



 сигнальная система")

 сигнальной системы")













 и «глобального» (TIR1)")



 и растений (B)")
















, связанные с закрытием устьиц")

 biology
biologySimilar presentations:










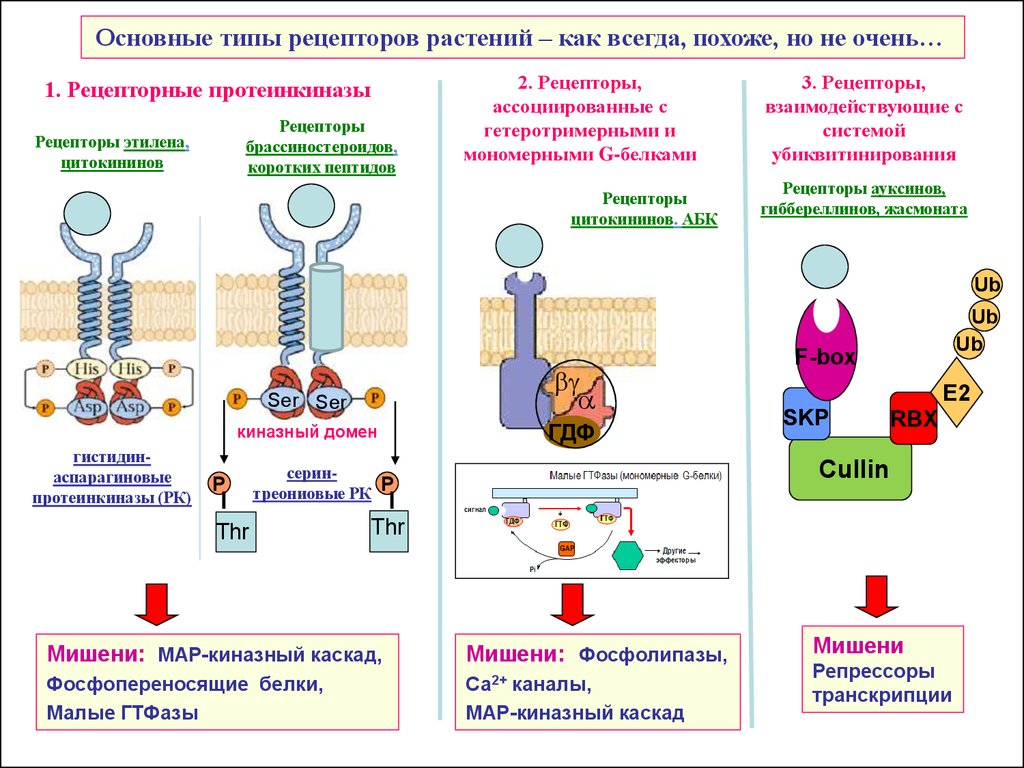
Основные типы рецепторов растений
1. И на все это надо как то реагировать...
2. Схема восприятия и трансдукции сигнала в изображении Бидструпа...
3. Три группы рецепторов сигналов в животной клетке
4.
Основные типы рецепторов растений – как всегда, похоже, но не очень…1. Рецепторные протеинкиназы
Рецепторы
брассиностероидов,
коротких пептидов
Рецепторы этилена,
цитокининов
2. Рецепторы,
ассоциированные с
гетеротримерными и
мономерными G-белками
Рецепторы
цитокининов. АБК
3. Рецепторы,
взаимодействующие с
системой
убиквитинирования
Рецепторы ауксинов,
гиббереллинов, жасмоната
Ub
bg
a
Ser Ser
киназный домен
гистидинаспарагиновые
протеинкиназы (РК)
P
ГДФ
F-box
E2
SKP
RBX
Cullin
серинтреониовые РК P
Thr
Thr
Ub
Ub
Thr
Мишени: МАР-киназный каскад,
Мишени: Фосфолипазы,
Фосфопереносящие белки,
Малые ГТФазы
Са2+ каналы,
МАР-киназный каскад
Мишени
Репрессоры
транскрипции
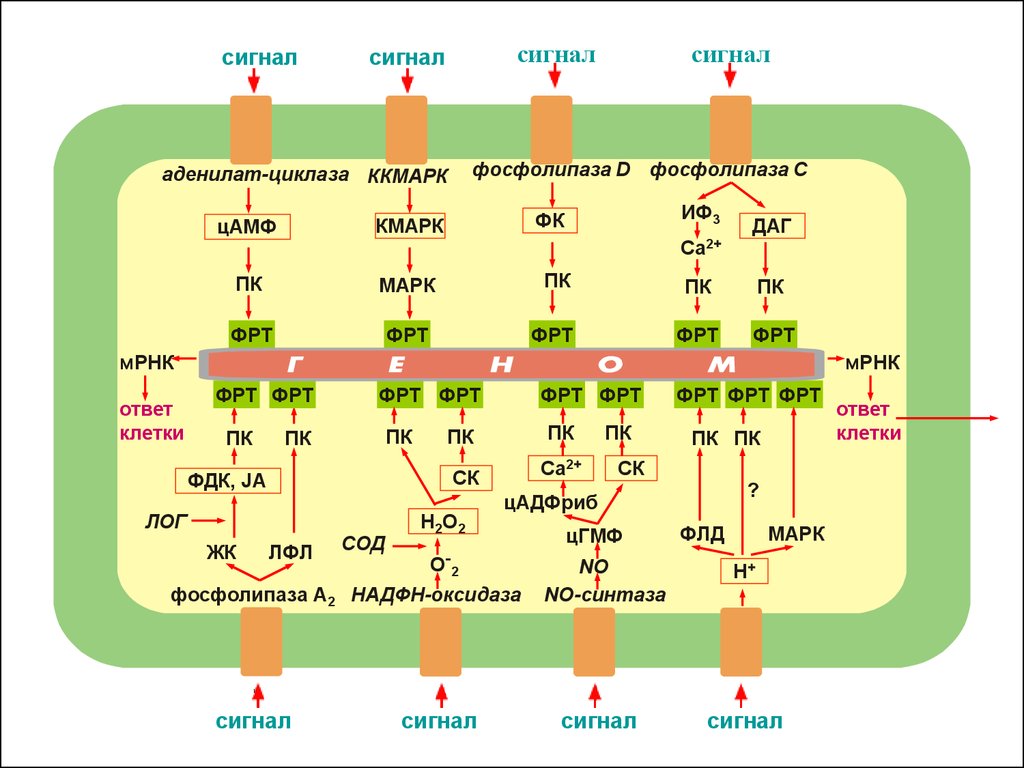
5. Вторичные мессенджеры сигналинга растительной клетки
6. Аденилат-циклазная (ц-АМФ-ная) сигнальная система
7. Работа каждой из фосфолипаз может приводить к образованию вторичных мессенджеров
Лизофософолипид8. Фосфолипаза С обеспечивает работу фосфоинозитольной (кальциевой) сигнальной системы
Кальмодулин9. «Депо» кальция в растительной клетке
10. Диацилглицерин также работает в качестве вторичного месенджера
11. Работа фосфолипазы D приводит к образованию фосфатидной кислоты – вторичного мессенджера фосфатидной сигальной системы.
Фосфатиднаякислота (PtdOH,
PA) играет
ключевую роль в
работе абсцизовой
кислоты по
закрыванию
устьиц
12. Работа фосфолипазы А2 проводит к образованию жасмонатов
Возможно, по этойпричине ряд авторов
относят жасмонаты
не к фитогормонам, а
ко вторичным
мессенджерам…
13. МАР-киназная сигнальная система и взаимодействие систем.
МАР-киназы –серин-треониновые киназы
В геноме Arabidopsis обнаружено
60 MAPKKK
10 MAPKK
23 MAPK.
Получается своеобразная
«сигнальная воронка»…
14.
сигналсигнал
сигнал
сигнал
фосфолипаза D фосфолипаза С
аденилат-циклаза ККМАРК
ИФ3
ФК
цАМФ
КМАРК
ПК
МАРК
ПК
ПК
ПК
ФРТ
ФРТ
ФРТ
ФРТ
ФРТ
Са2+
ДАГ
мРНК
ответ
клетки
мРНК
ФРТ ФРТ
ПК
ФРТ ФРТ
ПК
ПК
ФДК, JA
ЛОГ
ЖК
ЛФЛ
СОД
ФРТ ФРТ
ПК
ПК
СК
Са2+
Н2О2
-
О-2
сигнал
ПК ПК
СК
?
цАДФриб
фосфолипаза А2 НАДФН-оксидаза
сигнал
ПК
ФРТ ФРТ ФРТ
цГМФ
ФЛД
МАРК
NO
NO-синтаза
Н+
сигнал
сигнал
ответ
клетки
15. Убиквитины – метки смерти….
Убиквитин – небольшой белок, 76аминокислот.
Убиквитин (желтый) активизируется
убиквитин-активирующим
ферментом-E1 (красный).
Терминальный Gly на С-конце
убиквитина формирует тиоэфир с
цистеиновым остатком E1.
Активированный убиквитин
перемещается на один из нескольких
убиквитин-связывающих ферментов,
E2 (зеленый), снова формируя
тиоэфир. После этого убиквитин
перемещается на субстрат (белокмишень, голубой), связываясь с
внутренним Lys остатком этого белка.
Эта реакция обычно требует еще
одного фактора – убиквитин-белоклигазы E3 (синяя), который связывает
E2 с белком-мишенью. Обычно на
белок-мишень «навешивается»
несколько убиквитинов.
Убиквитированные белки
направляются в протеасому для
развала до аминокислот…
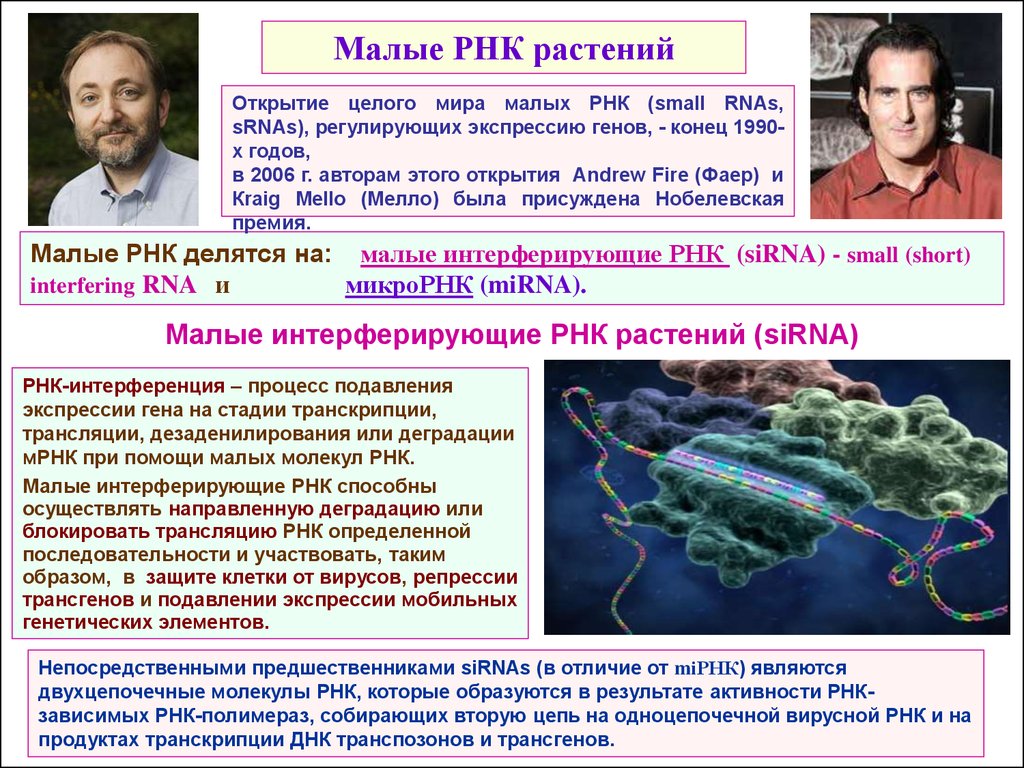
16.
Малые РНК растенийОткрытие целого мира малых РНК (small RNAs,
sRNAs), регулирующих экспрессию генов, - конец 1990х годов,
в 2006 г. авторам этого открытия Andrew Fire (Фаер) и
Кraig Mello (Мелло) была присуждена Нобелевская
премия.
Малые РНК делятся на: малые интерферирующие РНК (siRNA) - small (short)
interfering RNA и
микроРНК (miRNA).
Малые интерферирующие РНК растений (siRNA)
РНК-интерференция – процесс подавления
экспрессии гена на стадии транскрипции,
трансляции, дезаденилирования или деградации
мРНК при помощи малых молекул РНК.
Малые интерферирующие РНК способны
осуществлять направленную деградацию или
блокировать трансляцию РНК определенной
последовательности и участвовать, таким
образом, в защите клетки от вирусов, репрессии
трансгенов и подавлении экспрессии мобильных
генетических элементов.
Непосредственными предшественниками siRNAs (в отличие от miРНК) являются
двухцепочечные молекулы РНК, которые образуются в результате активности РНКзависимых РНК-полимераз, собирающих вторую цепь на одноцепочечной вирусной РНК и на
продуктах транскрипции ДНК транспозонов и трансгенов.
17.
siRNA растений и низших животныхорганизмов является важным
звеном внутриклеточного
иммунитета, позволяющего
распознавать и быстро расщепить
чужую РНК.
Если в клетку проникает РНК–
содержащий вирус, такая система
защиты осуществляет
направленную деградацию вирусной
РНК.
Если же вирус содержит ДНК,
система siRNA будет мешать ему
производить вирусные белки (так
как необходимая для этого мРНК
будет распознаваться и разрезаться).
siRNA из транспозонов или
трансгенных промоторов –
контролируют сайленсинг
транспозонов или трансгенов
путем модификации хроматина.
18. Микро РНК - посттранскрипционные репрессоры экспрессии генов.
Фактически - антисенс-регуляторы других РНКМогут быть страшными ядами…
Микро РНК (miRNA)
- 21-23 нуклеотида
формируются из
первичного транскрипта
(pri-mRNA,
70 нуклеотидов), который
находится в виде
«шпильки».
В геноме арабидопсса
около 100 локусов
pri-mRNA.
Процессинг у растений
отличается от животных,
у которых в цитозоле
работает фермент Dicer
(«шинкующий,
нарезающий кубиками»).
У растений - Dicer like
proteins, DLPs,
расположенные в ядре.
RISC – РНКнндуцируемый
«молчащий» комплекс.
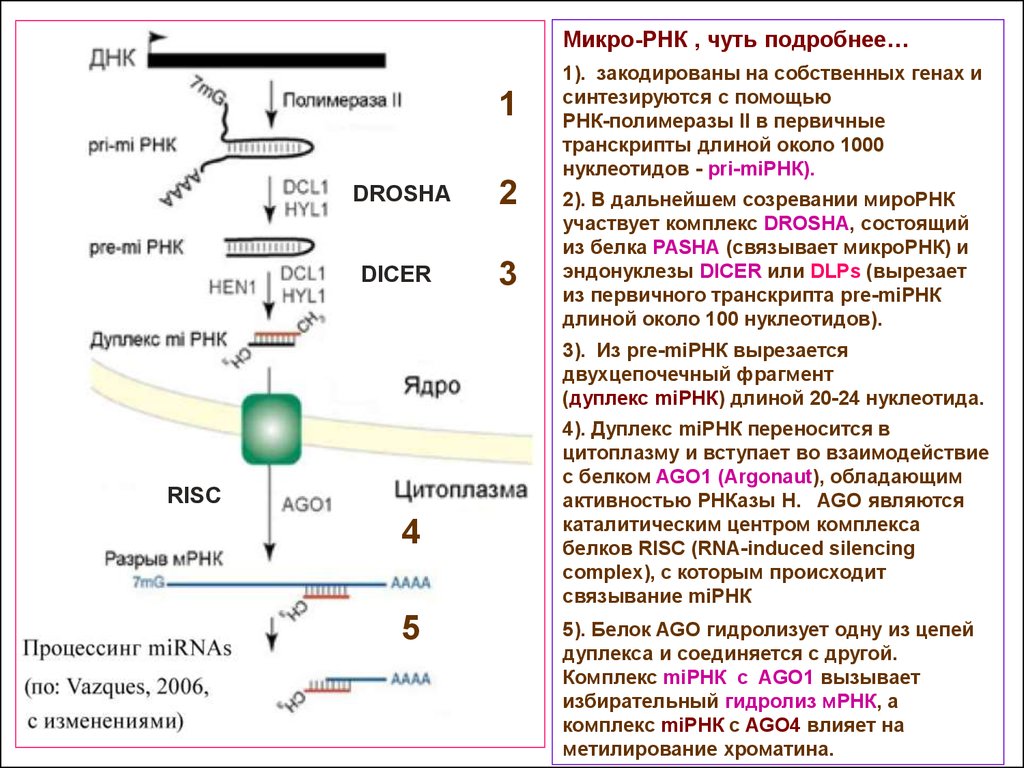
19.
Микро-РНК , чуть подробнее…1
DROSHA
2
DICER
3
1). закодированы на собственных генах и
синтезируются с помощью
РНК-полимеразы II в первичные
транскрипты длиной около 1000
нуклеотидов - pri-miРНК).
2). В дальнейшем созревании мироРНК
участвует комплекс DROSHA, состоящий
из белка PASHA (связывает микроРНК) и
эндонуклезы DICER или DLPs (вырезает
из первичного транскрипта pre-miРНК
длиной около 100 нуклеотидов).
3). Из pre-miРНК вырезается
двухцепочечный фрагмент
(дуплекс miРНК) длиной 20-24 нуклеотида.
RISC
4
5
4). Дуплекс miРНК переносится в
цитоплазму и вступает во взаимодействие
с белком AGO1 (Argonaut), обладающим
активностью РНКазы Н. AGO являются
каталитическим центром комплекса
белков RISC (RNA-induced silencing
complex), с которым происходит
связывание miРНК
5). Белок AGO гидролизует одну из цепей
дуплекса и соединяется с другой.
Комплекс miРНК с AGO1 вызывает
избирательный гидролиз мРНК, а
комплекс miРНК с AGO4 влияет на
метилирование хроматина.
20.
МикроРНК комплементарносвязывается с мРНК генов-мишеней,
что приводит к разрушению этой мРНК
(полная комплементарность), или к
ингибированию трансляции (частичная
комплементарность).
Таким способом микроРНК могут
контролировать уровень экспрессии
почти половины известных генов,
кодирующих синтез факторов
транскрипции.
МикроРНК могут являться
своеобразными «переключателями»
программы развития клетки.
МикроРНК могут регулировать:
• Эмбриогенез,
• Цветение,
• Морфогенез листа,
• Развитие семени,
• Формирование боковых корней
• Пролиферацию клеток,
• Трансдукцию ИУК-сигнала ,.
Обнаружены микро-РНК, нацеленные
на мРНК ферментов.
Содержание микроРНК резко
возрастает в условиях стресса.
21.
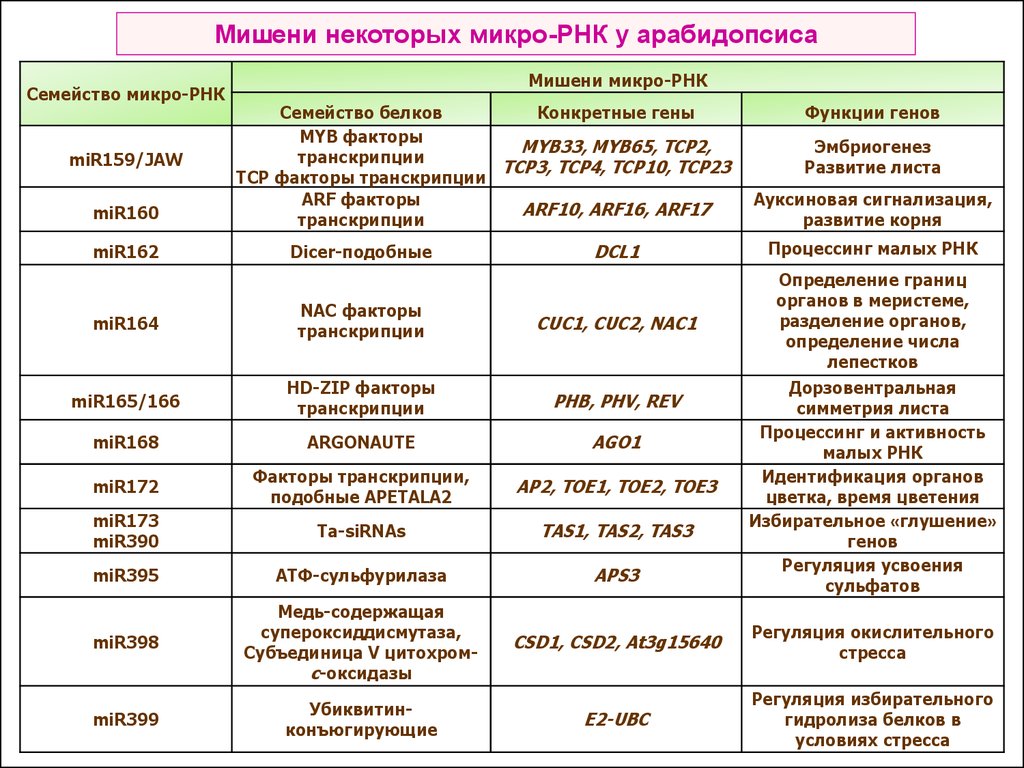
Мишени некоторых микро-РНК у арабидопсисаСемейство микро-РНК
miR159/JAW
miR160
miR162
Мишени микро-РНК
Семейство белков
Конкретные гены
MYB факторы
MYB33, MYB65, TCP2,
транскрипции
TCP3, TCP4, TCP10, TCP23
TCP факторы транскрипции
ARF факторы
ARF10, ARF16, ARF17
транскрипции
Dicer-подобные
Функции генов
Эмбриогенез
Развитие листа
Ауксиновая сигнализация,
развитие корня
DCL1
Процессинг малых РНК
Определение границ
органов в меристеме,
разделение органов,
определение числа
лепестков
miR164
NAC факторы
транскрипции
CUC1, CUC2, NAC1
miR165/166
HD-ZIP факторы
транскрипции
PHB, PHV, REV
miR168
ARGONAUTE
AGO1
miR172
Факторы транскрипции,
подобные APETALA2
AP2, TOE1, TOE2, TOE3
miR173
miR390
Ta-siRNAs
TAS1, TAS2, TAS3
miR395
АТФ-сульфурилаза
APS3
miR398
Медь-содержащая
супероксиддисмутаза,
Субъединица V цитохромс-оксидазы
CSD1, CSD2, At3g15640
Регуляция окислительного
стресса
miR399
Убиквитинконъюгирующие
E2-UBC
Регуляция избирательного
гидролиза белков в
условиях стресса
Дорзовентральная
симметрия листа
Процессинг и активность
малых РНК
Идентификация органов
цветка, время цветения
Избирательное «глушение»
генов
Регуляция усвоения
сульфатов
22. Рецепторы ауксинов – экстра- или внутриклеточные?
1. ABP1? Белок семейства GPCR (G-Proteins Coupled Receptors)Около 22 кDa. У кукурузы – 5 генов. На N-конце - сигнал для транспорта в
ЭР (38 а-к), на С-конце – КDEL, на N- конце – сайт связывания с ИУК:
His-Arg-His-Ser-Cys-Glu. Сайт гликозилирования (Asp33-Thr-Thr), До 90%
белка - в ЭР.
2. Транспортеры ИУК? Транспортеры ИУК в клетку (4 гена - AUX1 и
др.) из клетки (8 генов – PIN1, PIN2 и др).
3. ABP 60 kDa ? Гликозидаза цитокининовых конъюгатов. Взаимодействие
фитогормонов?
4. ABP 24-25 kDa? Глютатион-5-трансфераза (GST).
GST контролирует уровень глютатиона, влияет на уровень жасмонатов,
перекиси водорода.
Множественность рецепторов?
ИУК – особый гормон: специфичный полярный транспорт (от клетки к
клетке), эндогенный ритм образования и транспорта.
23. Система трансдукции ауксинового сигнала: AUX/IAA - ARF.
Aux/IAA – смейство (более 25) короткоживущихядерных ауксин-индуцируемых белков.
ARFs - Auxin-respons factor (ауксиновые
факторы транскрипции). Более 20 белков.
Aux/IAA – белки образуют
разнообразные димеры как
между собой, так и с ARF. Разные
димеры регулируют
транскрипцию генов с AuxRE
(Auxin-respons element - TGTCNC).
Димеры ARF – активируют,
Aux/IAA+ARF – тормозят.
Ауксин одновременно активирует
транскрипцию Aux/IAA и их
распад путем убиквитирования
и деградации в 26S протеосоме.
24. Существует как минимум два рецептора ауксинов. Один из них активирует убиквитинирование
Рецептор ауксина TIR1 (transportinhibitor response 1) имеет F-box мотив
(около 50 аминокислот для белокбелковых взаимодействий) и LRR
участок.
Он является компонентом
специфичного E3-убиквитинлигазного комплекса SCFTIR1 .
Когда ауксин связывается с SCFTIR1, то
он активирует его, в результате
происходит убиквитинирование и
протеолитическая деградация
AUX/IAA репрессорных белков.
25. Работа рецептора ауксина ядерной локализации.
А. При отсутствии (низкихконцентрациях) ауксина,
транскрипция аусинрегулируемых генов,
активирующихся ARF-белками,
блокирована репрессорами
транскрипции Aux/IAA .
В. При наличии (высоких
концентрациях) ауксин
связывается с белком TIR1,
или с другими AFB белками,
которые входят в SCFTIR1
комплекс (RBX1-Cullin-ASK1TIR1) убиквитирования.
Связывание ауксина
стимулирует взаимодействие
Aux/IAAs с SCFTIR1 что ведет к
убиквитированию Aux/IAAs
белков, и их разрушению в 26S
протеасоме. Это снимает
ингибирование ARFs и
запускает транскрипцию.
С. Димеризация ARF усиливает
транскрипцию.
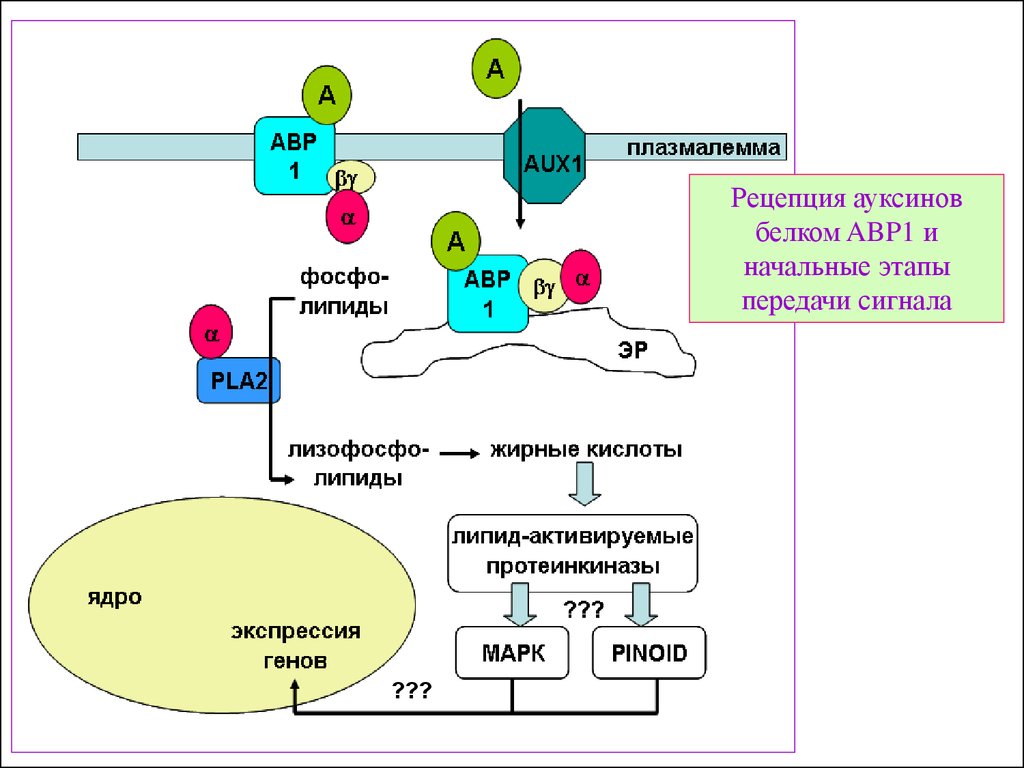
26. Принципиальная схема работы двух рецепторов ауксинов – «быстрого» (АВР1) и «глобального» (TIR1)
1. Активация гена:В норме промотор гена
Н+-АТФ-зы «закрыт»
репрессорным
комплексом, который
является и рецептором
ауксина. Ауксин вызывает
протеолитическую
деградацию репрессора.
Активация транскрипции
ведет к увеличению Н+АТФ-зы на плазменной
мембране.
2. Перемещение Degradation
и
встраивание белка:
Ауксин-связывающий
белок (рецептор ауксина )
ABP1 может увеличивать
перемещение H +-ATФзы
к плазмалемме.
3. Стабилизация H +ATФзы:
Обработка ауксином
вызывает сохранение Н+АТФ-зы в плазмалемме.
Это тоже может быть
обусловлено ABP1
27.
Рецепция ауксиновбелком ABP1 и
начальные этапы
передачи сигнала
???
28. Система трасдукции гиббереллинового сигнала очень похожа на ауксиновую…
В генах-мишениях: впромторной области (-289)
- GARE или GARC
• GARE: часто
UTAACAUANTCYGG (N
– A/G, Y – C/T)
• GARC: три участка:
пиримидиновый (-171),
TAACAAA- и TATCCAC
(-115)
Транскрипция гиббереллинзависимых генов
регулируется GA зависимой
деградацией DELLA
репрессоров. Гиббереллин
связывается с GID1 белком,
что ведет к его
взаимодействию с SLR1.
SCFGID2 комплекс
осуществляет
убиквитинирование
SLR1 и его деградацию.
Структура белков-репрессоров
гиббереллинового сигналинга.
DELLA – 5 аминокислот
(аспарагиновая кислота D,
глутамновая кислота E, лейцин
L, лейцин L, аланин A)
29. Сигнальная система гиббереллинов
1. GA1 из эмбриоида связывается с гипотетическиммембранным рецептором на поверхности клетки
алейронового слоя.
2. Комплекс GA-рецептор взаимодействует с
гетеротримерным G-белком и запускает две независимых
цепи трансдукции сигнала - Са 2+ -независимую и Са 2+ зависимую
3. Ca2+-независимый путь, вовлекающий cGMP и ряд
других компонентов, завершается активацией F-белка компонента SCF-убиквитин-лигазного комплекса.
4. GA1 может также войти в клетку непосредственно и
связаться с альтернативным рецепторным белком, который
расположен преимущественно в ядре.
5. Активизированный F-белок связывается DELLAдоменом белка репрессора, который блокирует
транскрипцию GAMYB генов или других активаторов
транскрипции.
6. Репрессор убиквитинируется через SCF-убиквитинлигазный комплекс и разрушается в протеасоме.
7. Деградация репрессора приводит к транскрипции
GAMYB и других генов раннего ответа.
8. Вновь синтезируемый GAMYB белок перемещается в
ядро и связывается с промотороами генов α-амилазы и
других гидролитических ферментов, запуская их
транскрипцию (9).
10. Амилаза и другие гидролазы синтезируются на
шероховатом ER, затем перемещаются в AG (11).
12. Для активации секреторного пути транспорта этих
белков необходима стимуляция гиббереллином кальциевой
сигнальной системы - Са 2+-зависимый путь
30.
Как нашли еtr1 –мутант у арабидопсиса- C2H4 + C2H4
31. Двухкомпонентные рецепторные системы: гистидинкиназы микроорганизмов (A) и растений (B)
У животныхгистидинкиназы не
обнаружены.
Консервативный участок
H (гистидин),
N (аспарагин),
G (глицин),
F (фенилаланин)
HPt (histidine
phosphotransfer protein) –
гистидинфосфотрансфкрный белок
- «дополнительный»
переносчик..
Введение дополнительных
переносчиков обусловлено,
скорее всего, размерами
эукариотической клетки и
необходимостью
передавать сигнал в
ядро…
32. Гистидинкиназы, схема трансдукции сигнала
Нpt – гистидин-фосфотрансферный белок, дляарабидопсиса – АНР.
ARRs – регуляторы ответа (Arabidopsis
response regulators) – около 20 штук..
Два типа ARR . Тип А содержит только
ресиверный домен, содержащий аспартат.
Тип В содержит дополнительный «выходной»
домен – трансфактор.
H – гистидин, D – аспартат.
33. Рецепторы цтокинина и этилена – гистидинкиназы.
Схемы строения рецепторовцитокининов и этилена
В геноме арабидопсиса 16 генов
гистидинкиназ. Из них
3 – рецепторы цитокининов,
5 – рецепторы этилена,
5 – фитохромы - но это
«бывщие» гистидинкиназы...
34. Сигнальная система цитокининов
1. Цитокинин связывается с CRE1, который, вероятно,существует как димер на мембране ЭР. Цитокинин
связывается с CHASE-доменом рецептора Две других
гибридных сенсорных киназы (AHK2 и AHK3)
содержащие CHASE-домены, также вероятно, являются
рецепторами цитокининов в Arabidopsis.
2. Связывание цитокинина с рецепторами активизирует
деятельность рецепторов в качестве гистидинкиназ.
Фосфат перемещается на аспартат (D) в слитый
ресиверный домен.
3. Фосфат перемещается на консервативный гистидин в
AHP белке.
4. Фосфорилирование «заставляет» AHP белок
перемещаться в ядро, где он передает фосфат на остаток
аспартата, находящийся в ресивером домене ARR типа В
5. Фосфорилирование ARR типа В активизирует его output
домен и взывает транскрипцию генов ARR типа А,.
6. ARR типа А, вероятно, также могут быть
фосфорилированы и AHP белками.
7. Фосфорилированые ARR типа А взаимодействуют с
различными эффекторами и вызывают «цитокининовыйе»
ответы клетки
ЭР
35. Этилен работает «наоборот» - его присутствие инактивирует рецепторы…
Пять рецепторов этилена у арабидопсиса. У трех их нихотсутствует «главный» гистидин…
GAF –
консервативный
cGMP-связывающий
домен.
Найденный у
различных белков.
Обычно его функции –
небольшой
регуляторный участок,
связывающий
молекулы.
36.
Схема трансдукции этиленового сигнала1. RAN1 белок с кофактором, содержащим медь
включается в рецептор этилена, ETR1, ETR2,
ERS1, ERS2 (ethylene resistant), EIN4 (ethylene
insensitive) находящийся на мембране ЭР или АГ.
Белок RTE активирует процесс.
2. В отсутствии этилена его рецепторы
связываются с киназой CTR1 (constitutive triple
response 1) и активизируют ее работу, что приводит
к репрессии ответа на этилен – выключает работу
белка EIN2, факторов транскрипции EIN3/EIL1 и
компонентов МАР-киназного каскада МКК9 и
МРК3/6.
3. Связывание этилена с рецептором (в виде
димера) приводит к инактивации рецептора, к
диссоциации и инактивация CTR1 за счет
связывания с фосфатидными кислотами.
Рецепторы этилена разваливаются в протеасоме..
4. Инактивация CTR1 «снимает запрет» с работы
сигнального каскада.
5. Активация EIN2 включает семейство факторов
транскрипции EIN3/EIL1 (или предотвращает их
распад через убиквитинирование?), которые в
свою очередь вызывают экспрессию
трансфакторов ERF1. Активация этого
транскрипционного каскада ведет к
крупномасштабным изменениям в экспрессии
генов «этиленового ответа».
6. Белки EBF запускают убиквитирование и
протеолиз трансфакторов EIN3/EIL1.
Эндоррибонуклеаза EIN5/XRN негативно
регулирует экспрессию генов EBF
37.
Схема трансдукции этиленового сигналаCTR1 - Ser\Thr протеинкиназа, негативный регулятор этиленового сигналлинга
Протеинкиназа, MAPKKK, гомолог Raf протеинкиназ млекопитающих. «Анитпод» обычных
MAPKKK – киназы «нижних» уровней она не активирует, а ингибирует и фосфатидные кислоты
ее не стимулируют, а выключают. В отсутствии этилена связана с его рецепторами, активна и
ответа на этилен нет. При связывании этилена CTR1 «уходит» из комплекса с рецепторами,
связывается с фостфатидными кислотами, деградирует - происходит запуск ответа на этилен
EIN2 - позитивный регулятор этиленового сигналлинга,
Белок с неизвестной функцией, у арабидопсиса кодируется одним геном. N-концевой
трансмембранный домен гомологичен металлотранспортерам Nramp, но ничего не
транспортирует.. С-концевой цитоплазматический домен – важен для функции, но неясно как…
EIN3, EIL1 и EIL2 - этилен-зависимые транскрипционные факторы
Позитивные регуляторы этиленового сигналинга, локализуются в ядре, короткоживущие белки
(время полужизни – 30 мин), ответ на этилен регулируется стабильностью EIN3 белков, но не
уровнем их экспрессии. Индуцируют экспрессию гена ERF1 (Ethylene Response Factor)
EIN5/XRN4 – рибонуклеаза, регулирует уровень транскриптов EBF1/2 - компонентов
убиквитин-лигазного комплекса
EIN5/XRN4
ETR
CTR1
EIN2
MAPK
EBF1, 2
EIN3,
EIL1, 2
Контроль
развития
ERF1
Ответ на
стресс
38. Сигнальные системы цитокинина и этилена взаимодействуют.
A. При низких концентрациях лиганда,?
этиленовые рецепторы активны и
стимулируют негативный регулятор
CTR1, который вызывает деградацию
EIN3
B. Связывание этилена инактивирует
рецепторы, они перестают
стимулировать CTR1-зависимое
ингибирование EIN2. Это предотвращает
деградацию EIN3, который запускает
транскрипцию этилен-зависимых генов.
C. Связывание цтокинина с рецептором
вызывает его автофосфорилирование,
сопровождаемое переносом фосфата (P)
на гистидин фосфат-переносящего белка
(AHP), и далее на регулятор ответа
(ARR), что приводит к транскрипции
цитокинин-зависимых генов.
Возможно взаимодействие между
этиленоым и цитокининовым сигналом
через перекрестное фосфорилирование.
39. BRI1: LRR-RLK - рецептор брассиностероидов.
LRR-RLK: leucine rich repeat receptor-likekinase - группа RLK, у Arabidopsis более 170
генов. LRR участок - из повторяющихся
лейцин-обогащенных фрагментов из 24 а-к.
На N-конце – «лейциновая молния»
BRI1 - в плазмалемме,
экстрацеллюлярный домен из 25 неполных
RLL. Между 21 и 22 повторами - «остров»
из 70 а-к, связывание брассиностероидов.
На С-конце – киназный домен, серинтреониновая киназа.
Автофосфорилирование происходит после
связывания с BL.
LRR рецепторы.
А. Сравнение четырех типов
LRR рецепторов, найденных в
растениях и животных:
BRI1 - Arabidopsis рецептор
брассиностероидов (BR)
CLV1 - рецептор участвующий в
связвании маленького пептида
CLV3, который определяет
размер меристемы у Arabidopsis;
Cf-9, рецептор участвующий в
ответе растения на атаку
патогенна у томатов
Toll - рецептор разметки
эмбриона и врожденного
иммунитета у дрозофилы.
B. Механизм восприятия BR
сигнала .
В норме BRI1 и BAK1 LRR
киназы – неактивные
гомодмеры. После связывания с
брассиностероидом BRI1 и BAK1
агрегируются, в результате чего
автофосфориллируются. Это
ведет к репрессии ингибитора
BIN2, и индукции BR-ответа
40. Сигнальная система брассиностероидов.
A. При отсутствиибрассиностероидов (BL),
BIN2-киназа быстро
фосфорилирует
брассиностероидзависимые регуляторы
транскрипции, BES1 и
BZR1, что приводит к их
убиквитированию и
деградации в 26Sпротеасоме
B. При связывании BR c
мембранным
гетеродимером BRI1-BAK1,
что ингибирует BIN2 пока
еще не установленным
способом. Это ведет к
накоплению
дефосфориллированных
BES1 и BZR1 в ядре, где
они регулируют
транскрипцию BRрегулируемых генов, активируя, либо запрещая
транскрипцию.
41. Брассиностериоды как активируют транскрипцию некоторых генов, так и тормозят, в том числе гены ферментов собственного синтеза.
42. Рецептор брассиностероидов одновременно является и рецептором системина
В. У дрозофилы,протеаза
(изогнутая
стрелка)
расщепляет
пропетид,
Образуется
полипептид
Spatzle, который
затем
связывается с
Toll-рецептором.
«Выбор» между
регулированием
развития
эмбриона или
врожденным
иммунным
определяется
регулируемым
стадией развития
расщеплением
pre-Spatzle белка.
А. У томатов, BRI1 рецептор (tBRI1) может связаться как с BR, так и с неболишим пептидом
системином. Системин образуется в результате процессинга из большего полипептида за счет работы
неохарактеризованной протеазы (изогнутая стрелка). Связывание системина c tBRI1, приводит к
гетеродимеризации tBRI1 с неохарактеризованным LRR белоком (?), вызывая защитный ответ.
Напротив, если tBRI1 связывается с BR, это ведет к BRI-BAK1 димеризациии и BR-зависимому росту.
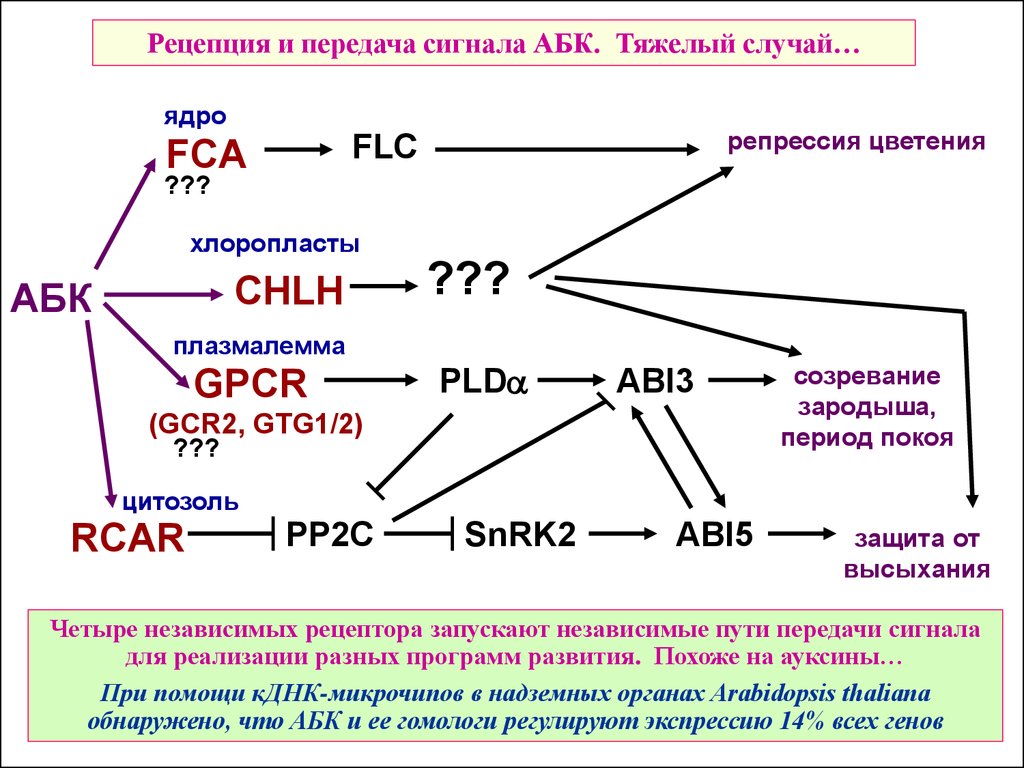
43.
Рецепция и передача сигнала АБК. Тяжелый случай…ядро
репрессия цветения
FLC
FCA
???
хлоропласты
АБК
CHLH
???
плазмалемма
GPCR
PLDa
ABI3
(GCR2, GTG1/2)
???
созревание
зародыша,
период покоя
цитозоль
RCAR
PP2C
SnRK2
ABI5
защита от
высыхания
Четыре независимых рецептора запускают независимые пути передачи сигнала
для реализации разных программ развития. Похоже на ауксины…
При помощи кДНК-микрочипов в надземных органах Arabidopsis thaliana
обнаружено, что АБК и ее гомологи регулируют экспрессию 14% всех генов
44. FCA – ядерный рецептор АБК
Сигнальная система АБК, регулирующая время цветения – внутриклеточный рецептор - ??FCA (Flowering time Control protein A) –
РНК-связывающий ядерный белок,
который запускает цветение, снимая
действие FLC (Flowering Locus C) мощного ингибитора цветения. FCA
афинно и специфично связывется с
АБК.
Без АБК, FCA образует совместно с
фактором полиаденилирования РНК FY
(Flowering Locus Y) комплекс
процессинга РНК, который
предотвращает образование mРНК
ингибитора FLC. Растение цветет.
При наличии АБК она связывается c Сконцевым участком FCA, рядом с
местом взаимодействия с FY. Это
разрушает их взаимодействие, что ведет
к накоплению FLC и задержке
цветения.
Транскрипция SOC1 происходит только
в растениях с низкими уровнями FLC
45.
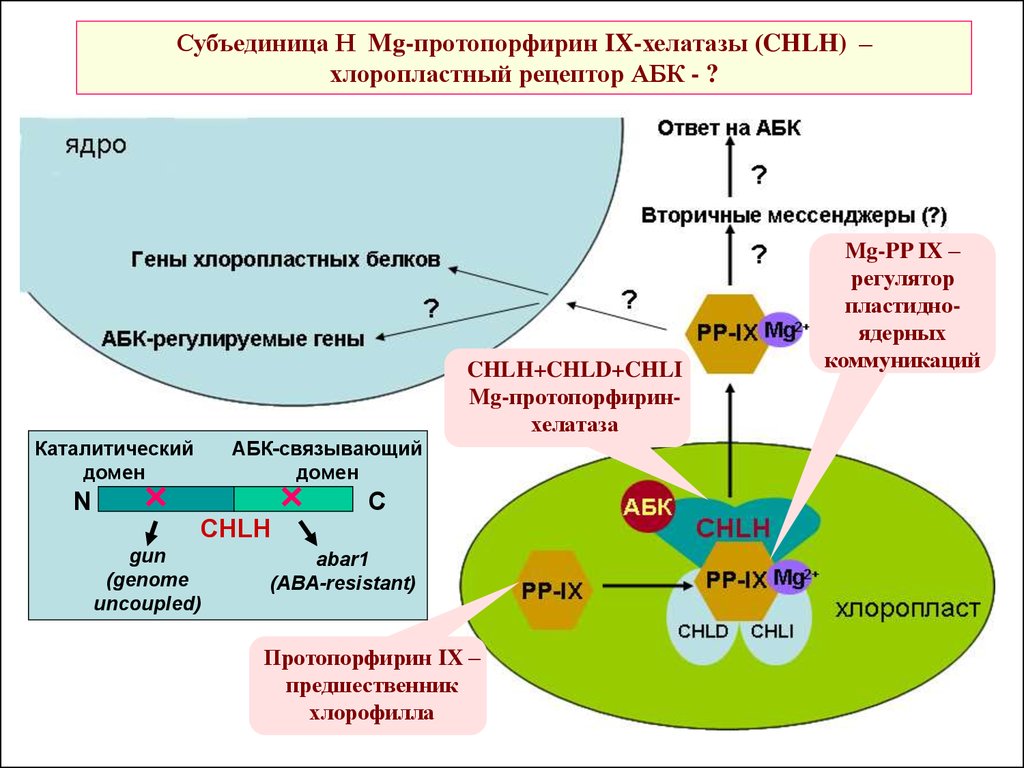
Субъединица Н Mg-протопорфирин IX-хелатазы (CHLH) –хлоропластный рецептор АБК - ?
CHLH+CHLD+CHLI
Mg-протопорфиринхелатаза
Каталитический
домен
АБК-связывающий
домен
N
C
CHLH
gun
(genome
uncoupled)
abar1
(ABA-resistant)
Протопорфирин IX –
предшественник
хлорофилла
Mg-PP IX –
регулятор
пластидноядерных
коммуникаций
46.
GPCR (G-Protein Coupled Receptors) – плазмалемные рецепторы АБКФосфолипаза Da1,
синтезирует
фосфатидные
кислоты (РА)
Киназа
фосфатидных
кислот и
липидфосфатфосфатаза
ГТФ
Липидные
сигнальные
молекулы
ГДФ
ТФ с В3 доменом:
ABI3 (а также
LEC2, FUS3)
Рецептор,
связанный с
комплексом G
белков
гистондеацетилаза
HDAC
Репрессор
транскрипции: ТФ с
В3 доменом
Гены запасных
белков
47.
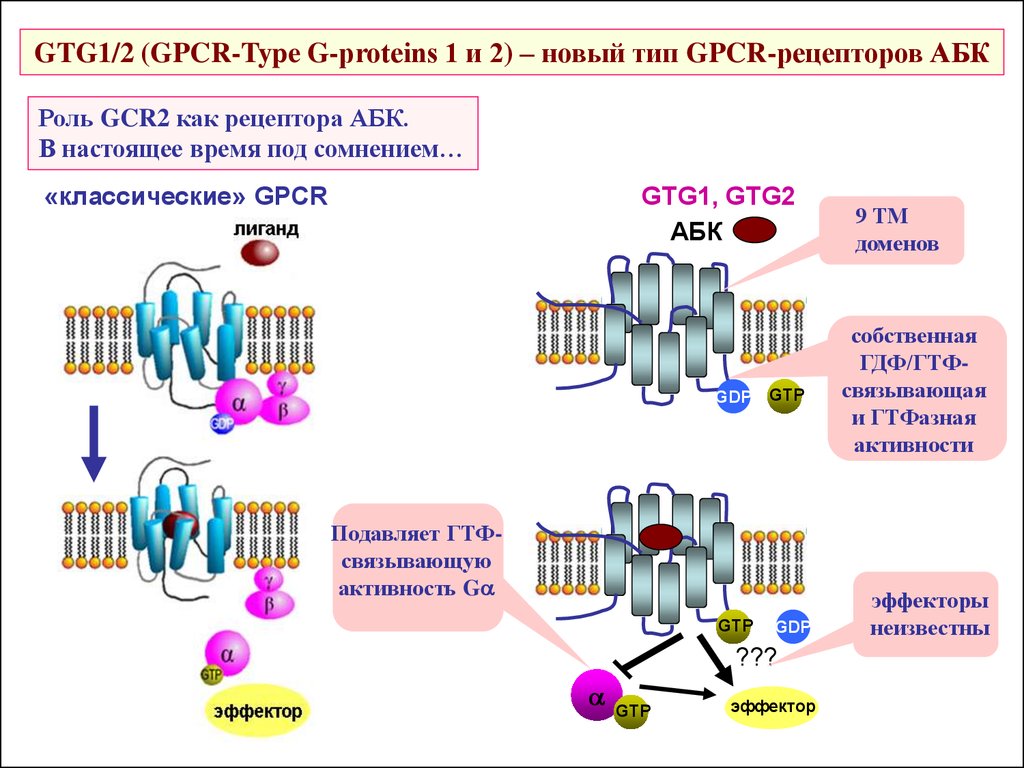
GTG1/2 (GPCR-Type G-proteins 1 и 2) – новый тип GPCR-рецепторов АБКРоль GCR2 как рецептора АБК.
В настоящее время под сомнением…
«классические» GPCR
GTG1, GTG2
АБК
GDP
GTP
Подавляет ГТФсвязывающую
активность Ga
GTP
GDP
???
a
GTP
эффектор
9 ТМ
доменов
собственная
ГДФ/ГТФсвязывающая
и ГТФазная
активности
эффекторы
неизвестны
48.
RCAR/PYR/PYL – цитозольные рецепторы АБКRegulatory Components of ABA Receptor/ Pyrobactin Resistant/ Pyrobactin Resistant-Like
Рецептор – белок
семейства
START
протеинфос
фатазы 2С
Образуют
сигналосомный
комплекс
ABI1, 2
Мономерная
убиквитинлигаза
Ser/Thr
протеинкиназы
ABI5
ABAResponsive
Elements
Трансфакторы
семейства
bZIP
SUMO
Гены ответа на
стресс (LEA, PR,
etc.)
49. Регуляторные механизмы и трансфакторы АБК-сигналинга
1. АБК и различные транс-факторыобразуются в ответ на разнообразные
сигналы (внешние или внутренние)
2. АБК запускает многие
трансфакторы за счет активации
ряда киназ. В передаче сигнала от
АБК участвует фосфолипаза D
(PLD), которая регулируется
microRNA (miRNA),
3. Промоторы АБК-регулируемых
генов имеют различные комбинации
сайтов узнавания (MYBR, MYCR, и
т.д.), которые могут связываться с
различными семействами
трансфакторов. Гены-мишени:
сдержат ABRE или ABRC (АБКрегулируемый элемент или
комплекс). В промоторной области G-бокс, часто с последовательностью
ACGT.
5. В отсутствии АБК, АБК нечувствительные факторы
транскрипции (ABI3 - 5) могут
разрушаться через протеасомы.
50.
Обобщенная схема АБК-сигналинга на 2015 год..Рецепторы АБК: плазмалемный GPCR (GPCRs), цитозольно-ядерный PYR/PYL/RCARs (PYLs), пластидный (CHLH).
Активация транскрипции АБК-зависимых генов («длинный сигналинг»)- через PYLs, PP2Cs, SnRK2s, (в левом квадрате).
Закрытие устьиц («короткий сигналинг») – через GPCRs и PYLs
В отсутствии АБК димерный PYLs не связывается с фосфатазами PP2Cs и они ингибируют SnRK2s. В присутствии АБК
мономерные PYLs ингибируют PP2Cs, SnRK2s акитвируется (автофосфорилирование) и активирует трансфакторы ( bZIP),
SLAC1 - SLOW ANION CHANNEL-ASSOCIATED 1 KAT1 - POTASSIUM CHANNEL IN ARABIDOPSIS THALIANA 1
51. Сигнальные системы АБК («короткий сигналинг»), связанные с закрытием устьиц
1. АБК связывается с ее рецепторами.2. Связывание АБК вызывает
формирование активных форм кислорода
(ROS), которые активизируют Ca2+
каналы на плазмалемме. Фосфатидная
кислота (PA), образующаяся за счет
работы фосфолипазы D (PLD)
увеличивает образование ROS.
3. Открываются Ca2+ каналы
плазмалеммы.
4. АБК стимулирует продукцию NO,
который повышает уровень циклической
АДФ-рибозы (cADPR).
5. АБК увеличивает уровень IP3 через
систему трансдукции сигналов,
включающую сфингозин-1-фосфат (S1P),
G-белки и фосфолипазы С и D (PLC и
PLD).
6. Повышение cADPR и IP3 активизирует
дополнительные каналы кальция на
тонопласте.
7. Повышение внутриклеточного кальция
блокирует K+-in каналы на плазмалемме.
8. Повышение внутриклеточного кальция
вызывает открытие хлоридных
(анионных) каналов на плазмалемме,
вызывая ее деполяризацию
9. Ингибируется Н-АТФ-за плазмалеммы
10. Деполяризация мембраны активирует
К+out каналы плазмалеммы
ROS
NO – IP3
52. Рецепторы трех гормонов работают по одному принципу..
Рецепторы, работающие на основе убиквитинлигазы(A) При низких концентрациях ауксина, гены
ответа на ауксин ингибируются Aux/IAA белками,
которые образуют гетеродимеры с ARF факторами транскрипции. В присутствии ауксина
TIR1 субединица F-бокс белка SCFTIR1 убиквитинлигазы связывается с ауксином, и в таком виде
способна присоединять Aux/IAA белки к SCF
комплексу для убиквитинирования. Деградация
Aux/IAA белков в 26S протеасоме дерепрессирует
ARF факторы транскрипции.
(B) JAZ белки негативно регулируют ответ на
воздействие жасмоната, подавляя MYC2
транскрипционные факторы. После связывания с
жасмонатом, SCFCOI1 убиквитин-лигаза
убиквитинирует JAZ белки и отправляет их на
протеолиз, дерепрессируя тем самым MYC2.
(C) DELLA белки подавляют GA ответ, негативно
регулируя PIF3, PIF4, и возможно другие факторы
транскрипции, которые контроллируют
экспрессию GA-индуцибельных генов. DELLA
белки также являются активаторами нескольких
GA-реперссируемых генов, некоторые из которых
кодируют ферменты синтез гиббереллинов и
компоненты сигнальных путей ответа на GA,
включая GID1 рецепторы. После связывания с
GA, GID1 рецептор взаимодействует с DELLA.
GID1-GA-DELLA комплекс узнается SCFGID2
убиквитин-лигазой, которая направляет DELLAбелки на убиквитинирование и соответствующую
деградацию.