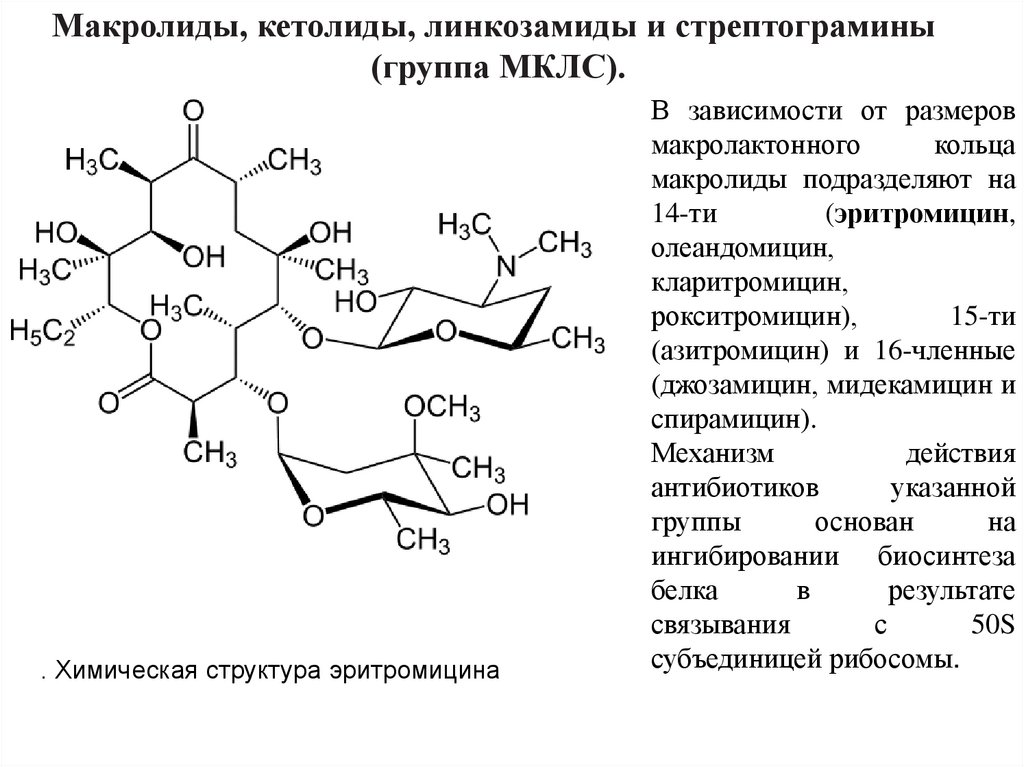
Макролиды, кетолиды, линкозамиды и стрептограмины(группа МКЛС).
. Химическая структура эритромицина
В зависимости от размеров
макролактонного
кольца
макролиды подразделяют на
14-ти
(эритромицин,
олеандомицин,
кларитромицин,
рокситромицин),
15-ти
(азитромицин) и 16-членные
(джозамицин, мидекамицин и
спирамицин).
Механизм
действия
антибиотиков
указанной
группы
основан
на
ингибировании биосинтеза
белка
в
результате
связывания
с
50S
cубъединицей рибосомы.
32.
Хотя актибиотики группы МКЛС активны в отношенииширокого спектра бактерий, однако на практике их
обычно применяют против ряда грамположительных
микроорганизмов – Staphylococcus spp., Streptococcus spp.,
атипичных патогенов – Chlamydia spp., Mycoplasma spp., а
из грамотрицательных бактерий только в случае
инфицирования Helicobacter pylori. Долгое время
считалось, что низкая активность в отношении
грамотрицательных
бактерий
связана
с
низкой
проницаемостью этих антибиотиков через пориновые
каналы, однако, очевидно, не менее важен и процесс их
активного выведения из организма.
33.
Препараты группы МКЛС связываются с доменами II и V 23SрРНК. Основным участком связывания антибиотиков является
домен V, причем основные точки связывания – нуклеотиды в
положениях А2058, А2059 и G2505, хотя недавно обнаружено
связывание и по некоторым другим позициям. В пределах
домена II антибиотик взаимодействует с нуклеотидом в
положении А752, что наиболее характерно для кетолидов, и
этим объясняют их более высокую антибактериальную
активность, а также отсутствие перекрестной устойчивости с
макролидами. Связь с 23S рРНК препятствует сборке 50S
субъединицы и процессу элонгации.
34.
Устойчивость к препаратам группы МКЛС можетбыть связана с модификацией мишени действия, их
активным выведением и быстрой инактивацией.
Однако наиболее распространенным и практически
важным
механизмом
устойчивости
является
модификация мишени действия. Модификация 23S
рРНК
осуществляется
либо
в
результате
метилирования аденина в положении 2058, либо в
результате ряда нуклеотидных замен. Из двух
приведенных
механизмов
наиболее
распространенным и клинически важным является
метилирование. Метилирование (моно- или ди-)
осуществляется
семейством N-метилтрансфераз,
соответствующие гены которых получили название
erm (erythromycin ribosome methylation).
35.
В настоящее время у различных бактерий описано около 40erm генов, локализованных обычно на плазмидах. Известны
два варианта экспрессии устойчивости, связанной с
метилированием: конститутивный и индуцибельный. При
конститутивном типе экспрессии синтез активной мРНК
метилазы происходит независимо от наличия индукторов.
Фенотипически это проявляется в перекрестной устойчивости
к макролидам, линкозамидам и стрептограминам В; кетолиды
при этом сохраняют активность. При индуцибельном типе
экспрессии в отсутствие индуктора синтез мРНК метилазы не
происходит до конца за счет образования шпильки. Синтез
полноразмерной мРНК становится возможным в результате
перестройки конформации мРНК после связывания с
индуктором аттенуатора экспрессии. Способность отдельных
антибиотиков индуцировать перестройку мРНК определяется
в основном структурой аттенуатора, различающейся у
отдельных видов бактерий.
36.
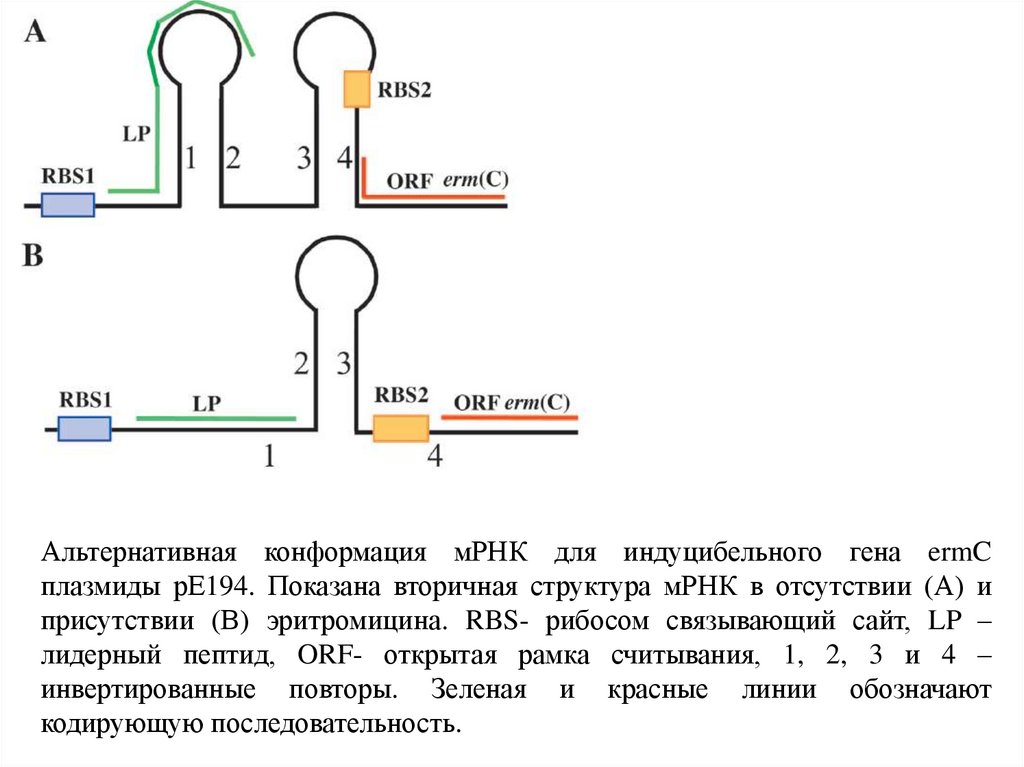
Альтернативная конформация мРНК для индуцибельного гена ermCплазмиды рЕ194. Показана вторичная структура мРНК в отсутствии (А) и
присутствии (В) эритромицина. RBS- рибосом связывающий сайт, LP –
лидерный пептид, ORF- открытая рамка считывания, 1, 2, 3 и 4 –
инвертированные повторы. Зеленая и красные линии обозначают
кодирующую последовательность.
37.
Для практики важно, что у Staphylococcus spp. индуцирующейактивностью обладают 14- и 15-членные макролиды. Таким
образом, штаммы с индуцибельным характером экспрессии
метилаз, устойчивые к указанным антибиотикам, сохраняют
чувствительность к 16-членным макролидам и стрептограминам.
У Streptococcus spp. индуцирующей активностью обладают все
антибиотики группы МКЛС, однако у 16-членных макролидов,
линкозамидов и стрептограминов индуцирующая активность
незначительна. На практике это приводит к значительному
разнообразию фенотипов.
38.
Вторым, относительно недавно расшифрованныммеханизмом модификации мишени действия
антибиотиков группы МКЛС, являются мутации в
генах рРНК и рибосомальных белков, приводящие к
конформационным
изменениям
пептидилтрансферазного центра и, соответственно, к
снижению аффинности препаратов. Детальная
информация об известных мутациях в генах рРНК
доступна на веб-сайте http://ribosome.fandm.edu.
Мутации в генах рРНК являются основным
механизмом устойчивости к макролидам у H. pylori.
39.
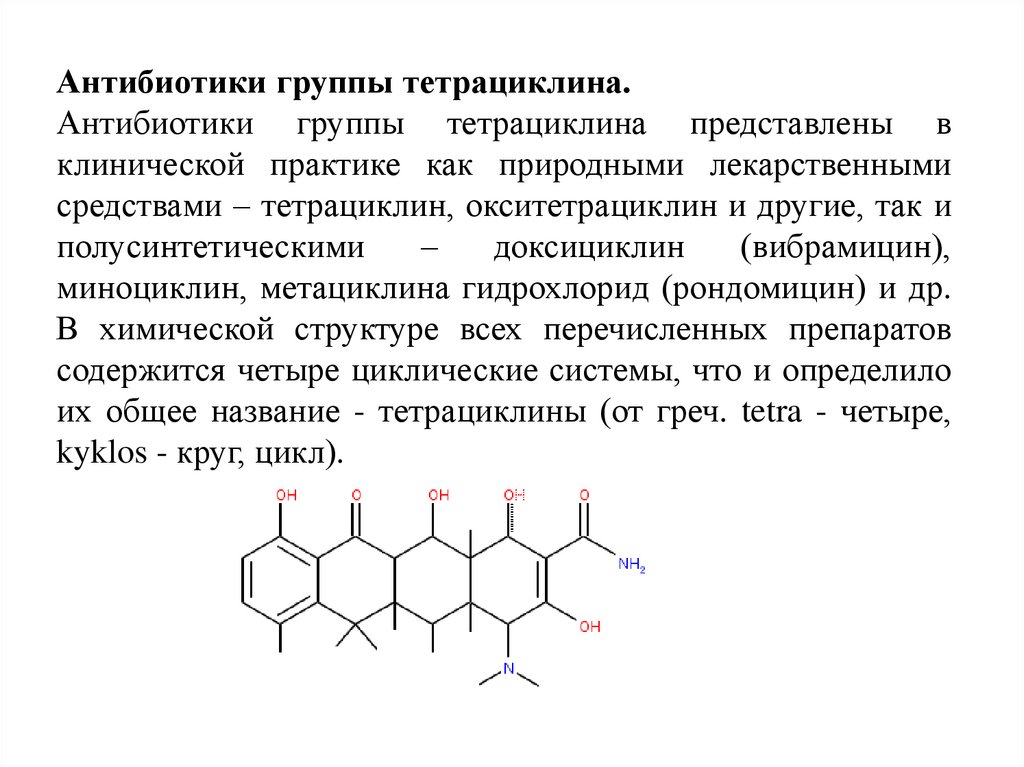
Антибиотики группы тетрациклина.Антибиотики группы тетрациклина представлены в
клинической практике как природными лекарственными
средствами – тетрациклин, окситетрациклин и другие, так и
полусинтетическими
–
доксициклин
(вибрамицин),
миноциклин, метациклина гидрохлорид (рондомицин) и др.
В химической структуре всех перечисленных препаратов
содержится четыре циклические системы, что и определило
их общее название - тетрациклины (от греч. tetra - четыре,
kyklos - круг, цикл).
40.
Первый представитель этой группы лекарств - хлортетрациклин был получен из грибов вида Streptomyces aureofaciens в 1948 г. Двагода спустя из грибов вида Streptomyces rimosus был выделен
препарат окситетрациклин.
В основе механизма действия лекарственных средств этой группы
лежит их способность обратимо связываться со специфичными
для них рецепторами, расположенными на бактериальных
рибосомах, в результате они препятствуют включению
аминокислот в строящуюся пептидную цепь и тем самым
нарушают синтез белка в микробной клетке, что в конечном итоге
препятствует их делению. Тетрациклины проникают в микробную
клетку в основном путем пассивной диффузии, но в некоторых
случаях и путем активного транспорта.
Наиболее часто гены резистентности к тетрациклину,
расположенные на плазмидах, обеспечивают активное выведение
тетрациклина из микробной клетки.
41.
Ассоциированные с мембраной транспортерыотносятся к одному из 5 семейств
1. АВС –семейство (или АТФ-связывающая
кассета)
2. MF иногда MFS –семейство (major facilitator
superfamily)
3. SMR-семейство (small multidrug resistance
family) малое семейство, обеспечивающее
устойчивости в различным лекарственным
препаратам)
4. MATE-семейство (multidrug and toxin extrusion
family)
5. RND-семейство (resistance-nodulation-division)
42.
Члены всех вышеперечисленных семейств, кромесемейства ABC (члены которого гидролизуют АТФ
для удаления антибиотика и других соединений из
клетки) действуют как вторичные переносчики, т.е.
выводят антибиотик из клетки и закачивают в клетку
ионы водорода (H +) или ионы натрия (Na +)
(антипорт-системы).
Системы
эффлюкс
обеспечивают выведения не только антибиотиков, но
играют ключевую роль в патогенности бактерий,
образовании биопленок, а также обеспечивают
устойчивость к стрессовым факторам среды
(например, от токсичного действия активных форм
кислорода, в частности, перекиси водорода).
43.
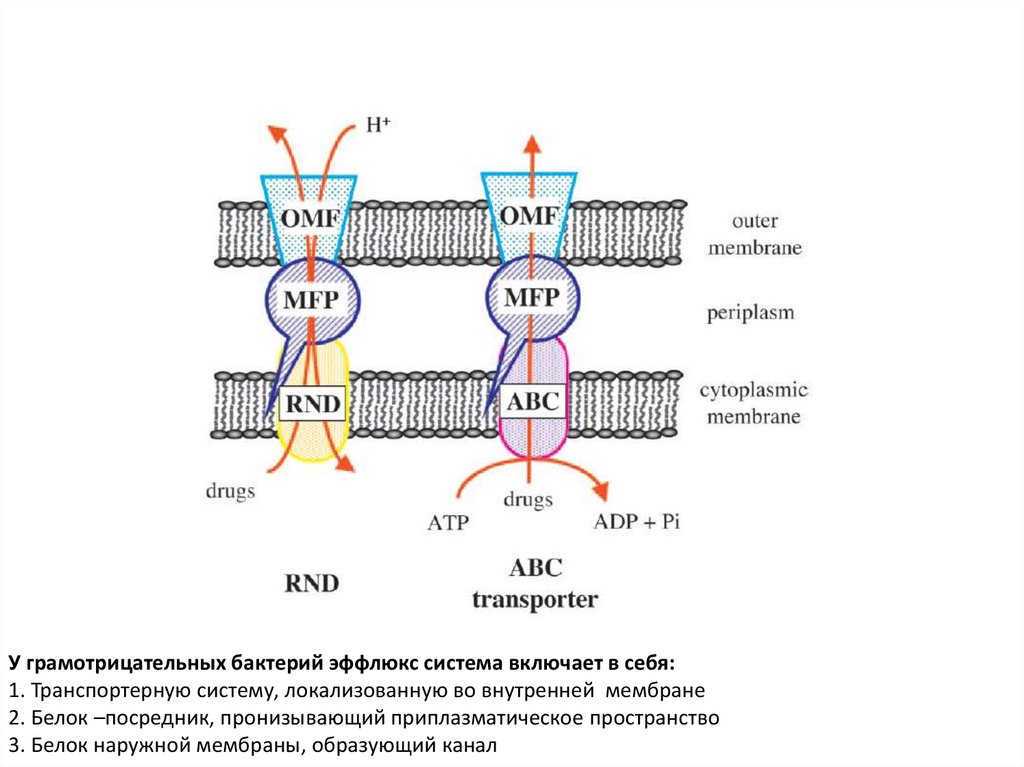
У грамотрицательных бактерий эффлюкс система включает в себя:1. Транспортерную систему, локализованную во внутренней мембране
2. Белок –посредник, пронизывающий приплазматическое пространство
3. Белок наружной мембраны, образующий канал
44.
У грамположительных бактерий в виду присутствия только однойцитоплазматической мембраны системы эффлюкс не содержат белковых
посредников, пронизывающих периплазматическое пространство
FQ – флюорокинолон, CM – хлорамфеникол, TC – тетрациклин, ML- макролиды, BL – βлактамы, Биоциды-антисептики, MD- полирезистентность
45.
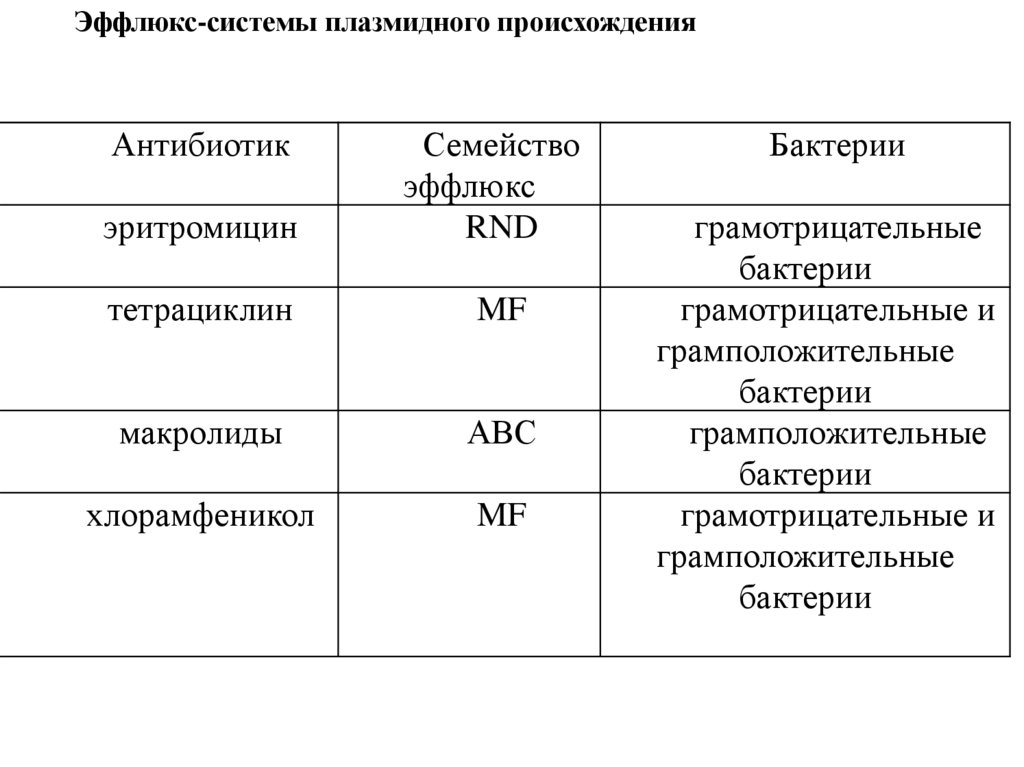
Эффлюкс-системы плазмидного происхожденияАнтибиотик
эритромицин
Семейство
эффлюкс
RND
тетрациклин
MF
макролиды
АВС
хлорамфеникол
MF
Бактерии
грамотрицательные
бактерии
грамотрицательные и
грамположительные
бактерии
грамположительные
бактерии
грамотрицательные и
грамположительные
бактерии
46.
Клеточный метаболизмСульфаниламиды и триметоприм блокируют различные
этапы одного метаболического пути бактерий – синтез
фолиевой кислоты, благодаря чему между ними отмечается
выраженный синергизм.
Сульфаниламиды, являющиеся структурным аналогом
парааминобензойной кислоты, являются конкурентными
ингибиторами дигидроптеоратсинтетазы.
Триметоприм подавляет активность дигидрофолатредуктазы.
Устойчивость связана с формированием метаболического
шунта. Устойчивость к триметоприму или сульфаниламидам
может
являться
результатом
приобретения
генов
дигидрофолатредуктазы
или
дигидроптеоратсинтетазы,
нечувствительной (или малочувствительной) к действию
антибиотика.
Следует отметить, что устойчивость к
триметоприму часто локализуется в составе транспозонов
(Tn7).
47.
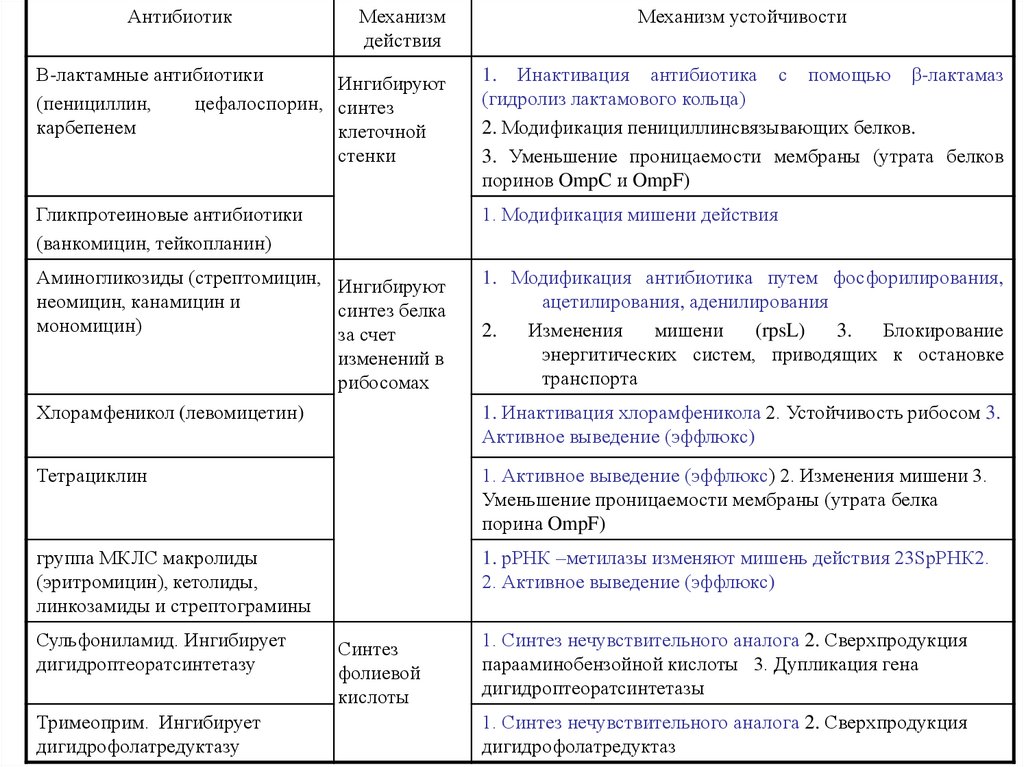
АнтибиотикМеханизм
действия
Механизм устойчивости
Β-лактамные антибиотики
Ингибируют
(пенициллин,
цефалоспорин, синтез
карбепенем
клеточной
стенки
1. Инактивация антибиотика с помощью β-лактамаз
(гидролиз лактамового кольца)
2. Модификация пенициллинсвязывающих белков.
3. Уменьшение проницаемости мембраны (утрата белков
поринов OmpC и OmpF)
Гликпротеиновые антибиотики
(ванкомицин, тейкопланин)
1. Модификация мишени действия
Аминогликозиды (стрептомицин, Ингибируют
неомицин, канамицин и
синтез белка
мономицин)
за счет
изменений в
рибосомах
Хлорамфеникол (левомицетин)
1. Модификация антибиотика путем фосфорилирования,
ацетилирования, аденилирования
2.
Изменения
мишени
(rpsL)
3.
Блокирование
энергитических систем, приводящих к остановке
транспорта
Тетрациклин
1. Активное выведение (эффлюкс) 2. Изменения мишени 3.
Уменьшение проницаемости мембраны (утрата белка
порина OmpF)
группа МКЛС макролиды
(эритромицин), кетолиды,
линкозамиды и стрептограмины
1. рРНК –метилазы изменяют мишень действия 23SрРНК2.
2. Активное выведение (эффлюкс)
Сульфониламид. Ингибирует
дигидроптеоратсинтетазу
Тримеоприм. Ингибирует
дигидрофолатредуктазу
Синтез
фолиевой
кислоты
1. Инактивация хлорамфеникола 2. Устойчивость рибосом 3.
Активное выведение (эффлюкс)
1. Синтез нечувствительного аналога 2. Сверхпродукция
парааминобензойной кислоты 3. Дупликация гена
дигидроптеоратсинтетазы
1. Синтез нечувствительного аналога 2. Сверхпродукция
дигидрофолатредуктаз
48.
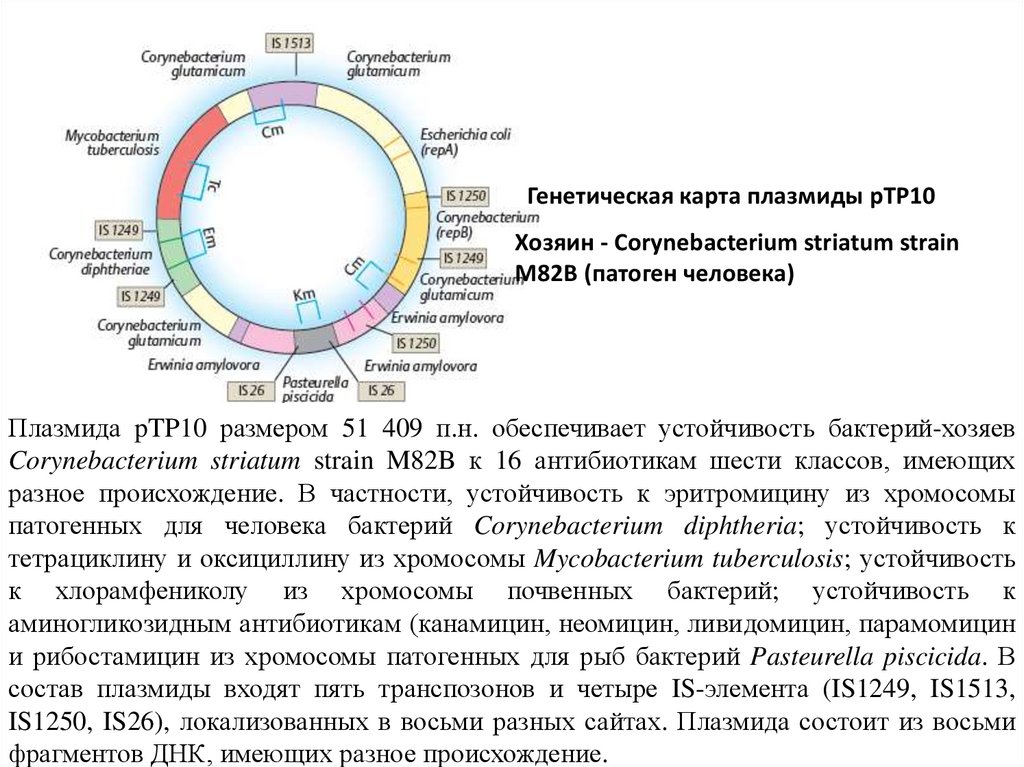
Генетическая карта плазмиды рТР10Хозяин - Corynebacterium striatum strain
M82B (патоген человека)
Плазмида pTP10 размером 51 409 п.н. обеспечивает устойчивость бактерий-хозяев
Corynebacterium striatum strain M82B к 16 антибиотикам шести классов, имеющих
разное происхождение. В частности, устойчивость к эритромицину из хромосомы
патогенных для человека бактерий Corynebacterium diphtheria; устойчивость к
тетрациклину и оксициллину из хромосомы Mycobacterium tuberculosis; устойчивость
к хлорамфениколу из хромосомы почвенных бактерий; устойчивость к
аминогликозидным антибиотикам (канамицин, неомицин, ливидомицин, парамомицин
и рибостамицин из хромосомы патогенных для рыб бактерий Pasteurella piscicida. В
состав плазмиды входят пять транспозонов и четыре IS-элемента (IS1249, IS1513,
IS1250, IS26), локализованных в восьми разных сайтах. Плазмида состоит из восьми
фрагментов ДНК, имеющих разное происхождение.
49.
Транспозоны имеют инвертированные концевые повторы.Внедрение транспозонов вызывает образование в сайте-мишени
прямых повторов фланкирующей ДНК.
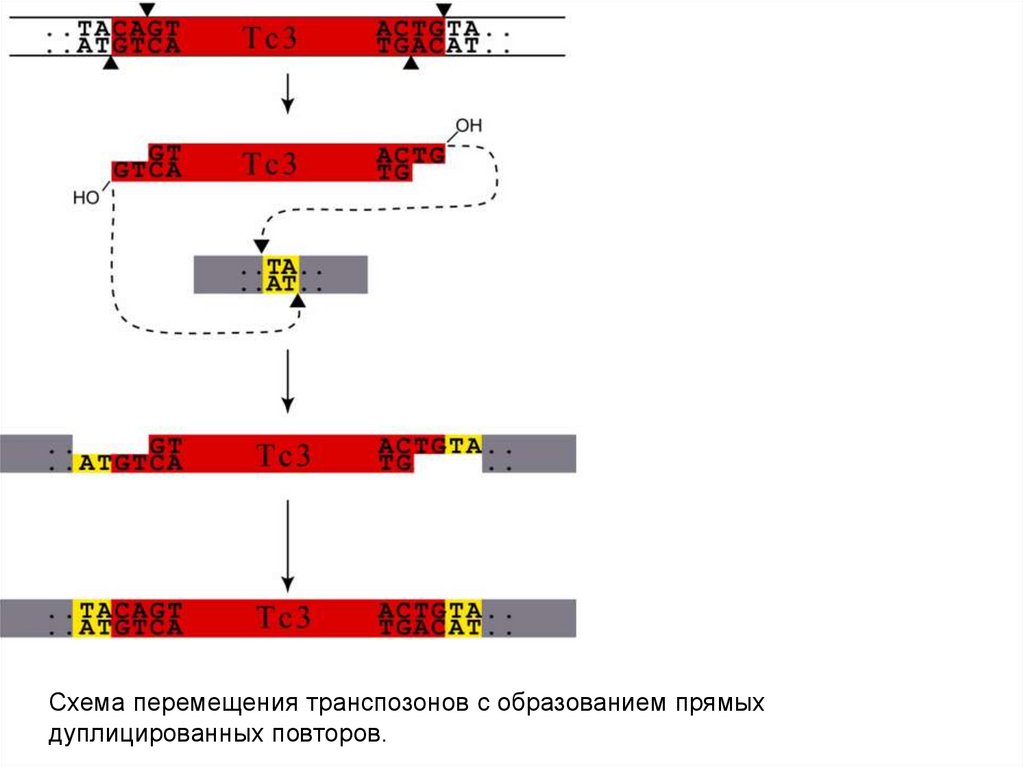
50.
51.
Схема перемещения транспозонов с образованием прямыхдуплицированных повторов.
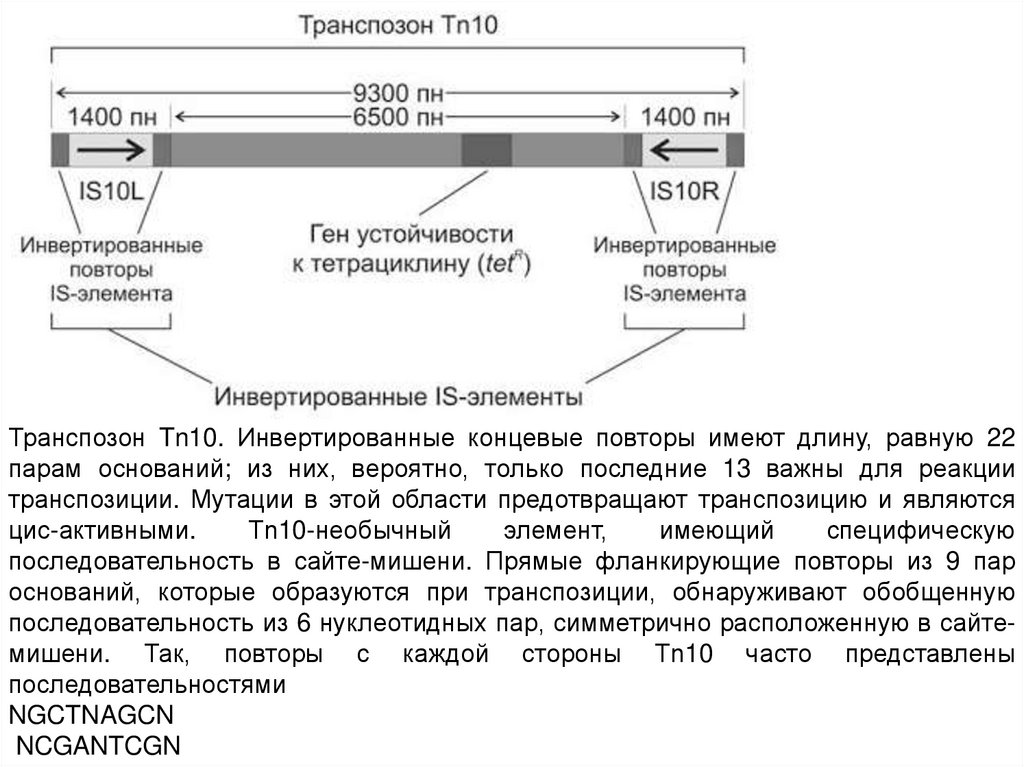
52.
Транспозон Tn10. Инвертированные концевые повторы имеют длину, равную 22парам оснований; из них, вероятно, только последние 13 важны для реакции
транспозиции. Мутации в этой области предотвращают транспозицию и являются
цис-активными.
Тn10-необычный
элемент,
имеющий
специфическую
последовательность в сайте-мишени. Прямые фланкирующие повторы из 9 пар
оснований, которые образуются при транспозиции, обнаруживают обобщенную
последовательность из 6 нуклеотидных пар, симметрично расположенную в сайтемишени. Так, повторы с каждой стороны Тn10 часто представлены
последовательностями
NGCTNAGCN
NCGANTCGN
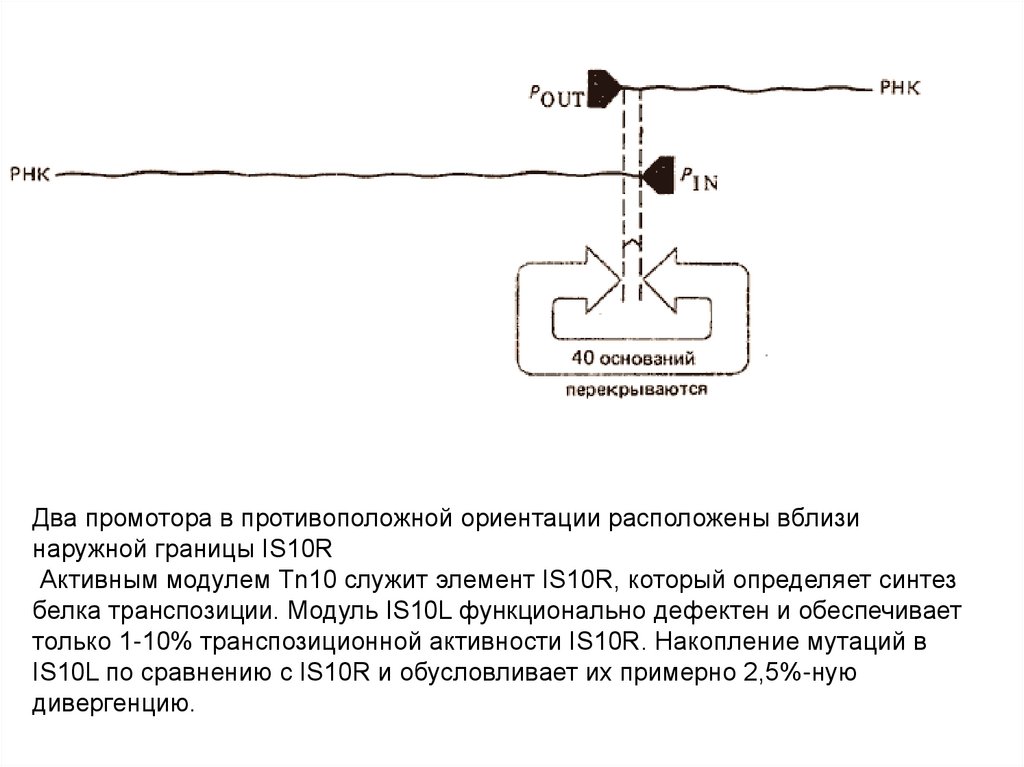
53.
Два промотора в противоположной ориентации расположены вблизинаружной границы IS10R
Активным модулем Тn10 служит элемент IS10R, который определяет синтез
белка транспозиции. Модуль IS10L функционально дефектен и обеспечивает
только 1-10% транспозиционной активности IS10R. Накопление мутаций в
IS10L по сравнению с IS10R и обусловливает их примерно 2,5%-ную
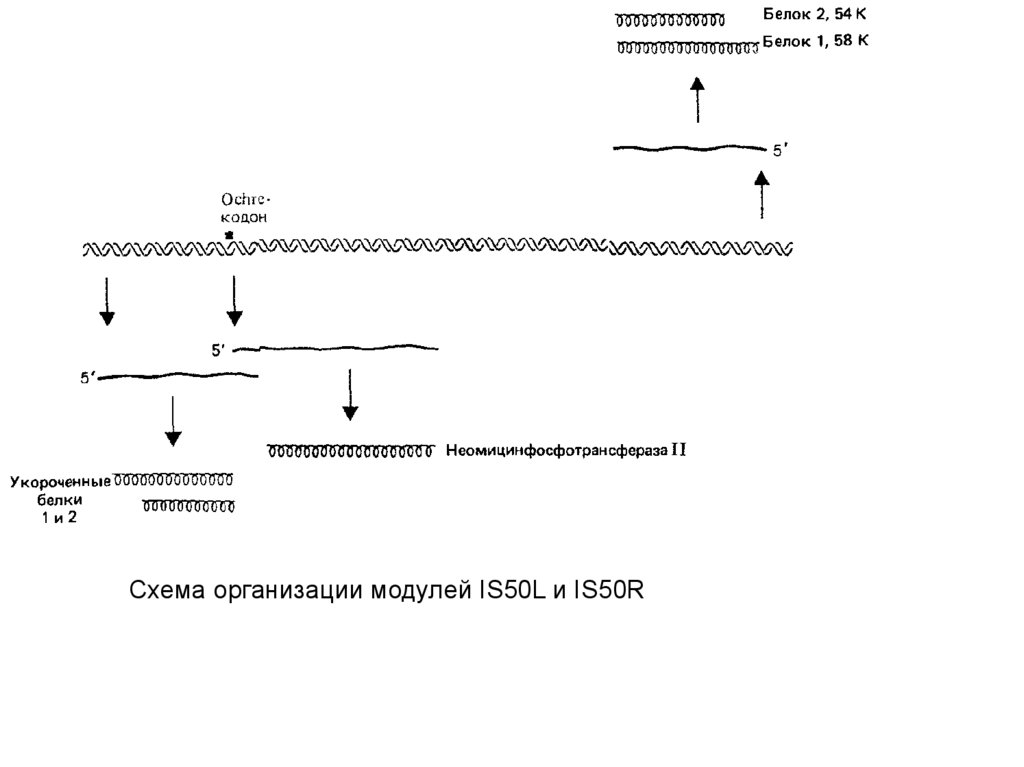
дивергенцию.