



































")










")






















































 biology
biologySimilar presentations:








")

Физиология высшей нервной деятельности
1.
Физиология высшейнервной деятельности
2.
Цель лекции:Дать механизмы развития сна и
сновидений, сознательного и
бессознательного
Мотивация лекции:
Данный раздел физиологии имеет
большое клиническое значение
3.
План лекции:- физиология сна, сновидений и гипноза
- физиология сознательного и
бессознательного
- физиология боли
4. Сон - физиологическое состояние неподвижности с ослабленным тонусом мышц и резко ограниченным сенсорным контактом с внешней
средой5. Сон – это состояние, характеризующееся:
отсутствием моторной активности,
пониженным уровнем сознания (у человека),
внешним покоем,
характерной позой (которая генетически
детерминирована),
• сенсорной изоляцией,
• определённым функциональным состоянием
мозга и внутренних систем, которые резко
отличаются от состояния при бодрствовании.
6. Функции сна:
покой и отдых,
восстановление сил организма,
переработка дневной информации и
перевод её в долговременную память,
поддержание творческой активности
личности,
оценивание уровня адаптационных
процессов и регуляция их
интенсивности
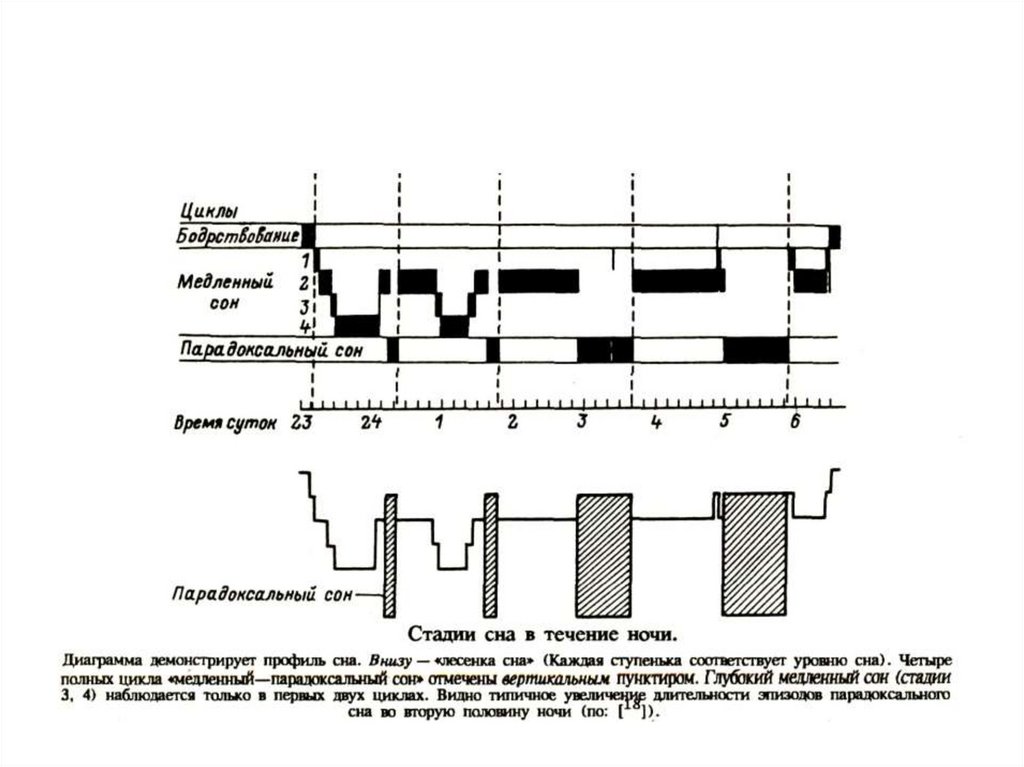
7. Виды сна
Ежесуточный естественный
– Монофазный и полифазный сон
– Медленный или ортодоксальный сон
– Быстрый или парадоксальный сон
Сезонный
Патологический
Наркотический
Гипнотический
8. Фазы сна
• Стадия дремоты - постепенное замещение альфаритма низкоамплитудными тета-волнами• Стадия сонных веретен - между двух-трехфазными
медленными колебаниями возникают сонные
веретена высокой амплитуды и частоты (12-16 гц)
• Стадия появления дельта-волн - до 50% ритмики
периодически занимают дельта-волны
• Стадия глубокого дельта-сна -более 50% ритмики
занимают дельта-волны
• Парадоксальный сон - десинхронизация ритмики
каждые 90-100 мин
9.
Соотношение сна и бодрствования, а также БДГ– и медленноволновогосна в различные периоды жизни человека. Наиболее существенное
изменение в раннем возрасте–уменьшение общей длительности сна и
значительное снижение в нем доли БДГ–фазы
10.
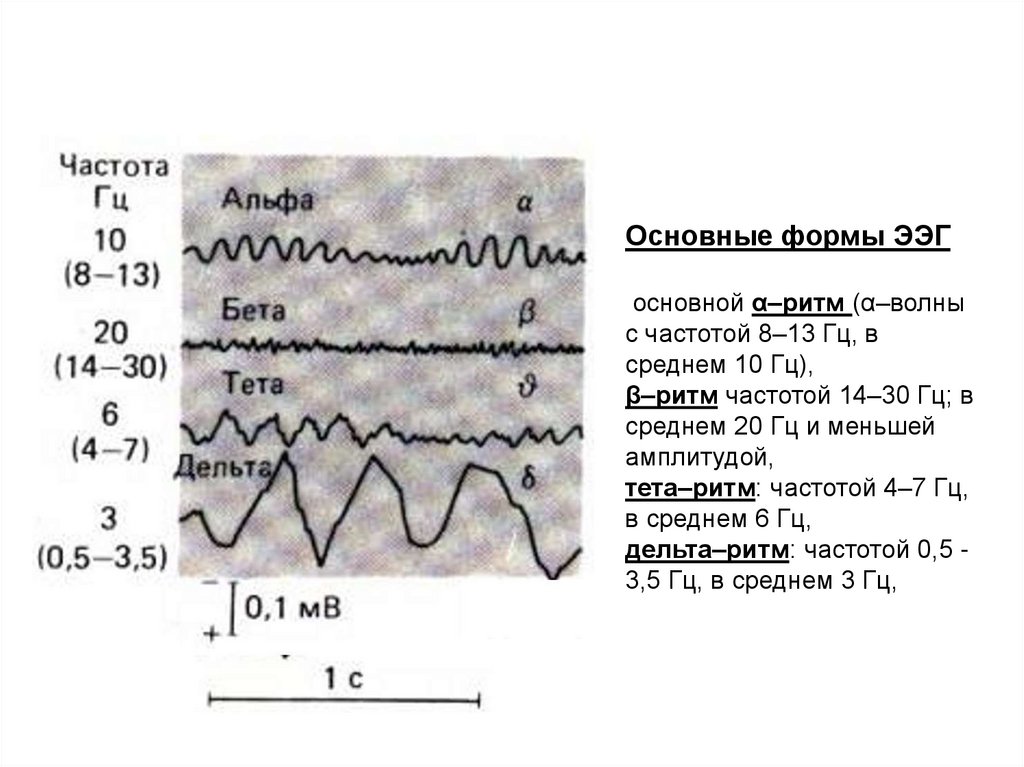
Основные формы ЭЭГосновной α–ритм (α–волны
с частотой 8–13 Гц, в
среднем 10 Гц),
β–ритм частотой 14–30 Гц; в
среднем 20 Гц и меньшей
амплитудой,
тета–ритм: частотой 4–7 Гц,
в среднем 6 Гц,
дельта–ритм: частотой 0,5 3,5 Гц, в среднем 3 Гц,
11.
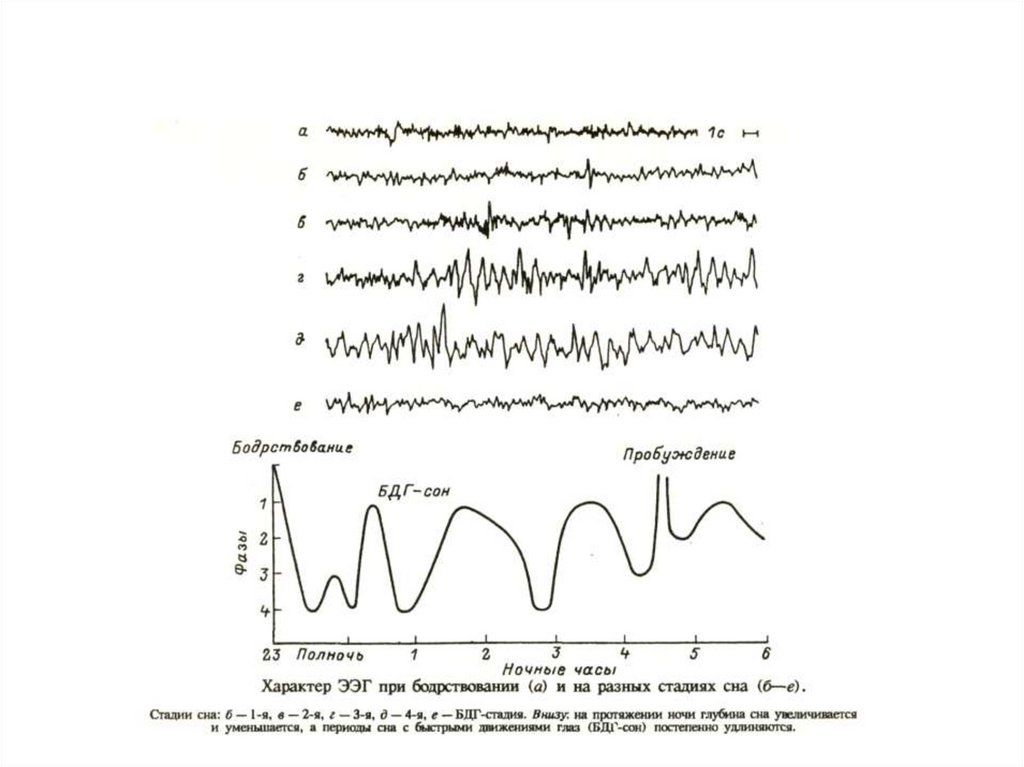
Классификация стадий сна у человека по особенностям ЭЭГ по ЛумисуСтадия W — бодрствование в расслабленном состоянии,
стадия А — переход от бодрствования ко сну,
стадия В — засыпание и самый поверхностный сон (острые вертекс—зубцы соответствуют
«моменту» засыпания),
стадия С — поверхностный сон,
стадия D — умеренно глубокий сон,
стадия Е — глубокий сон. Три нижние кривые представляют собой одновременную запись ЭЭГ,
электроокулограммы (ЭОГ) и электромиограммы указательного пальца (ЭМГ) во время сна с
быстрым движением глаз (БДГ) со сновидениями.
12.
13.
14.
15.
16. Центры сна
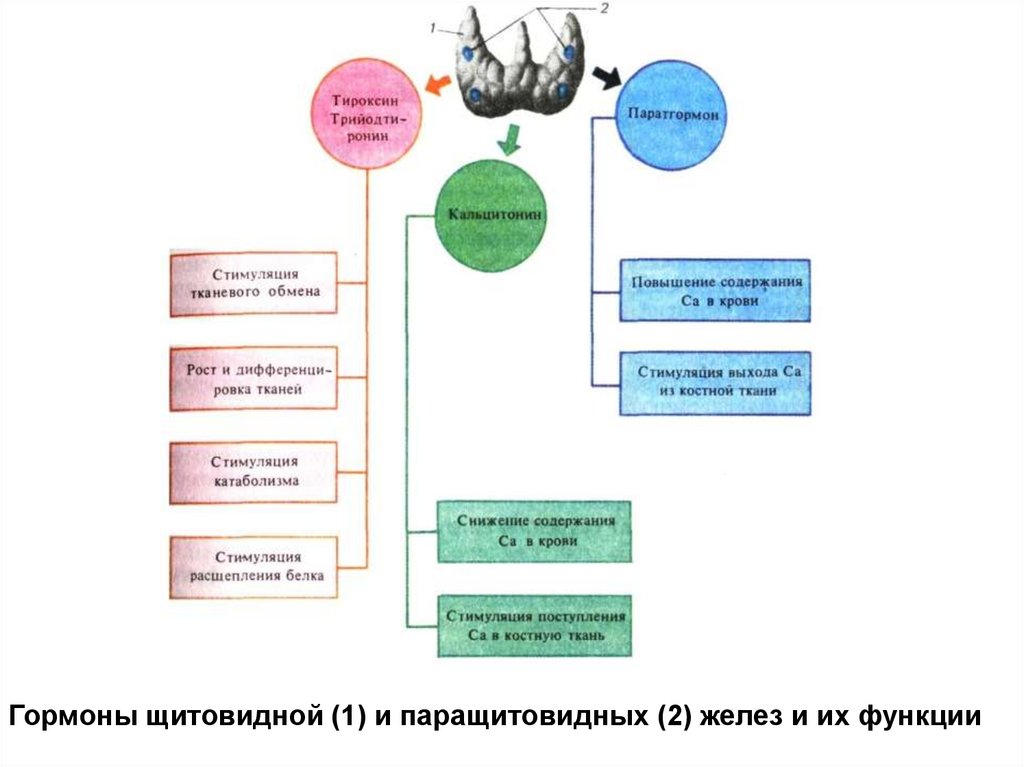
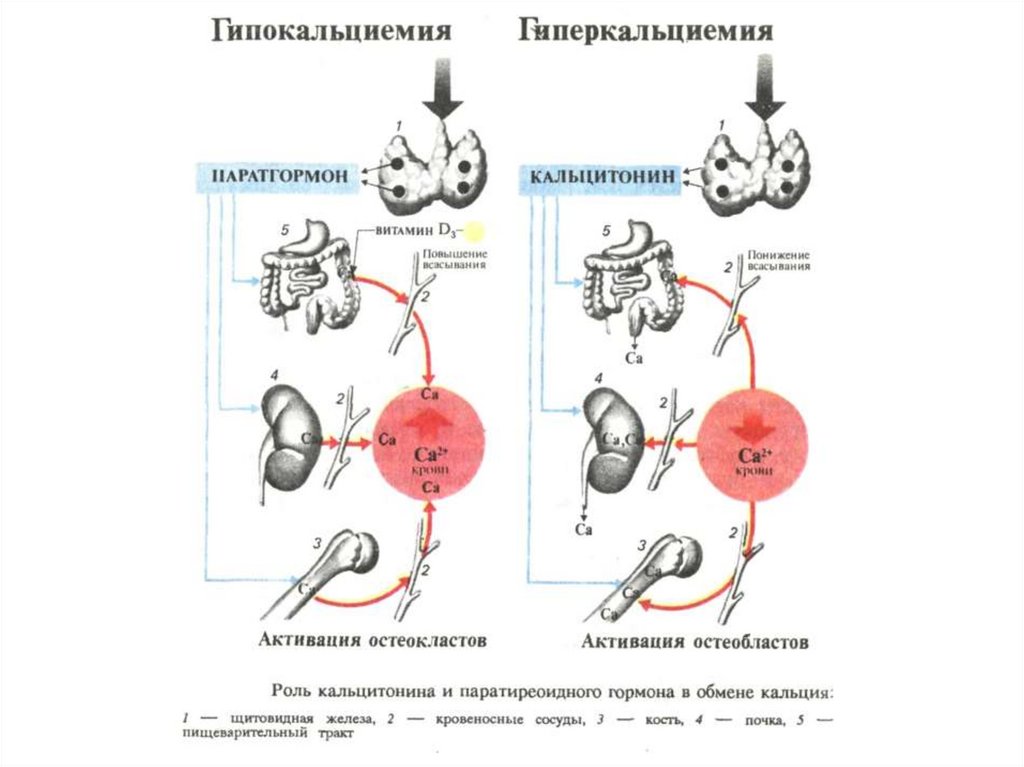
Синхронизирующие (сомногенные) структуры- структуры вокруг сильвиевого водопровода и
задней стенки IV желудочка,
- медиальный таламус,
- хвостатое ядро,
- базальные отделы переднего мозга
Десинхронизирующие (пробуждающие)
структуры
- РФ заднего и среднего мозга,
- ядра моста - голубое пятно и ядро шва,
- неспецифические ядра таламуса
17.
18. Нервные теории сна
Гесс: в ядрах гипоталамуса существует центр сна.Поражение гипоталамуса – увеличение сонливости.
Павлов: Сон – следствие иррадиации торможения,
распространяющееся по коре. Сон – особый вид
торможения, сходный с условным торможением, во
время которого происходит восстановление затраченных
нейронами ресурсов.
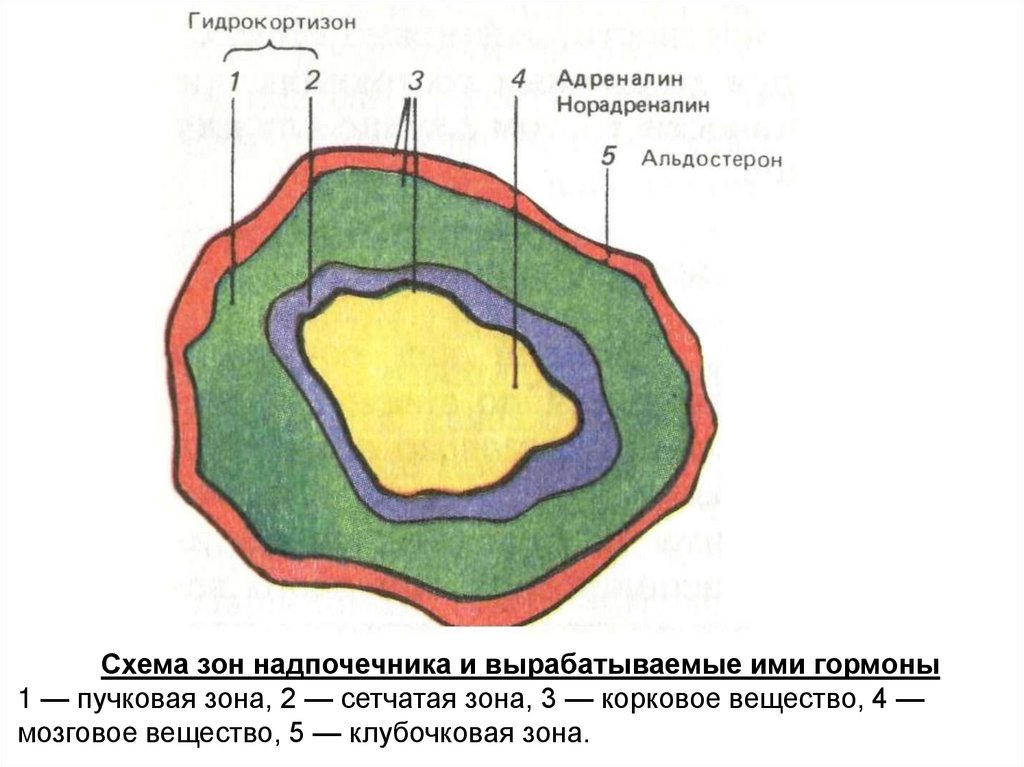
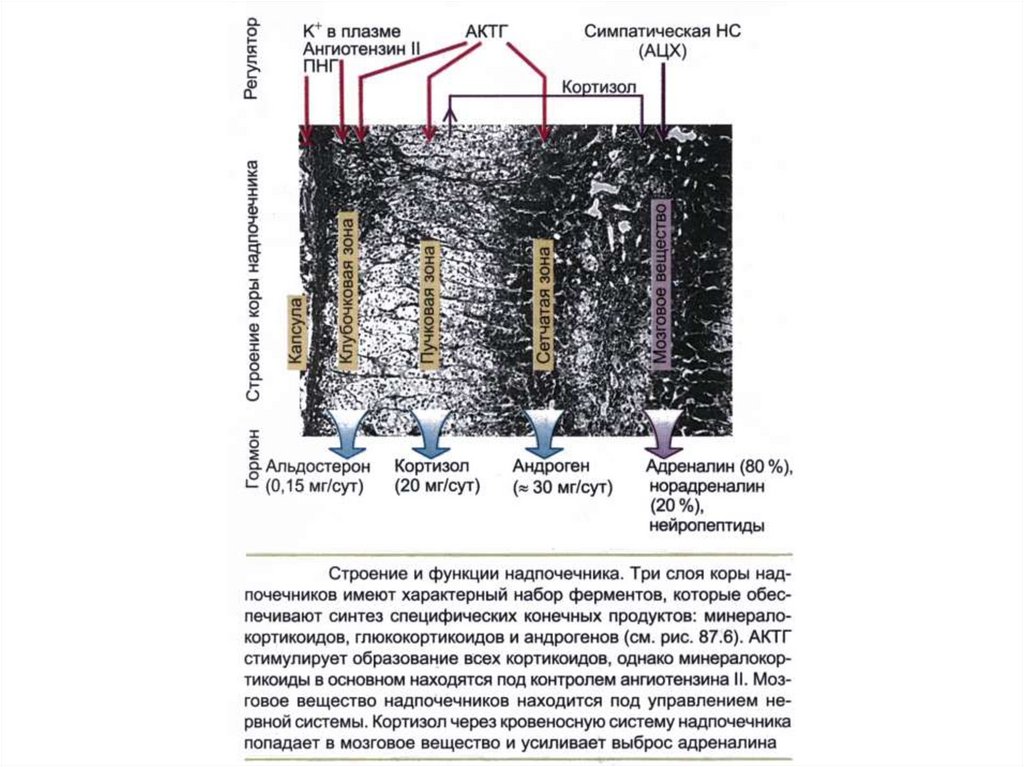
Анохин: лобные доли коры, заторможенные во сне,
высвобождают гипоталамус от своего тормозного
воздействия. Гипоталамус, активный в течение сна,
блокирует прохождение импульса через таламус, т.е.
возникает функциональная деафферентация коры.
19.
20.
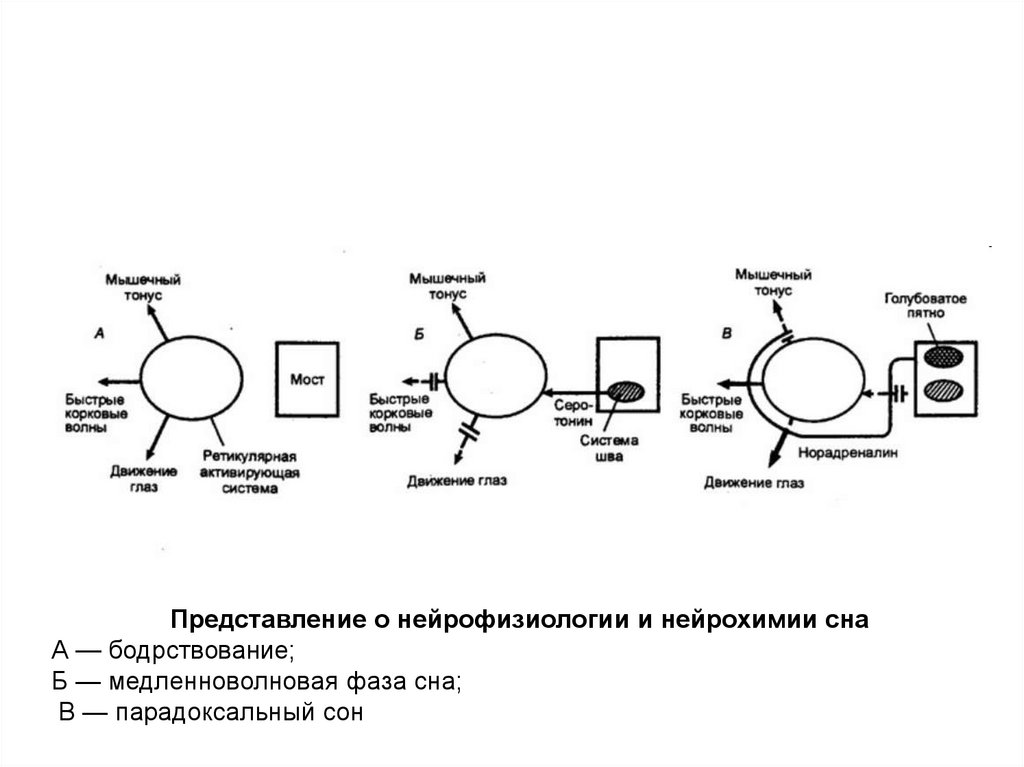
Представление о нейрофизиологии и нейрохимии снаА — бодрствование;
Б — медленноволновая фаза сна;
В — парадоксальный сон
21.
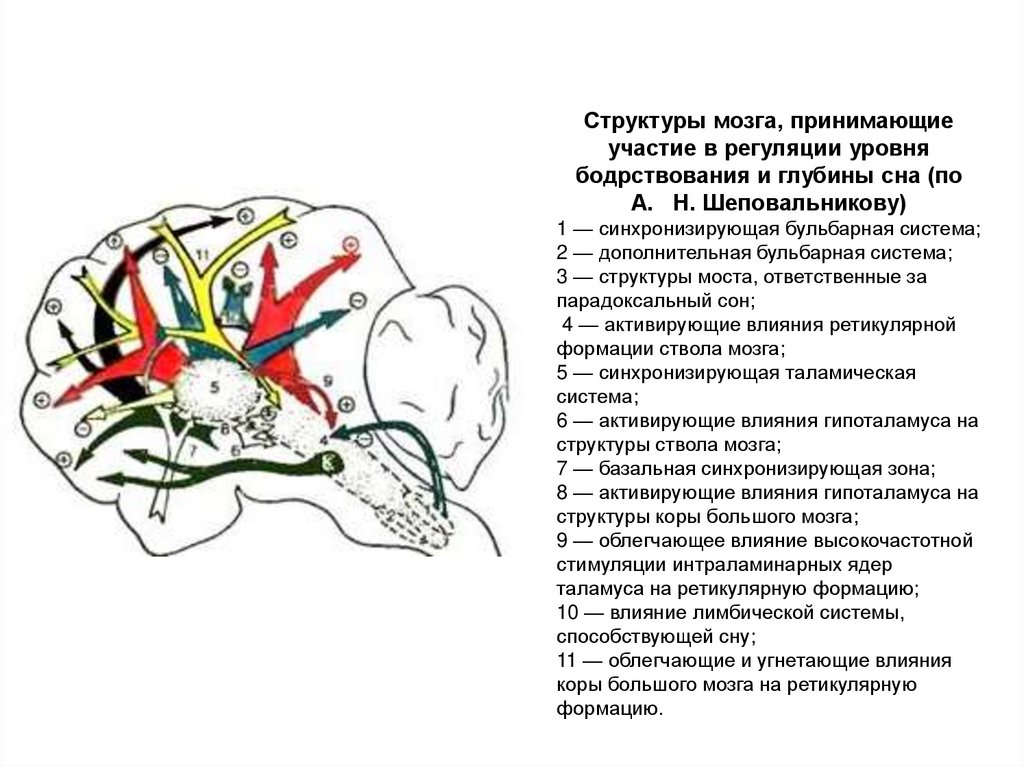
Структуры мозга, принимающиеучастие в регуляции уровня
бодрствования и глубины сна (по
А. Н. Шеповальникову)
1 — синхронизирующая бульбарная система;
2 — дополнительная бульбарная система;
3 — структуры моста, ответственные за
парадоксальный сон;
4 — активирующие влияния ретикулярной
формации ствола мозга;
5 — синхронизирующая таламическая
система;
6 — активирующие влияния гипоталамуса на
структуры ствола мозга;
7 — базальная синхронизирующая зона;
8 — активирующие влияния гипоталамуса на
структуры коры большого мозга;
9 — облегчающее влияние высокочастотной
стимуляции интраламинарных ядер
таламуса на ретикулярную формацию;
10 — влияние лимбической системы,
способствующей сну;
11 — облегчающие и угнетающие влияния
коры большого мозга на ретикулярную
формацию.
22.
Физиологияэндокринной
системы
23.
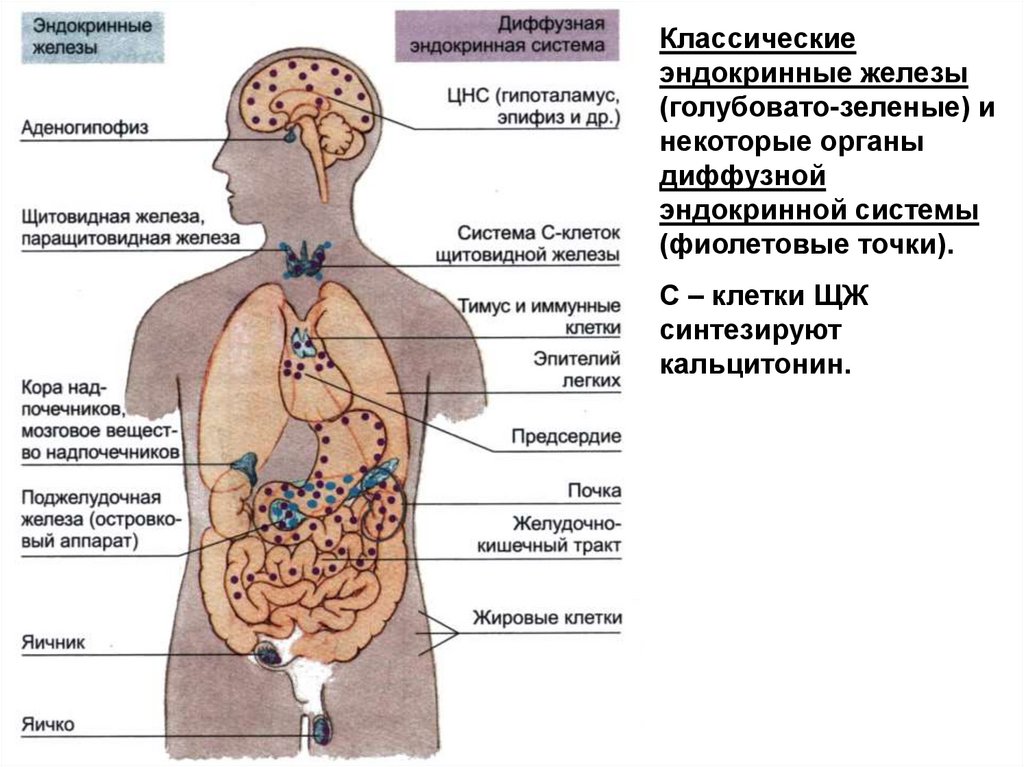
Классическиеэндокринные железы
(голубовато-зеленые) и
некоторые органы
диффузной
эндокринной системы
(фиолетовые точки).
С – клетки ЩЖ
синтезируют
кальцитонин.
24. Эндокринная система
• 1. Эндокринные железыГИПОФИЗ (аденогипофиз и нейрогипофиз)
НАДПОЧЕЧНИКИ (кора и мозговое в-во)
ЩИТОВИДНАЯ ЖЕЛЕЗА
ОКОЛОЩИТОВИДНЫЕ ЖЕЛЕЗЫ
ЭПИФИЗ
• 2. Органы с эндокринной тканью
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
ПОЛОВЫЕ ЖЕЛЕЗЫ
• 3. Органы с эндокринной функцией клеток
ПЛАЦЕНТА
ТИМУС
ПОЧКИ
СЕРДЦЕ
АПУД-система ЖКТ
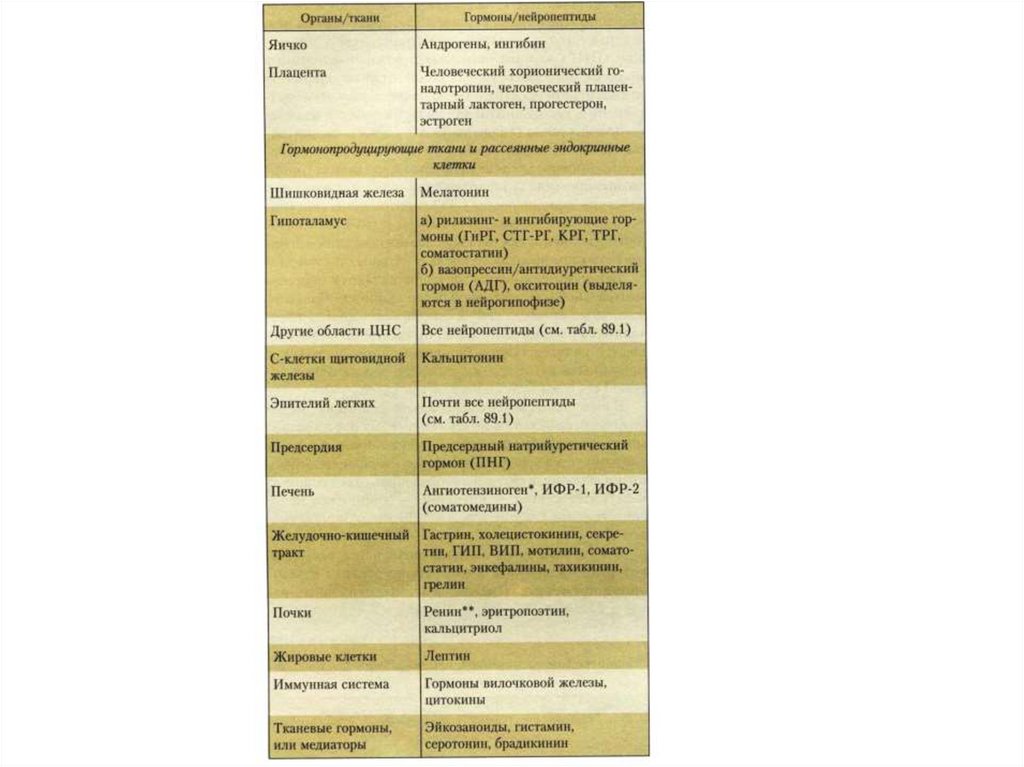
25. Железы внутренней секреции и их гормоны
26.
27.
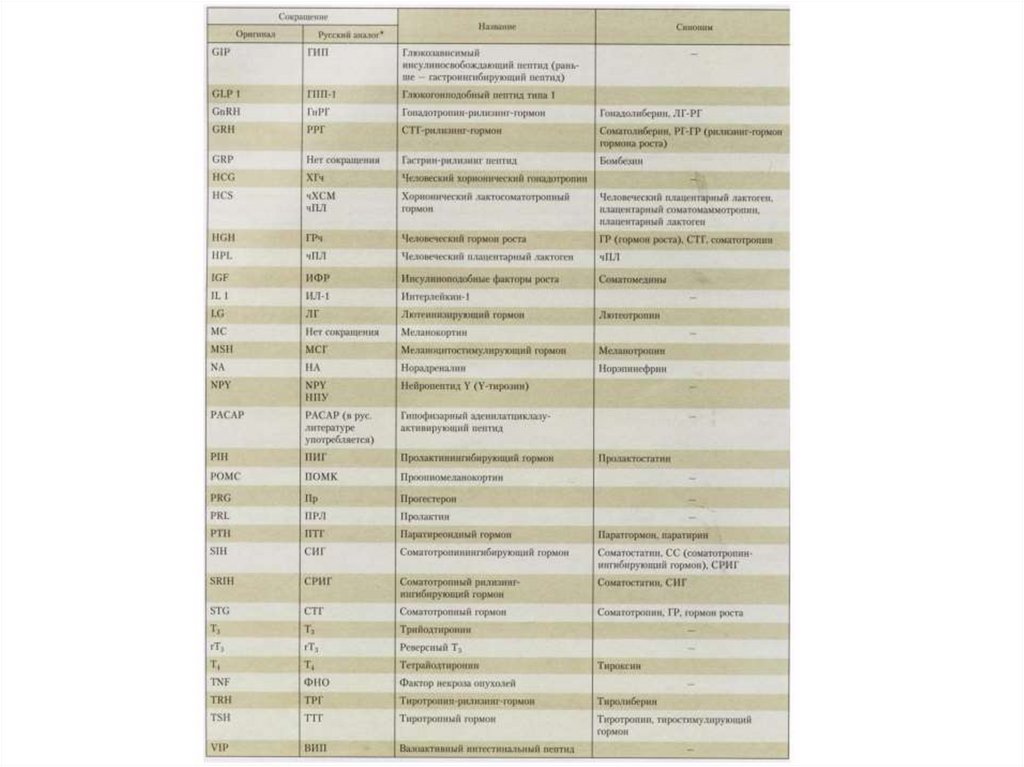
28. Химическая природа и пути действия гормонов
• Простые и сложные белки,пептиды - гормоны гипофиза, pancreas и др.
• Стероидные гормоны - коры
надпочечников, половые
стероиды
• Производные аминокислот
- адреналин, гормоны щитовидной железы, эпифиза
- через мембранные
рецепторы и вторичные посредники
- проникая в клетку,
через рецепторы циплазмы и ядра
- через мембранные
рецепторы и вторичные посредники
29.
30. Виды действия гормонов
• Метаболическое• Морфогенетическое
• Кинетическое
• Корригирующее
31.
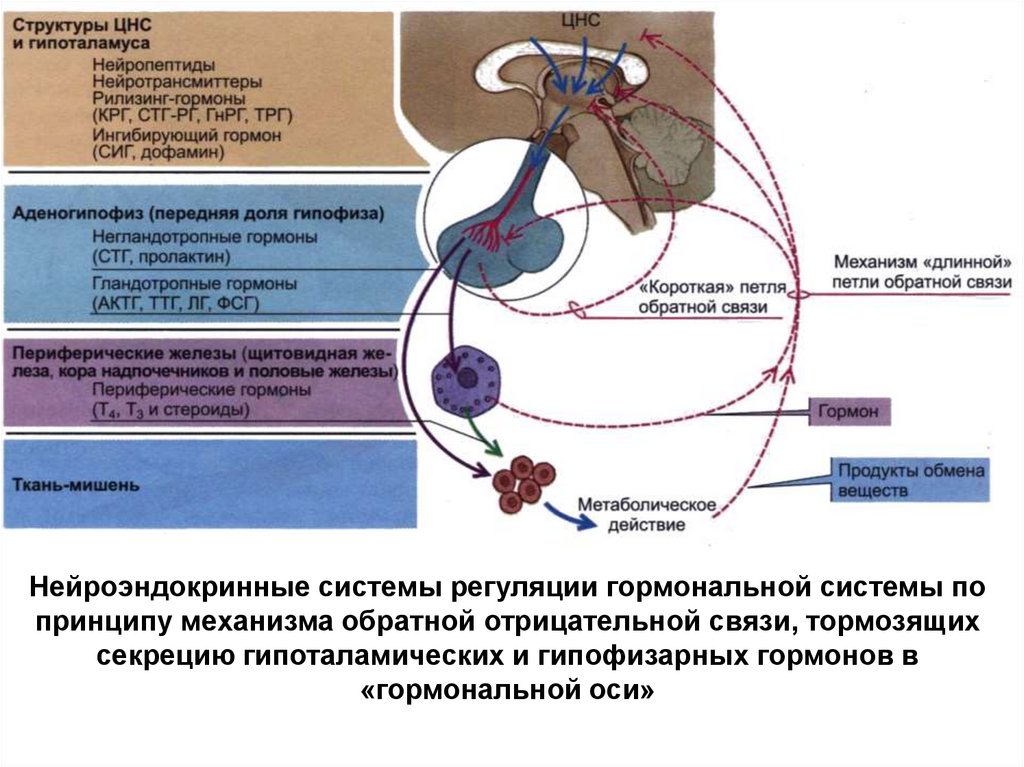
Нейроэндокринные системы регуляции гормональной системы попринципу механизма обратной отрицательной связи, тормозящих
секрецию гипоталамических и гипофизарных гормонов в
«гормональной оси»
32.
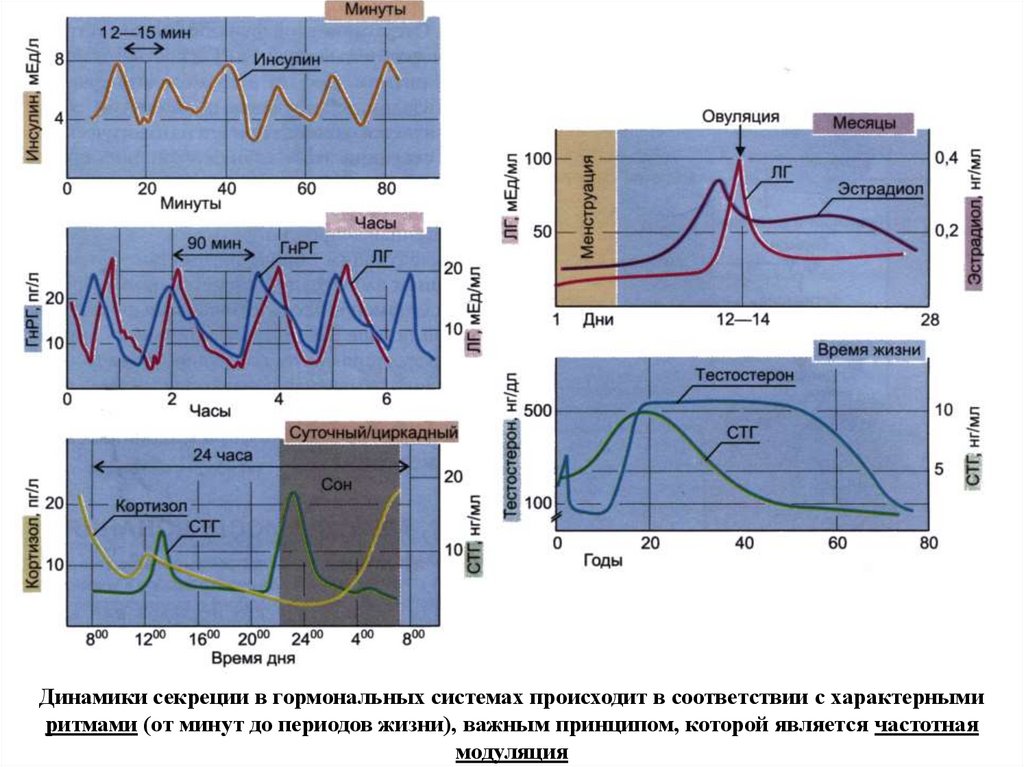
Динамики секреции в гормональных системах происходит в соответствии с характернымиритмами (от минут до периодов жизни), важным принципом, которой является частотная
модуляция
33.
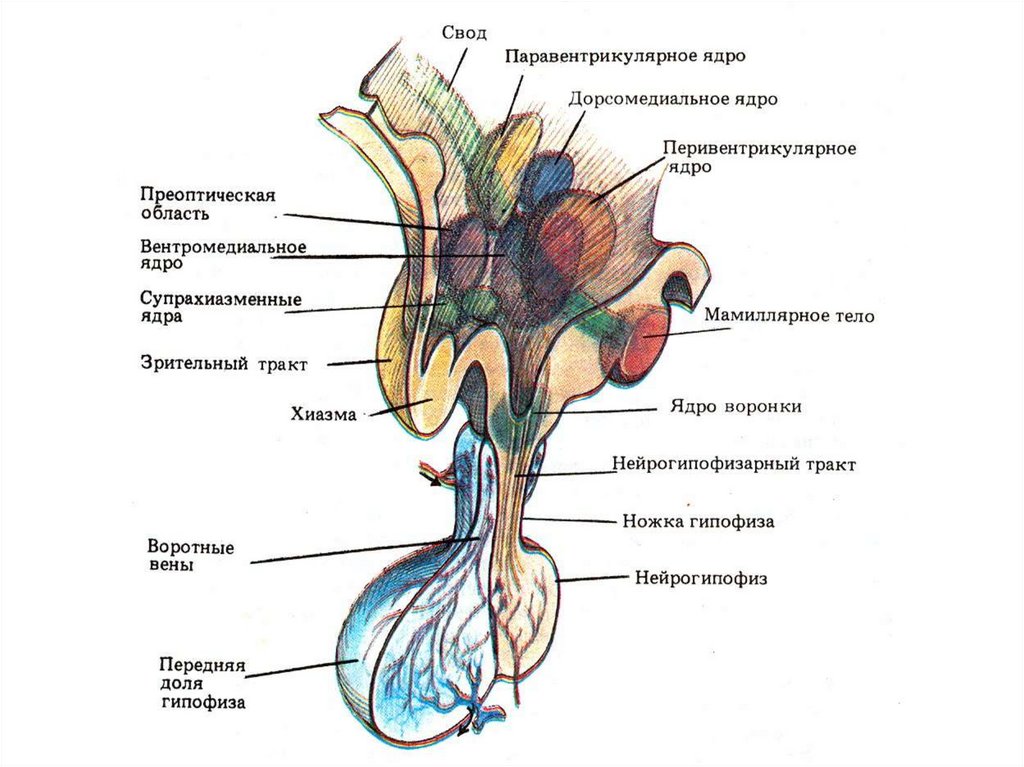
34. Нейроны гипоталамуса
Обладают рецепторной функцией и способны улавливатьизменения химического состава крови и
цереброспинальной жидкости, что достигается за счет:
1. Мощной сети капилляров (до 3000 на кв.мм.) и их
высокой проницаемости
2. Наличия клеток избирательно чувствительных к
изменению:
• рН крови
• содержанию ионов, особенно Na+ и K+
• осмотического давления (супраспинальное ядро)
• половых гормонов (передний гипоталамус)
• концентрации глюкозы (вентромедиальное ядро)
• температуры крови
«Рецепторные» нейроны гипоталамуса практически не
адаптируются и генерируют импульсы до тех пор, пока
показатель не нормализуется
35. Двусторонние связи гипоталамуса с:
• Таламусом• Лимбической системой
• КБП
• Центральным серым веществом среднего мозга
• Соматическими ядрами ствола мозга
Эти связи не только нервные, но и нервносекреторные с выделением нейропептидов:
• Опиоидов (энкефалины и эндорфины)
• Вещество Р
• Соматостатин
• Неротензин и др.
36. Гипоталамус
Вырабатывает собственные гормоны• Эффекторные: окситоцин и
вазопрессин (супраоптическое и
паравентрикулярное)
• Релизинг-гормоны: либерины и
статины
37.
38.
39.
Гормоны гипофизотропнойчасти гипоталамуса
выделяются в области
срединного возвышения и
через воротную систему
гипофиза
транспортируются в
аденогипофиз. В
нейрогипофизе
заканчиваются аксоны
нейронов (зеленые концы
стрелок; в срединном
возвышении АДГ также
выделяется в воротную
сосудистую систему).
Двусторонние связи
гипоталамуса с другими
ЦНС отмечены голубыми
стрелками.
40.
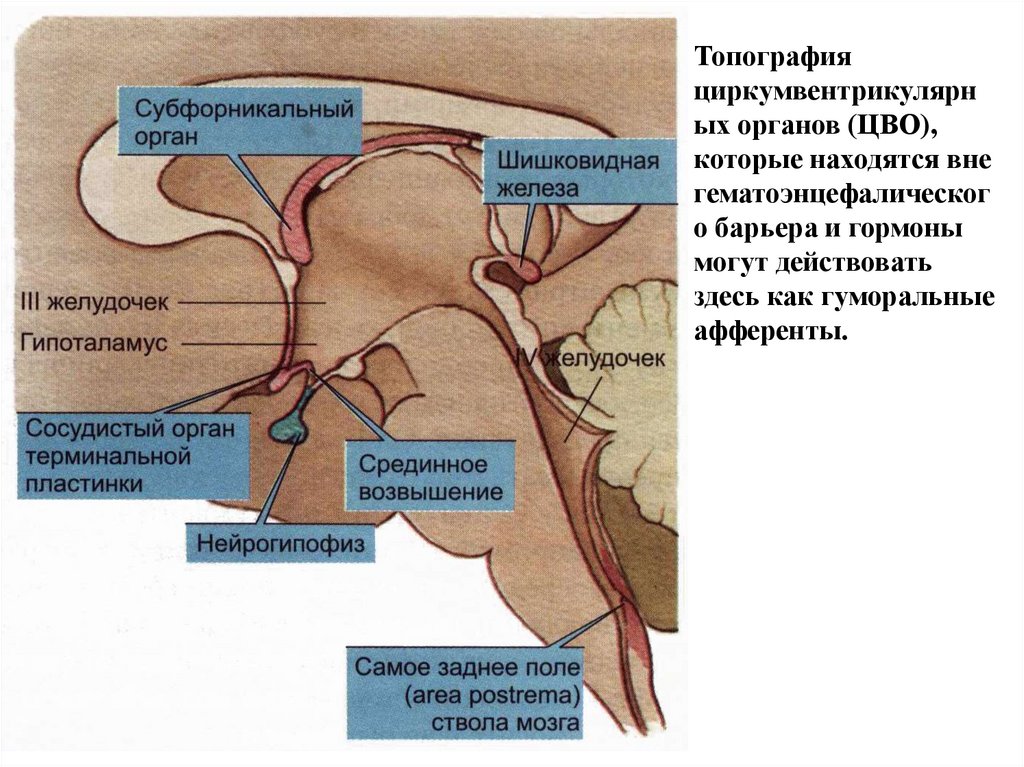
Топографияциркумвентрикулярн
ых органов (ЦВО),
которые находятся вне
гематоэнцефалическог
о барьера и гормоны
могут действовать
здесь как гуморальные
афференты.
41.
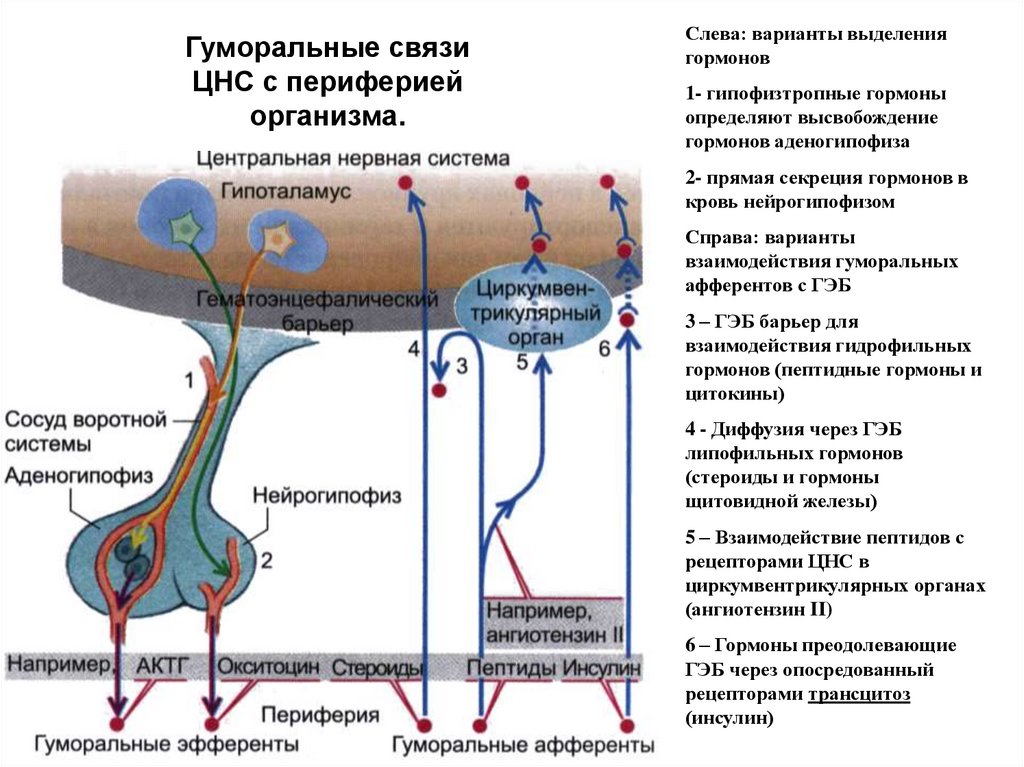
Гуморальные связиЦНС с периферией
организма.
Слева: варианты выделения
гормонов
1- гипофизтропные гормоны
определяют высвобождение
гормонов аденогипофиза
2- прямая секреция гормонов в
кровь нейрогипофизом
Справа: варианты
взаимодействия гуморальных
афферентов с ГЭБ
3 – ГЭБ барьер для
взаимодействия гидрофильных
гормонов (пептидные гормоны и
цитокины)
4 - Диффузия через ГЭБ
липофильных гормонов
(стероиды и гормоны
щитовидной железы)
5 – Взаимодействие пептидов с
рецепторами ЦНС в
циркумвентрикулярных органах
(ангиотензин II)
6 – Гормоны преодолевающие
ГЭБ через опосредованный
рецепторами трансцитоз
(инсулин)
42.
43.
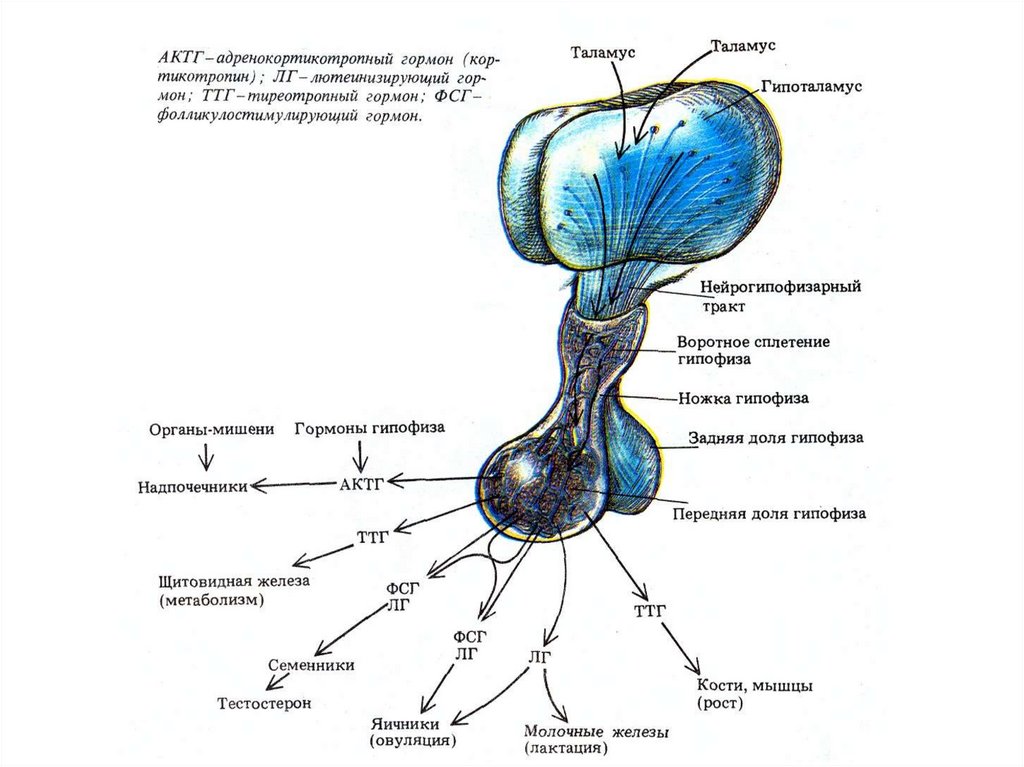
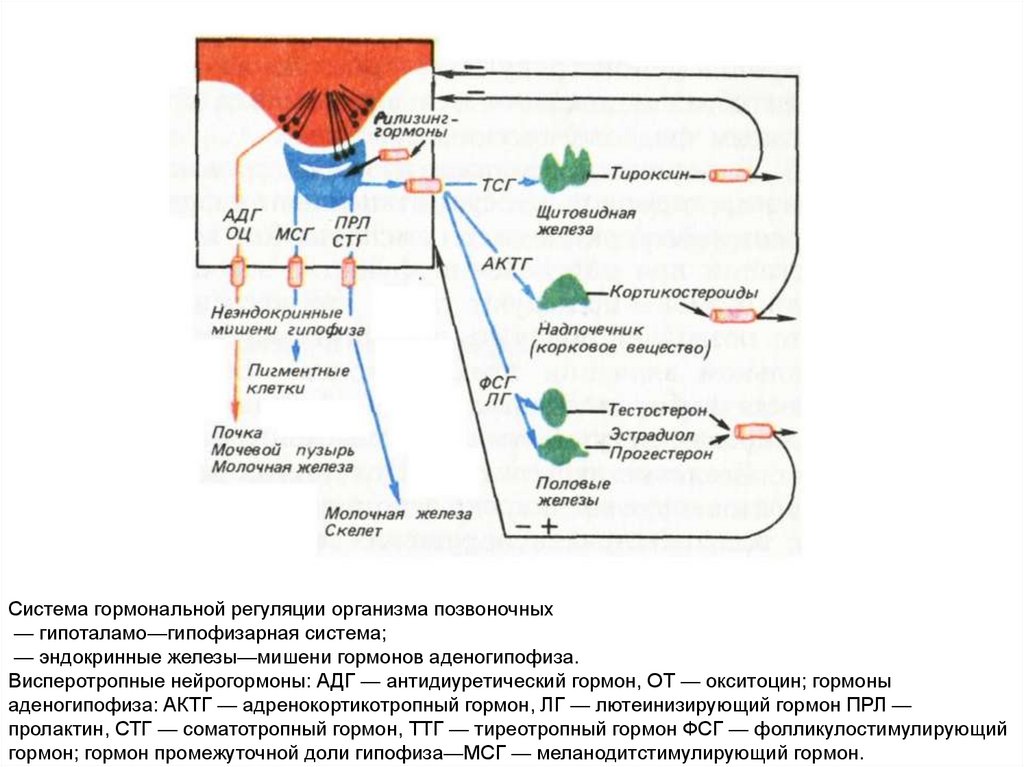
Система гормональной регуляции организма позвоночных— гипоталамо—гипофизарная система;
— эндокринные железы—мишени гормонов аденогипофиза.
Висперотропные нейрогормоны: АДГ — антидиуретический гормон, ОТ — окситоцин; гормоны
аденогипофиза: АКТГ — адренокортикотропный гормон, ЛГ — лютеинизирующий гормон ПРЛ —
пролактин, СТГ — соматотропный гормон, ТТГ — тиреотропный гормон ФСГ — фолликулостимулирующий
гормон; гормон промежуточной доли гипофиза—МСГ — меланодитстимулирующий гормон.
44.
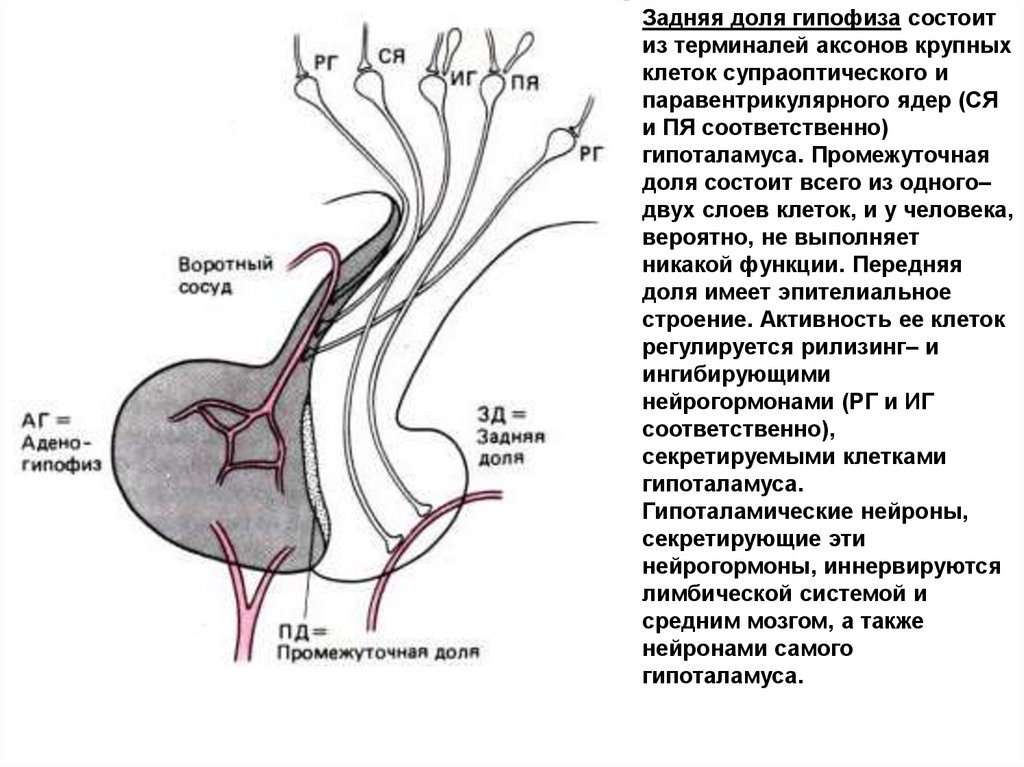
Задняя доля гипофиза состоитиз терминалей аксонов крупных
клеток супраоптического и
паравентрикулярного ядер (СЯ
и ПЯ соответственно)
гипоталамуса. Промежуточная
доля состоит всего из одного–
двух слоев клеток, и у человека,
вероятно, не выполняет
никакой функции. Передняя
доля имеет эпителиальное
строение. Активность ее клеток
регулируется рилизинг– и
ингибирующими
нейрогормонами (РГ и ИГ
соответственно),
секретируемыми клетками
гипоталамуса.
Гипоталамические нейроны,
секретирующие эти
нейрогормоны, иннервируются
лимбической системой и
средним мозгом, а также
нейронами самого
гипоталамуса.
45.
46.
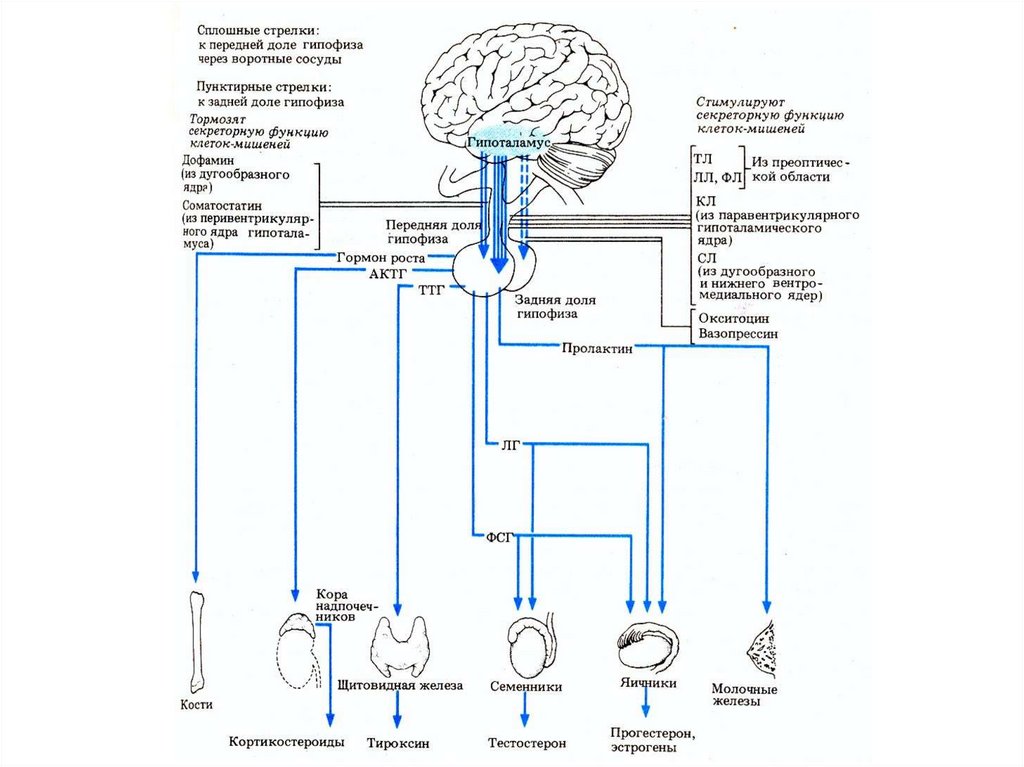
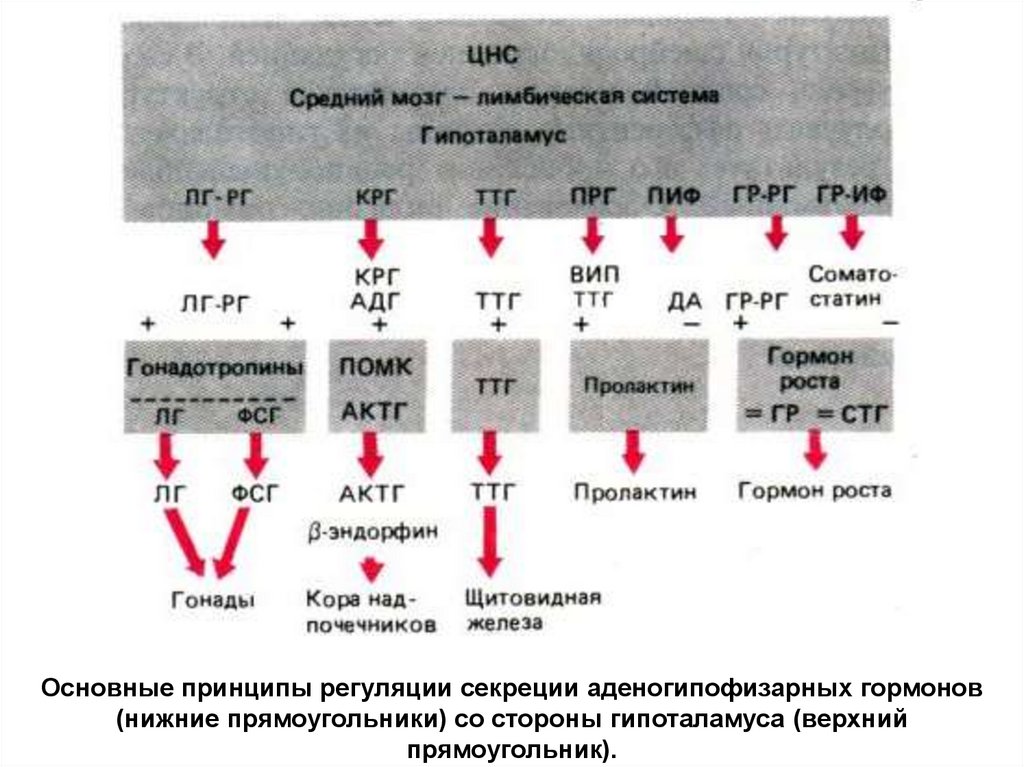
Основные принципы регуляции секреции аденогипофизарных гормонов(нижние прямоугольники) со стороны гипоталамуса (верхний
прямоугольник).
47.
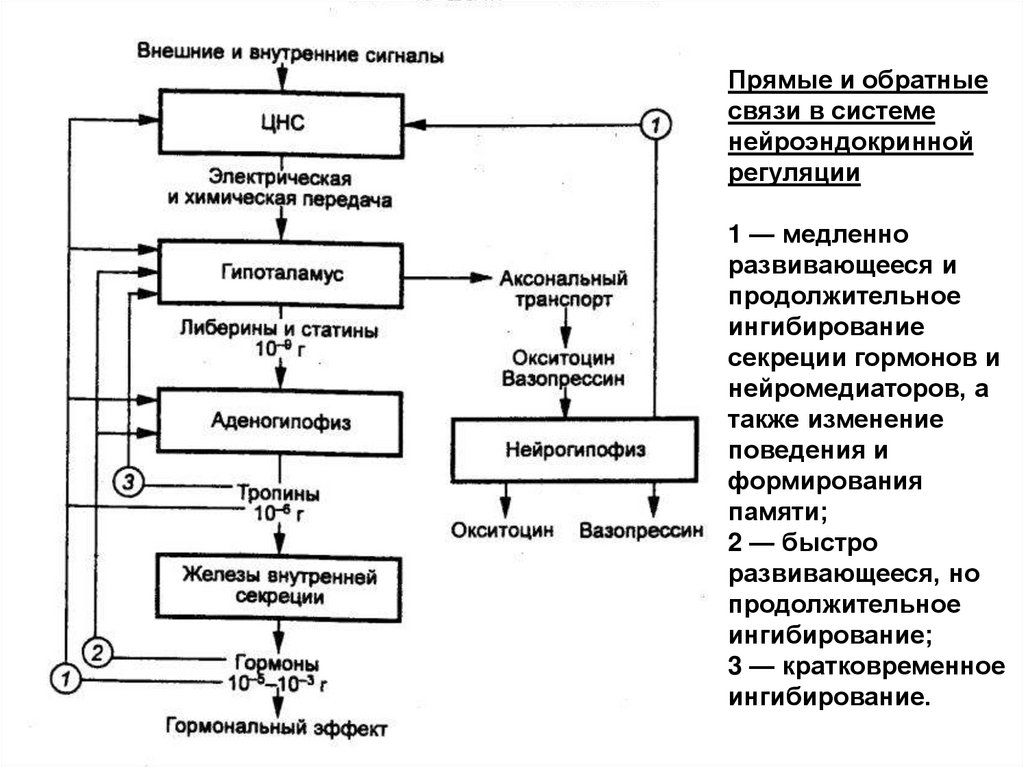
Прямые и обратныесвязи в системе
нейроэндокринной
регуляции
1 — медленно
развивающееся и
продолжительное
ингибирование
секреции гормонов и
нейромедиаторов, а
также изменение
поведения и
формирования
памяти;
2 — быстро
развивающееся, но
продолжительное
ингибирование;
3 — кратковременное
ингибирование.
48. Рилизинг–гормоны
группа нейрогормонов, мишенямикоторых являются эндокринные клетки
передней доли гипофиза.
Их подразделяют на либерины и статины.
К либеринам относятся соматолиберин,
гонадолиберин, тиреолиберин и
кортиколиберин,
К статинам - соматостатин и
пролактиностатин.
49.
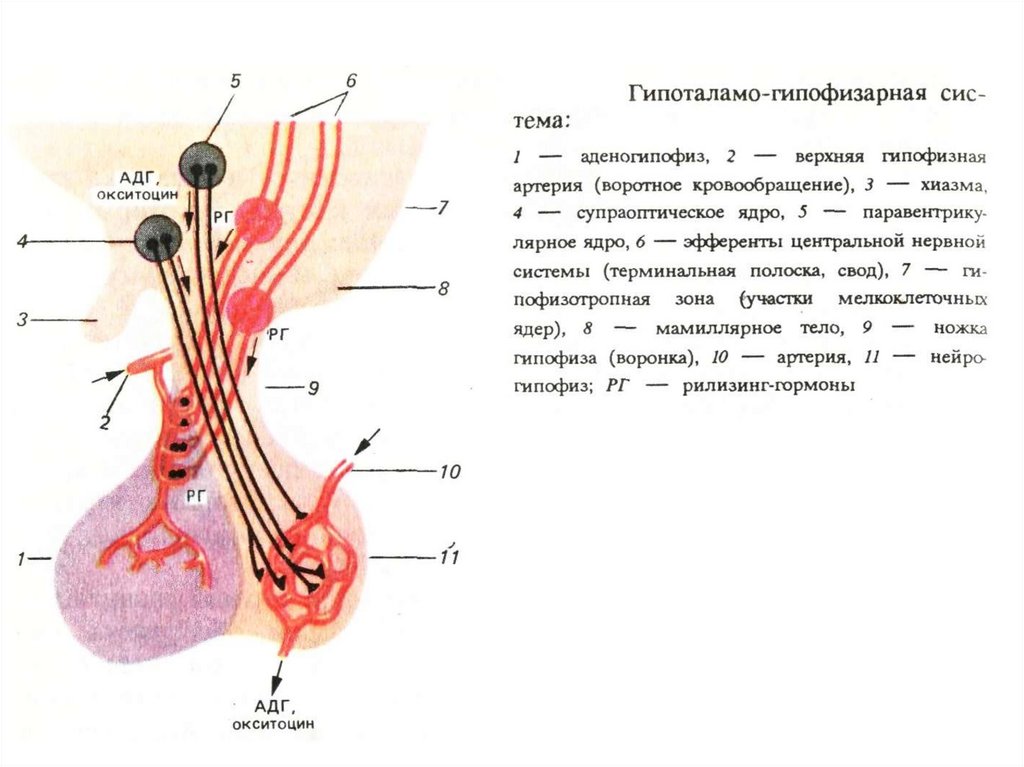
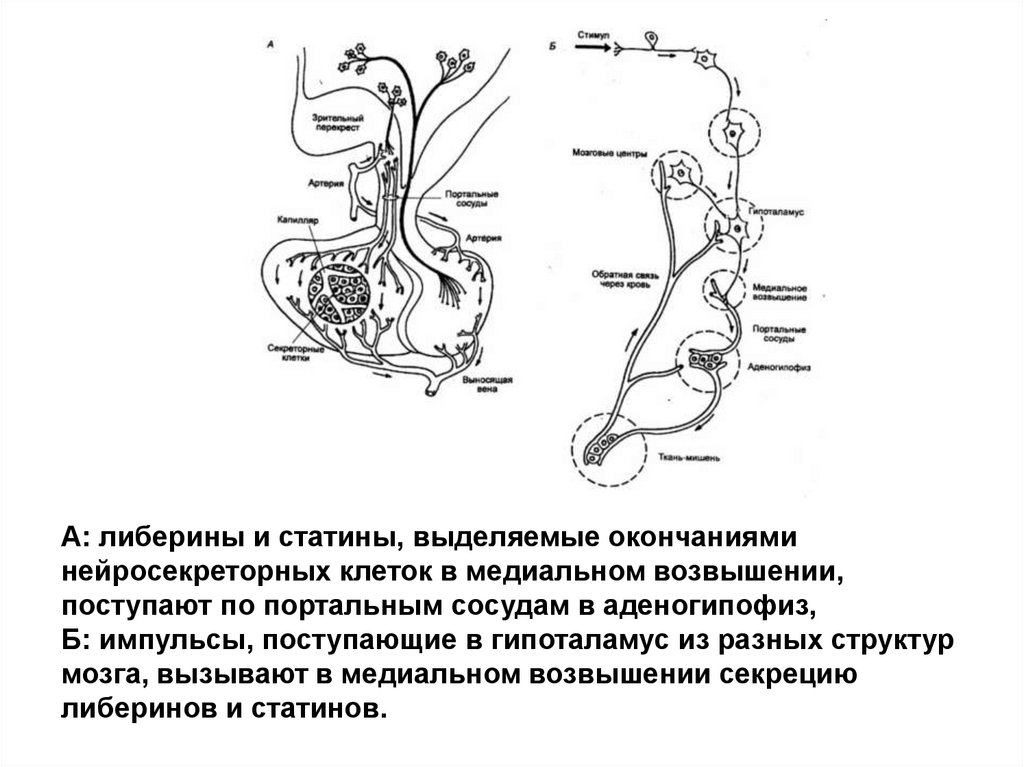
А: либерины и статины, выделяемые окончанияминейросекреторных клеток в медиальном возвышении,
поступают по портальным сосудам в аденогипофиз,
Б: импульсы, поступающие в гипоталамус из разных структур
мозга, вызывают в медиальном возвышении секрецию
либеринов и статинов.
50. Соматостатин
1 – в передней долегипофиза подавляет синтез и
секрецию СТГ, АКТГ и ТТГ;
2 – в островках Лангерганса
подавляет секрецию
инсулина и глюкагона;
3 – в ЖКТ ингибирует
секрецию гастрина,
холецистокинина, секретина;
4 – в почках подавляет
секрецию ренина;
5 – в желудке ингибирует
желудочную секрецию;
Механизм действия – через
ингибирование
аденилатциклазной системы.
51. Соматолиберин
синтезируется нейросекреторными нейронами дуговогоядра (n. arcuatus) гипоталамуса и некоторыми
опухолями островковых клеток поджелудочной
железы (соматолибриномы).
Функция - стимулирует секрецию гормона роста в
передней доле гипофиза.
Механизм действия: через связанные с G-белком
рецепторы типа секретина (стимуляция
аденилатциклазной системы).
52. Гонадолиберин
Мишени гонадолиберина — гонадотрофы переднейдоли гипофиза.
Функции:
Гонадолиберин — ключевой нейрорегулятор
репродуктивной функции, стимулирует синтез и
секрецию ФСГ и ЛГ в продуцирующих гонадотрофы
клетках,
Механизм действия: через связанные с G-белком
рецепторы (стимуляция образования
инозитолтрифосфата и диацилглицерола)
53. Пролактиностатин
Мишени пролактиностатина —лактотрофы передней доли гипофиза.
Функции:
Пролактиностатин подавляет секрецию
пролактина из лактотрофных клеток
передней доли гипофиза.
Механизм действия – через
ингибирование аденилатциклазной
системы.
54. Тиреолиберин
синтезируется многими нейронами ЦНС (в т.ч.нейросекреторными нейронами паравентрикулярного
ядра).
Мишени — тиреотрофы и лактотрофы передней доли
гипофиза:
стимулирует секрецию пролактина из лактотрофов
стимулирует секрецию тиреотропина из тиреотрофов.
стимулирует секрецию гормона роста из
соматотрофов (в патологических ситуациях).
Рецепторы. Связывание тиреолиберина с
рецепторами активирует в клетках–мишенях
G-белок, что приводит к активации фосфолипазы A2
и синтезу производных арахидоновой кислоты.
55. Кортиколиберин
синтезируется внейросекреторных
нейронах
паравентрикулярного ядра
гипоталамуса, плаценте, Тлимфоцитах.
Глюкокортикоиды
подавляют синтез
гипоталамического
кортиколиберина и
стимулируют синтез
плацентарного
кортиколиберина.
56. Кортиколиберин
Функции:1. Передняя доля гипофиза - стимуляция
синтеза и секреции АКТГ
2. Во время стресса - координатор
эндокринных, нейровегетативных и
поведенческих ответов по цепочке «кора–
гипоталамус–гипофиз–надпочечник».
Механизм действия кортиколиберина
связан с стимуляцией связанных с
G-белком рецепторов типа секретина
(стимуляция аденилатциклазной системы -
57. Меланостатин
подавляет образование меланотропинов.58. Дофамин
Промежуточный продукт метаболизматирозина и предшественник
норадреналина и адреналина —
поступающий к клеткам передней доли
гипофиза через кровь — ингибирует
секрецию ФСГ, ЛГ, ТТГ и пролактина.
59.
60. Гормоны задней доли гипофиза
Вазопрессин и окситоцин синтезируются внейросекреторных нейронах паравентрикулярного и
супраоптического ядер гипоталамуса,
транспортируются по их аксонам в составе
гипоталамо-гипофизарного пути в заднюю долю
гипофиза, где и секретируются в кровь. Сигналом к
секреции является импульсная активность этих же
самых нейросекреторных нейронов.
61. Вазопрессин (АДГ)
Оказываетантидиуретический
и
сосудосуживающий
эффекты
(повышает
системное АД).
Главная функция
АДГ — регуляция
обмена воды.
62. Регуляция секреции АДГ
Стимулируют секрецию:1. гиповолемия,
2. гиперосмоляльность,
3. переход в вертикальное положение,
4. стресс, состояние тревоги.
Подавляют секрецию :
1. алкоголь,
2. -адренергические агонисты,
3. глюкокортикоиды.
63. Окситоцин:
- стимулирует сокращениеГМК миометрия.
- стимулирует сокращение
миоэпителиальных клеток
альвеол лактирующей
молочной железы (рефлекс
молокоотделения).
Рецептор окситоцина —
связан с G-белком
(стимуляция фосфолипазыС –увеличение содержания
внутриклеточного Ca2+).
64. Гормоны передней доли гипофиза
По химической структуре гормоныаденогипофиза являются либо
пептидами, либо гликопротеинами.
Пептидные гормоны: СТГ, АКТГ и
пролактин.
Гликопротеины: ТТГ, ЛГ и ФСГ.
65. СТГ
Рецепторы СТГотносятся к
семейству
мембранных
цитокиновых
рецепторов.
Регуляция синтеза
СТГ происходит за
счет обратной связи
через АК и глюкозу.
66. Метаболические эффекты СТГ
1. Начальная фаза (инсулиноподобный эффект):А) СТГ увеличивает поглощение глюкозы мышцами и жировой
тканью,
Б) СТГ увеличивает поглощение АК и синтез белка мышцами и
печенью.
В) СТГ угнетает липолиз в жировой ткани.
2. Отсроченная фаза (диабетогенный эффект).
А) Через несколько десятков минут происходит угнетение
поглощения и утилизации глюкозы и усиление липолиза.
Б) При длительном воздействии СТГ развивается
инсулинорезистентность (мышцы, жировая ткань, печень),
которая приводит к нарушениям углеводного и жирового
обмена, и появляются симптомы сахарного диабета II типа.
67. Секрецию СТГ
стимулируют:тормозят:
1. Уменьшение глюкозы в крови,
2. Уменьшение свободных ЖК в
крови,
3. Голодание, белковая
недостаточность,
4. Травма, стресс, мышечная
работа,
5. Тестостерон, эстрогены,
6. Глубокий сон,
7. Соматолиберин.
1. Увеличение глюкозы в крови,
2. Увеличение свободных ЖК в
крови,
3. Возраст,
4. Экзогенный СТГ,
5. Ожирение,
6. Соматомедины,
7. Соматостатин.
68. Регуляция секреции АКТГ
Синтез и секрецию АКТГ стимулирует кортиколиберин.Глюкокортикоиды в высоких дозах - ингибируют
секрецию АКТГ, а в низких - стимулируют секрецию
АКТГ.
Стресс стимулирует секрецию АКТГ.
Секреция АКТГ начинает расти после засыпания и
достигает пика при пробуждении.
69. Функции АКТГ
АКТГ стимулирует синтез и секрецию гормонов корынадпочечников (преимущественно кортизола).
Гиперсекреция АКТГ ведёт к гиперплазии коры надпочечников с
увеличением секреции глюкокортикоидов и
минералокортикоидов (болезнь Иценко–Кушинга)
Дефицит АКТГ вызывает эндокринную недостаточность
надпочечников.
Рецепторы АКТГ относятся к мембранным, связанным с G-белком
(активирует аденилатциклазу, цАМФ активирует ферменты
синтеза глюкокортикоидов).
70. Секрецию пролактина
стимулируют:тиреолиберин,
стимуляция соска и
околососкового
поля.
стресс.
подавляют:
пролактиностатин
дофамин.
71. Функции пролактина
Главная функция пролактина — стимуляция функциимолочной железы.
Рецептор пролактина — мембранный цитокиновый
рецептор.
Рецептор пролактина также связывает СТГ, что объясняет
лактогенный эффект СТГ.
Гиперсекреция пролактина:
У женщин - менструальные нарушения и галакторея
(произвольные выделения из молочных желез)
У мужчин -галакторея, импотенция и снижение либидо.
У детей - задержка полового созревания.
Дефицит пролактина - послеродовая недостаточность
лактации.
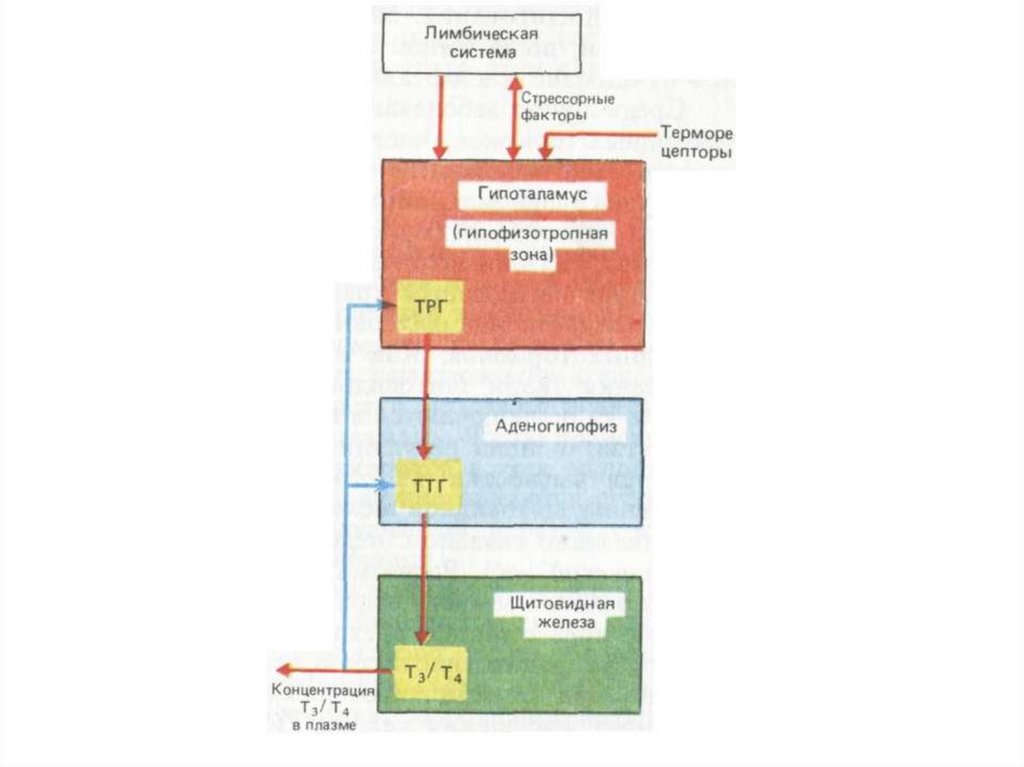
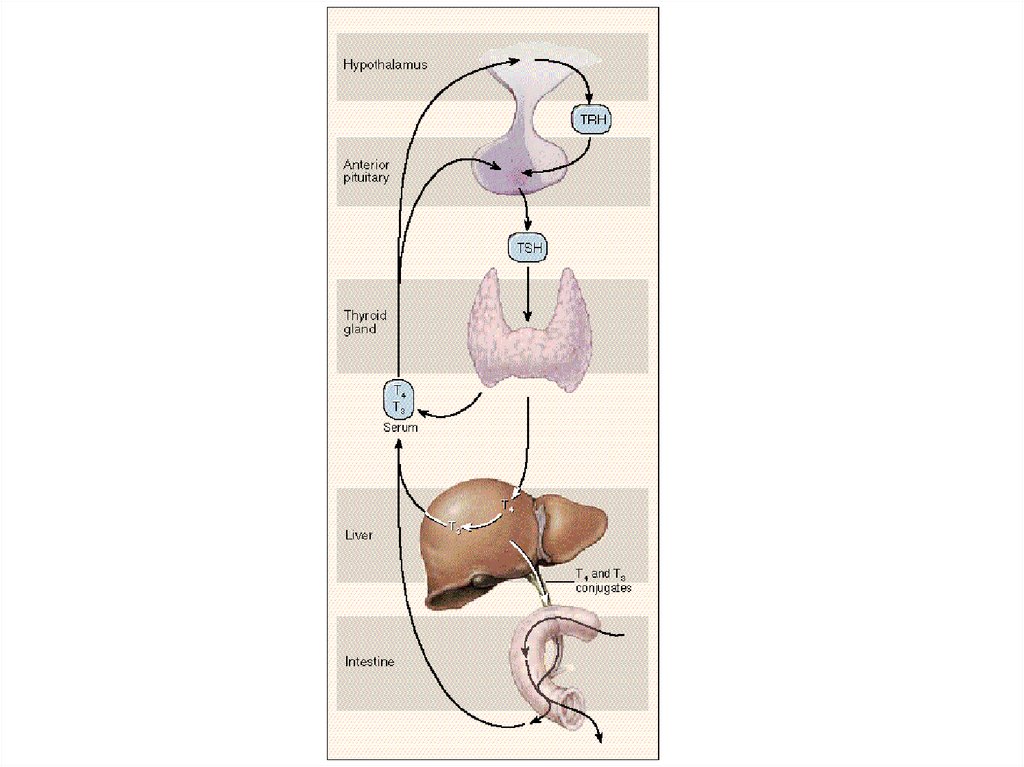
72. Тиреотропный гормон (ТТГ)
Соматостатин подавляет секрецию ТТГ.Тиреолиберин стимулирует синтез и секрецию ТТГ.
Гормоны щитовидной железы (Т3 и Т4), подавляют
секрецию ТТГ по принципу отрицательной обратной
связи.
ТТГ стимулирует дифференцировку эпителиальных
клеток щитовидной железы и их функциональное
состояние.
При дефиците ТТГ развивается вторичный гипотиреоз.
Рецептор ТТГ — трансмембранный гликопротеин,
связанный с G-белком (активация аденилатциклазы).
73. Гонадотропные гормоны:
1. фолликулостимулирующий гормон (ФСГ)2. лютеинизирующий гормон (ЛГ),
3. хорионический гонадотропин (ХГТ) плаценты.
ФСГ:
- у женщин вызывает рост фолликулов яичника,
- у мужчин регулирует сперматогенез.
ЛГ стимулирует:
- синтез тестостерона,
- синтез эстрогенов и прогестерона,
- овуляцию и образование жёлтого тела.
ХГТ синтезируется клетками трофобласта с 10–12 дней развития.
При беременности ХГТ взаимодействует с клетками жёлтого тела
и стимулирует синтез и секрецию прогестерона.
74.
Гонадолиберин стимулирует синтез и секрецию ФСГ и ЛГ.a- и b-ингибины (фолликулы яичника и клетки Сертоли яичка)
подавляют секрецию ФСГ.
Рецепторы гонадотропинов — трансмембранные
гликопротеины, связанные с G-белком (активация
аденилатциклазы).
Избыток гонадотропинов вызывает преждевременный
пубертат. Преждевременное изосексуальное половое
созревание:
девочки — менáрхе (первое менструальное кровотечение) до
8,5 лет,
мальчики — маскулинизация (накопления вторичных половых
признаков мужского пола) до 10 лет.
Недостаточность гонадотропинов приводит к развитию
гипогонадизма.
75.
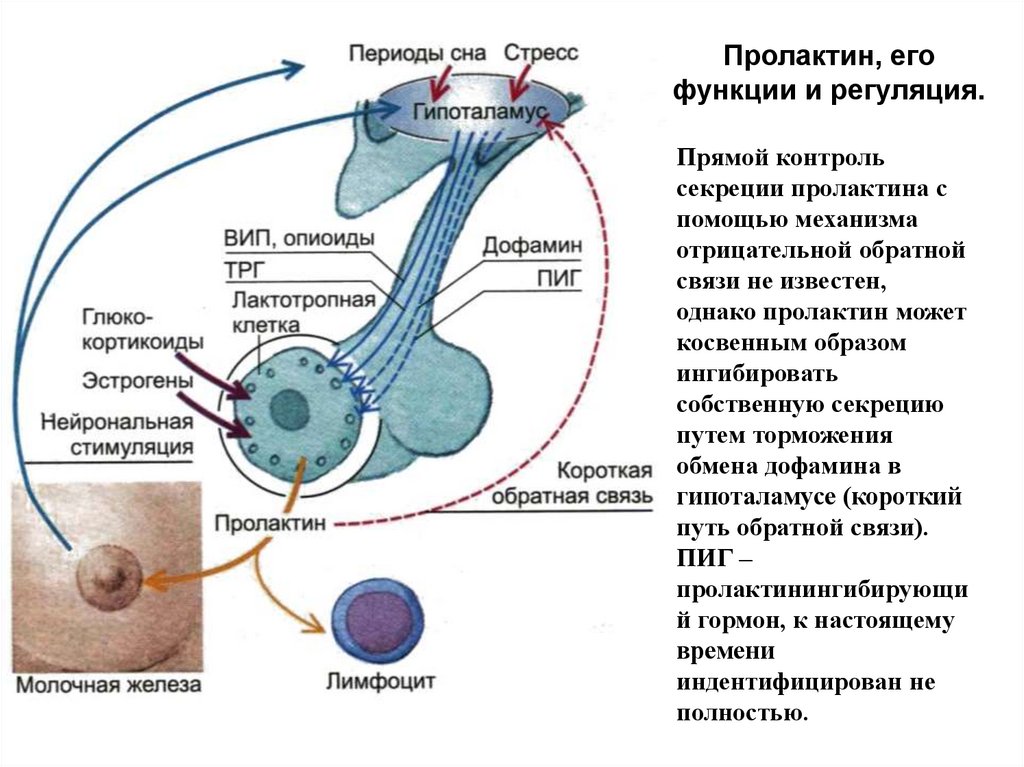
Пролактин, егофункции и регуляция.
Прямой контроль
секреции пролактина с
помощью механизма
отрицательной обратной
связи не известен,
однако пролактин может
косвенным образом
ингибировать
собственную секрецию
путем торможения
обмена дофамина в
гипоталамусе (короткий
путь обратной связи).
ПИГ –
пролактинингибирующи
й гормон, к настоящему
времени
индентифицирован не
полностью.
76.
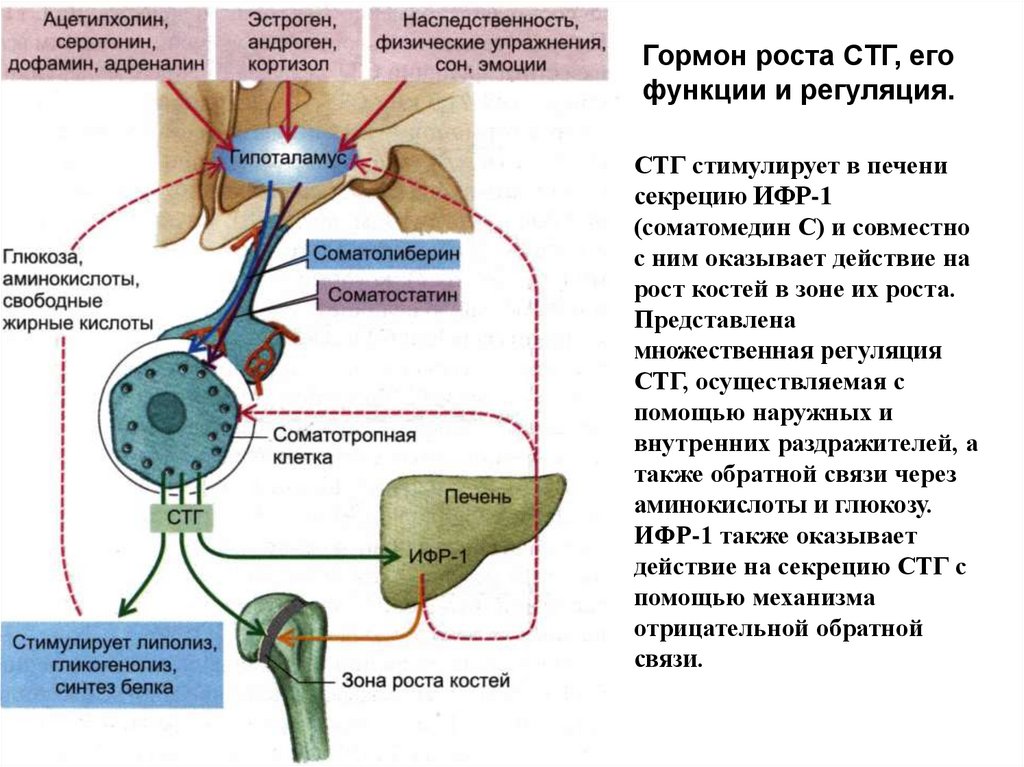
Гормон роста СТГ, егофункции и регуляция.
СТГ стимулирует в печени
секрецию ИФР-1
(соматомедин С) и совместно
с ним оказывает действие на
рост костей в зоне их роста.
Представлена
множественная регуляция
СТГ, осуществляемая с
помощью наружных и
внутренних раздражителей, а
также обратной связи через
аминокислоты и глюкозу.
ИФР-1 также оказывает
действие на секрецию СТГ с
помощью механизма
отрицательной обратной
связи.
77.
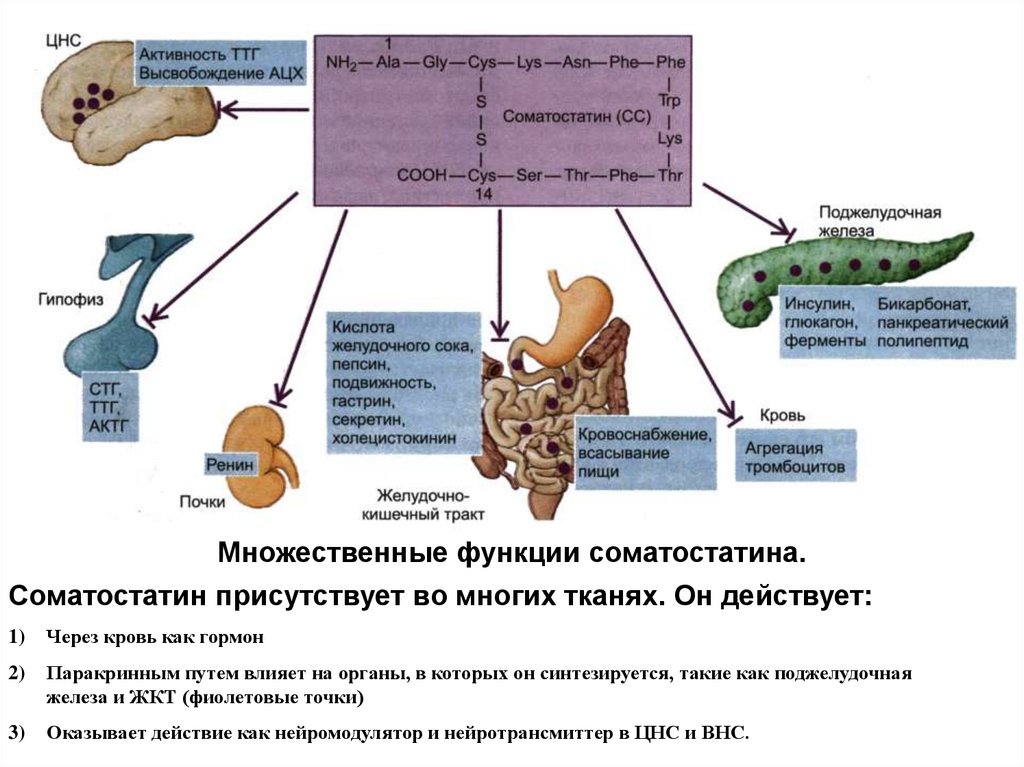
Множественные функции соматостатина.Соматостатин присутствует во многих тканях. Он действует:
1)
Через кровь как гормон
2)
Паракринным путем влияет на органы, в которых он синтезируется, такие как поджелудочная
железа и ЖКТ (фиолетовые точки)
3)
Оказывает действие как нейромодулятор и нейротрансмиттер в ЦНС и ВНС.
78.
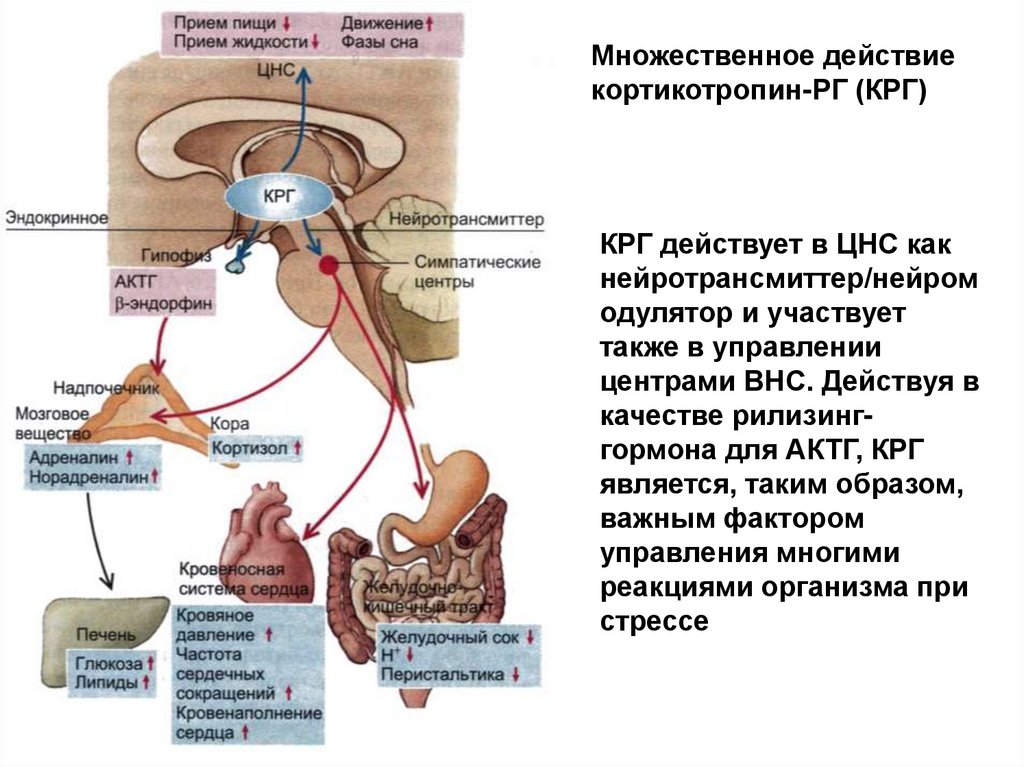
Множественное действиекортикотропин-РГ (КРГ)
КРГ действует в ЦНС как
нейротрансмиттер/нейром
одулятор и участвует
также в управлении
центрами ВНС. Действуя в
качестве рилизинггормона для АКТГ, КРГ
является, таким образом,
важным фактором
управления многими
реакциями организма при
стрессе
79.
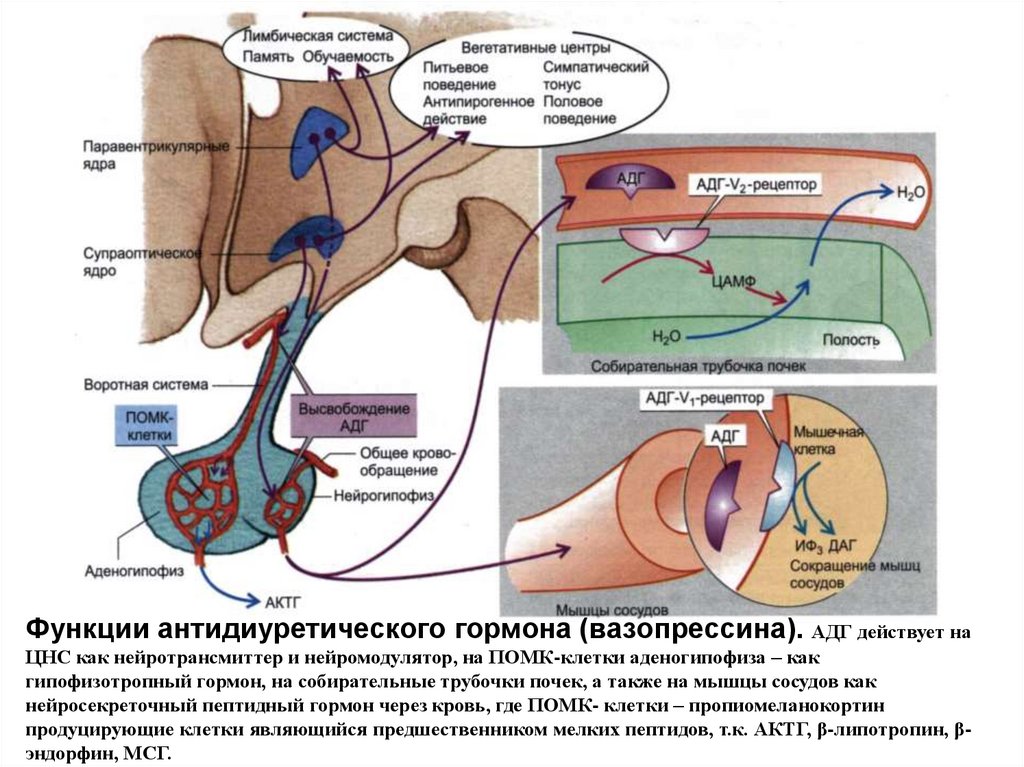
Функции антидиуретического гормона (вазопрессина). АДГ действует наЦНС как нейротрансмиттер и нейромодулятор, на ПОМК-клетки аденогипофиза – как
гипофизотропный гормон, на собирательные трубочки почек, а также на мышцы сосудов как
нейросекреточный пептидный гормон через кровь, где ПОМК- клетки – пропиомеланокортин
продуцирующие клетки являющийся предшественником мелких пептидов, т.к. АКТГ, β-липотропин, βэндорфин, МСГ.
80. Взаимодействие гипоталамуса и гипофиза
81. Гормоны гипофиза и их функции
82. Щитовидная железа
83.
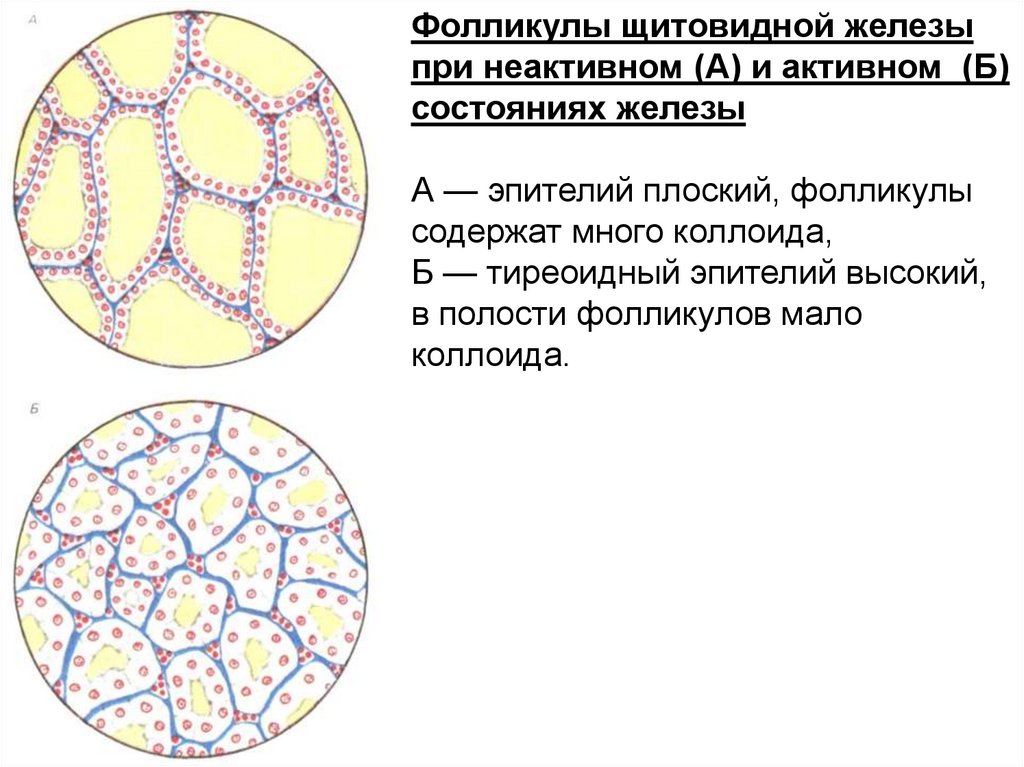
Фолликулы щитовидной железыпри неактивном (А) и активном (Б)
состояниях железы
А — эпителий плоский, фолликулы
содержат много коллоида,
Б — тиреоидный эпителий высокий,
в полости фолликулов мало
коллоида.
84.
85.
В клетках щитовидной железыпроисходит синтез двух классов
гормонов:
йодсодержащих и кальцитониновых.
1. Йодсодержащие гормоны,
2. Кальцитонин,
3. Катакальцин,
4. Пептиды a и b.
86.
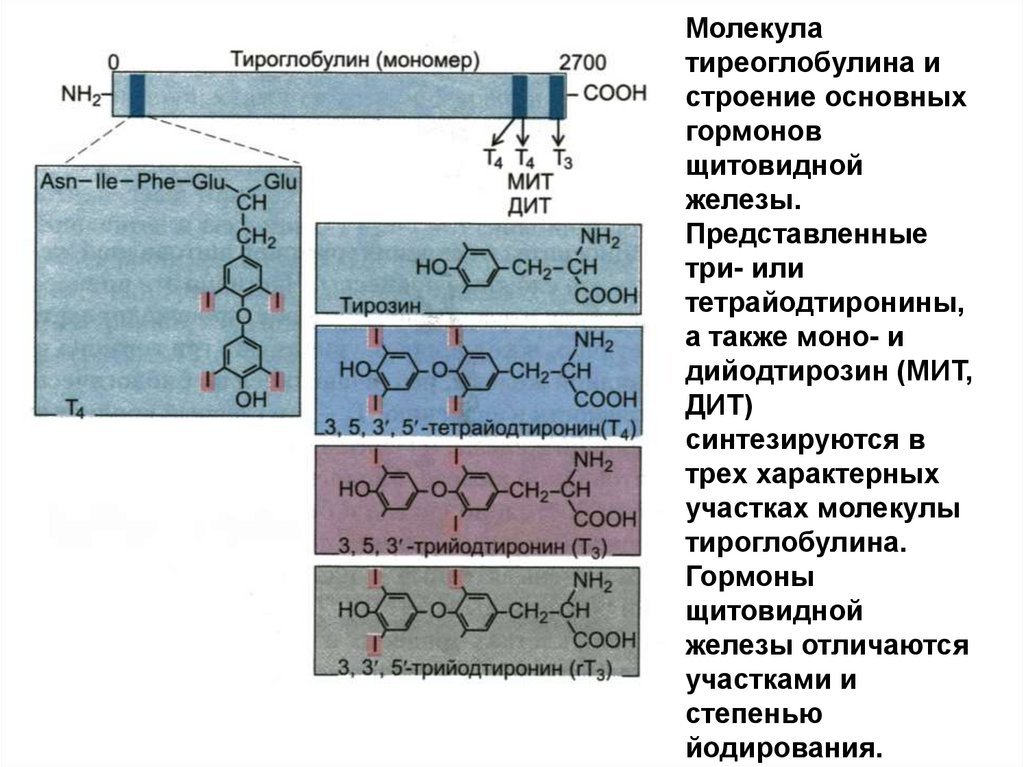
Молекулатиреоглобулина и
строение основных
гормонов
щитовидной
железы.
Представленные
три- или
тетрайодтиронины,
а также моно- и
дийодтирозин (МИТ,
ДИТ)
синтезируются в
трех характерных
участках молекулы
тироглобулина.
Гормоны
щитовидной
железы отличаются
участками и
степенью
йодирования.
87. Тироксин и трийодтиронин
Йодсодержащие гормоны - тироксин(T4) и трийодтиронин (Т3)
Образование Т3 из тироксина частично
происходит в щитовидной железе, а
преимущественно в печени.
Рецепторы тиреоидных гормонов ядерные рецепторы - факторы
транскрипции.
Сродство Т3 к рецепторам клеток–
мишеней в 10 раз выше, чем у Т4.
88. Функции йодсодержащих гормонов
1. Необходимы для нормального развития ЦНС,2. Увеличивают обменные процессы, ускоряют катаболизм
белков, жиров и углеводов,
3. Увеличивают ЧСС и сердечный выброс,
4. Увеличивают потребление О2 в результате увеличения
активности Na+,K+-АТФазы.
5. В печени ускоряют гликолиз, синтез холестерола и желчных
кислот.
6. Т3 в печени и жировой ткани повышает чувствительность клеток
к адреналину (стимуляция липолиза в жировой ткани и
мобилизация гликогена в печени).
7. Т3 увеличивает потребление глюкозы, стимулирует синтез
белков и увеличение мышечной массы, повышает
чувствительность к действию адреналина.
8. Участвуют в теплопродукции, повышая чувствительность к
симпатической нервной системе и стимулируя секрецию
норадреналина.
89. Кальцитонин и катакальцин
Их функции антагонистичны эффектамПТГ:
кальцитонин уменьшает [Са2+] в крови,
стимулирует минерализацию кости,
усиливает почечную экскрецию Са2+,
фосфатов и Na+ (уменьшает их
реабсорбцию).
90. Пептиды a и b
относящиеся к кальцитониновому гену,участвуют в ноцицепции, пищевом
поведении, а также в регуляции тонуса
сосудов.
Рецепторы к этим пептидам найдены в
ЦНС, сердце, плаценте.
91.
Уровень гормонов щитовиднойжелезы и связывающих белков
в плазме и функциональные
тесты.
А) приведены характерные для различных
функциональных состояний щитовидной
железы доли свободного, несвязанного Т4 и
свободных и занятых мест связывания
тироксинсвязывающего глобулина (ТСГ), а
также соответствующие значения ТТГ в
крови. Очевидно, что для гормональной
активности ЩЖ очень важным яв-ся
наличие свободного гормона.
Б) представлен стимулирующий тест с
ТРГ. Для гипотиреоза характерен высокий
уровень ТТГ в сочетании с повышенной
стимулирующей способностью ТРГ. При
гипертиреозе секреция ТТГ не может
стимулироваться с помощью ТРГ.
92.
Связи щитовиднойжелезы у млекопитающих
93.
94.
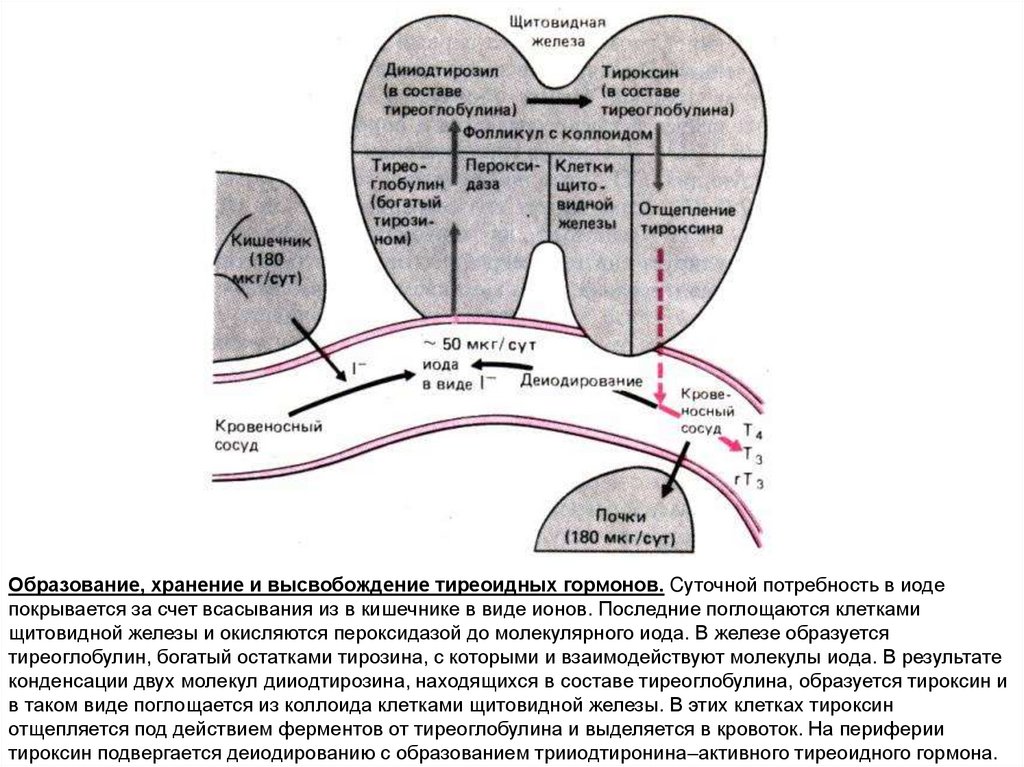
Образование, хранение и высвобождение тиреоидных гормонов. Cуточной потребность в иодепокрывается за счет всасывания из в кишечнике в виде ионов. Последние поглощаются клетками
щитовидной железы и окисляются пероксидазой до молекулярного иода. В железе образуется
тиреоглобулин, богатый остатками тирозина, с которыми и взаимодействуют молекулы иода. В результате
конденсации двух молекул дииодтирозина, находящихся в составе тиреоглобулина, образуется тироксин и
в таком виде поглощается из коллоида клетками щитовидной железы. В этих клетках тироксин
отщепляется под действием ферментов от тиреоглобулина и выделяется в кровоток. На периферии
тироксин подвергается деиодированию с образованием трииодтиронина–активного тиреоидного гормона.
95.
96.
97.
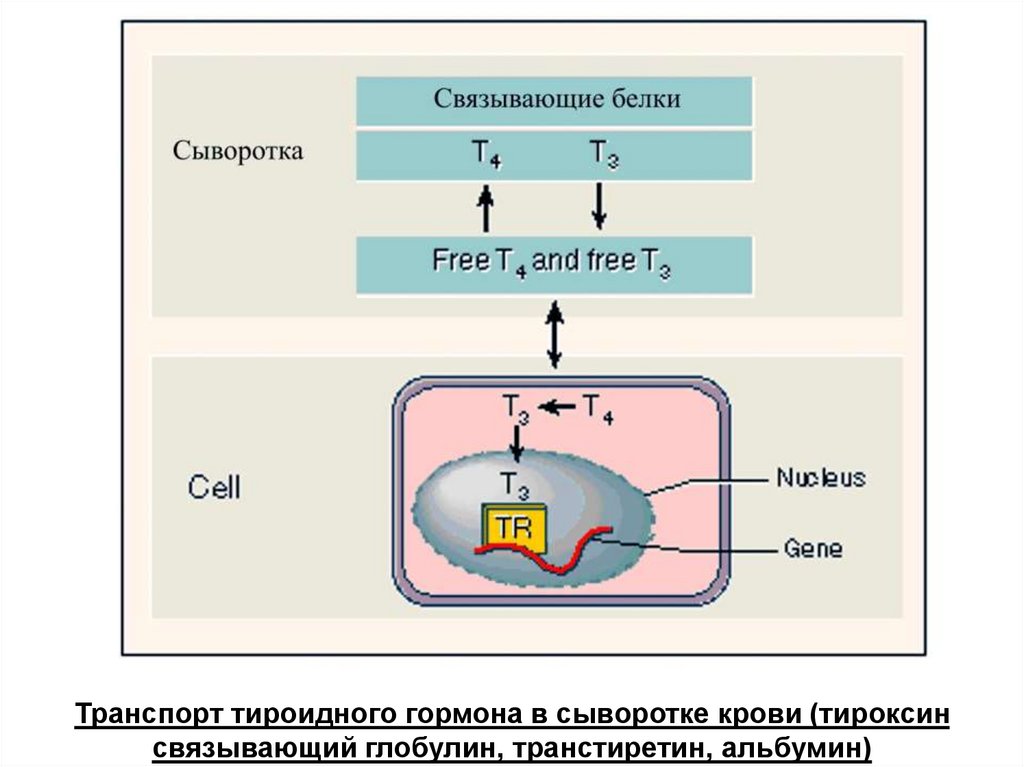
Транспорт тироидного гормона в сыворотке крови (тироксинсвязывающий глобулин, транстиретин, альбумин)
98.
Гормоны щитовидной (1) и паращитовидных (2) желез и их функции99.
• Cуточная потребность в йоде покрывается за счет всасывания вкишечнике в виде ионов.
• Последние поглощаются клетками щитовидной железы и
окисляются пероксидазой до молекулярного йода.
• В железе образуется тиреоглобулин, богатый остатками
тирозина, с которыми и взаимодействуют молекулы йода.
• В результате конденсации двух молекул дийодтирозина,
находящихся в составе тиреоглобулина, образуется тироксин и
в таком виде поглощается из коллоида клетками щитовидной
железы.
• В этих клетках тироксин отщепляется под действием
ферментов от тиреоглобулина и выделяется в кровоток.
• На периферии тироксин подвергается дейодированию с
образованием трийодтиронина–активного тиреоидного гормона.
100.
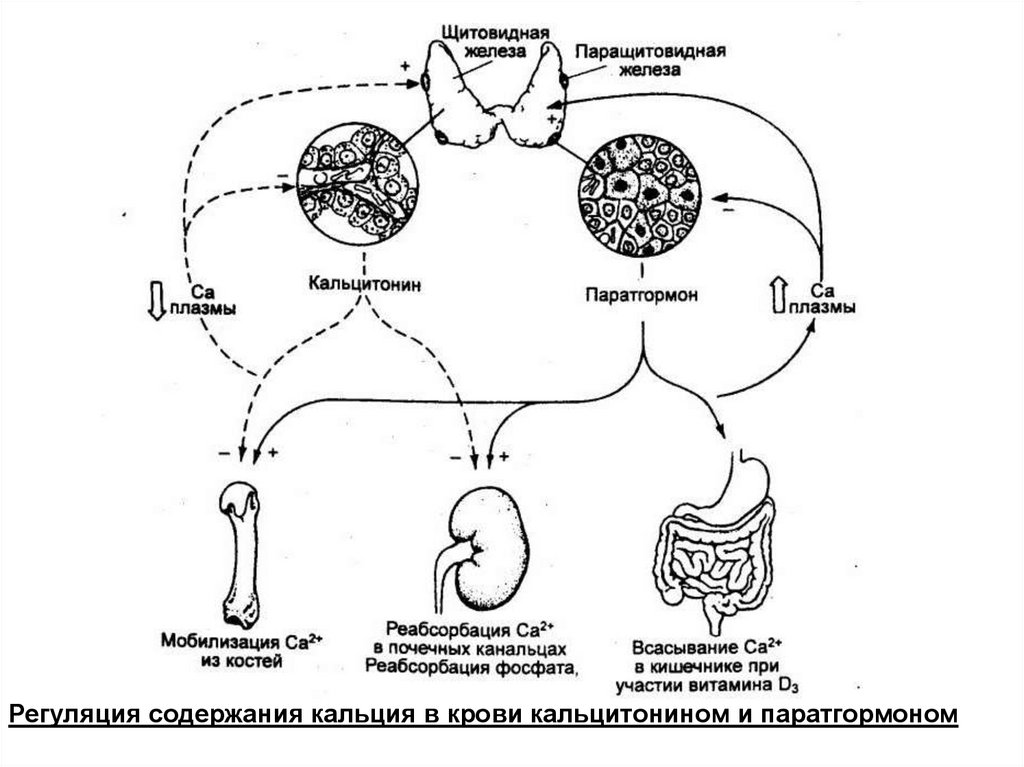
Регуляция содержания кальция в крови кальцитонином и паратгормоном101.
102. Эпифиз
103. Эпифиз
Гормоны эпифиза представлены в основном индоламинами (серотонин имелатонин), пептидами и белками.
Выделяясь в ликвор III желудочка, серотонин:
1.
действует на гипоталамо—гипофизарную систему в условиях стресса, где
увеличивает секрецию соматотропина, кортикотропина и пролактина,
запускающих защитные реакции организма, и подавляет выход в кровь
гонадотропинов и тиреотропина.
2.
увеличивает секрецию тормозного медиатора ГАМК, что оказывает
сомногенный эффект.
3.
непосредственно влияет на активацию иммунной системы, облегчая течение
воспалительных реакций.
4.
может соединяться с гемоглобином, препятствуя диссоциации его молекулы
под влиянием ультрафиолета.
Группа пептидов и белков эпифиза регулирует репродуктивные функции
посредством контроля секреции гонадотропинов гипофиза и половых
стероидов гонад. Гонадотропин, гонадолиберин и антигонадотропины
эпифиза оказывают на репродуктивную систему реципрокное действие
104. Гормон эпифиза — мелатонин
Кроме того:1.
снижение метаболизма в
светочувствительных
нейронах СХЯ,
иннервирующих и
активирующих все центры
нейрогипоталамуса.
2.
подавляет секрецию
гонадолиберина и окситоцина
и активирует секрецию
соматостатина
3.
обеспечивает регуляцию
биоритмов эндокринных
функций и метаболизма для
приспособления организма к
разным условиям
освещенности
4.
в радужной оболочке глаза
повышает
светочувствительность
фоторецепторов
105. НАДПОЧЕЧНИКИ
106.
Схема зон надпочечника и вырабатываемые ими гормоны1 — пучковая зона, 2 — сетчатая зона, 3 — корковое вещество, 4 —
мозговое вещество, 5 — клубочковая зона.
107. Строение и функции надпочечника
Три слоя надпочечников имеютхарактерный набор
ферментов обеспечивающий
синтез специфических
конечных продуктов:
глюкокортикоидов,
минералокортикоидов и
андрогенов.
Мозговое вещество синтезирует
катехоламины.
108.
109. ГОРМОНЫ НАДПОЧЕЧНИКА
Кора надпочечникаМозговое вещество
АДРЕНАЛИН
НОРАДРЕНАЛИН
КОРТИЗОЛ
КОРТИКОСТЕРОН
АЛЬДОСТЕРОН
ДЕЗОКСИКОРТИКОСТЕРОН
ДЕГИДРОЭПИАНДРОСТЕРОН
АНДРОСТЕНДИОН
110. Глюкокортикоиды
Основной глюкокортикоид – кортизолРегуляция секреции
Активирующие (нисходящие) влияния.
Непосредственный активатор синтеза и
секреции кортизола — АКТГ. АКТГ выделяется
клетками передней доли гипофиза под
действием кортиколиберина.
Стрессовые стимулы активируют всю нисходящую
систему влияний, вызывая быстрое выделение
кортизола. Кортизол вызывает различные
метаболические эффекты, направленные на
снятие повреждающей природы состояния
стресса.
Тормозные (восходящие) влияния по принципу
отрицательной обратной связи реализует
кортизол, подавляя секрецию АКТГ в передней
доле гипофиза и кортиколиберина в
гипоталамусе. Это уменьшает концентрацию
кортизола в плазме в то время, когда организм
не подвергается воздействию стресса.
111. Функции глюкокортикоидов
Углеводный обмен.1. Стимуляция глюконеогенеза и уменьшение
потребления глюкозы внутренними органами (кроме
головного мозга). Основной эффект — увеличение
концентрации глюкозы в крови.
2. Глюконеогенез за счёт аминокислот, лактата и
жирных кислот.
3. В печени стимулируют синтез ключевых ферментов
обмена аминокислот — субстратов глюконеогенеза.
4. Синтез гликогена усиливается за счёт активации
гликогенсинтетазы. Запасаемый гликоген легко
превращается в глюкозу путём гликогенолиза.
112. Функции глюкокортикоидов
Липидный обмен.1. Липолиз усиливается в конечностях.
2. Липогенез усиливается в других частях тела
(туловище и лицо) (синдром Кушинга).
Белки и нуклеиновые кислоты
1. Анаболический эффект в печени.
2. Катаболический эффект в других органах, в
особенности в скелетных мышцах, что приводит к
их атрофии и мышечной слабости. Образующиеся
аминокислоты поступают в печень.
113. Функции глюкокортикоидов
- В высоких дозах выступают как иммунодепрессанты(применяют для предупреждения отторжения
трансплантатов).
- имеют выраженный противовоспалительный эффект.
- при длительном применении ингибируют
синтетическую активность фибробластов и
остеобластов, в результате развиваются истончение
кожи и остеопороз.
- введение глюкокортикоидов может уменьшить отёк
слизистой оболочки.
114. Основной минералокортикоид - альдостерон
Основнойминералокортикоид альдостерон
Регуляция секреции альдостерона :
Ангиотензин II — главный регулятор, стимулирует выброс
альдостерона.
Сердечный натрийуретический фактор (атриопептид)
ингибирует синтез альдостерона.
Na+ Эффекты гипо- и гипернатриемии реализуются через ренинангиотензиновую систему.
К+ Эффекты ионов калия не зависят от содержания в крови Na+ и
ангиотензина II. Гиперкалиемия стимулирует секрецию
альдостерона. Гипокалиемия тормозит секрецию альдостерона.
Простагландины E1 и E2 стимулируют синтез альдостерона. F1a
и F2a тормозят секрецию минералокортикоидов.
Травмы и стрессовые состояния увеличивают секрецию
альдостерона за счёт активирующего влияния АКТГ на кору
надпочечников.
115. Функции альдостерона:
- увеличивает реабсорбцию Na+. Задержка Na+приводит к увеличению содержания воды в
организме и повышению АД.
- увеличивает экскрецию ионов К+ вызывает
гипокалиемию, мышечную слабость.
- увеличивает реабсорбцию Cl-, HCO3- и экскрецию H+.
Гипоальдостеронизм
потеря Na+приводит к гиповолемии, снижению
сердечного выброса и кровотока в почках, слабости,
артериальной гипотензии, а задержка K+ ведёт к
гиперкалиемии и нарушениям сердечного ритма.
116. Гормоны надпочечников
117.
118.
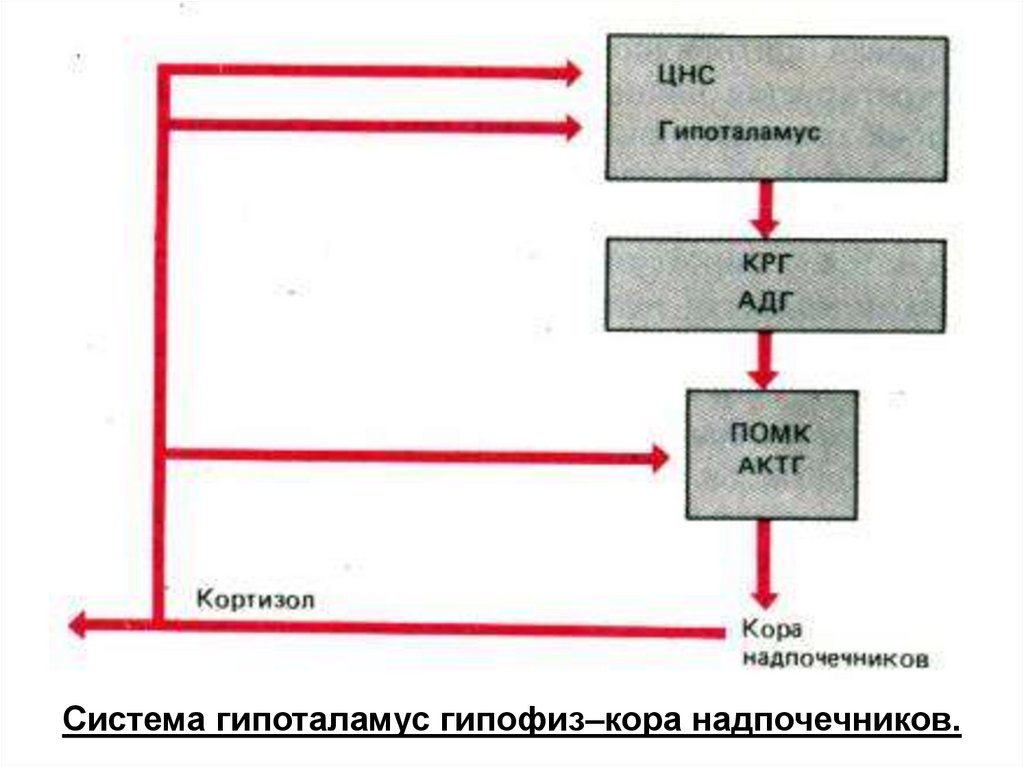
Система гипоталамус гипофиз–кора надпочечников.119.
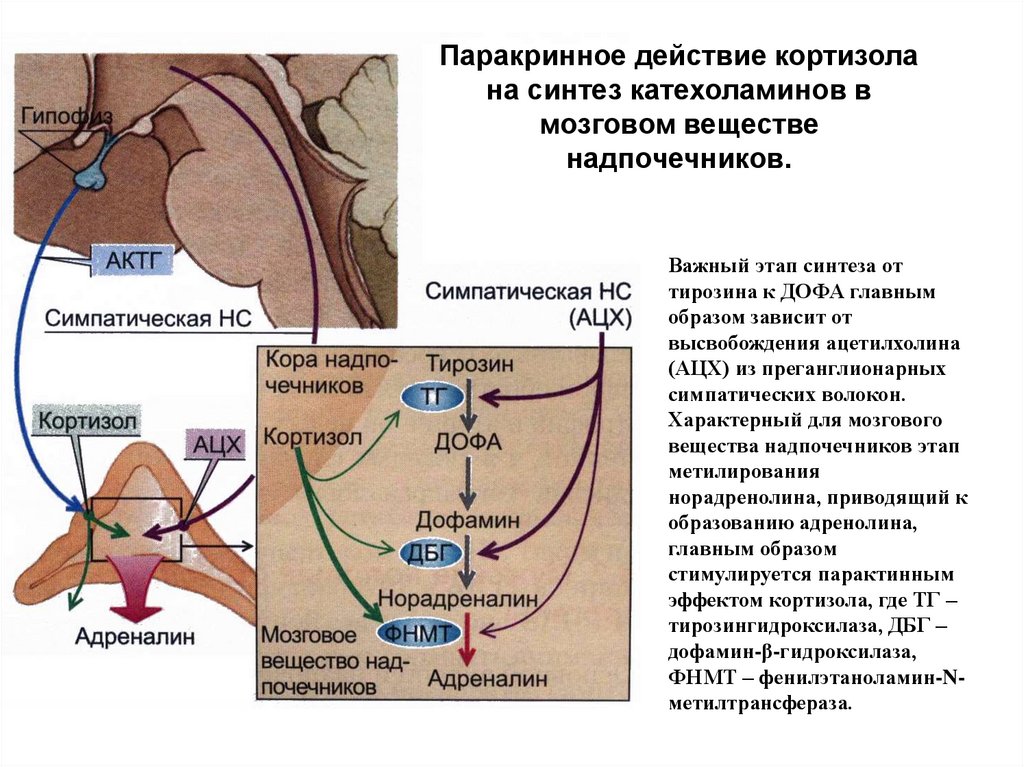
Паракринное действие кортизолана синтез катехоламинов в
мозговом веществе
надпочечников.
Важный этап синтеза от
тирозина к ДОФА главным
образом зависит от
высвобождения ацетилхолина
(АЦХ) из преганглионарных
симпатических волокон.
Характерный для мозгового
вещества надпочечников этап
метилирования
норадренолина, приводящий к
образованию адренолина,
главным образом
стимулируется парактинным
эффектом кортизола, где ТГ –
тирозингидроксилаза, ДБГ –
дофамин-β-гидроксилаза,
ФНМТ – фенилэтаноламин-Nметилтрансфераза.
120.
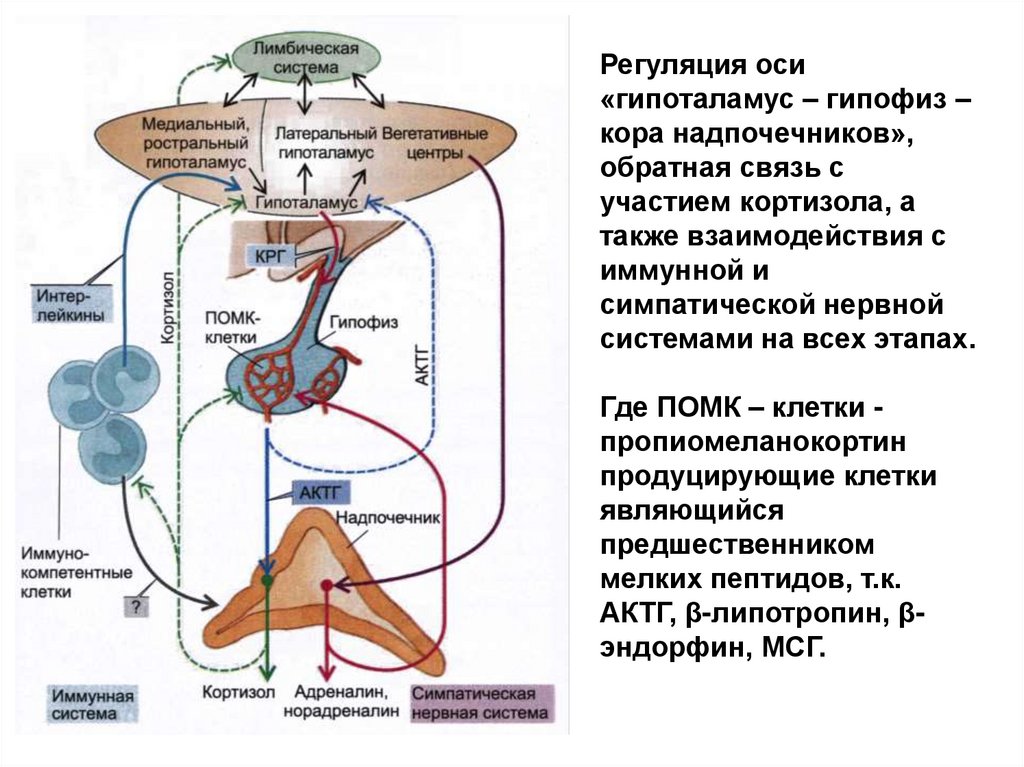
Регуляция оси«гипоталамус – гипофиз –
кора надпочечников»,
обратная связь с
участием кортизола, а
также взаимодействия с
иммунной и
симпатической нервной
системами на всех этапах.
Где ПОМК – клетки пропиомеланокортин
продуцирующие клетки
являющийся
предшественником
мелких пептидов, т.к.
АКТГ, β-липотропин, βэндорфин, МСГ.
121.
Множественное действие КРГ.КРГ действует в ЦНС как
нейротрансмиттер/нейромо
дулятор и участвует в
управлении центрами ВНС.
Действуя в качестве
рилизинг-гормона для АКТГ.
КРГ яв-ся важным
фактором управления
многими реакциями
организма при стрессе.
122. МОРФОЛОГИЧЕСКАЯ ТРИАДА СТРЕССА
• ГИПЕРТРОФИЯ КОРЫ НАДПОЧЕЧНИКОВ• ИНВОЛЮЦИЯ ТИМУСА И ЛИМФАТИЧЕСКИХ УЗЛОВ
• КРОВОИЗЛИЯНИЯ И ИЗЪЯЗВЛЕНИЕ СЛИЗИСТОЙ
ОБОЛОЧКИ ЖЕЛУДКА
123. СТАДИИ СТРЕССА ПО Г.СЕЛЬЕ
Реакциятревоги
СТАДИЯ
РЕЗИСТЕНТНОСТИ
ИСХОДНЫЙ СРЕДНИЙ
УРОВЕНЬ
УСТОЙЧИВОСТИ
СТАДИЯ
ИСТОЩЕНИЯ
124. ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
125.
В островках Лангерганса идентифицировано несколькотипов эндокринных клеток, синтезирующих и
секретирующих пептидные гормоны:
1. инсулин (b-клетки, 70% от всех островковых клеток),
2. глюкагон (a-клетки, 15%),
3. соматостатин (d-клетки),
4. панкреатический полипептид (PP-клетки, F-клетки)
5. у детей младшего возраста — гастрины (G-клетки,
D-клетки).
126. Действие инсулина на углеводный обмен
1. активация утилизации глюкозыклетками,
2. усиление фосфорилирования;
3. подавление распада гликогена;
4. стимуляция синтеза гликогена;
5. угнетение глюконеогенеза;
6. активация процессов гликолиза;
7. гипогликемия.
127. Действие инсулина на белковый обмен
1. повышение проницаемости мембрандля аминоокислот;
2. усиление синтеза иРНК;
3. активация в печени синтеза
aминокислот;
4. повышение синтеза и подавление
распада белка.
128. Действие инсулина на липидный обмен
1. стимуляция синтеза свободных жирных кислот изглюкозы;
2. стимуляция синтеза липопротеиновой липазы в
клетках эндотелия сосудов
3. активация гидролиза связанных с липо-протеинами
крови триглицеридов и поступления жирных кислот
в клетки жировой ткани;
4. стимуляция синтеза триглицеридов;
5. подавление распада жира;
6. активация окисления кетоновых тел в печени.
129. Скорость проявления эффектов инсулина
Физиологические эффекты инсулина по скорости их наступленияподразделяют на:
Быстрый (секунды)
Увеличение транспорта глюкозы (исключение нейроны
головного мозга), аминокислот и K+ внутрь инсулинчувствительных клеток
Медленный (минуты)
Стимуляция белкового синтеза
Торможение распада белков
Активация ферментов (например, гликогенсинтазы и
гликолитических)
Ингибирование гликоген фосфорилазы и ферментов
глюконеогенеза
Отсроченный (часы)
Транскрипция генов, трансляция мРНК, синтез белка, рост и
размножение клеток (инсулин – гормон-митоген)
130. Регуляция
Глюкоза - ведущий регулятор секреции инсулина: приучастии Са2+ стимулирует секрецию инсулина и
тормозит секрецию глюкагона.
Глюкагон является стимулятором секреции как
соматостатина, так и инсулина.
Специфическим ингибитором продукции инсулина
является соматостатин (образуется в кишечнике,
где тормозит всасывание глюкозы). Активация его
секреции происходит под влиянием всасывающихся
в кишечнике аминокислот и гастроинтестинальных
гормонов.
131. Гомеостаз глюкозы
поддерживают следующие механизмы:1. Печень демпфирует колебания концентрации глюкозы за счет
обратимой реакции глюкоза - гликоген.
2. Инсулин и глюкагон реципрокно регулируют нормальное
содержание глюкозы в крови.
3. Гипогликемия оказывает прямое влияние на гипоталамус,
который возбуждает симпатическую нервную систему. В
результате адреналин секретируется из надпочечников и
увеличивает выделение глюкозы печенью.
4. Продолжительная гипогликемия стимулирует выделение СТГ и
кортизола, которые уменьшают скорость потребления глюкозы
большинством клеток организма, что помогает возвращению
концентрации глюкозы в крови к нормальному уровню.
132. Функциональная организация островков Лангерганса
сплошные стрелки — стимуляция, пунктирные — подавление гормональных секретов.Ведущий регулятор — глюкоза — при участии кальция стимулирует секрецию инсулина b-клетками и, напротив,
тормозит секрецию глюкагона а-клетками. Всасывающиеся в желудке и кишечнике аминокислоты
являются стимуляторами функции всех клеточных элементов.
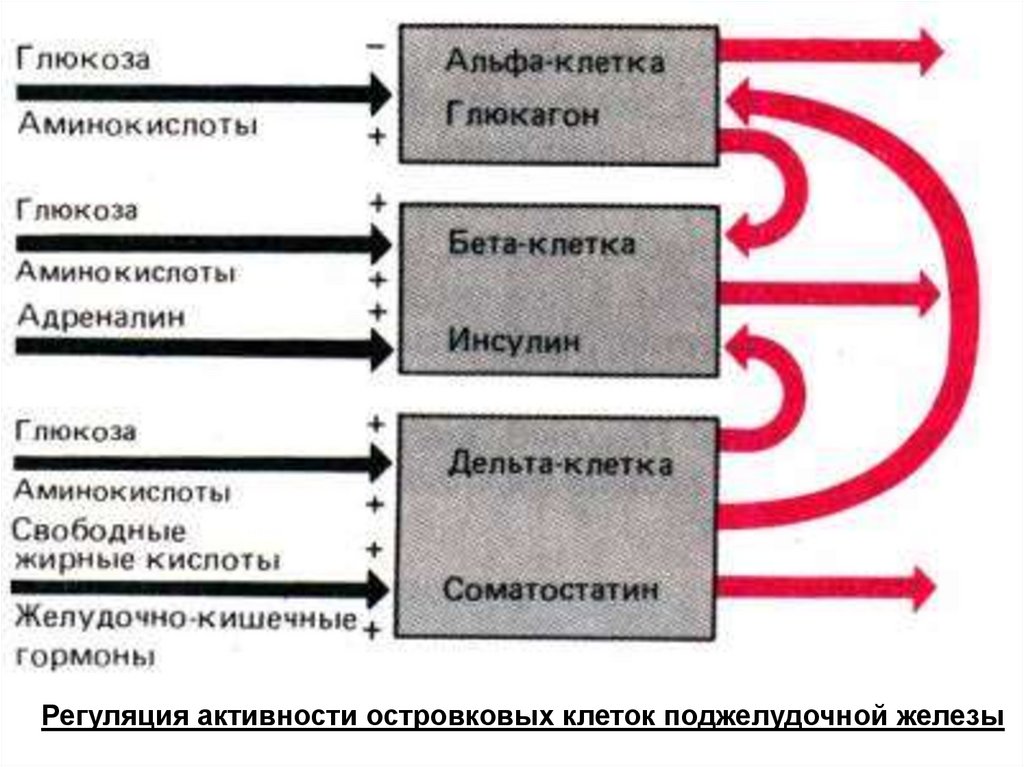
Ведущий «внутриорганный» ингибитор секреции инсулина и глюкагона — соматостатин, активация его
секреции происходит под влиянием всасывающихся в кишечнике аминокислот и гастроинтестинальных
гормонов при участии ионов Са2+. Глюкагон является стимулятором секреции как соматостатина, так и
инсулина.
133. Сахарный диабет типа I
развивается в результате аутоиммунной деструкции b-клетокостровков Лангерганса и значительно реже вследствие
мутаций гена инсулина и генов, принимающих участие в синтезе
и секреции инсулина.
Сопровождается:
поражением почек (диабетическая нефропатия),
сетчатки (диабетическая ретинопатия),
артериальных сосудов (диабетическая ангиопатия),
периферических нервов (диабетическая невропатия)
осмотическим диурезом (избыток кетонов и глюкозы )
метаболическим кетоацидозом (кетоны – органические кислоты)
134. Сахарный диабет типа II
развивается в результате :1. либо нарушения секреции инсулина (избыточное
содержание сахара в крови не увеличивает секрецию
инсулина),
2. либо извращения реакции клеток–мишеней на
инсулин (развивается резистентность к инсулину),
3. либо имеют значение оба фактора.
Поскольку дефицита инсулина нет, то вероятность
развития метаболического кетоацидоза низка.
135. Тимус
Тимус (вилочковая железа) является центральным органом иммунитета,обеспечивающим продукцию специфических Т-лимфоцитов.
Тимоциты секретируют в кровь гормоны (тимозины и тимопоэтины), которые:
• обеспечивают пролиферацию Т-лимфоцитов,
• обеспечивают их созревание (дифференцировка),
• обеспечивают отбор пригодных для организма клеток и гибель непригодных,
• влияют на процессы синтеза клеточных рецепторов к медиаторам и гормонам,
• стимулируют разрушение ацетилхолина в нервно-мышечных синапсах,
• регулируют состояние углеводного и белкового обмена,
• регулируют состояние обмена кальция,
• регулируют функции щитовидной и половых желез,
• модулируют эффекты глюкокортикоидов, тироксина (антагонизм) и соматотропина
(синергизм).
В целом вилочковая железе рассматривается как орган интеграции иммунной и
эндокринной систем организма.
136. Яички
В яичках синтезируются стероидные андрогены и a-ингибин.Тестостерон — основной циркулирующий андроген. В эмбриогенезе андрогены
контролируют развитие плода по мужскому типу. В период полового созревания
они стимулируют становление признаков мужского пола. С наступлением
половой зрелости тестостерон необходим для поддержания сперматогенеза,
вторичных половых признаков, секреторной активности предстательной железы
и семенных пузырьков.
Дигидротестостерон образуется из тестостерона в клетках Лейдига, простате,
семенных пузырьках. Это самый сильный естественный андроген. Он
определяет физическое развитие во время полового созревания у мужчин,
регулирует сексуальное поведение и эректильность, развитие гениталий у
мужчин и предстательной железы.
a-Ингибин. Это гликопротеидный гормон синтезируется в клетках Сертоли извитых
семенных канальцев и блокирует синтез гипофизарного ФСГ
137. Яичники
В яичниках синтезируются стероидные женские половые гормоны, гликопротеиновые гормоныингибины и пептидной природы релаксины.
Эстрогены в период полового созревания стимулируют становление признаков женского
пола. У женщин детородного возраста эстрогены активируют пролиферацию
фолликулярных клеток, а в эндометрии контролируют пролиферативную фазу
менструального цикла.
Эстрадиол, эстрон и эстриол— обладают выраженной эстрогенной активностью.
Прогестерон синтезируют клетки жёлтого тела, а также клетки хориона при наступлении
беременности. Прогестерон в эндометрии контролирует секреторную фазу менструального
цикла и существенно увеличивает порог возбудимости ГМК миометрия. Стимулируют
синтез прогестерона ЛГ и ХГТ.
Релаксины — синтезируется клетками жёлтого тела и цитотрофобластом, при
беременности оказывают расслабляющий эффект на ГМК миометрия, а перед родами
приводят к размягчению лонного сочленения и шейки матки.
Ингибины - подавляют синтез и секрецию гипоталамического гонадолиберина и
гипофизарного ФСГ.
138. Гормоны, вырабатываемые в плаценте
Плацента тесно анатомически и функционально связана с
организмами матери и плода. Синтез в плаценте эстриола происходит
не только из эстрадиола матери, но и из дегидроэпиандростерона,
образуемого надпочечниками плода.
В плаценте образуется прогестерон, действующий преимущественно
на мускулатуру матки.
В наибольших количествах при беременности плацентой
продуцируется хорионический гонадотропин, оказывающий
регуляторные эффекты не только на процессы дифференцировки и
развития плода, но и на метаболизм в организме матери. Гормон
обеспечивает в организме матери задержку солей и воды,
необходимых для растущего плода, стимулирует секрецию
вазопрессина, активирует механизмы иммунитета у матери.
139. Гормоны, вырабатываемые в почке
Ренин не является гормоном, этот фермент — начальное звено в ренин-ангиотензиновойсистеме, важнейшего регулятора системного АД. Ренин синтезируется в видоизменённых
(эпителиоидных) ГМК стенки приносящих артериол почечных телец.
Регуляторы синтеза и секреции ренина:
1. симпатическая иннервация (стимуляция), опосредуемая b-адренорецепторами
2. ангиотензины (по принципу отрицательной обратной связи);
3. рецепторы плотного пятна (регистрация содержания NaCl в канальцах нефрона);
4. барорецепторы в стенке приносящей артериолы почечных телец.
Кальцитриол — активная форма витамина D3 — синтезируется в митохондриях
проксимальных извитых канальцев, способствует всасыванию кальция и фосфатов в
кишечнике, стимулирует остеобласты (ускоряет минерализацию костей).
Образование кальцитриола стимулируют ПТГ и гипофосфатемия, подавляет —
гиперфосфатемия.
Эритропоэтин —синтезируется интерстициальными клетками, в ответ на гипоксию
стимулирует эритропоэз на стадии формирования проэритробластов.
Вазодилататоры — в частности, брадикинин и некоторые простагландины синтезируются в
интерстициальных клетках мозгового вещества почки.
140. Гормоны, вырабатываемые в сердце
Натрийуретический фактор синтезируюткардиомиоциты правого предсердия и некоторые
нейроны ЦНС.
Мишени — клетки почечных телец, собирательных
трубочек почки, клубочковой зоны коры
надпочечников, ГМК сосудов.
Функции — контроль объёма внеклеточной жидкости и
гомеостаза электролитов (угнетение синтеза и
секреции альдостерона, ренина, вазопрессина). Эти
пептиды оказывают мощный сосудорасширяющий
эффект и снижают АД.
141. Гормоны, вырабатываемые в ЖКТ
В стенке трубчатых органов ЖКТ присутствует огромное количествосекретирующих гормоны разнообразных эндокринных клеток
(энтероэндокринные клетки):
Гастрин стимулирует секрецию HCl париетальными клетками слизистой
оболочки желудка.
Секретин стимулирует выделение бикарбоната и воды из секреторных
клеток желёз двенадцатиперстной кишки и поджелудочной железы. Он
регулирует рН в проксимальном отделе тонкой кишки
Холецистокинин стимулирует сокращения жёлчного пузыря и выделение
ферментов из поджелудочной железы
Вазоактивный интестинальный полипептид (ВИП) Из оказывает
многочисленные антигастриновые и проглюкагоновые эффекты. Кроме
того, через свои рецепторы гормон в основном подавляет функции Т—
и В—лимфоцитов, моноцитов и нейтрофилов на уровне
иммунокомпетентных структур.
142. АПУД-система
APUD — аббревиатура, образованная из первых букв англ. словamines - амины, precursor - предшественник, uptake - усвоение,
поглощение, decarboxylation -декарбоксилирование.
Система клеток, способных к выработке и накоплению биогенных
аминов и (или) пептидных гормонов и имеющих общее
эмбриональное происхождение. АПУД-систему составляют
около 40 типов клеток, обнаруживаемых в ЦНС (гипоталамусе,
мозжечке), железах внутренней секреции (гипофизе,
шишковидном теле, щитовидной железе, островках
поджелудочной железы, надпочечниках, яичниках), в
желудочно-кишечном тракте, легких, почках и мочевых путях,
параганглиях и плаценте.
143. АПУД-система
Предполагают, что единым эмбриональнымпредшественником клеток АПУД-системы является
так называемый нейроэндокриннопрограммированный эпибласт. Кроме этого
апудоциты имеют еще одну общую черту - наличие в
них особого фермента - нейронспецифической
енолазы
Наиболее изученной является АПУД-система
желудочно-кишечного тракта и поджелудочной
железы, объединяемая в отдельную
гастроэнтеропанкреатическую эндокринную систему,
на долю которой приходится около половины всех
апудоцитов
144. Гастроэнтеропанкреатическая эндокринная система
Гастроэнтеропанкреатическая эндокринная система
Тип апудоцитов
Основная локализация апудоцитов
Основные продуцируемые вещества
D
Желудок, тонкая кишка, поджелудочная железа
Соматостатин
D1
Желудок, тонкая кишка, поджелудочная железа
Вазоактивный интестинальный полипептид (ВИП)
ЕС
Желудок, тонкая и толстая кишка
Субстанция Р, серотонин
ECL
Желудок
Серотонин, гистамин, дофамин
G
Желудок
Гастрин, энкефалины, эндорфины
I
Тонкая кишка
Холецистокинины
К
Тонкая кишка
Гастроингибирующий полипептид (ГИП)
L—EG
Тонкая и толстая кишка
Глюкагоноподобный иммунореактивный пептид
Мо
Тонкая кишка
Мотилин, серотонин
N
Тонкая кишка
Нейротензин
Р
Желудок, тонкая кишка
Гастрин-рилизинг-пептид
S
Тонкая кишка
Секретин
А
Поджелудочная железа
Глюкагон, энкефалины, эндорфины
В
Поджелудочная железа
Инсулин
PP (F)
Поджелудочная железа
Панкреатический полипептид (ПП)
145. Гормоны поджелудочной железы и их функции: α- альфа-клетки, продуцирующие глюкогон, β – бетта-клетки, продуцирующие инсулин, D
– клетки – соматостатин.146.
Регуляция активности островковых клеток поджелудочной железы147.
Спасибоза ваше
внимание