















































 biology
biologySimilar presentations:








")

Нуклеиновые кислоты
1. Нуклеиновые кислоты
2.
• Нуклеиновые кислоты являютсявысокомолекулярными
органическими соединениями.
Существует два типа НК:
рибонуклеиновые кислоты (РНК) и
дезоксирибонуклеиновые кислоты
(ДНК). Эти соединения играют
важную роль в биологических
системах, начиная с вирусов и
заканчивая клетками высших
животных, участвуя в хранении (ДНК)
и реализации(РНК) генетической
информации.
3.
• Следует иметь ввиду, что одни НК немогут быть достаточно стабильными
хранителями генетической
информации, и тем более , одни они
не в состоянии обеспечить перевод
этой информации в конкретные
биологические структуры: клетки,
ткани, органы. Даже вирусы
представляют собой комплекс НК и
белков, т.е. нуклеопротеидный
комплекс.
4.
• Молекулы НК являются самымибольшими биологическими
полимерами. Так молекулярная
масса РНК может достигать
10 – 14 106 ,а молекулярная масса
ДНК – 1010- 1011.
Длина такой молекулы ДНК может
доходить до 8см при диаметре всего
в 2 нм (соотношение длины
молекулы и её диаметра
приближается к 107).
5.
• Такая молекула крайне нестабильнаи в отсутствии стабилизирующих её
белков легко разламывается.
• При всех своих гигантских размерах
молекулы НК устроены
сравнительно просто: они, подобно
белкам, состоят из весьма
ограниченного числа мономерных
единиц, соединённых в длинные
неразветвлённые полимерные цепи.
6.
• При полном гидролизе НК вгидролизате могут быть обнаружены
соединения трёх типов:
• фосфорная кислота;
• углевод ( рибоза или дезоксирибоза)
• органические соединения;
представляющие собой различные
производные гетероциклов пурина
или пиримидина.
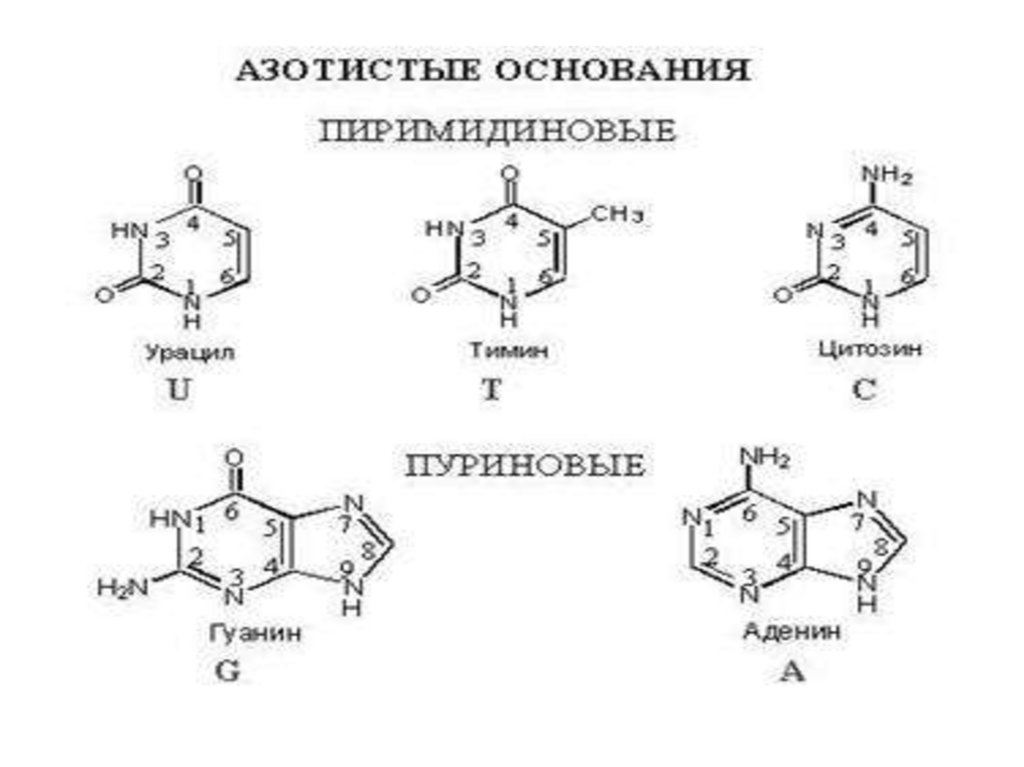
7.
8.
• На долю аденина (А), гуанина (Г),цитозина (Ц) и тимина (Т)
приходится 99 -98% от общего их
количества.
Эти основания получили название
главных азотистых оснований ДНК.
9. Минорные азотистые основания
• Минорные азотистые основания –эторедко встречающиеся азотистые
основания. Они составляют 1 -2% в
составе ДНК и до 15% в составе РНК.
• По своей химической природе они
обычно являются метилированными
или гидроксиметилированными
производными главных азотистых
оснований.
10. Минорные азотистые основания
NH2СН2ОН
N
O
NH
5-гидроксиметилцитозин
11.
• Азотистые основанияпиримидинового или пуринового
ряда, имеющие в своём составе
кислород, могут существовать в
двух формах: лактамной (оксоформа) и лактимной (гидроксиформа).
• В составе НК азотистые основания
всегда присутствуют в лактамной
форме.
12. Лактим-лактамная таутомерия азотистых оснований
13. Нуклеозиды
14. Нуклеозиды
15. Нуклеотиды
• Если к остатку моносахарида внуклеозиде присоединён остаток
фосфорной кислоты, то такая
структура получила название
мононуклеотида (нуклеотида)
16. Нуклеотиды
17.
18. Нуклеотиды
Мононуклеотиды в клетках могут:• входить в состав НК;
• функционировать самостоятельно,
выступая или в качестве коферментов,
или в качестве регуляторов обменных
процессов. Так, АТФаденозинтрифосфорная кислота,
выполняет в клетках роль переносчика
энергии из реакций, идущих с
выделением энергии, в реакции,
идущих с поглощение энергии.
19. Нуклеотиды
• Энергия временно аккумулируется впирофосфатных связях молекулы
АТФ.
20.
• Примером нуклеотида,выполняющего в клетках роль
регулятора обменных процессов,
является циклическая
аденозинмонофосфорная кислота
или цАМФ.
• Циклические нуклеотиды участвуют
в регуляции содержания ионов Мg2+
в организме.
21. цАМФ
22. цАМФ
• В структуре цАМФ один остатокфосфорной кислоты соединён
одновременно и с третьим(С3), и с
пятым (С5) атомами углерода рибозы
сложноэфирными связями.
• Иногда в качестве коферментов
выступают не мононуклеотиды, а их
димеры. Типичным примером такого
димера является НАД Н2/НАД+ никотинамиддинуклеотид:
23. НАД Н2/НАД+
24. Первичная структура НК
• В процессе образованияполинуклеотида 2 мононуклеотида
соединяются друг с другом за счёт
взаимодействия остатка фосфорной
кислоты одного мононуклеотида у С5
с гидроксильной группой у С3 –
рибозы (или дезоксирибозы)
второго мононуклеотида:
25. Первичная структура НК
26. Первичная структура НК
• После выделения молекулы воды 2мононуклеотида оказываются
соединёнными сложноэфирной связью
между остатком фосфорной кислоты
одного мононуклеотида и С3-рибозы
другого. Такая связь часто называется
фосфодиэфирной связью, поскольку
остаток фосфорной кислоты образует
2 сложноэфирные связи-с остатком
углевода своего мононуклеотида и
остатком пентозы другого.
27. Первичная структура НК
• Образовавшийся нуклеотид имеетна одном конце остаток фосфорной
кислоты, а на другом - свободный
гидроксил у С3- рибозы.
• За счёт их может идти
присоединение следующих
мононуклеотидов, т.е. существует
принципиальная основа для
образования полимера –
полинуклеотида.
28. Первичная структура НК
• Количество мономерных звеньев вполимерной цепи может достигать
108 -109 единиц.
• Мононуклеотиды связаны в
полинуклеотид последовательно
так, что разветвлённых структур не
образуется.
• Последовательность соединения
мононуклеотидов в полинуклеотид
носит название первичной структуры
НК.
29. Первичная структура НК
• Полинуклеотидная цепь имеетначало и окончание.
• Началом считается тот конец
полинуклеотида, на котором
находится или свободная
гидроксильная группа С5-рибозы,
или же остаток фосфорной кислоты,
связанный с этим гидроксилом.
30. Первичная структура НК
• Окончанием полинуклеотидасчитается его конец, содержащий
или свободный гидроксил у С3пентозы или же остаток фосфорной
кислоты, связанный с этим
гидроксилом.
• 5,
3,- прямое направление
• 3,
5, - обратное направление.
31. Первичная структура НК
• Полинуклеотидную цепь принятоизображать в виде
последовательности сокращённых
названий азотистых оснований,
входящих в её состав нуклеотидов:
• рГ – А- А – Г – Ц –Т –А – Ц ОН
• 5,
3,
32.
• Изучение нуклеотидного состава ДНКпоказало, что:
- количество пиримидинов в молекуле
эквивалентно количеству пуринов
( Т+Ц=А+Г );
- А = Т;
- Г=Ц;
- А + Ц = Г + Т.
33.
• Тем не менее ДНК каждогоотдельного вида животного,
растения или микроорганизма
обладает характерным для данного
вида составом, который не зависит
от возраста, условий роста,
различных факторов развития и т.д.
• В то же время ДНК, выделенная из
различных клеток или органов
животного, имеет одинаковый
состав, характерный для данного
вида.
34. Вторичная структура ДНК
• Согласно современнымпредставлениям молекула ДНК
представляет собой не одиночную, а
двойную полинуклеотидную цепь,
спиралеобразно скрученную в
пространстве. Углеводнофосфатные основы цепей
расположены по периферии
структуры, а азотистые основания
упакованы внутри неё.
35. Вторичная структура ДНК
36. Вторичная структура ДНК
• Спираль имеет правую закрутку и накаждый виток спирали приходится
10 пар нуклеотидов, расстояние
между которыми составляет 0,34 нм.
• Таким образом, один виток спирали
занимает по длине 3,4 нм, диаметр
спирали составляет 2,0 нм.
• Направление полинуклеотидных
цепей антипараллельно, т.е. одна
цепь имеет направление 5
3, а
другая 3
5.
37. Вторичная структура ДНК
• Азотистые основания одной цепивзаимодействуют с азотистыми
основаниями другой цепи за счёт
образования водородных связей
между ними. Азотистое основание
тимин одной цепи всегда
взаимодействует с аденином другой, а
цитозин одной цепи взаимодействует с
гуанином в другой цепи.
• Эти пары азотистых оснований
называют комплементарными АО.
38. Вторичная структура ДНК
• Долгое время считалось, чтостабильность двойной спирали ДНК
обусловлена только водородными
связями между комплементарными
азотистыми основаниями
полинуклеотидных цепей. Однако,
выяснено, что одних водородных
связей для стабилизации недостаточно.
В настоящее время обнаружен ещё один
вид взаимодействия – стэкинг
взаимодействие.
39. Стэкинг –взаимодействие -
Стэкинг –взаимодействие • -суть его состоит в том, что плоскостиазотистых оснований молекулы ДНК,
перпендикулярные длинной оси
молекулы и параллельные друг другу,
образуют внутри спирали что-то вроде
монетного столбика. Каждое из
азотистых оснований взаимодействует
с выше и ниже лежащим азотистым
основанием за счёт своих
делокализованных систем пиэлектронов.
40. Стэкинг –взаимодействие
• В результата вся молекулаоказывается «прошитой» по длине
особым типом связей, которые
получили название стекинг –
взаимодействием.
41. Вторичная структура ДНК Стэкинг -взаимодействие
42. Вторичная структура ДНК
• Поскольку все азотистые основанияодной полинуклеотидной цепи ДНК
комплементарны азотистым
основаниям её цепи, то и в целом
одна полинуклеотидая цепь
комплементарна другой цепи.
43. Вторичная структура ДНК
• Регулярная пространственнаяструктура, возникающая в
результате взаимодействия двух
антипараллельных
комплементарных
полинуклеотидных цепей и
представляющая собой двойную
правозакрученную спираль, носит
название вторичной структуры ДНК.
44. РНК
• В отличие от ДНК молекулы РНК неимеют каких-либо выраженных
закономерностей в отношении
количественного состава входящих в
них азотистых оснований.
• К тому же молекулы РНК состоят из
одной полинуклеотидной цепи и
выраженной вторичной структуры на
всём протяжении молекулы РНК не
имеют.
45. РНК
• В качестве элементов вторичнойструктуры РНК обычно рассматривают
так называемые «шпильки»,
представляющие собой участки одной
и той же полинуклеотидной цепи,
перегнутые в пространстве и
закрученные сами на себя.
Стабилизация таких структур
осуществляется за счёт водородных
связей между азотистыми основаниями
восходящего и нисходящего участков
полинуклеотидной цепи.
46. РНК
• Полной комплементарности междувосходящим и нисходящим
участками в полинуклеотидной цепи
в такой шпильке нет и структура её
не так регулярна и прочна, как
аналогичная структура ДНК, но тем
не менее в некоторых классах
молекул РНК в образовании такого
рода элементов вторичной
структуры может участвовать до 60%
всех мононуклеотидных остатков.
47. РНК
• В то время как состав и структураДНК постоянны для клеток данного
организма, молекулу РНК даже в
пределах одной и той же клетки
гетерогенны по структуре и составу.
Кроме того, они выполняют
различные функции в сложном
процессе реализации генетической
информации. По функциональному
принципу клеточные РНК можно
разделить на 4 класса:
48. РНК
• Рибосомальная РНК – входит в составрибосом, клеточных органелл, на
которых происходит сборка
полипептидных цепей белков в
процессе их биосинтеза. На долю рРНК
приходится 60-65% всей клеточной РНК.
• Информационная(матричная
РНК).Функция этого класса РНК-перенос
генетической информации из ядра к
месту синтеза белка-рибосомам.Она
составляет 5%от всей клеточной РНК.
49.
• Транспортная РНК- её функциясостоит в отборе и транспортировке
аминокислот из цитоплазмы к
рибосомам. тРНК составляет до 10%
всей клеточной РНК.
• Ядерная РНК. Эта фракция состоит
главным образом из
предшественников молекул первых
трёх классов РНК, образующихся в
процессе синтеза РНК на ДНК. Эта
фракция составляет около 20% всей
клеточной РНК.