
- процесс биосинтеза полипептидных цепей белков в живых клетках")















 biology
biologySimilar presentations:

")

")
")
")




Трансляция
1. Трансляция
МЛ-1162. Трансляция (от лат. translatio-перевод)- процесс биосинтеза полипептидных цепей белков в живых клетках
Трансляция (от лат. translatioперевод)- процесс биосинтезаполипептидных цепей белков в
живых клетках
3. План презентации:
1. Строение рибосомы2. Принципы и условия трансляции
3. Образование аминоацил-тРНК
4. Этапы трансляции:
Инициация
Элонгация
Терминация
5. Основные отличия и особенности трансляции у
эукариот и прокариот
6. Биологическое значение Трансляции.
4. Строение рибосомы
Рибосома — важнейший органоид живой клетки сферической илислегка эллипсоидной формы, диаметром 100—200 ангстрем, состоящий
из большой и малой субъединиц. Рибосомы служат для биосинтеза
белка из аминокислот по заданной матрице на основе генетической
информации, предоставляемой матричной РНК, или мРНК.
В эукариотических клетках рибосомы располагаются на мембранах
эндоплазматического ретикулума, хотя могут быть локализованы и в
неприкрепленной форме в цитоплазме. Нередко с одной молекулой
мРНК ассоциировано несколько рибосом, такая структура называется
полирибосомой. Синтез рибосом у эукариот происходит в специальной
внутриядерной структуре — ядрышке. Рибосомы эукариот включают
четыре молекулы рРНК, из них 18S, 5.8S и 28S рРНК синтезируются в
ядрышке РНК полимеразой I в виде единого предшественника (45S),
который затем подвергается модификациям и нарезанию. 5S рРНК
синтезируется РНК полимеразой III в другой части генома и не
нуждаются в дополнительных модификациях.
5.
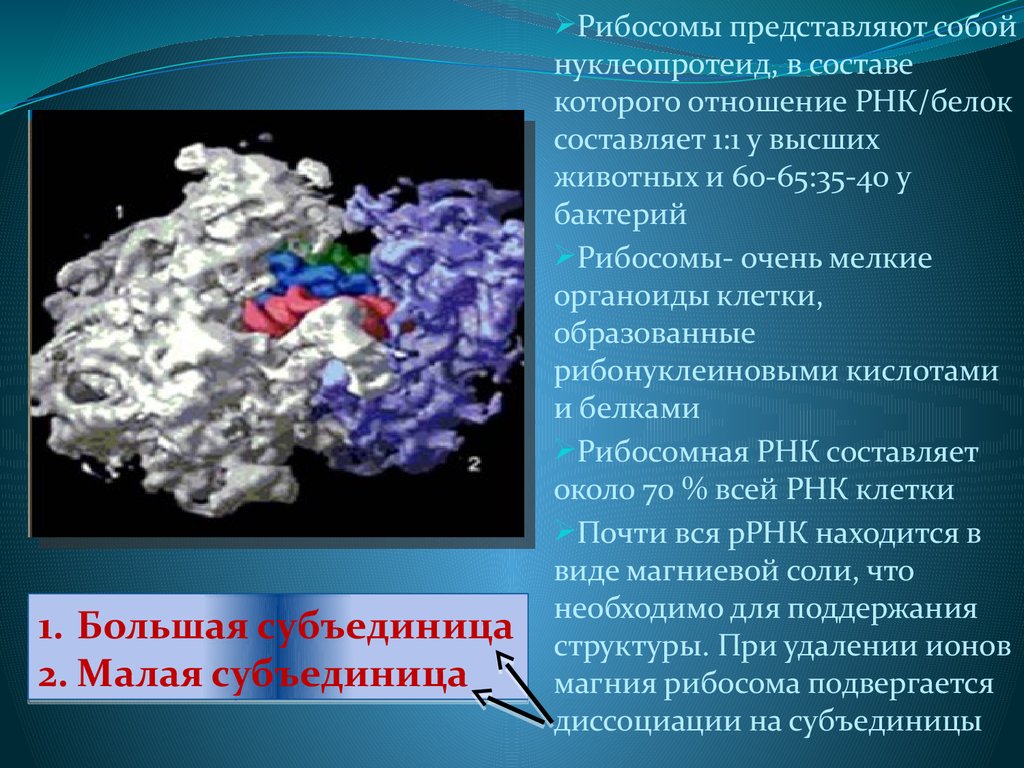
Рибосомы представляют собой1. Большая субъединица
2. Малая субъединица
нуклеопротеид, в составе
которого отношение РНК/белок
составляет 1:1 у высших
животных и 60-65:35-40 у
бактерий
Рибосомы- очень мелкие
органоиды клетки,
образованные
рибонуклеиновыми кислотами
и белками
Рибосомная РНК составляет
около 70 % всей РНК клетки
Почти вся рРНК находится в
виде магниевой соли, что
необходимо для поддержания
структуры. При удалении ионов
магния рибосома подвергается
диссоциации на субъединицы
6.
Каждая рибосома состоит из двух частиц- малой ибольшой. Малая субъединица имеет два участка
связывания: А-участок - аминоацил-тРНК-связывающий (для
поступающих аминокислот), Р-участок аминоацилпептидил-тРНК-связывающий (для
синтезируемой полипептидной цепочки. На большой
субъединице находится центр, катализирующий образование
пептидной связи. Так как пептидная связь образуется путем
переноса синтезируемого пептида на аминоацил-тРНК, он
носит название пептидилтрансферазного центра. На большой
субъединице находится также участок, связывающий
вспомогательные белки, называемыефакторами трансляции.
Субъединицы рибосомы способны разделяться и
объединяться при участии дополнительных белков.
Основной функцией рибосом является синтез белков
7. Трансляция
Принципы :Условия:
Матричность
мРНК
Рибосомы
Аминокислоты
тРНК
Ферменты(факторы
трансляции IF)
Энергия(АТФ;ГТФ)
Среда.
8. Схема образования аминоацил-тРНК
На первом этапе происходит активация аминокислоты АТРазой:аминокислота+ АТР→аминоацил-AMP +РР(пирофосфат).
На втором этапе активированная аминокислота соединяется с соответствующей
тРНК:
аминоацил-AMP + тРНК→аминоацил-тРНК + АМР.
Суммарное уравнение двух реакций:
аминокислота + тРНК + ATP →аминоацил-тРНК + AMP + PPi
9. Этапы трансляции
В сильно упрощённом виде трансляция включает следующие стадии. Синтезированная вклеточном ядре в ходе транскрипции молекула иРНК поступает в цитоплазму,
претерпевает ряд модификаций и соединяется с рибосомой (в клетках прокариот, не
разделённых на ядро и цитоплазму, иРНК связывается с рибосомой сразу). Находящиеся
в цитоплазме аминокислоты активируются взаимодействием с богатым энергией
соединением – АТФ. Поскольку аминокислоты и иРНК в силу их химического строения
«не соответствуют» друг другу (не могут взаимодействовать), между ними существует
своего рода переходник – транспортные РНК (тРНК). Активированные специальным
ферментом аминокислоты с участием этого же фермента (для каждого вида
аминокислоты – своего) соединяются т-РНК, также только со своей. Далее тРНК, несущая
аминокислоту, поступает на рибосому и своим антикодоном (тройкой нуклеотидов),
узнав на иРНК свой кодон (комплентарную тройку нуклеотидов), закрепляется на иРНК
на единственном свободном месте рядом со строящейся полипептидной цепью.
Специальный фермент рибосомы образует пептидную связь между аминокислотой и
синтезируемым полипептидом, а рибосома сдвигается по цепи иРНК на один кодон,
освобождая место для присоединения следующей тРН К. Так происходит наращивание
полипептидной цепи до тех пор, пока рибосома не дойдёт до «стоп-кодона». Получив
сигнал окончания синтеза, белковые факторы терминации освобождают полипептидную
цепь от рибосомы. Таким образом кодоны иРНК определяют последовательность
аминокислот в белке, а следовательно, его строение, свойства и активность.
10. Инициация
Стадия инициации, являющаяся «точкой отсчета» начала синтеза белка, требуетсоблюдения ряда условий, в частности наличия в системе, помимо 70S (или 80S) рибосом,
инициаторной амино-ацил-тРНК (аа-тРНК), инициирующих кодонов в составе мРНК и
белковых факторов инициации. Экспериментально доказано, что синтез белка инициирует
единственная аминокислота – метионин. Соответственно эти тРНК принято обозначать
тРНКфМет и тРНКМет. Укажем также, что эукариотическая клетка не нуждается в
формилировании метионина. У прокариот синтез N-формилметионил-тРНК протекает в
две стадии:
Данную стадию катализирует метионил-тРНК-синтетаза. Реакция нуждается в
доставке энергии гидролиза АТФ.
11.
Катализирующая II стадию трансформилаза оказалась более специфичной, чем метионил-тРНКсинтетаза: она не формилирует ни свободный метионин, ни метионин в комплексе с тРНКМет. Такимобразом, N-формилметионил-тРНК является первой аа-тРНК, которая определяет включение N-концевого
остатка аминокислоты и тем самым начало трансляции. К настоящему времени выяснена природа
белковых факторов инициации. У Е. coli открыты три таких инициирующих фактора, обозначаемых
соответственно IF-1, IF-2, IF-3. Все они получены в высокоочищенном состоянии с примерными
молекулярными массами 9000, 10000 и 22000 соответственно. IF-3 обеспечивает узнавание участка на
молекуле мРНК, к которому присоединяется формилметионил-тРНК. Данный белковый фактор первым
связывается со свободной 30S субчастицей рибосомы и препятствует ассоциации 30S и 50S субчастиц в
70S рибосому без молекулы мРНК. IF-1 способствует связыванию инициатор-ной формилметионил-тРНК с
комплексом 30S субчастицы и мРНК. Белковый фактор IF-2, вероятнее всего, способствует объединению
30S и 50S субчастиц после того, как на первой субчастице уже присутствуют инициирующие кодоны мРНК,
N-формилметионил-тРНК, IF-3, IF-1 и ГТФ. Этот белок рассматривают как фактор стабилизации всего
инициаторного 70S комплекса. Аналогичные белковые факторы инициации обнаружены также в эукариотических клетках. Открыто около 10 эукариотических белковых факторов инициации, их принято
обозначать eIF. Все они, по-видимому, важны для инициации, однако только три из них абсолютно
необходимы и существенны для белкового синтеза: eIF-2, eIF-3 и eIF-5. Они получены в чистом виде: eIF-2
состоит из α-, β- и γ-субъединиц (мол. масса 38000, 47000 и 50000 соответственно), eIF-3 (мол. масса
500000–700000) и eIF-5 (мол. масса 125000).
12.
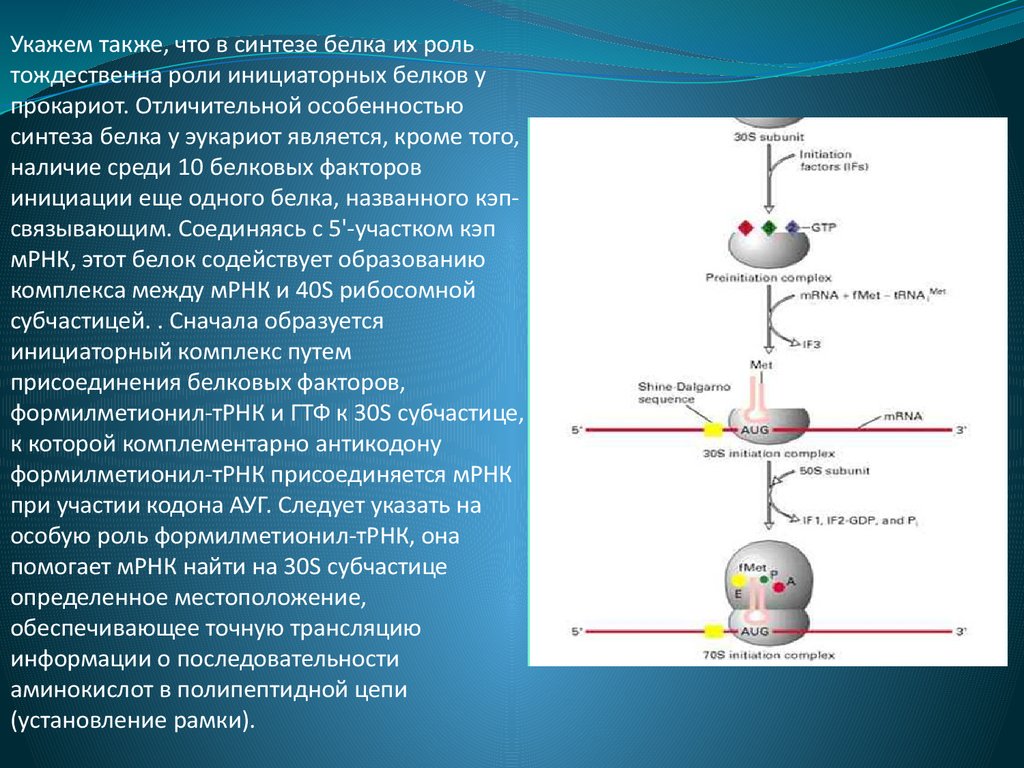
Укажем также, что в синтезе белка их рольтождественна роли инициаторных белков у
прокариот. Отличительной особенностью
синтеза белка у эукариот является, кроме того,
наличие среди 10 белковых факторов
инициации еще одного белка, названного кэпсвязывающим. Соединяясь с 5'-участком кэп
мРНК, этот белок содействует образованию
комплекса между мРНК и 40S рибосомной
субчастицей. . Сначала образуется
инициаторный комплекс путем
присоединения белковых факторов,
формилметионил-тРНК и ГТФ к 30S субчастице,
к которой комплементарно антикодону
формилметионил-тРНК присоединяется мРНК
при участии кодона АУГ. Следует указать на
особую роль формилметионил-тРНК, она
помогает мРНК найти на 30S субчастице
определенное местоположение,
обеспечивающее точную трансляцию
информации о последовательности
аминокислот в полипептидной цепи
(установление рамки).
13. Элонгация
Процесс элонгации полипептидной цепи у Е. coli начинается собразования первой пептидной связи и непосредственно, точнее
топографически, связан с большой субчастицей (50S) рибосомы,
содержащей два центра для связывания тРНК, один из них называется
аминоацильным (А), другой – пептидильным (П)
В процессе элонгации у Е. coli также участвует три белковых фактора –
элонгационные факторы трансляции, сокращенно обозначаемые Tu, Ts и
G . У эукариот также открыты три таких фактора, названных
эукариотическими элонгационными факторами трансляции и
обозначаемых соответственно eEF-1α, eEF-1αβ и eEF-2, почти все они
получены в чистом виде, для ряда из них установлена первичная
структура.
Процесс элонгации принято делить на 3 стадии: узнавание кодона и
связывание аминоацил-тРНК, образование пептидной связи и
транслокация.
14.
На I стадии в соответствии с природой кодона мРНК в свободный А-участокрибосомы доставляется аминоацил-тРНК при участии фактора элонгации Tu.
Этот процесс требует затраты энергии и сопряжен с гидролизом ГТФ и
образованием прочно связанного комплекса Тu–ГТФ. Образовавшийся
комплекс подвергается диссоциации только в присутствии второго фактора
элонгации Ts, при котором освободившийся фактор Tu может вновь,
соединяясь с молекулой ГТФ, принять участие в доставке аа-тРНК в рибосому.
Таким образом, в транслирующей 70S рибосоме в пептидильном центре
располагается формилметионил-тРНК, а в А-центре – аминоацил-тРНК
(первая аминокислота после метионина). С этого момента начинается II
стадия элонгации – образование первой пептидной связи. Для этого в
рибосоме осуществляется ферментативная реакция транспептидирования
между формилметионил-тРНК в П-центре и новой аа-тРНК в А-центре. В
процессе этой реакции остаток формилметионина переносится на свободную
NH2-группу аа-тРНК и замыкается первая пептидная связь в будущей
полипептидной цепи. Параллельно из пептидильного центра освобождается
тРНКфМет в цитозоль. Фермент, катализирующий реакцию
транспептирования, получил название пептидил-трансферазы. Таким
образом, в процессе транспептидазной реакции в А-центре образуется
дипептидил-тРНК, а П-центр остается свободным («вакантным»).
15.
На III стадии процесса элонгации необходимо иметь свободный аминоацильный центр для присоединения следующей аа-тРНК. Для этогоблагодаря процессу транслокации образовавшийся фрагмент
дипептидил-тРНК переносится от аминоацильного на пептидильный
центр. Достигается транслокация благодаря передвижению рибосомы
относительно мРНК при участии фермента транслоказы (функцию ее
выполняет фактор элонгации G у Е. coli и eEF-2 у эукариот) за счет
использования энергии распада еще одной молекулы ГТФ. В результате
транслокации дипептидил-тРНК занимает место в пептидильном
центре рибосомы, а аминоацильный центр освобождается для нового
цикла узнавания и может присоединить новую следующую аа-тРНК,
соответствующую кодону мРНК. В процессе транслокации рибосома
перемещается вдоль мРНК по направлению к ее 3'-концу на расстояние в
один кодон, т.е. точно на один триплет. Таким образом, на стадии
элонгации происходит последовательное наращивание полипептидной
цепи по одной аминокислоте в строгом соответствии с
последовательностью триплетов (кодонов) в молекуле мРНК.
16.
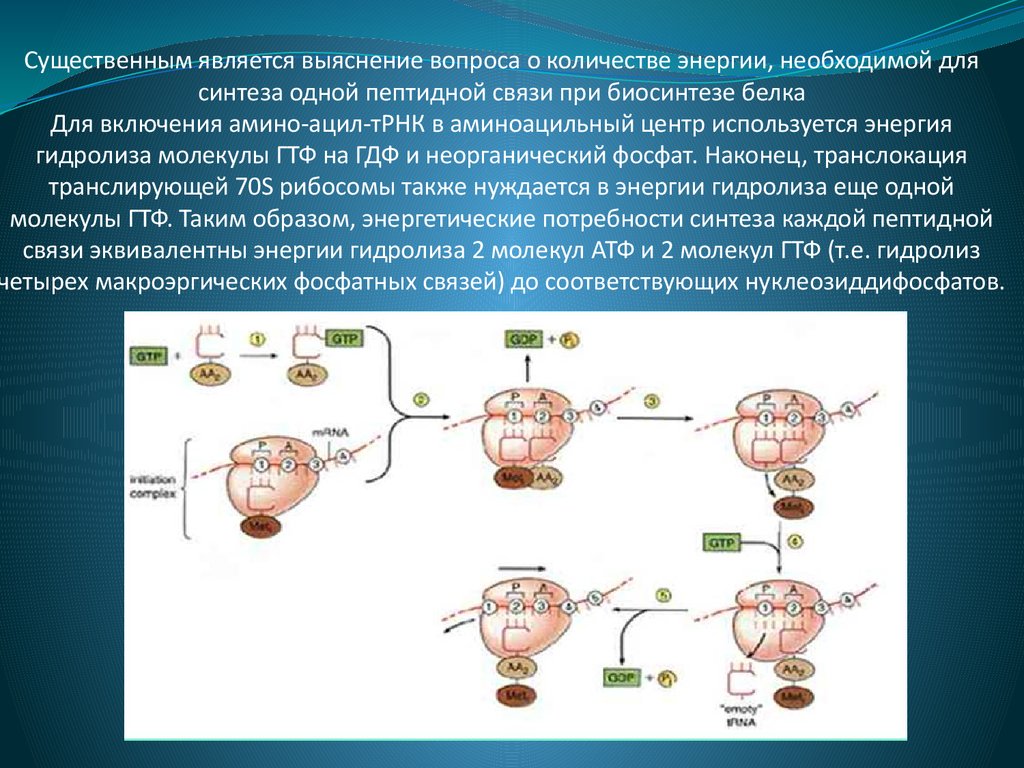
Существенным является выяснение вопроса о количестве энергии, необходимой длясинтеза одной пептидной связи при биосинтезе белка
Для включения амино-ацил-тРНК в аминоацильный центр используется энергия
гидролиза молекулы ГТФ на ГДФ и неорганический фосфат. Наконец, транслокация
транслирующей 70S рибосомы также нуждается в энергии гидролиза еще одной
молекулы ГТФ. Таким образом, энергетические потребности синтеза каждой пептидной
связи эквивалентны энергии гидролиза 2 молекул АТФ и 2 молекул ГТФ (т.е. гидролиз
четырех макроэргических фосфатных связей) до соответствующих нуклеозиддифосфатов.
17. Терминация
Терминация - синтез полипептидной цепи в 70S рибосоме при участии трех белковыхфакторов терминации (рилизинг-факторов). Эти белки обозначаются RF-1 (мол. масса
47000), RF-2 (мол. масса 35000–48000) и RF-3 (мол. масса 46000) у прокариот. В клетках
животных открыт один-единственный белок с аналогичным свойством – рилизингфактор R (eRF, мол. масса 56000–105000). У Е. coli RF-1 наделен свойством узнавания в
молекуле мРНК терминирующих кодонов УАГ и УАА , a RF-2 – соответственно УГА и УАА.
Эукариотический рилизинг-фактор eRF узнает все три терминирующих кодона (нонсенскодоны) и индуцирует освобождение синтезированного полипептида опосредованно
через пептидил-трансферазу. После того как терминирующий кодон мРНК занимает
свое место в аминоацильном центре рибосомы, к нему присоединяется не тРНК,
поскольку отсутствуют соответствующие антикодоны тРНК, узнающие этот
терминальный сигнал, а один из белковых факторов терминации и блокируется
дальнейшая элонгация цепи. Считают, что терминирующие кодоны и белковые факторы
индуцируют изменение специфичности пеп-тидилтрансферазной активности таким
образом, что она катализирует перенос растущей пептидной цепи, скорее, к молекуле
воды, вызывая гидролиз, чем к аминогруппе аминокислоты.
18.
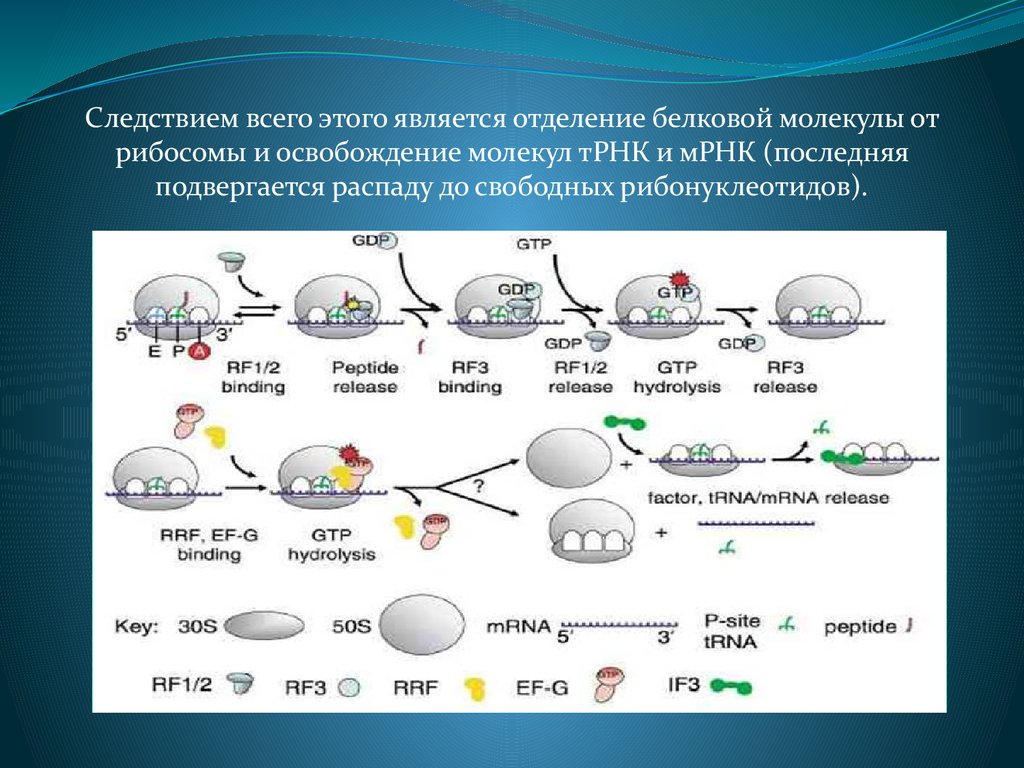
Следствием всего этого является отделение белковой молекулы отрибосомы и освобождение молекул тРНК и мРНК (последняя
подвергается распаду до свободных рибонуклеотидов).
19. Особенности трансляции у прокариот и эукариот
Прокариоты. Синтез белка происходит нарибонуклеопротеиновом комплексе - рибосоме, в
процессе трансляции mRNA. Рибосома состоит из
большой и малой субъединиц, которые соединены
в области инициации трансляции (translation
initiation region -TIR) mRNA во время стадии
инициации трансляции. Во время элонгации
рибосома скользит вдоль mRNA и синтезирует
полипептидную цепь. Элонгация продолжается до
тех пор, пока рибосома не достигает стоп-кодона на
mRNA - терминация трансляции. После
терминации рибосома отделяется от
синтезированного полипептида и способна снова
повторить цикл трансляции mRNA.
Каждая стадия трансляции имеет свои
регуляторные факторы, но у эукариот этих
факторов гораздо больше, чем у прокариот.
20.
Эукариоты. Вместо комплементарного РНК-РНКузнавания, в которое вовлечена прединициирующая
последовательность Шайна-Дальгарно прокариотических
мРНК, эукариотические мРНК узнаются
эукариотическими рибосомами по кэпированному 5'концу с обязательным участием белка, например, eIF-4F
инициаторного фактора. Предполагается, что этот белок
участвует в расплавлении вторичных структур 5'- областей
мРНК, облегчая их связывание с малыми субчастицами
рибосом. В отличие от прокариот, эукариотическая мРНК
образует комплексы с белками , что обусловливает ее
метаболическую стабильность. Вследствие этого у
эукариот отсутствует постоянная интенсивная деградация
и интенсивный ресинтез мРНК, которые, как правило,
моноцистронны и имеют специфически
модифицированный (кэпированный) 5'-конец. Все это
обусловливает целый ряд особенностей инициации
трансляции и ее регуляции у эукариотических
организмов. Естественно, что метаболическая
стабильность эукариотической мРНК делает регуляцию на
уровне трансляции особенно важной в общей картине
регуляции биосинтеза белка.
21. Биологическое значение трансляции
Трансляция крайне важный процесс, и нарушение или выпадение любогозвена, участвующего в синтезе белка, почти всегда приводит к развитию
патологии, причем клинические проявления болезни будут определяться
природой и функцией белка, синтез которого оказывается нарушенным
(структурный или функциональный белок). Иногда синтезируются так
называемые аномальные белки как результат действия мутагенных факторов
и, соответственно, изменения генетического кода (например, гемоглобин
при серповидно-клеточной анемии). Последствия этих нарушений могут
выражаться в развитии самых разнообразных синдромов или заканчиваться
летально. Следует отметить, что организм располагает мощными
механизмами защиты: подобные изменения генетического аппарата быстро
распознаются специфическими ферментами — рестриктазами, измененные
последовательности вырезаются и вновь замещаются соответствующими
нуклеотидами при участии полимераз и лигаз.