











 (модуляторный):")
 ДНК")





")

 medicine
medicine biology
biologySimilar presentations:










Механизмы действия сигнальных соединений. Ядерные рецепторы
1. Механизмы действия сигнальных соединений
2. Этапы инициации эффектов сигнальных соединений
Селективная генерация межклеточного сигнала(эндокринная клетка/железа)
Клеточное узнавание сигнала
(рецепторы)
Внутриклеточная интерпретация сигнала
(рецепторы+сигнальные каскады)
Селективный ответ клетки
Начальный
Ранний
Поздний
Биохимические, пролиферативные, физиологические, морфологические
эффекты
3. Классификация гормонкомпетентных тканей
ТканиНемишени
Мишени
(рецепторов нет)
(рецепторы есть)
Гормонзависимые
Общетрофические
эффекты+селективные
эффекты
Гормончувствительные
Селективные эффекты
4.
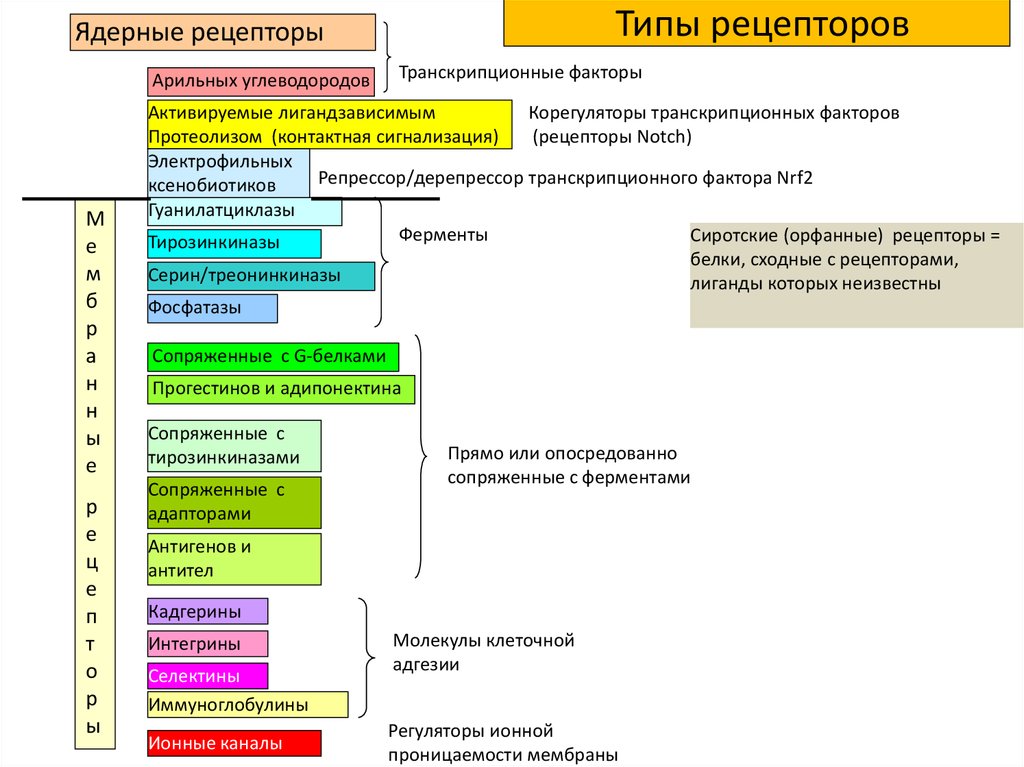
Типы рецепторовЯдерные рецепторы
Арильных углеводородов
М
е
м
б
р
а
н
н
ы
е
р
е
ц
е
п
т
о
р
ы
Транскрипционные факторы
Активируемые лигандзависимым
Корегуляторы транскрипционных факторов
Протеолизом (контактная сигнализация)
(рецепторы Notch)
протеолизом
Электрофильных
Репрессор/дерепрессор транскрипционного фактора Nrf2
ксенобиотиков
Гуанилатциклазы
Ферменты
Сиротские (орфанные) рецепторы =
Тирозинкиназы
белки, сходные с рецепторами,
Серин/треонинкиназы
лиганды которых неизвестны
Фосфатазы
Сопряженные с G-белками
Прогестинов и адипонектина
Сопряженные с
тирозинкиназами
Сопряженные с
адапторами
Прямо или опосредованно
сопряженные с ферментами
Антигенов и
антител
Кадгерины
Интегрины
Селектины
Иммуноглобулины
Ионные каналы
Молекулы клеточной
адгезии
Регуляторы ионной
проницаемости мембраны
5. Модель структурно-функциональной организации рецептора
Гормонсвязывающий доменСопрягающий
домен
Эффекторный
домен
6. Уни- и мультирецепторные модели действия рецепторов
Рецептор 1Рецептор 1
Рецептор 2
Рецептор 3
Эффекты 1
Эффекты 2
Эффекты (n)
Эффекты 1
Эффекты 2
Эффекты 3
7.
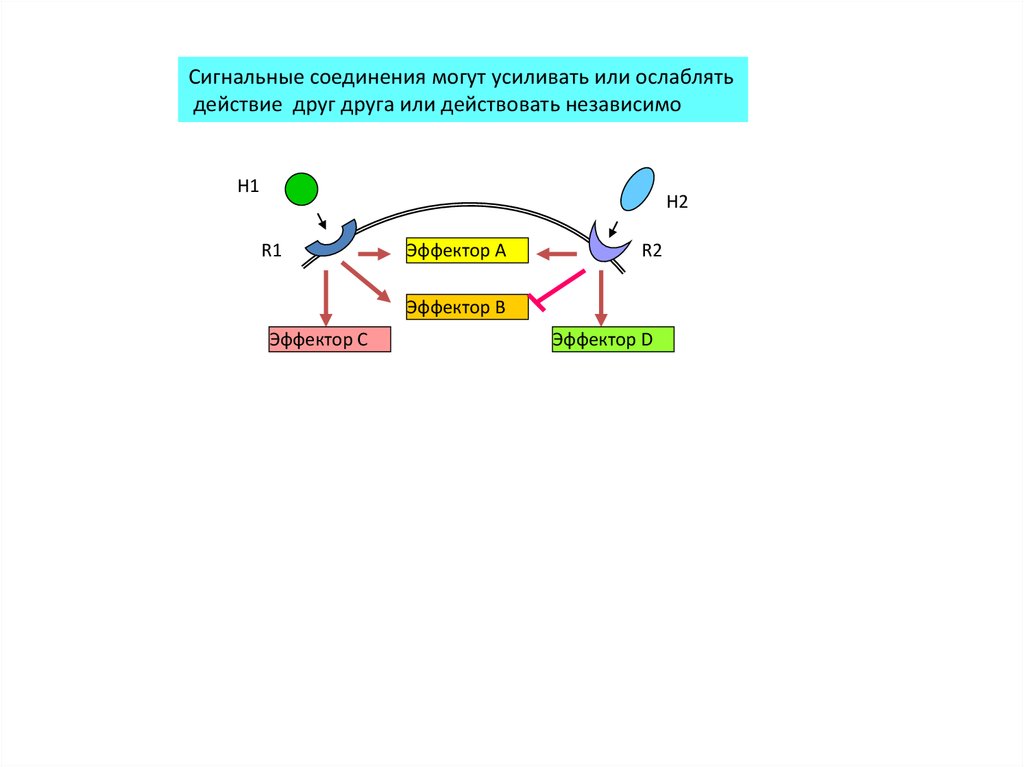
Сигнальные соединения могут усиливать или ослаблятьдействие друг друга или действовать независимо
H1
H2
R1
Эффектор A
R2
Эффектор B
Эффектор C
Эффектор D
8.
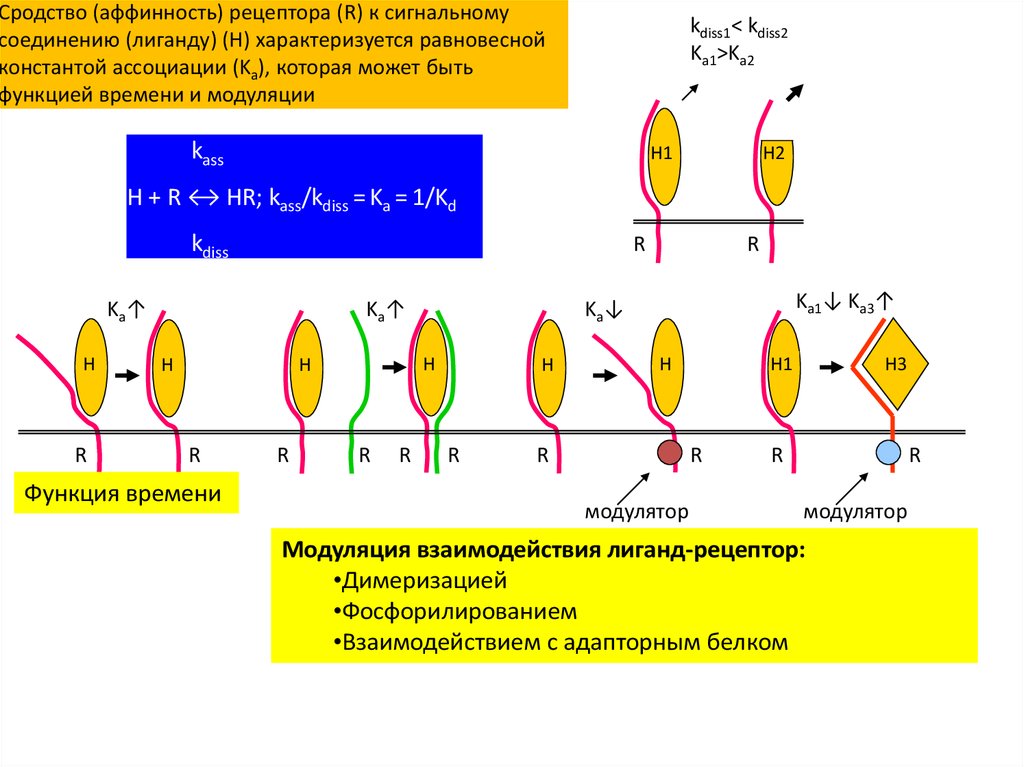
Сродство (аффинность) рецептора (R) к сигнальномусоединению (лиганду) (H) характеризуется равновесной
константой ассоциации (Ka), которая может быть
функцией времени и модуляции
kdiss1< kdiss2
Ka1>Ka2
kass
H1
H2
H + R ↔ HR; kass/kdiss = Ka = 1/Kd
kdiss
R
Ka↑
H
R
Ka↑
H
R
Функция времени
R
R
R
Ka1↓ Ka3↑
Ka↓
H
H
R
H
R
H
R
H1
R
модулятор
H3
R
R
модулятор
Модуляция взаимодействия лиганд-рецептор:
•Димеризацией
•Фосфорилированием
•Взаимодействием с адапторным белком
9.
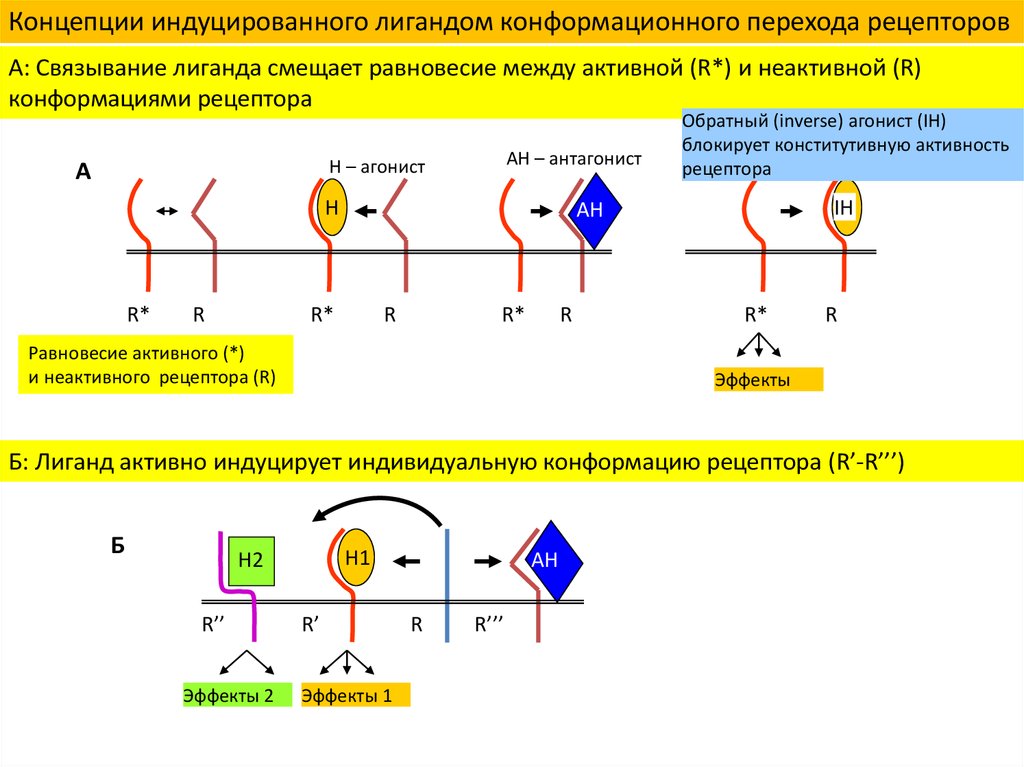
Концепции индуцированного лигандом конформационного перехода рецепторовА: Связывание лиганда смещает равновесие между активной (R*) и неактивной (R)
конформациями рецептора
AH – антагонист
H – агонист
А
H
R*
R
Обратный (inverse) агонист (IH)
блокирует конститутивную активность
рецептора
IH
AH
R*
R
R*
R
Равновесие активного (*)
и неактивного рецептора (R)
R*
R
Эффекты
Б: Лиганд активно индуцирует индивидуальную конформацию рецептора (R’-R’’’)
Б
H1
H2
R’’
Эффекты 2
R’
Эффекты 1
AH
R
R’’’
10.
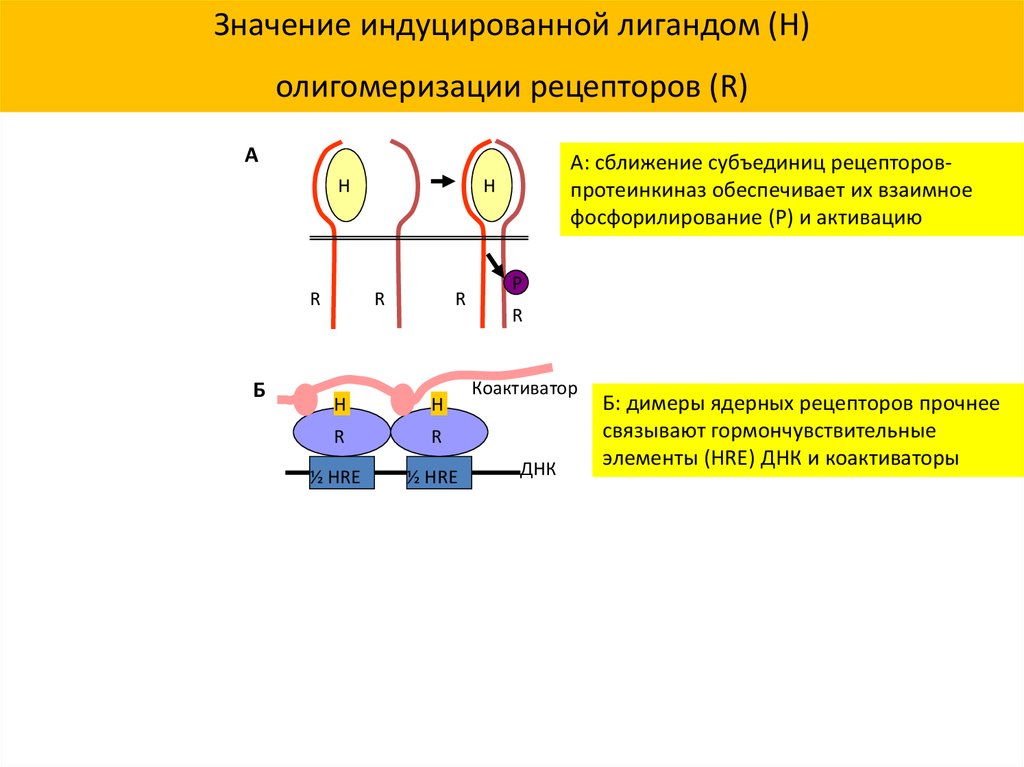
Значение индуцированной лигандом (H)олигомеризации рецепторов (R)
А
H
R
Б
А: сближение субъединиц рецепторовпротеинкиназ обеспечивает их взаимное
фосфорилирование (P) и активацию
H
R
R
H
H
R
R
½ HRE
½ HRE
P
R
Коактиватор
ДНК
Б: димеры ядерных рецепторов прочнее
связывают гормончувствительные
элементы (HRE) ДНК и коактиваторы
11.
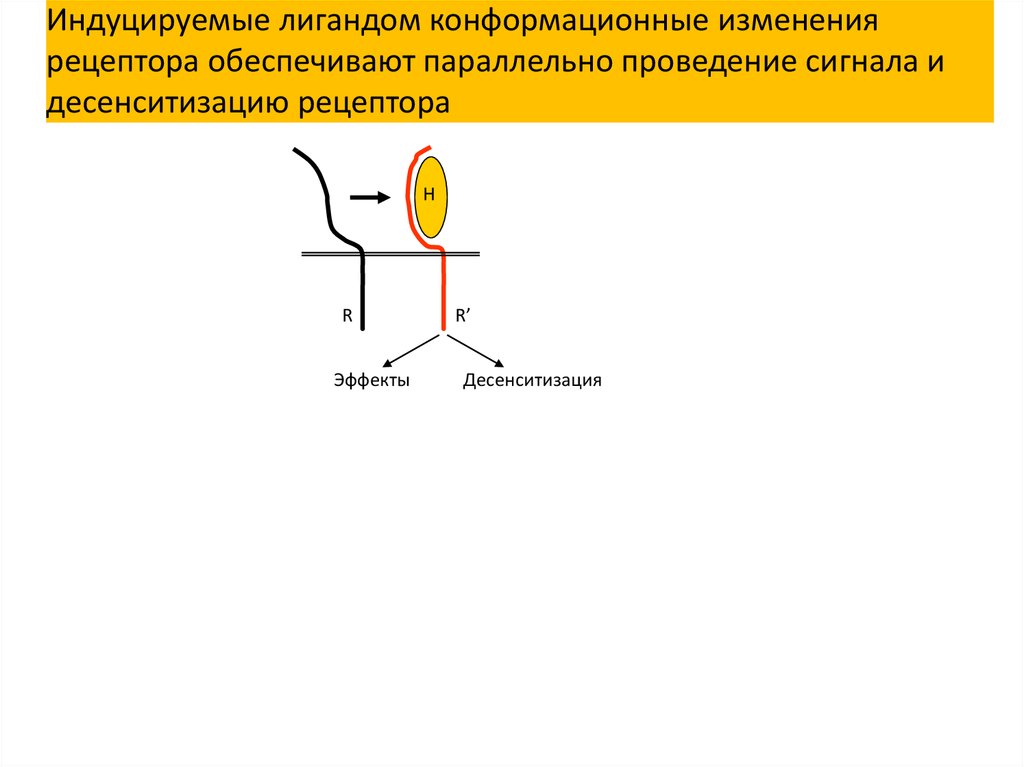
Индуцируемые лигандом конформационные изменениярецептора обеспечивают параллельно проведение сигнала и
десенситизацию рецептора
H
R
Эффекты
R’
Десенситизация
12. Ядерные рецепторы и системы передачи сигнала
13. Специфические факторы транскрипции
• Конститутивные факторы транскрипции =факторы транскрипции базального транскрипционого комплекса:
• Обеспечение инициации транскрипции
• Обеспечение базального уровня транскрипции
• Обеспечение тканеспецифичности транскрипции
(тканеспецифические конститутивные факторы транскрипции)
• Адаптивные факторы транскрипции:
Регуляция скорости транскрипции гена в зависимости от условий внешней и
внутренней среды
в результате взаимодействия с:
• коактиваторами (семейство SRC-1, SRA, HATs, TIF1, p68,
CBP/p200 и т.д.)
• конститутивными факторами транскрипции (TAFs, TBP, TFIIB, SAGA,
HATs и т.д.)
• Регуляция транскрипции в результате взаимодействия с ДНК очень
редко
14.
Адаптивные факторы транскрипцииСпособ передачи:
на входе:
Взаимодействие с гормоном
Взаимодействие с другим сигнальным соединением
Фосфорилирование
На выходе:
Изменение скорости транскрипции гена
Перестройка (деконденсация или гиперконденсация) хроматина – изменение
доступности для РНК-полимераз
Изменение конформации ДНК, оптимизирующее ее для действия других
транскрипционных факторов
15.
Передача сигнала набазальный транскрипционный комплекс
Корегуляторы ядерных рецепторов:
1. Корепрессоры
• Помогают подавлять действие рецептора без лиганда
• Связывание лиганда ведет к диссоциации комплекса рецептор-корепрессор и
дает возможность рецептору связаться с коактиватором
2. Коактиваторы
• Связывание ГРК с основными транскрипционными факторами
• Связывание ГРК с РНК-полимеразой 2
16. Регуляция экспрессии гормонзависимого гена млекопитающих
Сборка базального транскрипционного комплексаэ
н
х
а
н
с
е
р
с
а
й
л
е
н
с
е
р
Корегулятор
Гормон
5’
ЯР
ГЧЭ
ЯР
Тканеспецифические
факторы транскрипции
Активация
транскрипции
Базальный
транскрипционный
комплекс
Кодирующая
область
3’
мРНК
3’-некодирующая
область
Регуляторная область
Промоторная
область
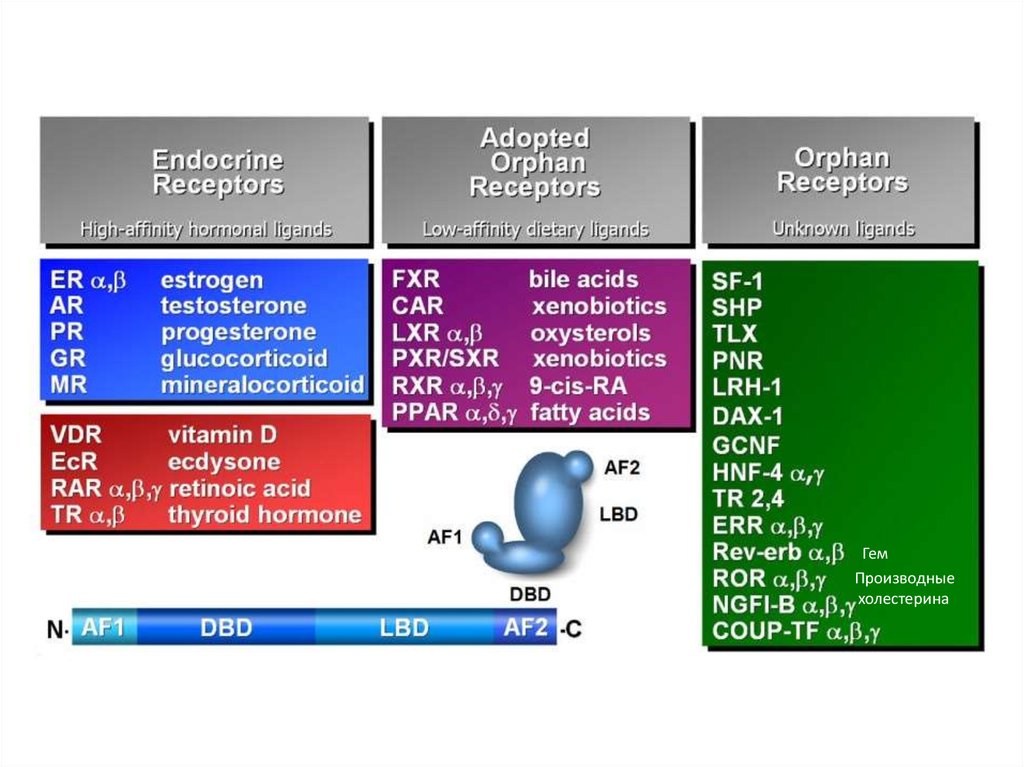
17. Лиганды ядерных рецепторов
––
Гормоны (стероидные, тиреоидоные)
Гормональные формы жирорастворимых
витаминов (Д и А)
Сиротские:
–
Сенсоры липидов
Сенсоры ксенобиотиков
Оставшиеся сиротскими
18.
ГемПроизводные
холестерина
19.
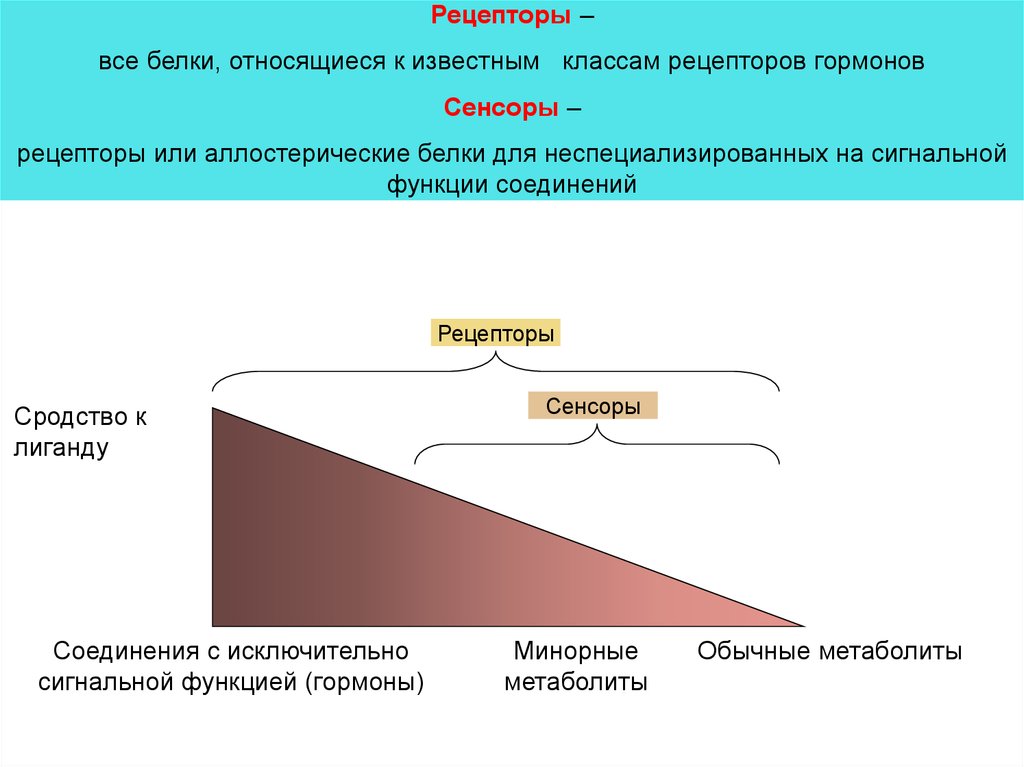
Рецепторы –все белки, относящиеся к известным классам рецепторов гормонов
Сенсоры –
рецепторы или аллостерические белки для неспециализированных на сигнальной
функции соединений
Рецепторы
Сродство к
лиганду
Соединения с исключительно
сигнальной функцией (гормоны)
Сенсоры
Минорные
метаболиты
Обычные метаболиты
20.
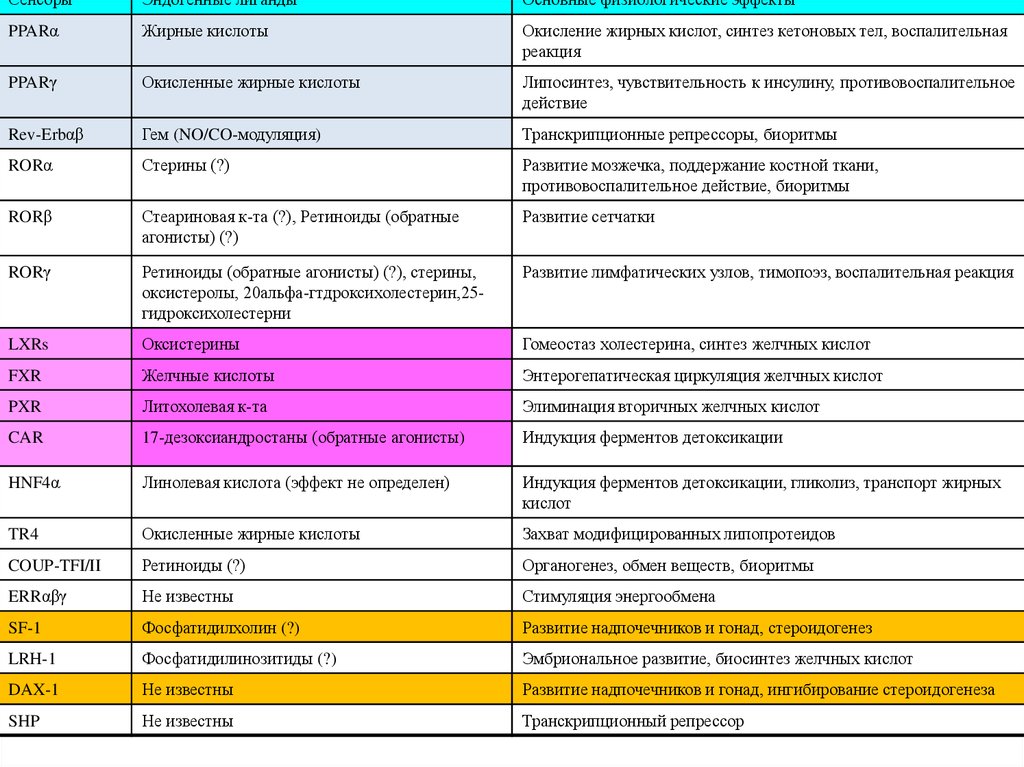
СенсорыЭндогенные лиганды
Основные физиологические эффекты
PPARα
Жирные кислоты
Окисление жирных кислот, синтез кетоновых тел, воспалительная
реакция
PPARγ
Окисленные жирные кислоты
Липосинтез, чувствительность к инсулину, противовоспалительное
действие
Rev-Erbαβ
Гем (NO/CO-модуляция)
Транскрипционные репрессоры, биоритмы
RORα
Стерины (?)
Развитие мозжечка, поддержание костной ткани,
противовоспалительное действие, биоритмы
RORβ
Стеариновая к-та (?), Ретиноиды (обратные
агонисты) (?)
Развитие сетчатки
RORγ
Ретиноиды (обратные агонисты) (?), стерины,
оксистеролы, 20альфа-гтдроксихолестерин,25гидроксихолестерни
Развитие лимфатических узлов, тимопоэз, воспалительная реакция
LXRs
Оксистерины
Гомеостаз холестерина, синтез желчных кислот
FXR
Желчные кислоты
Энтерогепатическая циркуляция желчных кислот
PXR
Литохолевая к-та
Элиминация вторичных желчных кислот
CAR
17-дезоксиандростаны (обратные агонисты)
Индукция ферментов детоксикации
HNF4α
Линолевая кислота (эффект не определен)
Индукция ферментов детоксикации, гликолиз, транспорт жирных
кислот
TR4
Окисленные жирные кислоты
Захват модифицированных липопротеидов
COUP-TFI/II
Ретиноиды (?)
Органогенез, обмен веществ, биоритмы
ERRαβγ
Не известны
Стимуляция энергообмена
SF-1
Фосфатидилхолин (?)
Развитие надпочечников и гонад, стероидогенез
LRH-1
Фосфатидилинозитиды (?)
Эмбриональное развитие, биосинтез желчных кислот
DAX-1
Не известны
Развитие надпочечников и гонад, ингибирование стероидогенеза
SHP
Не известны
Транскрипционный репрессор
21. Активность ядерных рецепторов в отсутствие лиганда
1. Транскрипционно неактивные без лиганда :некоторые рецепторы стероидов
2. Оказывающие репрессорное действие без
лиганда: рецепторы тиреоидных гормонов,
рецепторы гормональной формы витамина Д
3. Оказывающие стимулирующее действие без
лиганда : CAR (рецепторы 5альфа-андростанов,
фенобарбитураты – конкурентные антагонисты;
андростановые производные - обратные агонисты)
22.
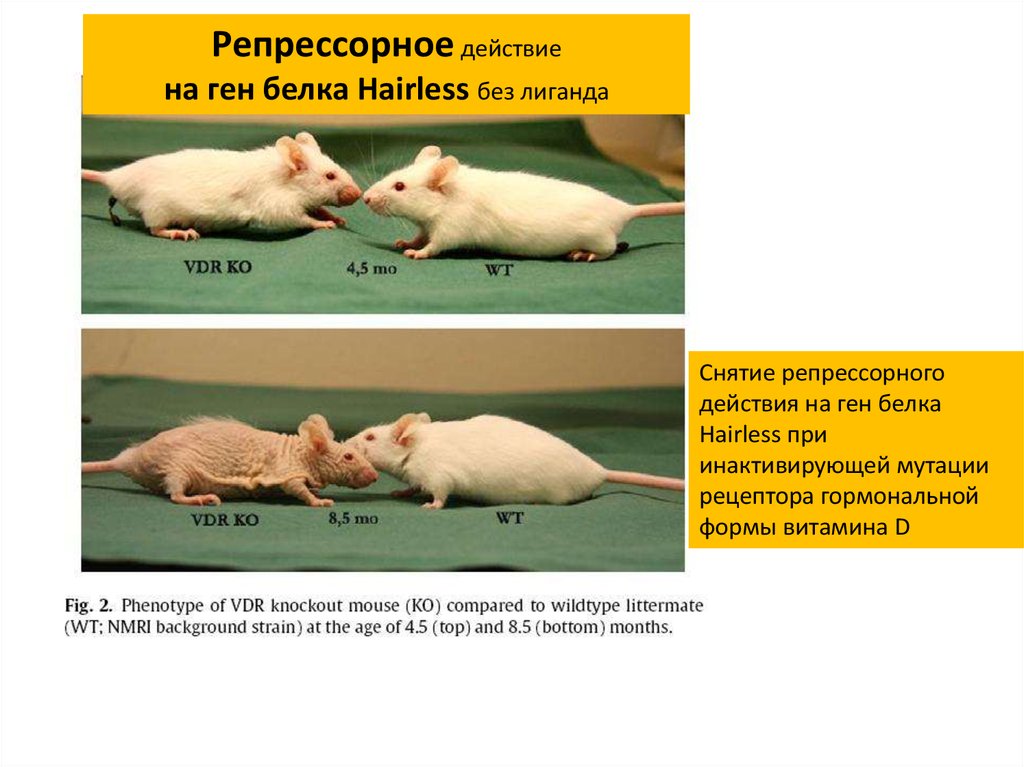
Репрессорное действиена ген белка Hairless без лиганда
Снятие репрессорного
действия на ген белка
Hairless при
инактивирующей мутации
рецептора гормональной
формы витамина D
23.
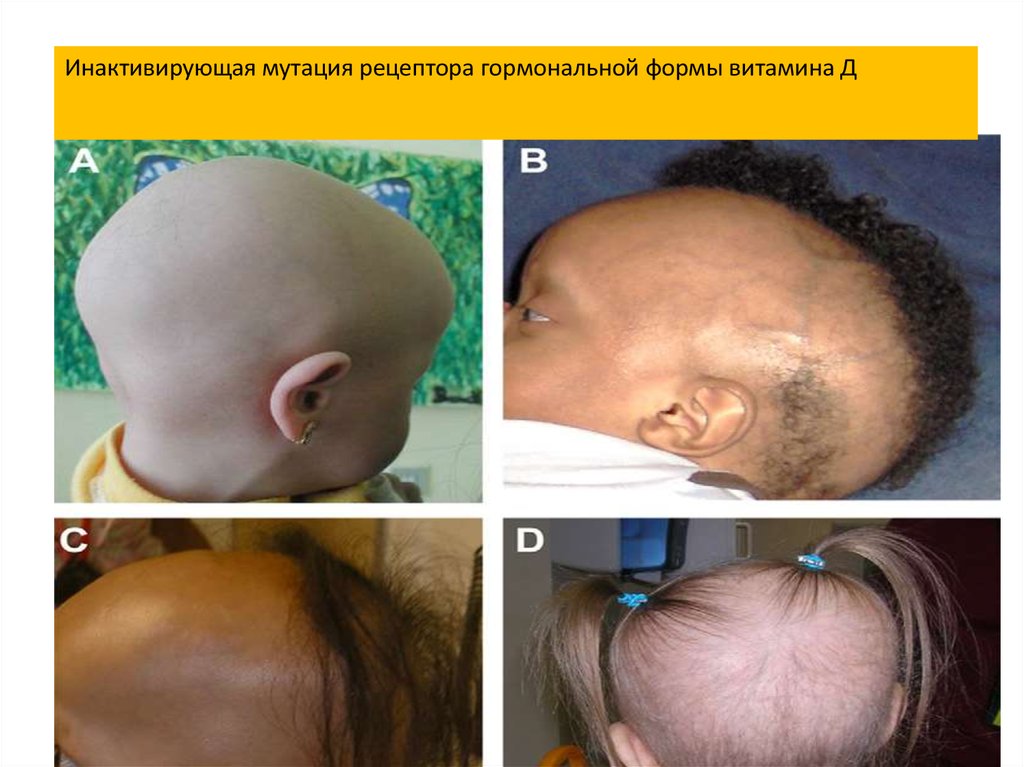
Инактивирующая мутация рецептора гормональной формы витамина Д24.
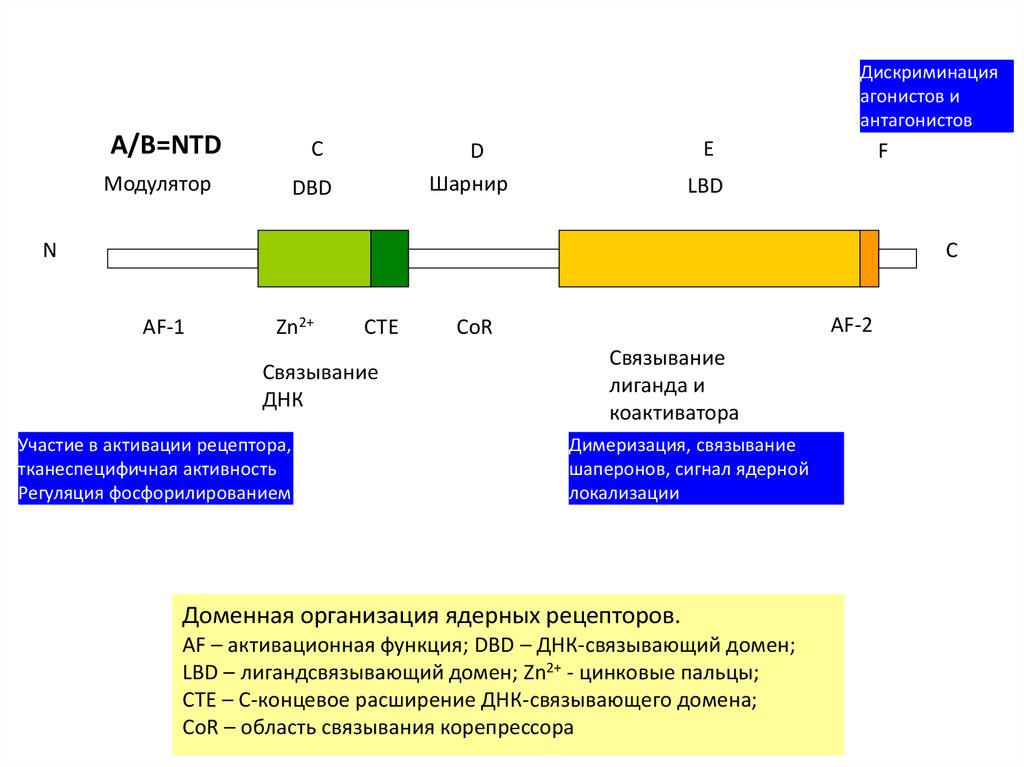
Дискриминацияагонистов и
антагонистов
A/B=NTD
C
Модулятор
E
D
Шарнир
DBD
F
LBD
N
C
AF-1
Zn2+
CTE
Связывание
ДНК
Участие в активации рецептора,
тканеспецифичная активность
Регуляция фосфорилированием
AF-2
CoR
Связывание
лиганда и
коактиватора
Димеризация, связывание
шаперонов, сигнал ядерной
локализации
Доменная организация ядерных рецепторов.
AF – активационная функция; DBD – ДНК-связывающий домен;
LBD – лигандсвязывающий домен; Zn2+ - цинковые пальцы;
CTE – C-концевое расширение ДНК-связывающего домена;
CoR – область связывания корепрессора
25. Домен NTD (A/B) (модуляторный):
––
–
–
Наиболее вариабельный по длине и послед-ти а/к
Сайты фосфорилирования MAP-киназами и другими
протеинкиназами
ГормонНЕзависимая активаторная функция AF-1
(взаимодействие с тканеспецифеческими
корегуляторами (коактиваторами)
Участвует в гормонзависимой активации
рецепторов стероидов
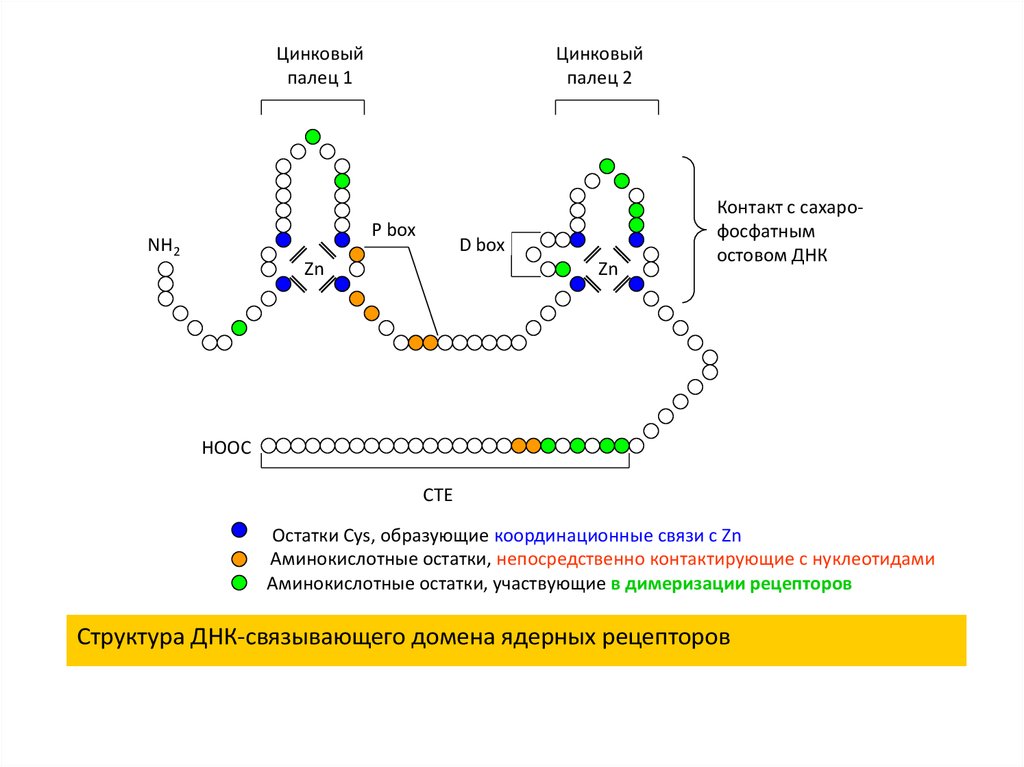
26.
Цинковыйпалец 1
Цинковый
палец 2
P box
NH2
D box
Zn
Zn
Контакт с сахарофосфатным
остовом ДНК
HOOC
CTE
Остатки Cys, образующие координационные связи с Zn
Аминокислотные остатки, непосредственно контактирующие с нуклеотидами
Аминокислотные остатки, участвующие в димеризации рецепторов
Структура ДНК-связывающего домена ядерных рецепторов
27. Гормончувствительные элементы (ГЧЭ) ДНК
Особенности взаимодействия ГЧЭ с ядерными рецепторами (ЯР):Полусайты –связывание мономера ЯР
Сдвоенные полусайты – связывание гомо- и гетеродимеров ЯР
Варианты сдвоенных полусайтов:
•Палиндромы
•Прямые повторы
•Обратные повторы
•Содержат спейсеры
Активность ГЧЭ:
•Позитивные – усиление экспрессии
•Негативные – подавление экспрессии
•Кол-во ГЧЭ – усиление эффекта
28.
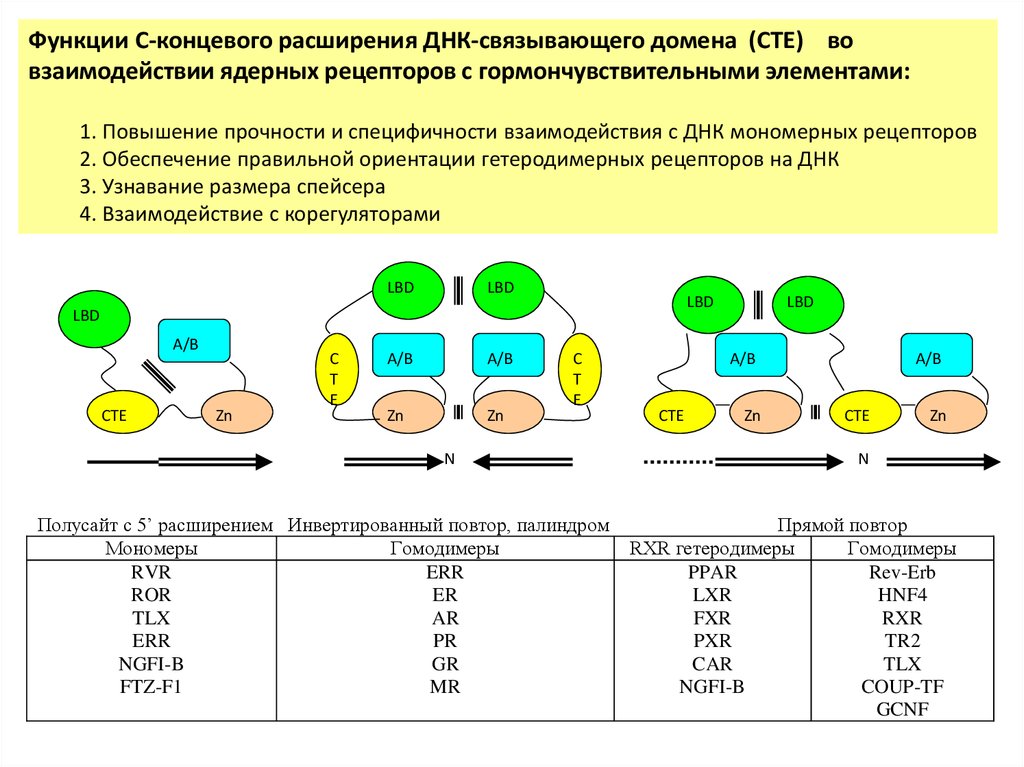
Функции C-концевого расширения ДНК-связывающего домена (СТЕ) вовзаимодействии ядерных рецепторов с гормончувствительными элементами:
1. Повышение прочности и специфичности взаимодействия с ДНК мономерных рецепторов
2. Обеспечение правильной ориентации гетеродимерных рецепторов на ДНК
3. Узнавание размера спейсера
4. Взаимодействие с корегуляторами
LBD
LBD
A/B
A/B
Zn
Zn
LBD
LBD
A/B
CTE
Zn
C
T
E
C
T
E
N
Полусайт с 5’ расширением Инвертированный повтор, палиндром
Мономеры
Гомодимеры
RVR
ERR
ROR
ER
TLX
AR
ERR
PR
NGFI-B
GR
FTZ-F1
MR
LBD
A/B
CTE
Zn
A/B
CTE
Zn
N
Прямой повтор
RXR гетеродимеры
Гомодимеры
PPAR
Rev-Erb
LXR
HNF4
FXR
RXR
PXR
TR2
CAR
TLX
NGFI-B
COUP-TF
GCNF
29.
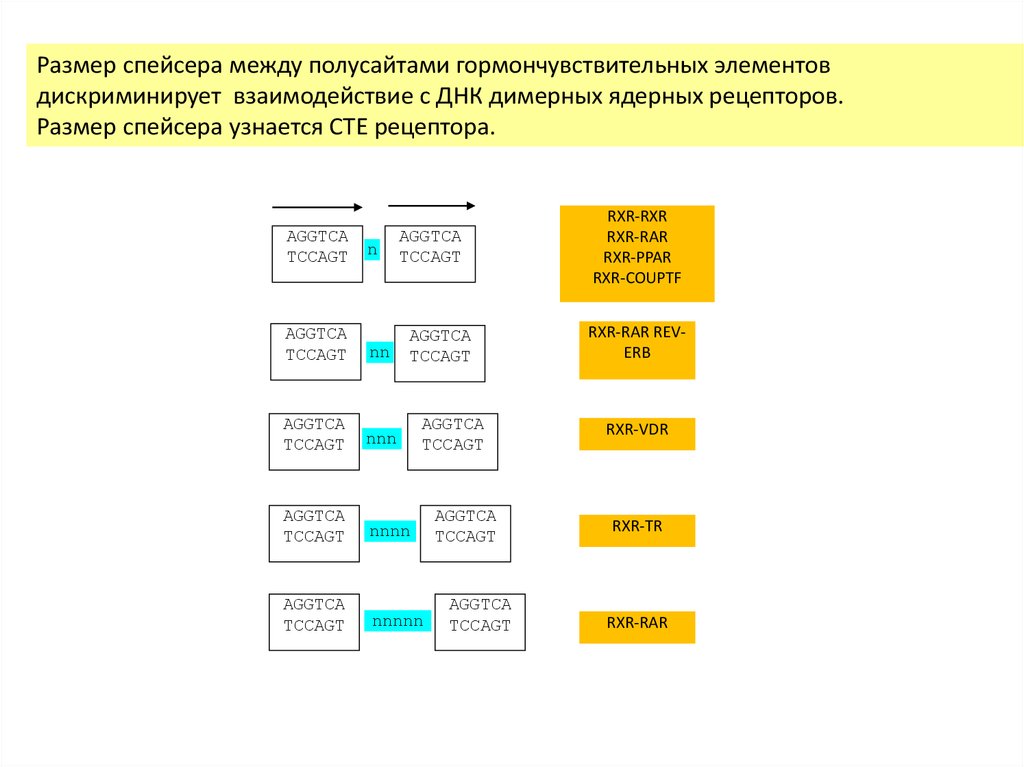
Размер спейсера между полусайтами гормончувствительных элементовдискриминирует взаимодействие с ДНК димерных ядерных рецепторов.
Размер спейсера узнается CTE рецептора.
AGGTCA
TCCAGT
AGGTCA
TCCAGT
n
AGGTCA
TCCAGT
nn
AGGTCA
TCCAGT
nnn
AGGTCA
TCCAGT
nnnn
AGGTCA
TCCAGT
nnnnn
AGGTCA
TCCAGT
AGGTCA
TCCAGT
AGGTCA
TCCAGT
AGGTCA
TCCAGT
RXR-RXR
RXR-RAR
RXR-PPAR
RXR-COUPTF
RXR-RAR REVERB
RXR-VDR
RXR-TR
RXR-RAR
30.
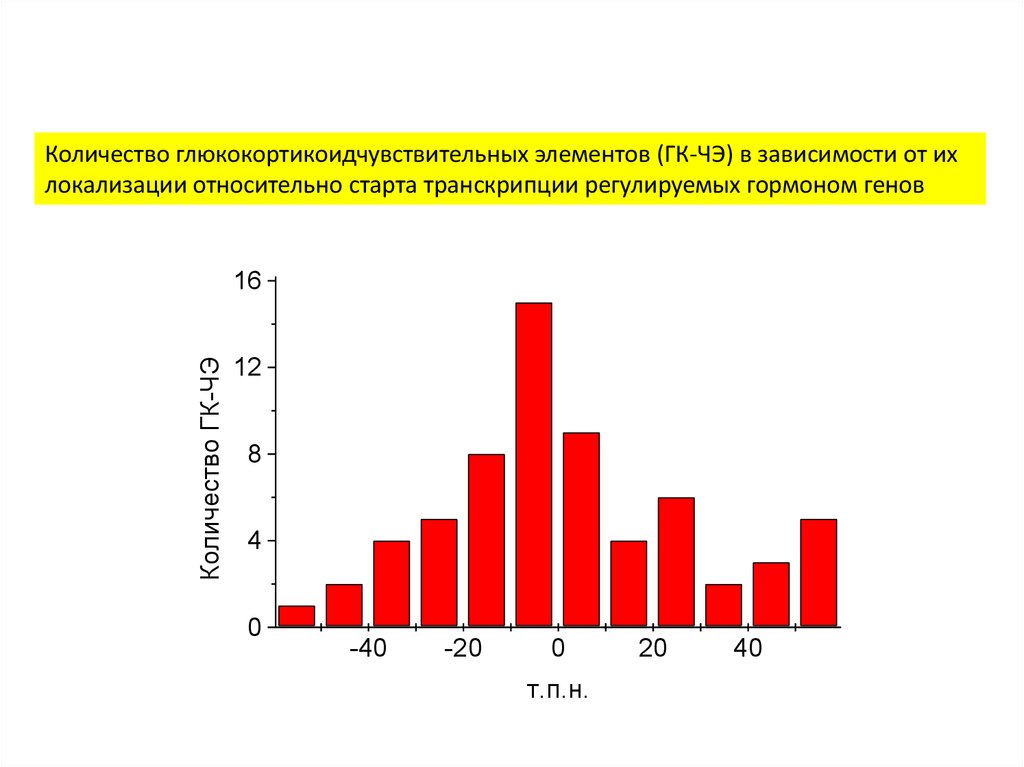
Количество глюкокортикоидчувствительных элементов (ГК-ЧЭ) в зависимости от ихлокализации относительно старта транскрипции регулируемых гормоном генов
Количество ГК-ЧЭ
16
12
8
4
0
-40
-20
0
т.п.н.
20
40
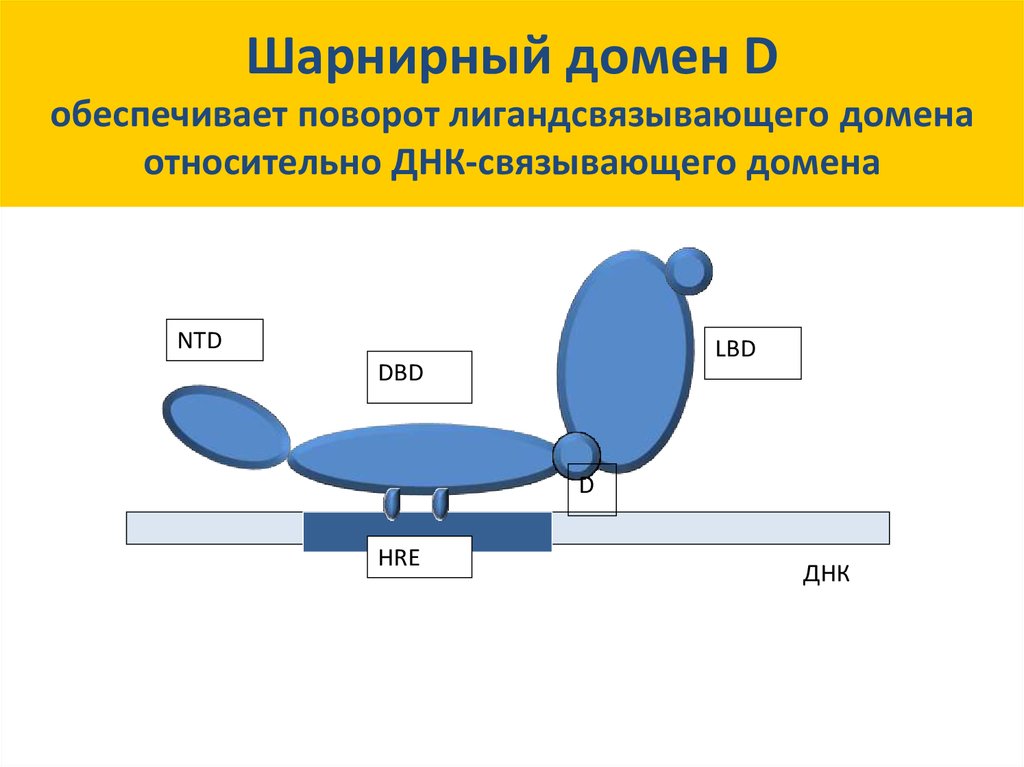
31. Шарнирный домен D
– Вариабелен по размеру ипоследовательности а/к
• Гибкий (поворот на 180° по отношению к
домену Е и возможность совмещения
связывания с ДНК и димеризации)
• Поверхность взаимодействия ЯР с
корепрессорами
32.
Шарнирный домен Dобеспечивает поворот лигандсвязывающего домена
относительно ДНК-связывающего домена
NTD
LBD
DBD
D
HRE
ДНК
33.
Лигандсвязывающий домен Е:Умеренно консервативен
Лигандсвязывающий карман:
• Связывание лиганда
• Участие в димеризации
• Взаимодействие с белками теплового шока
• Сигнал ядерной транслокации
Лигандзависимая активаторная функция AF2:
• Поверхность взаимодействия с коактиваторами
• Работа по типу мышеловки
• Сайт димеризации
Домен F:
Дополнительная дискриминация агонистов и антагонистов,
Связь с корепрессорами
34. Работа гормонзависимой активаторной функции AF2
Появление сайтавзаимодействия с
коактиватором
Гормон
AF2
AF2
AF2
AF2
AF2
AF2
Лигандсвязывающий карман
Конкурентный
антагонист
35.
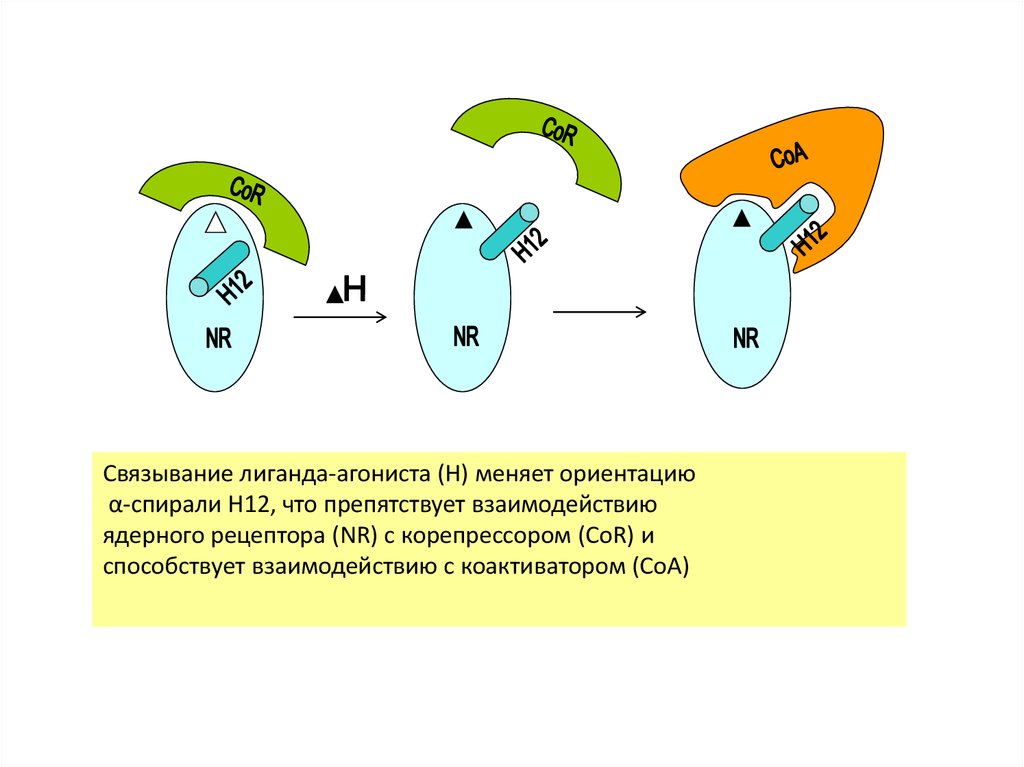
Связывание лиганда-агониста (H) меняет ориентациюα-спирали H12, что препятствует взаимодействию
ядерного рецептора (NR) с корепрессором (CoR) и
способствует взаимодействию с коактиватором (CoA)
36.
Роль димеризации37.
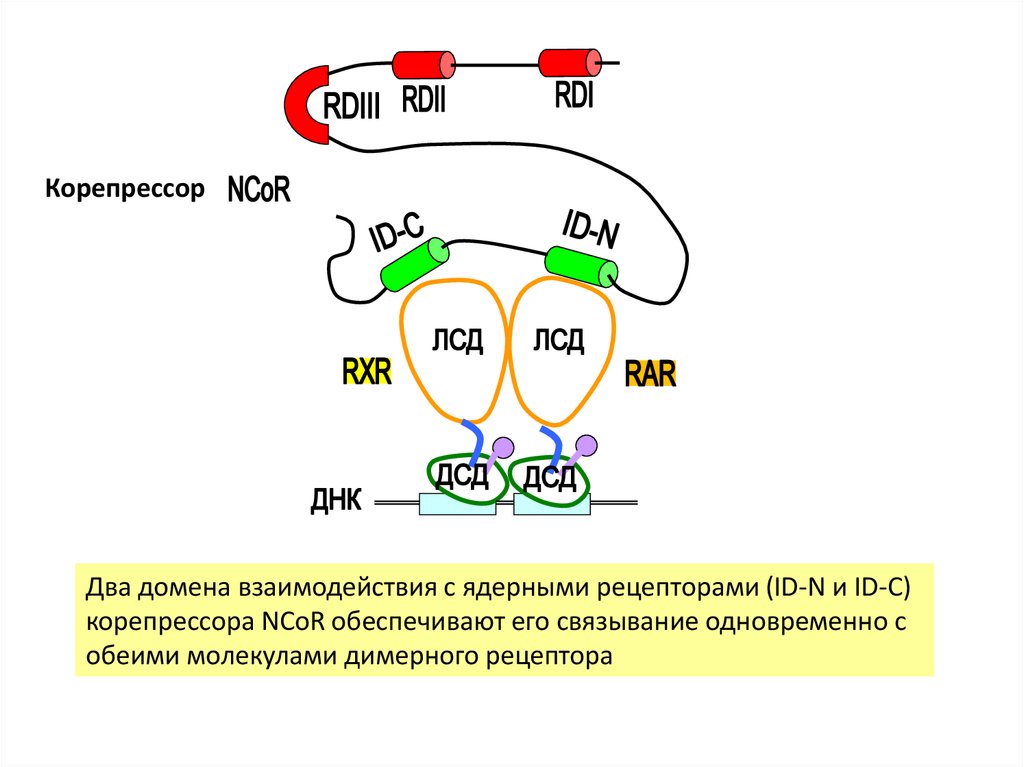
КорепрессорДва домена взаимодействия с ядерными рецепторами (ID-N и ID-C)
корепрессора NCoR обеспечивают его связывание одновременно с
обеими молекулами димерного рецептора
38.
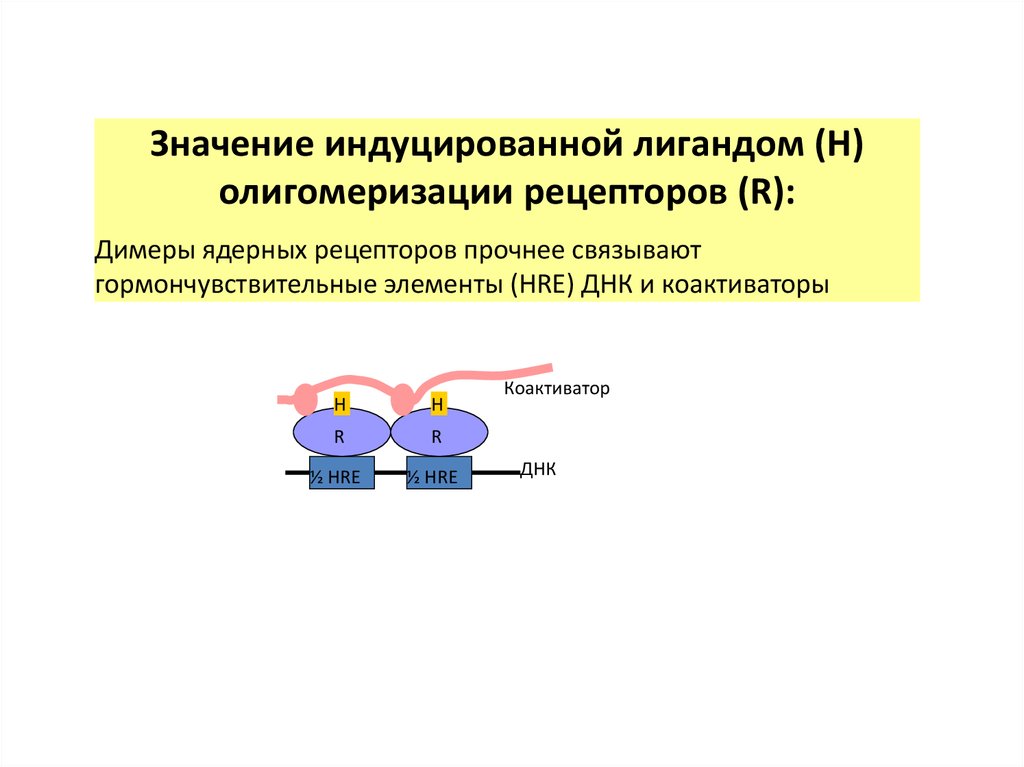
Значение индуцированной лигандом (H)олигомеризации рецепторов (R):
Димеры ядерных рецепторов прочнее связывают
гормончувствительные элементы (HRE) ДНК и коактиваторы
H
H
R
R
½ HRE
½ HRE
Коактиватор
ДНК
39.
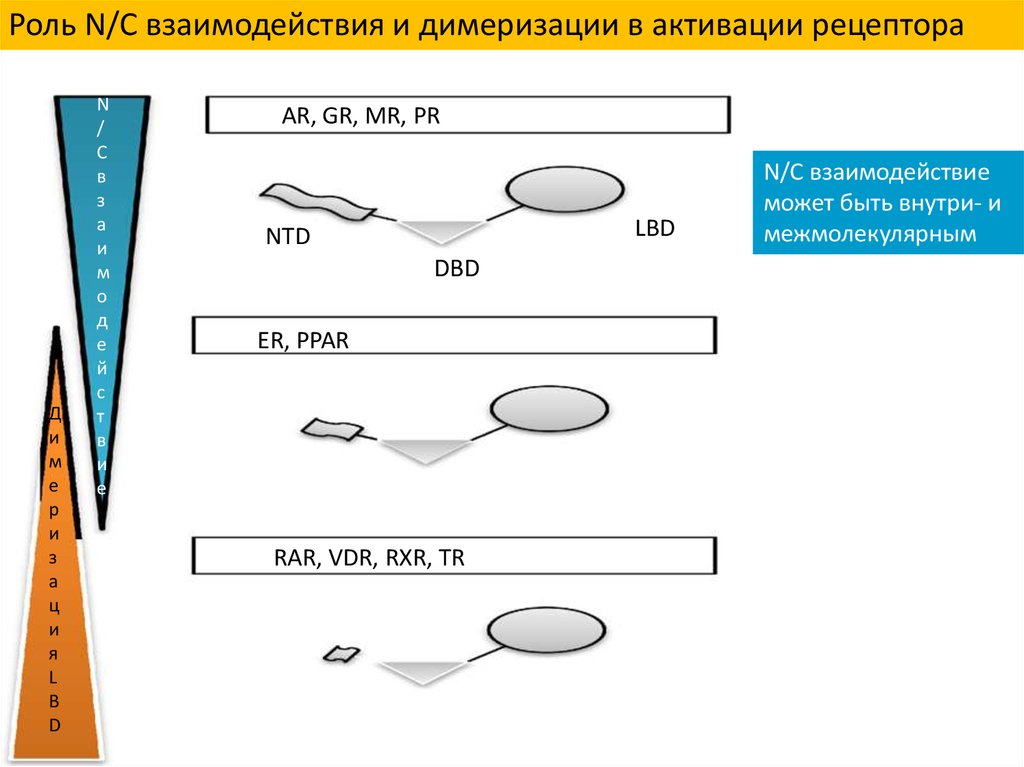
Роль N/C взаимодействия и димеризации в активации рецептораД
и
м
е
р
и
з
а
ц
и
я
L
B
D
N
/
C
в
з
а
и
м
о
д
е
й
с
т
в
и
е
AR, GR, MR, PR
LBD
NTD
DBD
ER, PPAR
RAR, VDR, RXR, TR
N/C взаимодействие
может быть внутри- и
межмолекулярным
40. Партнеры по димеризации
В зависимости от партнера по димеризации
чаще узнаются разные типы ГЧЭ
Способствуют удержанию ЯР в ядре
–
Одинаковый для разных ЯР партнер RXR:
Помогает поиску сайта с максимальным сродством, т.к.
количество ГЧЭ для RXR больше, чем для других ЯР
Пермиссивный эффект: 9-цисRA – RXR – пермиссирует
взаимодействие лиганда с рецептором
Непермиссивный эффект:
кооперативное усиление эффекта
41.
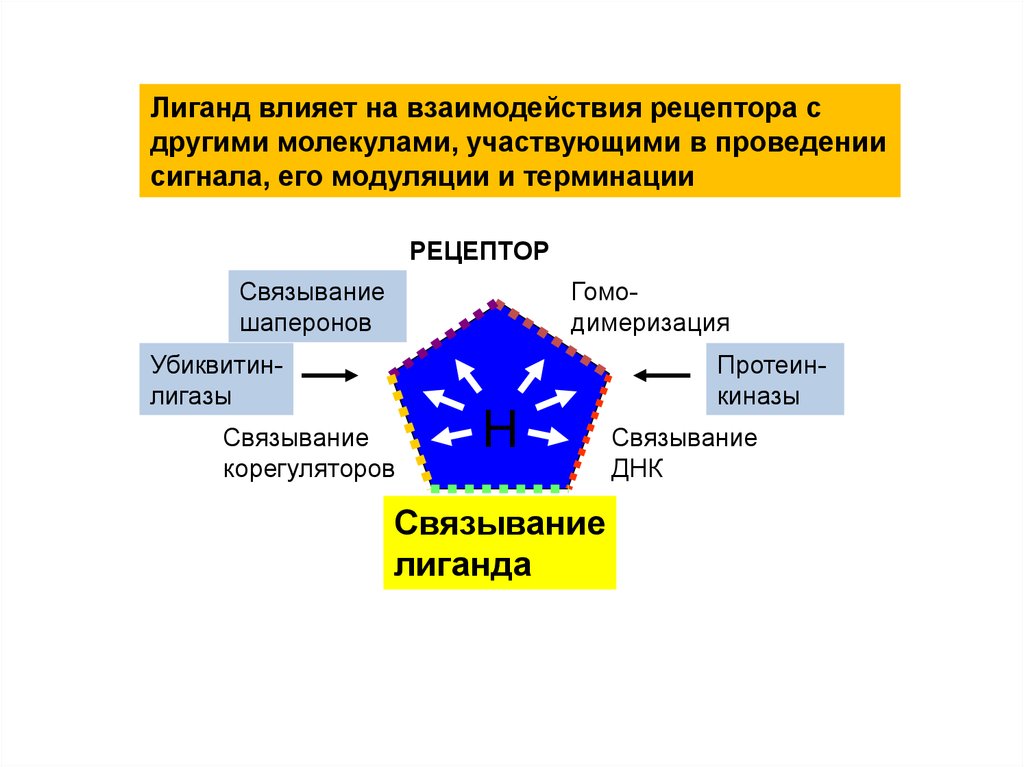
Лиганд влияет на взаимодействия рецептора сдругими молекулами, участвующими в проведении
сигнала, его модуляции и терминации
РЕЦЕПТОР
Связывание
шаперонов
Гомодимеризация
Убиквитинлигазы
Протеинкиназы
Связывание
корегуляторов
H
Связывание
лиганда
Связывание
ДНК
42. Роль комплексирования рецепторов стероидов с белками теплового шока (hsp90)
В цитоплазме гетеродимеры:1 молекула ЯР+2 молекулы hsp90
Роль hsp90:
• Комплексирование с гормонсвязывающим доменом
• Поддержание и усиление аффинности
гормонсвязывающего центра к гормону
• Блокирование ДНК-связывающего домена в отсутствие
гормона
• Транспорт вновь синтезированных рецепторов к ядру
43.
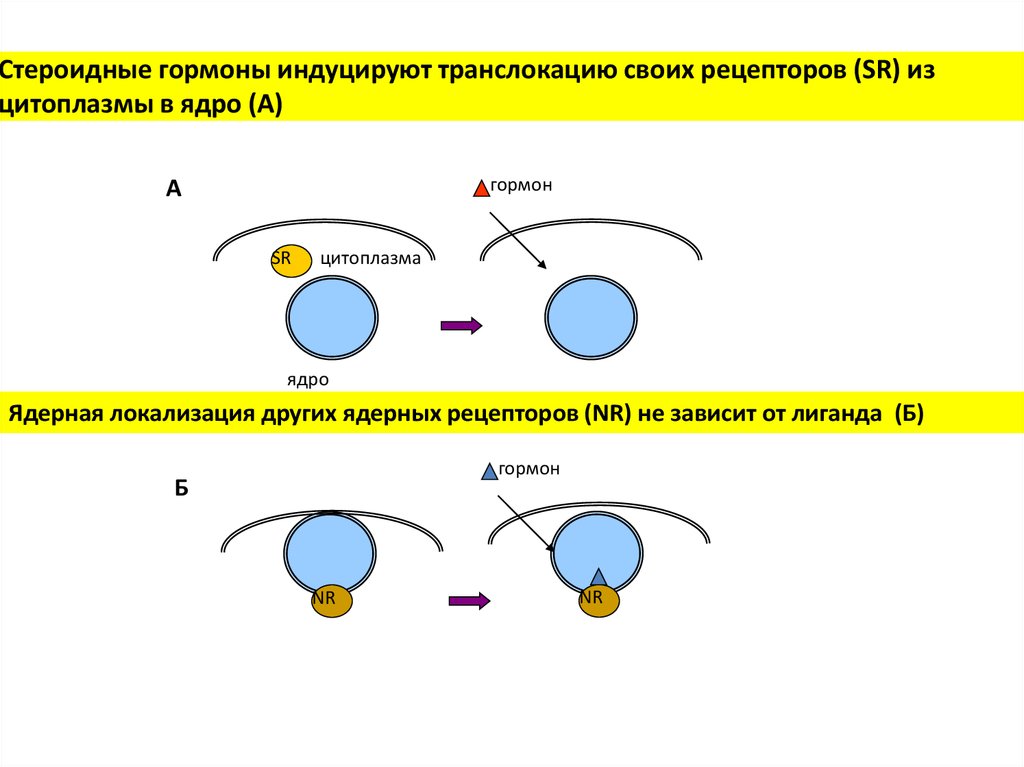
Стероидные гормоны индуцируют транслокацию своих рецепторов (SR) изцитоплазмы в ядро (А)
гормон
А
SR
цитоплазма
SR
ядро
Ядерная локализация других ядерных рецепторов (NR) не зависит от лиганда (Б)
гормон
Б
NR
NR
44.
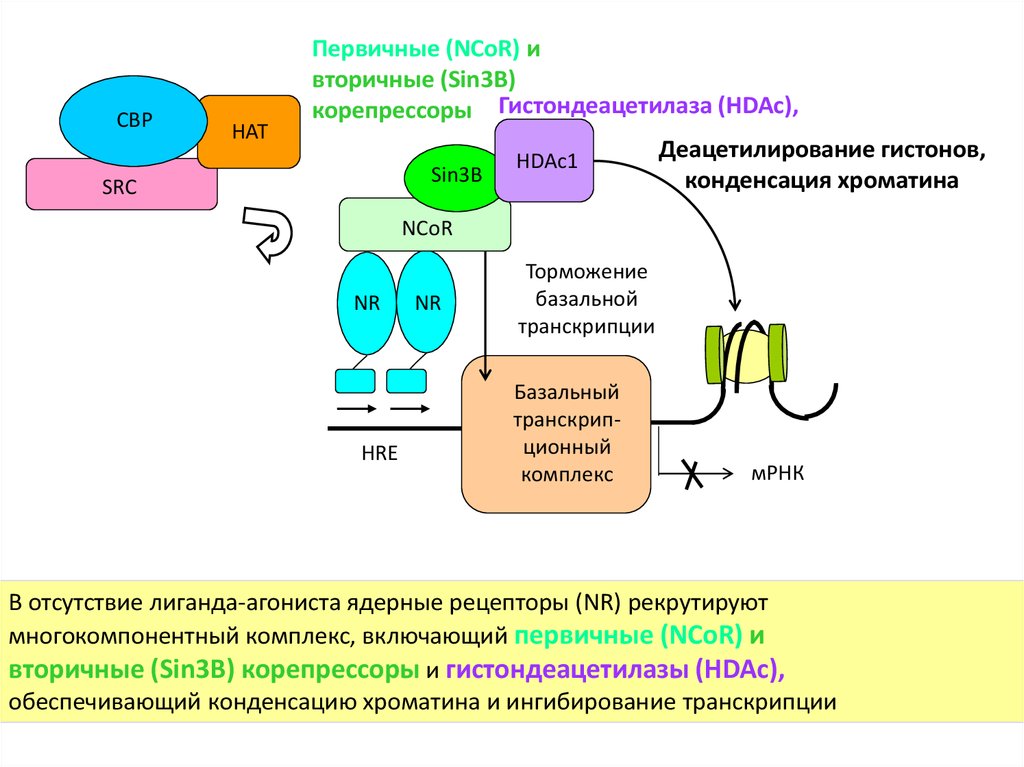
CBPHAT
Первичные (NCoR) и
вторичные (Sin3B)
корепрессоры Гистондеацетилаза (HDAc),
Sin3B
SRC
HDAc1
Деацетилирование гистонов,
конденсация хроматина
NCoR
NR
HRE
NR
Торможение
базальной
транскрипции
Базальный
транскрипционный
комплекс
мРНК
В отсутствие лиганда-агониста ядерные рецепторы (NR) рекрутируют
многокомпонентный комплекс, включающий первичные (NCoR) и
вторичные (Sin3B) корепрессоры и гистондеацетилазы (HDAc),
обеспечивающий конденсацию хроматина и ингибирование транскрипции
45.
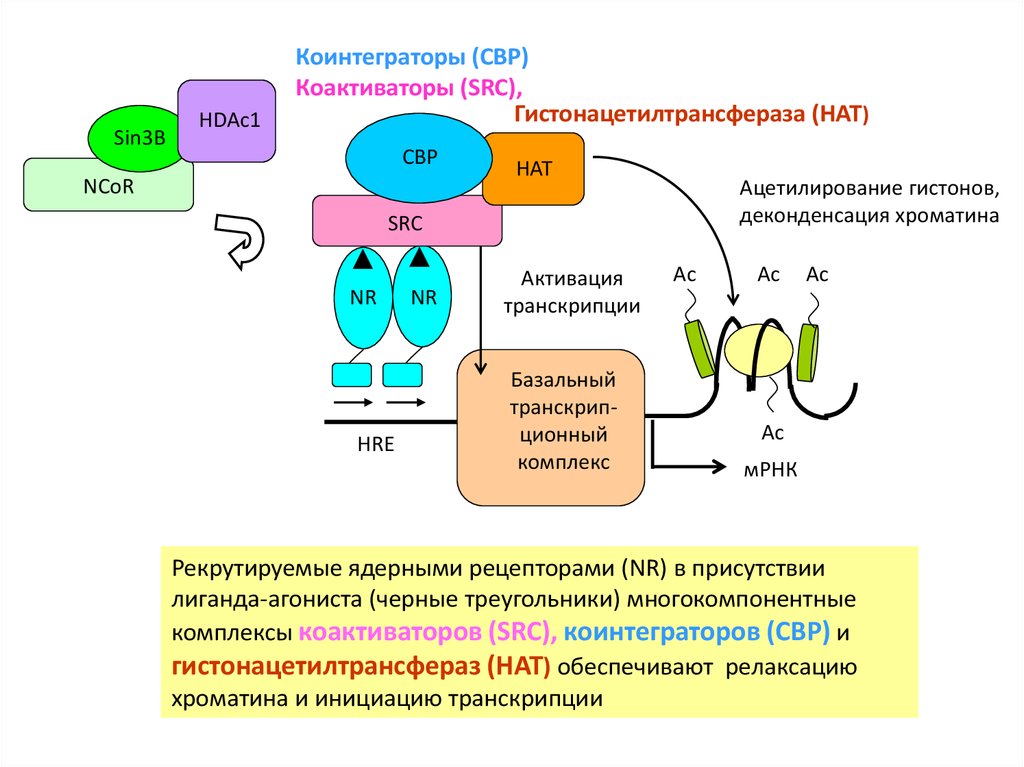
Sin3BHDAc1
Коинтеграторы (CBP)
Коактиваторы (SRC),
Гистонацетилтрансфераза (HAT)
CBP
NCoR
HAT
Ацетилирование гистонов,
деконденсация хроматина
SRC
NR
HRE
NR
Активация
транскрипции
Базальный
транскрипционный
комплекс
Ac
Ac
Ac
Ac
мРНК
Рекрутируемые ядерными рецепторами (NR) в присутствии
лиганда-агониста (черные треугольники) многокомпонентные
комплексы коактиваторов (SRC), коинтеграторов (CBP) и
гистонацетилтрансфераз (HAT) обеспечивают релаксацию
хроматина и инициацию транскрипции
46.
Роль нуклеосомКоличество глюкокортикоидчувствительных элементов (ГК-ЧЭ) в зависимости от их
локализации относительно старта транскрипции регулируемых гормоном генов
Количество ГК-ЧЭ
16
12
8
4
0
-40
-20
0
т.п.н.
20
40
47.
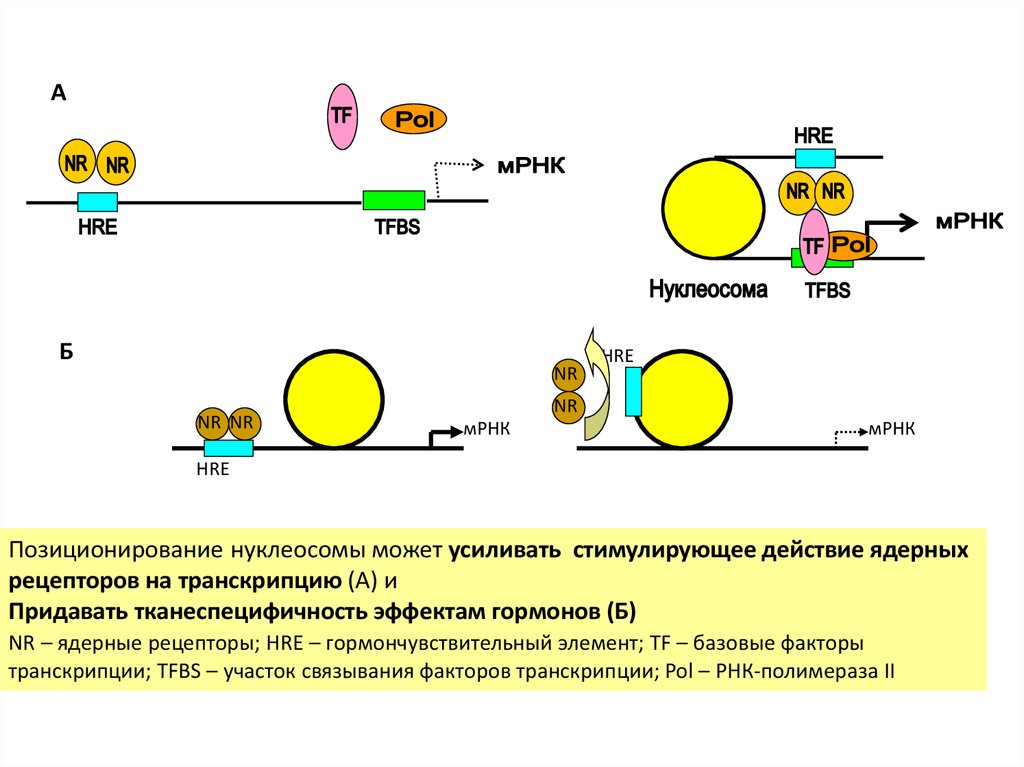
АБ
NR
NR NR
HRE
NR
мРНК
мРНК
HRE
Позиционирование нуклеосомы может усиливать стимулирующее действие ядерных
рецепторов на транскрипцию (А) и
Придавать тканеспецифичность эффектам гормонов (Б)
NR – ядерные рецепторы; HRE – гормончувствительный элемент; TF – базовые факторы
транскрипции; TFBS – участок связывания факторов транскрипции; Pol – РНК-полимераза II
48.
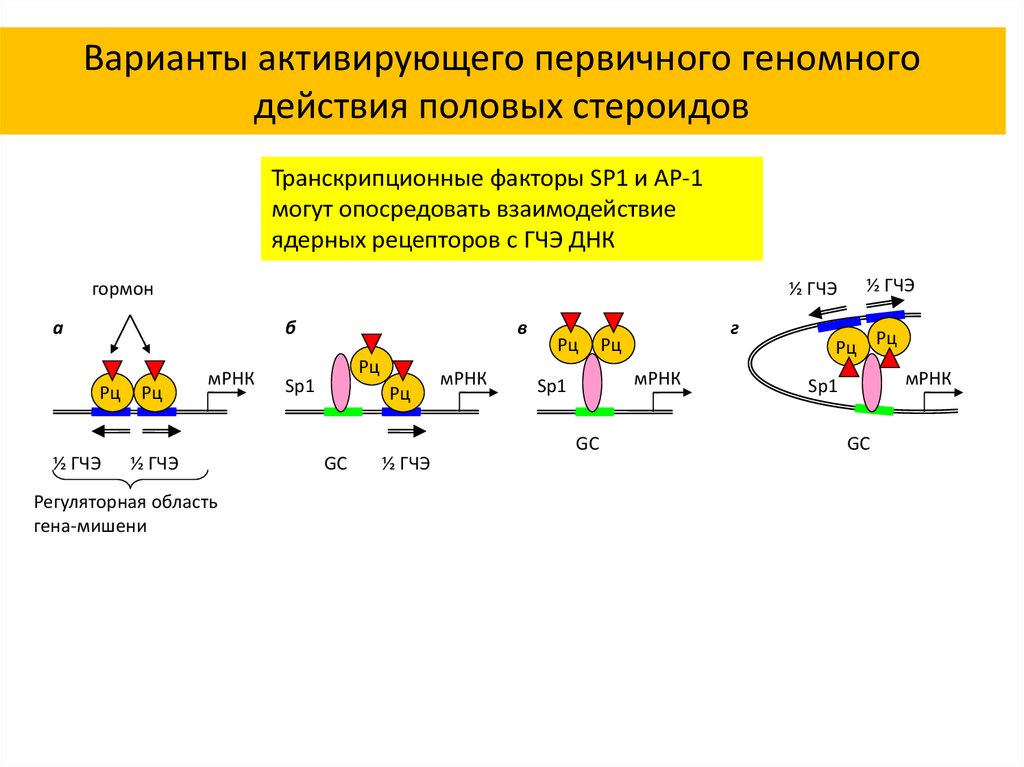
Варианты активирующего первичного геномногодействия половых стероидов
Транскрипционные факторы SP1 и AP-1
могут опосредовать взаимодействие
ядерных рецепторов с ГЧЭ ДНК
гормон
½ ГЧЭ
а
б
Рц
½ ГЧЭ
Рц
мРНК
½ ГЧЭ
Регуляторная область
гена-мишени
в
Рц
Sp1
Рц
GC
½ ГЧЭ
мРНК
Рц
г
Рц
мРНК
Sp1
GC
½ ГЧЭ
Рц Рц
мРНК
Sp1
GC