
 — биологически активные вещества органической природы, вырабатывающиеся в")



































")














")

")














")































")
")















")


![ЭФФЕКТЫ КАЛЬЦИТРИОЛА [1,25 (ОН)2 Д3]](https://cf4.ppt-online.org/files4/slide/q/QasAfjmXFpqT7c1nNv6MdPoxw4rk5HViUY3zGJ/slide-142.jpg "ЭФФЕКТЫ КАЛЬЦИТРИОЛА [1,25 (ОН)2 Д3]")





 biology
biologySimilar presentations:







")


Химическая природа, биосинтез, секреция и катаболизм гормонов
1. Химическая природа, биосинтез, секреция и катаболизм гормонов.
2. Гормо́ны (др.-греч. ὁρμάω — возбуждаю, побуждаю) — биологически активные вещества органической природы, вырабатывающиеся в
специализированных клетках желёзвнутренней секреции, поступающие в
кровь и оказывающие регулирующее
влияние на обмен веществ и
физиологические функции.
Гормоны служат гуморальными
(переносимыми с кровью) регуляторами
определённых процессов в различных
органах и системах.
3. Регуляторные вещества
ВнутриклеточныеУтилизоны
(неспецифический
межклеточный
контроль)
Межклеточные
Информоны (цитомедины)
(специальный межклеточный контроль)
Нейромедиаторы
Гистогормоны
(тканевые)
Гормоны
4. ОСНОВНЫЕ ПРИЗНАКИ ГОРМОНОВ
1. Высокая специфичность действия.Гормон оказывает своё действие только после
связывания с рецептором. Рецептор – это сложный
белок-гликопротеин, состоящий из белковой и
углеводной частей. Гормон связывается именно с
углеводной частью рецептора. Причём строение
углеводной части имеет уникальную химическую
структуру и соответствует пространственному
строению гормона. Поэтому гормон безошибочно,
точно, специфично связывается только со своим
рецептором, несмотря на малую концентрацию
гормона в крови.
5. ОСНОВНЫЕ ПРИЗНАКИ ГОРМОНОВ
2. Секретируемость - способностьпреодолевать клеточный барьер
6. ОСНОВНЫЕ ПРИЗНАКИ ГОРМОНОВ
3. Высокая биологическая активность(физиологическая концентрация 10-12 ммоль)
– Концентрация гормонов в крови очень
мала, но их действие сильно выражено,
поэтому даже небольшое увеличение или
уменьшение уровня гормона в крови
вызывает различные, часто значительные,
отклонения в обмене веществ и
функционировании органов и может
привести к патологии
7. ОСНОВНЫЕ ПРИЗНАКИ ГОРМОНОВ
4. Дистантность действия Гормонывырабатываются в одних органах
(эндокринных железах), а действуют в
других (тканях мишенях).
8. КЛАССИФИКАЦИЯ ГОРМОНОВ:
1. по химическому строению2. по биохимическим действиям,
биологическим функциям
3. по месту образования
4. по механизму действия
5. по функции гормонов
9. КЛАССИФИКАЦИЯ ГОРМОНОВ по химическому строению:
КЛАССИФИКАЦИЯ ГОРМОНОВПО ХИМИЧЕСКОМУ СТРОЕНИЮ:
10. 1. Классификация гормонов и БАВ по химической структуре:
1. Производные аминокислот:производные тирозина: тироксин, трийодтиронин, дофамин,
адреналин, норадреналин;
производные триптофана: мелатонин, серотонин;
производные гистидина: гистамин.
2. Белково-пептидные гормоны:
полипептиды: глюкагон, кортикотропин, меланотропин,
вазопрессин, окситоцин, пептидные гормоны желудка и кишечника;
простые белки (протеины): инсулин, соматотропин, пролактин,
паратгормон, кальцитонин;
сложные белки (гликопротеиды): тиреотропин, фоллитропин,
лютропин.
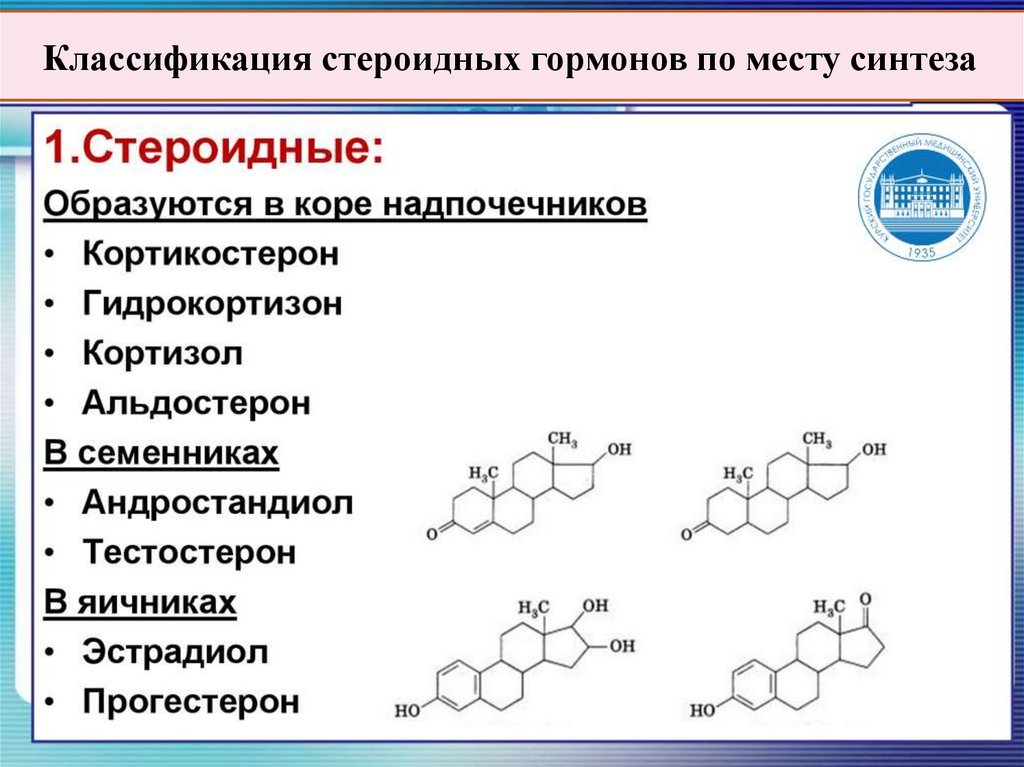
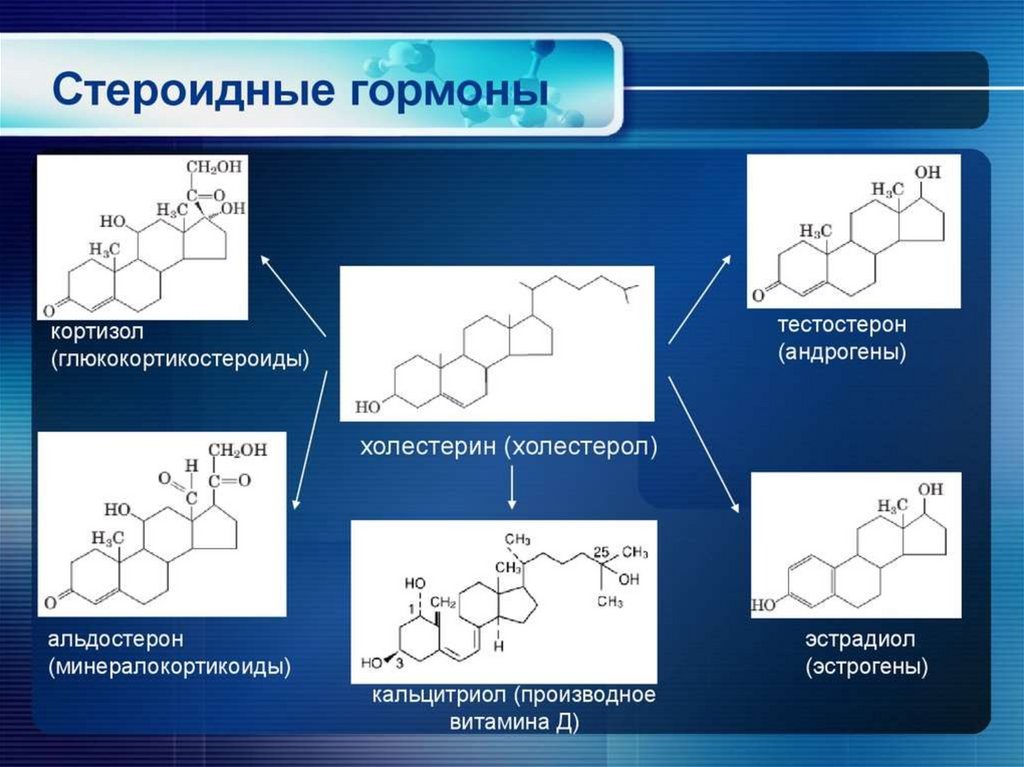
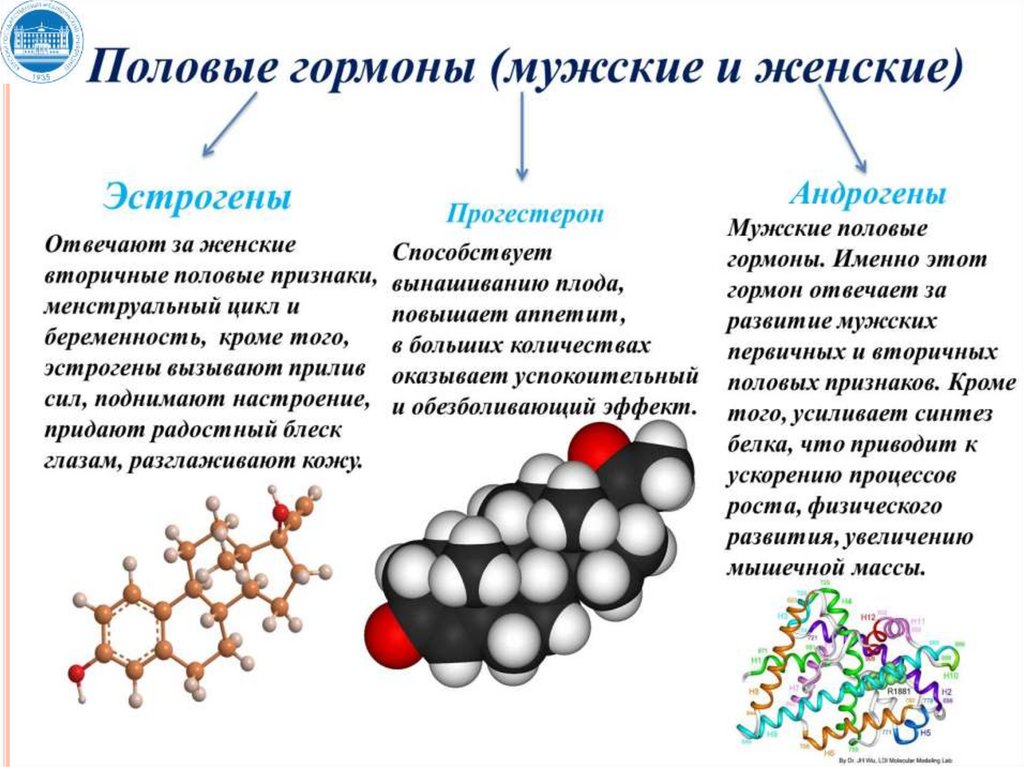
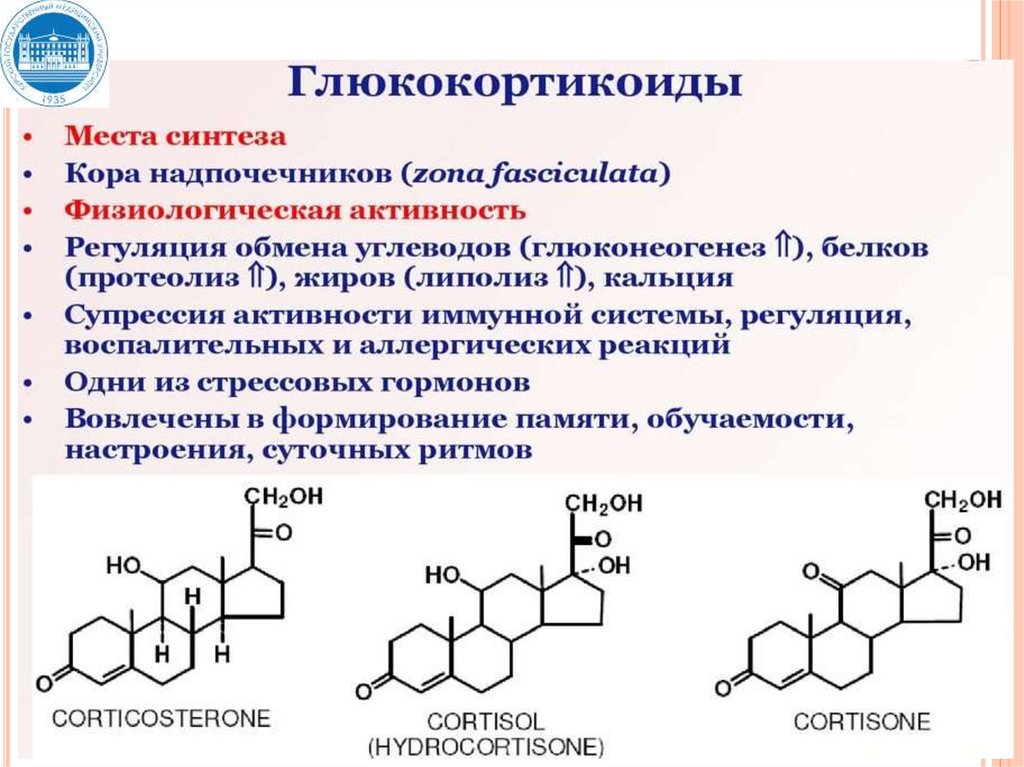
3. Стероидные гормоны:
кортикостероиды (альдостерон, кортизол, кортикостерон);
половые гормоны: андрогены (тестостерон), эстрогены и
прогестерон.
4. Производные жирных кислот:
арахидоновая кислота и ее производные: простагландины:
простациклины, тромбоксаны, лейкотриены.
11. 2. По биохимическим действиям, биологическим функциям различают 5 видов гормонов:
1.2.
3.
4.
5.
гормоны, регулирующие обмен белков, углеводов,
липидов: инсулин, глюкагон, адреналин, кортизол.
гормоны, регулирующие водно-солевой обмен в
организме: альдостерон, вазопрессин.
гормоны, регулирующие обмен ионов кальция и
фосфатов в организме: половые гормоны:
паратгормон, кальцитонин, кальцитриол.
гормоны, регулирующие репродуктивную функцию в
организме: половые гормоны (мужские и женские).
гормоны, регулирующие функции эндокринных
желез: АКТГ, тиреотропный, ЛГ, ФСГ, соматотропин,
меланотропный.
12. 3. По месту образования гормонов:
13. 3. По месту образования гормонов:
14. 4. По механизму действия гормонов:
15. 5. По функции гормонов:
Эффекторные гормоны — гормоны, которыеоказывают влияние непосредственно на органмишень.
Тропные гормоны — гормоны, основной
функцией которых является регуляция синтеза
и выделения эффекторных гормонов.
Выделяются аденогипофизом.
Рилизинг-гормоны — гормоны, регулирующие
синтез и выделение гормонов аденогипофиза,
преимущественно тропных. Выделяются
нервными клетками гипоталамуса.
16. ФУНКЦИОНАЛЬНЫЕ ФРАГМЕНТЫ В СТРУКТУРЕ ГОРМОНОВ
1. Адресный фрагмент (гаптомер) – обеспечивает поискмест
специфического
действия,
избирательно
связывается с рецепторами клеток-мишеней, не
производит биологический эффект гормона;
2. Актон (эффектомер) – фрагмент, обеспечивающий
включение гормональных эффектов, плохо связывается с
рецепторами клеток-мишеней;
3. Вспомогательный (дополнительный) фрагмент – отвечает
за:
- конформацию гормона;
- его стабильность;
- регулирует его активность;
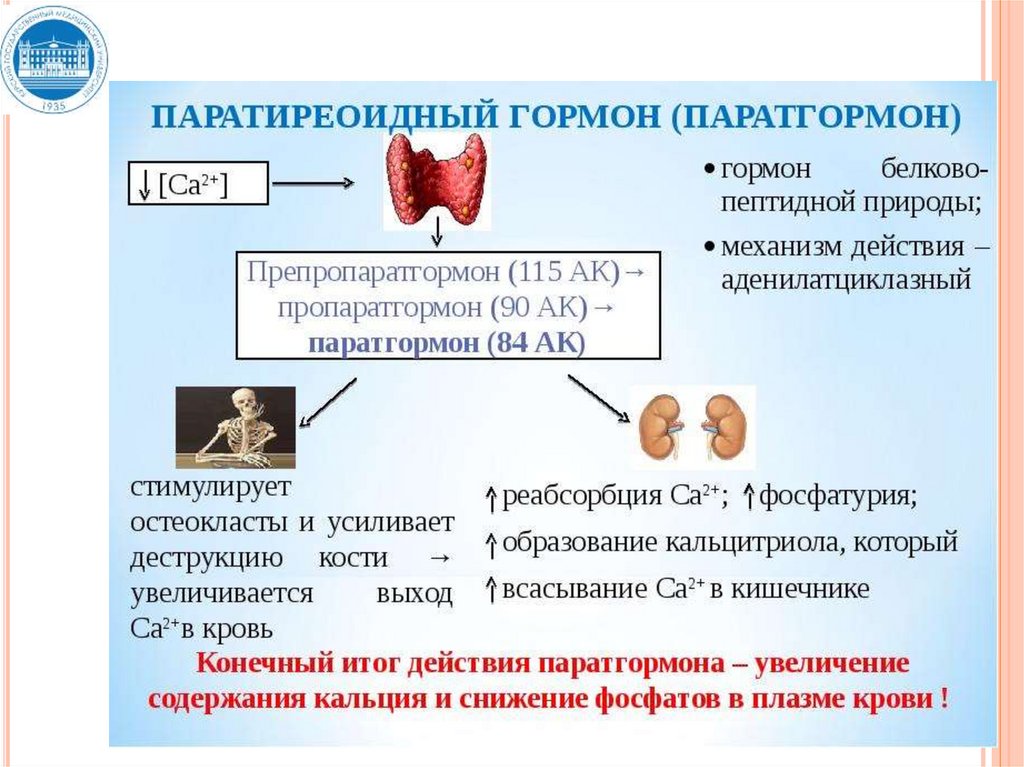
- иммунологические свойства (видовая принадлежность
17. Физиологическая организация эндокринной функции
ФИЗИОЛОГИЧЕСКАЯ ОРГАНИЗАЦИЯЭНДОКРИННОЙ ФУНКЦИИ
1. Синтез и секреция гормона
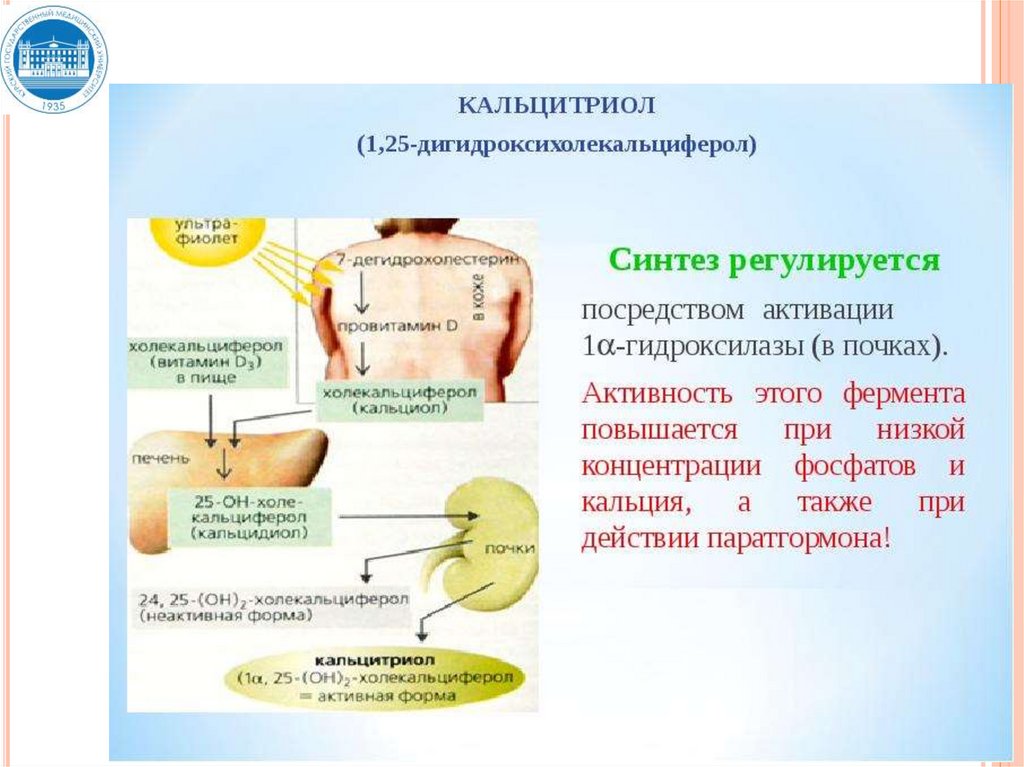
2. Регуляция и саморегуляция
функции эндокринной железы
3. Транспорт
4. Взаимодействие с клеткоймишенью
5. Периферический метаболизм и
выведение
18. Виды секреции гормонов (освобождение гормонов из эндокринных желез в венозную кровь или лимфу, что поддерживает их уровень в
ВИДЫ СЕКРЕЦИИ ГОРМОНОВ(ОСВОБОЖДЕНИЕ ГОРМОНОВ ИЗ ЭНДОКРИННЫХ ЖЕЛЕЗ В ВЕНОЗНУЮ КРОВЬ ИЛИ
ЛИМФУ, ЧТО ПОДДЕРЖИВАЕТ ИХ УРОВЕНЬ В ЦИРКУЛИРУЮЩИХ ЖИДКОСТЯХ)
1. Освобождение гормона из клеточных секреторных
гранул, которые способны перемещаться в клетках
эндокринных желез (белково-пептидные гормоны,
катехоламины);
2. Освобождение гормона из белковосвязанной
формы (тиреоидные гормоны);
3. Относительно свободная диффузия гормона через
клеточные мембраны (стероидные гормоны).
Последний тип секреции наиболее сопряжен во
времени с процессом синтеза гормонов.
19. Этапы обмена гормонов
ЭТАПЫ ОБМЕНА ГОРМОНОВ1) Синтез и секреция гормона
2) Регуляция и саморегуляциия
функции эндокринной железы
3) Транспорт
4) Взаимодействие с клеткоймишенью
5) Периферический метаболизм и
выведение
20. Взаимосвязь регуляторных систем организма
21. Для нормального функционирования многоклеточного организма необходима взаимосвязь между отдельными клетками, тканями и
ДЛЯ НОРМАЛЬНОГО ФУНКЦИОНИРОВАНИЯМНОГОКЛЕТОЧНОГО ОРГАНИЗМА НЕОБХОДИМА
ВЗАИМОСВЯЗЬ МЕЖДУ ОТДЕЛЬНЫМИ КЛЕТКАМИ,
ТКАНЯМИ И ОРГАНАМИ. ЭТУ ВЗАИМОСВЯЗЬ
ОСУЩЕСТВЛЯЮТ:
нервная система (центральная и периферическая) через
нервные импульсы и нейромедиаторы;
эндокринная система через эндокринные железы и
гормоны, которые синтезируются
специализированными клетками этих желез,
выделяются в кровь и транспортируются к различным
органам и тканям;
паракринная и аутокринная системы посредством
различных соединений, которые секретируются в
межклеточное пространство и взаимодействуют с
рецепторами либо близлежащих клеток, либо той же
клетки (простагландины, гормоны желудочнокишечного тракта, гистамин и др.);
иммунная система через специфические белки
22. Транспорт гормонов
ТРАНСПОРТ ГОРМОНОВФорменные
элементы крови
15-20%
Плазма крови
80-85%
Гормонспецифические
белки
80%
КСГ – кортикосвязывающий глобулин
ССГ – секссвязывающий глобулин
ТСГ – тиреосвязывающий глобулин
ИСГ – инсулинсвязывающий глобулин
и др.
В свободном виде
10%
Гормоннеспецифические
белки
10%
Альбумины, α-кислый
гликопротеид, γ-глобулины,
трансферрин, трипсин и др.
Эритроциты
80%
Лейкоциты
20%
23. Транспорт гормонов
ТРАНСПОРТ ГОРМОНОВ24.
25.
26.
27.
28.
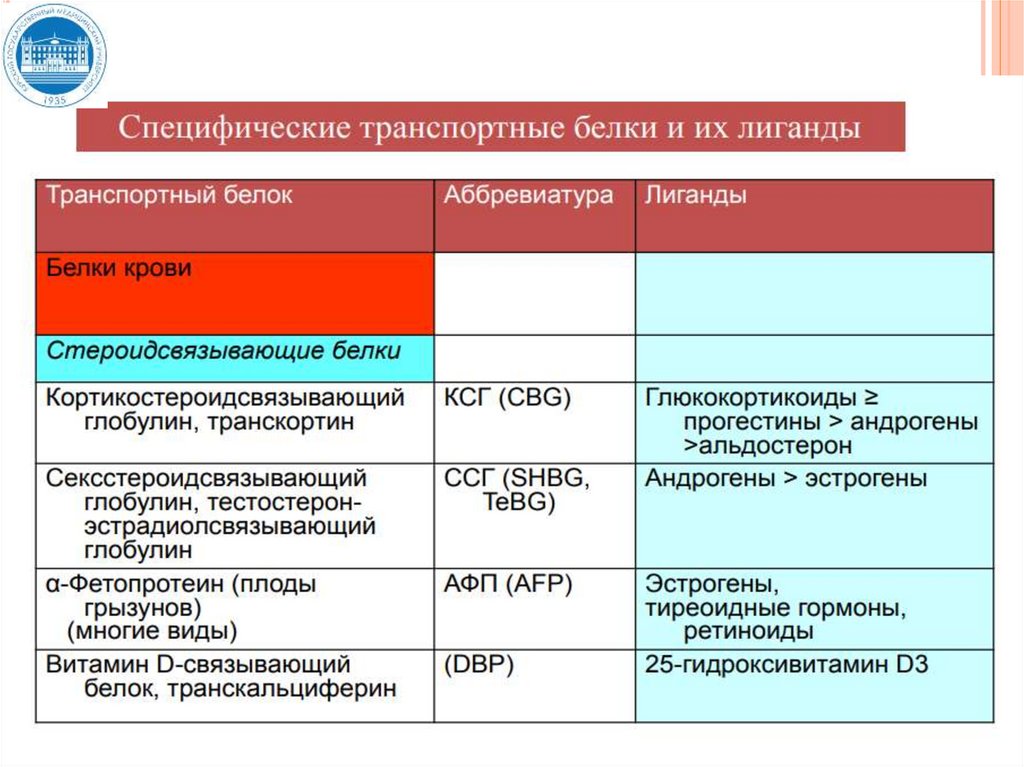
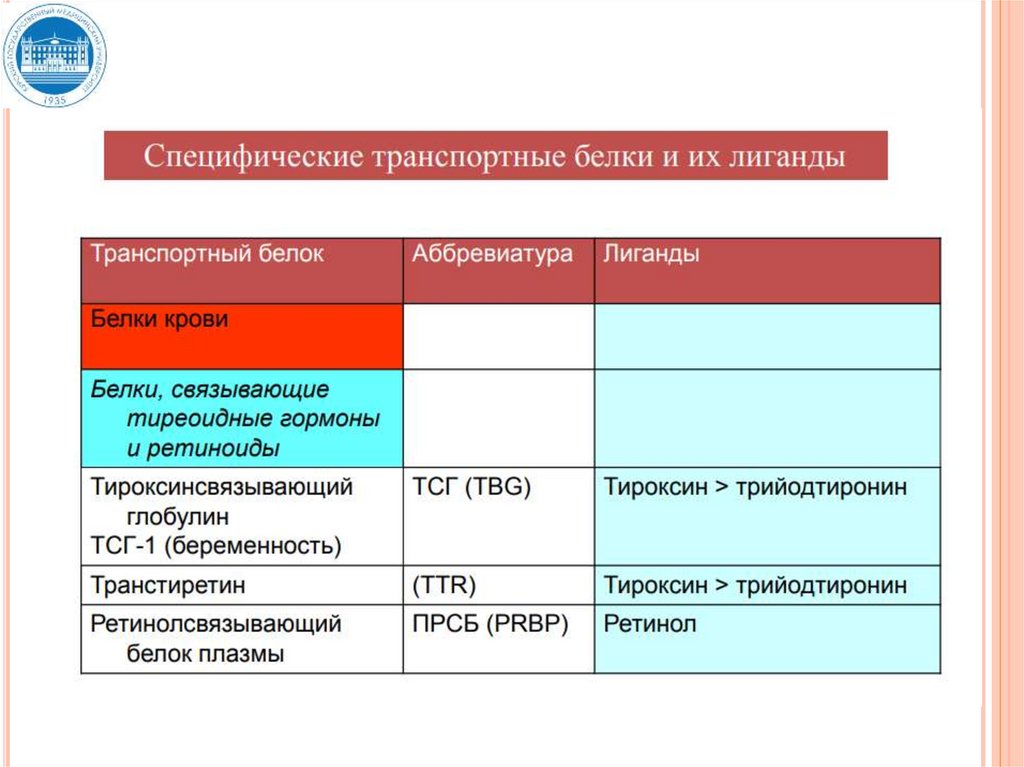
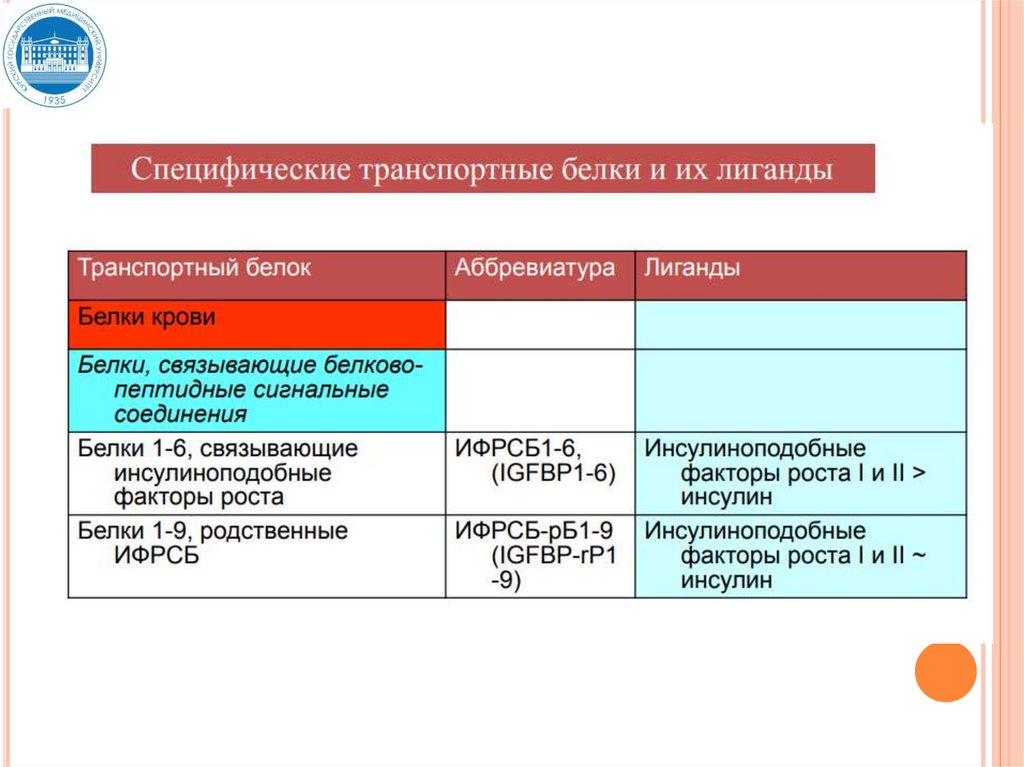
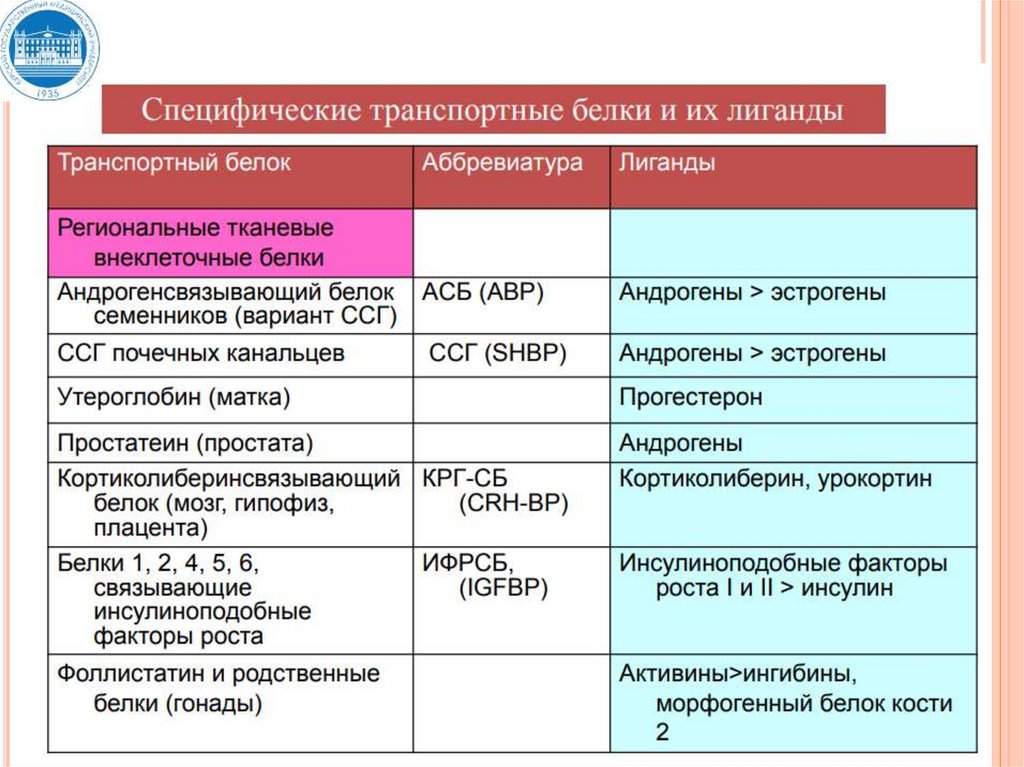
29. Связывание гормонов с транспортными белками
СВЯЗЫВАНИЕ ГОРМОНОВ СТРАНСПОРТНЫМИ БЕЛКАМИ
30. Метаболизм гормонов
МЕТАБОЛИЗМ ГОРМОНОВ31. Метаболизм гормонов
МЕТАБОЛИЗМ ГОРМОНОВ32. Метаболизм гормонов
МЕТАБОЛИЗМ ГОРМОНОВ33. Катаболизм гормонов
КАТАБОЛИЗМ ГОРМОНОВБелково-пептидные гормоны подвергаются протеолизу,
распадаются до отдельных аминокислот. Эти аминокислоты
дальше вступают в реакции дезаминирования,
декарбоксилирования, трансаминирования и распадаются до
конечных продуктов: NH3, CO2 и Н2О.
Гормоны - производные аминокислот подвергаются
окислительному дезаминированию и дальнейшему окислению
до СО2 и Н2О.
Стероидные гормоны не подвергаются распаду, так как в
организме нет ферментных систем, которые обеспечивали бы их
распад. В основном происходит модификация боковых
радикалов путем введения дополнительных гидроксильных
групп. Гормоны становятся более гидрофильными и в таком
виде выводятся с мочой.
34. Реакции периферического метаболизма гормонов различной химической природы
РЕАКЦИИ ПЕРИФЕРИЧЕСКОГО МЕТАБОЛИЗМАГОРМОНОВ РАЗЛИЧНОЙ ХИМИЧЕСКОЙ ПРИРОДЫ
Стероидные
Восстановление
двойной связи в
кольце,
конъюгирование
Производные
аминокислот
Белковопептидные
Протеолиз
Гидроксилирование
углеродных атомов
Метилирование
гидроксила
(катехоламины,
мелатонин)
Деиодирование
(тиреоидные)
Окислительное
дезаминирование
(катехоламины)
35. механизмы регуляции синтеза гормонов
МЕХАНИЗМЫ РЕГУЛЯЦИИ СИНТЕЗАГОРМОНОВ
1. Регуляция через гипоталамус
(например либерины, статины)
2. Регуляция через горомоны
(например АКТГ, СТГ)
3. Регуляция через метаболиты
(например глюкоза,
аминокислоты, ионы)
36. Динамика и механизмы реализации гормональных эффектов в клетке
ДИНАМИКА И МЕХАНИЗМЫ РЕАЛИЗАЦИИГОРМОНАЛЬНЫХ ЭФФЕКТОВ В КЛЕТКЕ
Начальные
(сек-до 2 часов)
Г
Г
+ Р
Р
Химическая
модификация
белков
Изменение
активности
белков
Эффекты
Ранние
(меньше 24 ч-48 ч)
Изменение
транскрипции
Изменение
трансляции
Эффекты
Поздние
(более 48 ч)
Изменение
репликации
Эффекты
37. Синтез белково – пептидных гормонов
СИНТЕЗБЕЛКОВО – ПЕПТИДНЫХ
ГОРМОНОВ
38. СТРОЕНИЕ И БИОСИНТЕЗ БЕЛКОВО-ПЕПТИДНЫХ ГОРМОНОВ
СТРОЕНИЕ И БИОСИНТЕЗ БЕЛКОВОПЕПТИДНЫХ ГОРМОНОВ1. Пептидные гормоны синтезируются в процессе
трансляции из аминокислот. Некоторые пептидные гормоны это короткие пептиды; например, гормон гипоталамуса
тиреотропин - либерин - трипептид. Большинство гормонов
передней доли гипофиза - гликопротеины.
Некоторые пептидные гормоны являются продуктами общего
гена. Большинство полипептидных гормонов синтезируется в
виде неактивных предшественников - препрогормонов.
Образование активных гормонов происходит путем
частичного протеолиза.
2. Инсулин - полипептид, состоящий из двух полипептидных
цепей. Цепь А содержит 21 аминокислотный остаток, цепь В 30 аминокислотных остатков. Обе цепи соединены между
собой двумя дисульфидными мостиками. Молекула инсулина
содержит также внутримолекулярный дисульфидный мостик в
А-цепи.
39. Схема биосинтеза белково-пептидных гормонов
СХЕМА БИОСИНТЕЗАБЕЛКОВО-ПЕПТИДНЫХ ГОРМОНОВ
Ген (ДНК)
Полирибосомы
Протеаза I
м-РНК
Пропрегормон
Пептид 1
Прогормон
Протеаза II
Гормон
Пептид 2
Низкомолекулярные гормоны синтезируются в цитоплазме под
влиянием соответствующих ферментов (которые в свою
очередь образуются по приведенной схеме, что
предопределяет видовую принадлежность гормона)
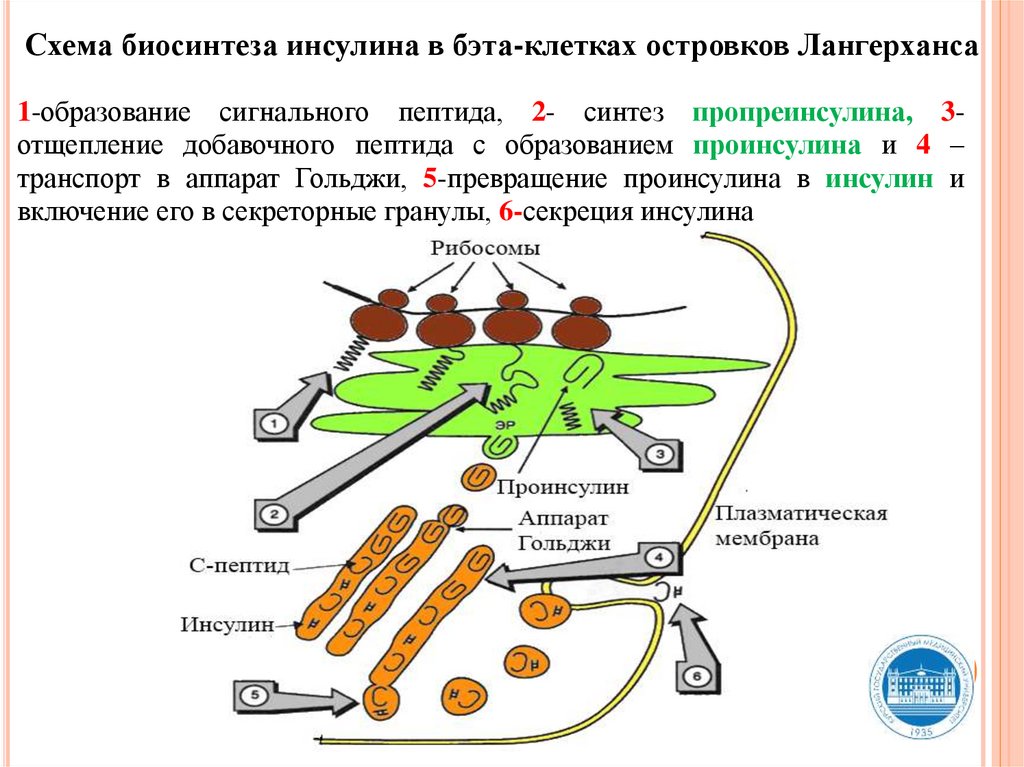
40. Схема биосинтеза инсулина в бэта-клетках островков Лангерханса
СХЕМА БИОСИНТЕЗА ИНСУЛИНА ВБЭТА-КЛЕТКАХ ОСТРОВКОВ ЛАНГЕРХАНСА
• Синтез пропреинсулина (ММ - 11,5 кД) на полирибосомах,
прикрепленных
к
наружной
поверхности
мембраны
эндоплазматического ретикулума (104-110 аминокислот)
N
23-24
добавочный пептид
30 (бэта-цепь)
32-37
21 (А-цепь)
C
вставочный
пептид
• пропреинсулин за счет добавочного N-пептида (23-24 в основном
гидрофобных аминокислот) проникает из ЭР в аппарат Гольджи
цитоплазмы - образование проинсулина
• вырезание вставочного пептида (32-37 аминокислот) из проинсулина образование инсулина (2-х цепочечная структура из 51 аминокислоты
бэта и А-цепочек, соединенных двумя дисульфидными связями, ММ 5,7 кД), включение в секреторные гранулы
41.
Схема биосинтеза инсулина в бэта-клетках островков Лангерханса1-образование сигнального пептида, 2- синтез пропреинсулина, 3отщепление добавочного пептида с образованием проинсулина и 4 –
транспорт в аппарат Гольджи, 5-превращение проинсулина в инсулин и
включение его в секреторные гранулы, 6-секреция инсулина
42. Анаболические эффекты инсулина
АНАБОЛИЧЕСКИЕ ЭФФЕКТЫ ИНСУЛИНА1. Начальные эффекты (в течении нескольких секундминут,
изменение трансмембранного транспорта,
фосфорилирование и дефосфорилирование белков,
активация и ингибирование ферментов)
2. Ранние, поздние эффекты (через несколько часов –
суток, активация синтеза РНК, ДНК, белков, усиление
пролиферации, дифференцировки, усиление синтеза
СТГ)
Инсулин наиболее всего активирует анаболические
процессы в мышцах, печени, почках, соединительной
ткани.
Кроме этого инсулин обеспечивает анаболизм белков
энергией за счет регуляции углеводного обмена.
43. Синтез гормонов производных аминокислот (формулы, ферменты, коферменты, функции)
СИНТЕЗ ГОРМОНОВ ПРОИЗВОДНЫХАМИНОКИСЛОТ (ФОРМУЛЫ, ФЕРМЕНТЫ,
КОФЕРМЕНТЫ, ФУНКЦИИ)
44. Гормоны - производные тирозина
ГОРМОНЫ - ПРОИЗВОДНЫЕ ТИРОЗИНАИз тирозина синтезируются;
гормоны мозгового слоя надпочечников
адреналин и норадреналин
йодсодержащие гормоны щитовидной железы.
1. В ходе синтеза адреналина и норадреналина
тирозин подвергается гидроксилированию,
декарбоксилированию и метилированию с
участием активной формы аминокислоты
метионина.
2. В щитовидной железе происходит синтез
йодсодержащих гормонов трийодтиронина и
тироксина (тетрайодтиронина).
45. Строение и биосинтез йодтиронинов
СТРОЕНИЕ И БИОСИНТЕЗЙОДТИРОНИНОВ
• Стимул – ТТГ гипофиза
• Необходим белок тиреоглобулин (Это гликопротеин, 115
остатков тирозина. Синтезируется в базальной части
клетки. Хранится во внеклеточном коллоиде.
J
ОН
ОН H O
2 2
ОН
J
J
J+
ОН
J
Конденсация
J
J
О
J
ОН
J
46. биосинтез йодтиронинов
БИОСИНТЕЗ ЙОДТИРОНИНОВЭР - эндоплазматический ретикулум;
ДИТ - дийодтиронин;
Тг - тиреоглобулин;
Т3 - трийодтиронин,
Т4 - тироксин.
Тиреоглобулин синтезируется на
рибосомах, далее поступает в комплекс
Гольджи, а затем во внеклеточный
коллоид, где он хранится и где
происходит иодирование остатков
тирозина.
Этапы образование йодтиронинов:
1.
2.
3.
4.
5.
транспорт иода в клетки
щитовидной железы,
окисление йода,
йодирование остатков тирозина,
образование йодтиронинов,
транспорт йодтиронинов в кровь
47. Регуляция синтеза и секреции йодтиронинов
РЕГУЛЯЦИЯ СИНТЕЗА И СЕКРЕЦИИЙОДТИРОНИНОВ
1 - тиреотропин-либерин
стимулирует освобождение
ТТГ;
2 - ТТГ стимулирует синтез и
секрецию йодтиронинов;
3, 4 - иодтиронины тормозят
синтез и секрецию ТТГ
Йодтиронины регулируют
процессы двух типов:
1. рост и дифференцировку
тканей;
2. энергетический обмен.
48. Йодтиронины
ЙОДТИРОНИНЫ1. синтез и секреция регулируется гипоталамогипофизарной системой по принципу обратной
связи
2. стимул для секреции тиреолиберина и
тиреостатина – изменение концентрации
йодтиронинов в крови
3. метаболические эффекты дозозависимые
4. клетки-мишени для прямого действия – печень,
мышцы, костный мозг
49. Заболевания щитовидной железы
ЗАБОЛЕВАНИЯ ЩИТОВИДНОЙ ЖЕЛЕЗЫ1. Гипотиреоз у новорожденных приводит к развитию
2.
3.
кретинизма
–
тяжелое,
необратимое
нарушение
умственного развития. Причина – недостаточность функций
щитовидной железы, заболевания гипофиза, гипоталамуса.
Гипотиреоз у взрослых – микседема. Наблюдается
снижение
основного
обмена,
скорости
гликолиза,
мобилизация гликогена, жиров, уменьшение мышечной
массы, теплопродукции.
Гипертиреоз – повышенная продукция йодтиронинов.
Отмечается увеличение размеров щитовидной железы,
увеличение концентрации Т3, Т4 в 2-5 раз и развитие
тиреотоксикоза. Признаки: увеличение основного обмена,
тахикардия, снижение массы тела (несмотря на повышенный
аппетит), потливость, увеличение температуры тела,
усиление процессов катаболизма, о чем свидетельствует
отрицательный азотистый баланс.
50. Синтез гормонов стероидной природы
СИНТЕЗ ГОРМОНОВСТЕРОИДНОЙ ПРИРОДЫ
51. Основные классы биологически активных стероидных гормонов
ОСНОВНЫЕ КЛАССЫ БИОЛОГИЧЕСКИАКТИВНЫХ СТЕРОИДНЫХ ГОРМОНОВ
1.
2.
3.
4.
5.
Глюкокортикоиды
Минералокортикоиды
Андрогены
Эстрогены
Гестогены
52.
Классификация стероидных гормонов по месту синтеза53.
54. Биосинтез стероидных гормонов
БИОСИНТЕЗ СТЕРОИДНЫХ ГОРМОНОВПечень
Ацетил-КоА
Холестерин
Холестерид
ЛПОНП
Ацил-КоА
Кровь
Эндокринная
железа
ЛПОНП
ЛНП
Цитоплазма
ЛНП
- кора надпочечников
- семенники
- яичники
- плацента
Холестерид
Эстераза
+
АКТГ
Ацил-КоА
+
Холестерин
Белок внутренней
мембраны митохондрий
(С-27) Холестерин-белок
Митохондрия
АКТГ
+
Прегненолон (С-21)
АКТГ
Кортикостероиды
+
Андрогены
Эстрогены
55.
56. Механизм действия гормонов
МЕХАНИЗМДЕЙСТВИЯ
ГОРМОНОВ
57. По степени чувствительности к гормонам клетки-мишени делятся на три группы:
ПО СТЕПЕНИ ЧУВСТВИТЕЛЬНОСТИ КГОРМОНАМ КЛЕТКИ-МИШЕНИ ДЕЛЯТСЯ НА
ТРИ ГРУППЫ:
• Гормонзависимые – дифференцировка, рост и
функционирование зависит от присутствия гормона:
(АКТГ
кора надпочечников,
половые гормоны
половые органы)
• Гормончувствительные – дифференцировка, рост и
функционирование возможны без гормона, но в его
присутствии эти процессы значительно изменяются
(АКТГ
клетки мышц, жировой ткани)
• Гормоннезависимые (гормоннечувствительные) – в
физиологических концентрациях гормон влияния не
оказывает.
(Половые гормоны
клетки мышц
Кортикостероиды
клетки миокарда)
58. Основные свойства рецепторов
ОСНОВНЫЕ СВОЙСТВАРЕЦЕПТОРОВ
1. Высокое сродство рецепторов к связывающему
гормону
2. Высокая избирательность - рецепторы
связывают определенную группу природных и
синтетических гормонов
3. Ограниченная связывающая емкость –
ограничивает взаимодействие клетки с
гормонами в рамках физиологических или
умеренных фармакологических концентраций
4. Специфическая тканевая локализация – отсюда
деление тканей на гормонзависимые,
гормончувствительные и гормоннезависимые.
59.
ВИДЫ РЕЦЕПТОРОВДЛЯ ГОРМОНОВ ПО МЕХАНИЗМУ
ДЕЙСТВИЯ ДЕЛЯТСЯ НА:
Мембранные
(белково-пептидные
гормоны,
катехоламины)
Внутриклеточные
(стероидные и тиреоидные
гормоны)
60.
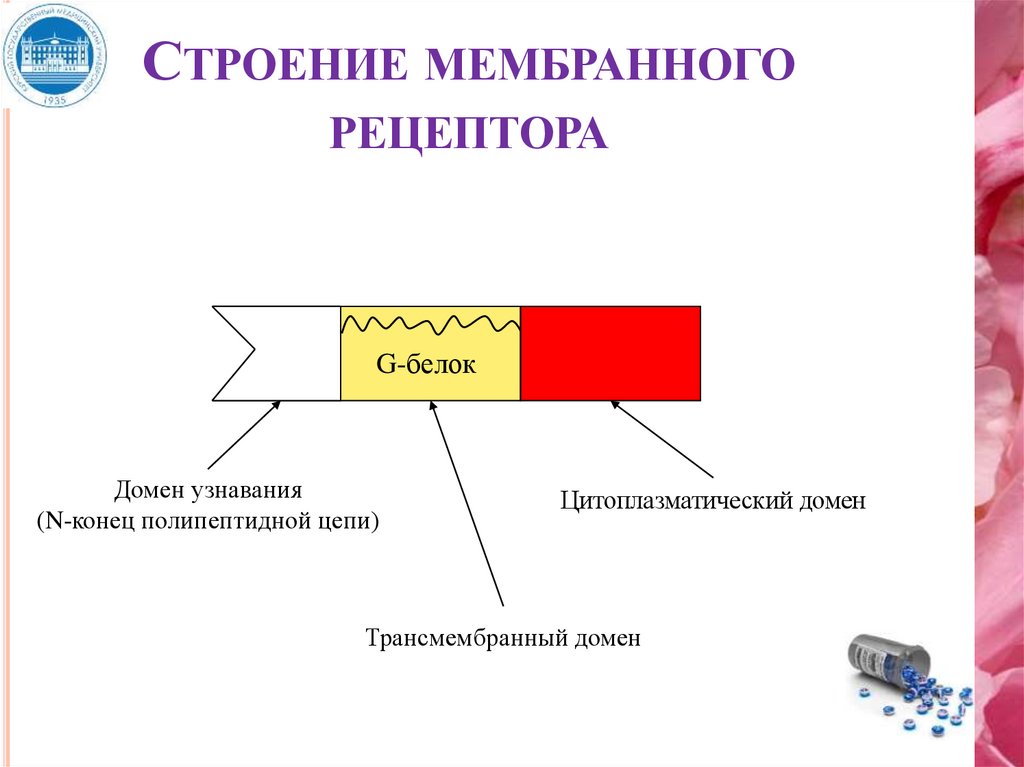
СТРОЕНИЕ МЕМБРАННОГОРЕЦЕПТОРА
G-белок
Домен узнавания
(N-конец полипептидной цепи)
Цитоплазматический домен
Трансмембранный домен
61. Рецепция белково-пептидных гормонов и катехоламинов (мембранная)
РЕЦЕПЦИЯ БЕЛКОВО-ПЕПТИДНЫХ ГОРМОНОВИ КАТЕХОЛАМИНОВ (МЕМБРАННАЯ)
Клеточная
мембрана
Цитоплазма
Ядро
М
ДНК
Рецептор
+
Рецепторная
часть
Э
Предшественник
и-РНК
Медиатор
Гормон
-
Эффекторная часть
Аденилатциклаза
Гуанилатциклаза
Протеаза
Цистерна (Са2+)
АТФ
ГТФ
Белок
Са2+
цАМФ
цГМФ
Пептиды
Са2+
Метаболические
эффекты
62. Строение внутриклеточного рецептора
СТРОЕНИЕ ВНУТРИКЛЕТОЧНОГОРЕЦЕПТОРА
NH2
COOH
С – домен узнавания и
связывания гормона
Вариабельная область связывает
белки и регулирует
транскрипцию
Центральная часть для
связывания ДНК
63. Рецепция стероидных и тиреоидных гормонов (внутриклеточная)
РЕЦЕПЦИЯ СТЕРОИДНЫХ И ТИРЕОИДНЫХ ГОРМОНОВ(ВНУТРИКЛЕТОЧНАЯ)
Клеточная мембрана
Г
Г
+
Цитоплазма
Р1
Р1
Г
Р1
Тепмература
рН
Г
Р2
Ядро
Г
Р2
ДНК
Метаболические
эффекты
и-РНК
64. Передача гормональных сигналов через мембранные рецепторы
ПЕРЕДАЧА ГОРМОНАЛЬНЫХ СИГНАЛОВ ЧЕРЕЗМЕМБРАННЫЕ РЕЦЕПТОРЫ
65. Передача гормональных сигналов через внутриклеточные рецепторы
ПЕРЕДАЧА ГОРМОНАЛЬНЫХ СИГНАЛОВЧЕРЕЗ ВНУТРИКЛЕТОЧНЫЕ РЕЦЕПТОРЫ
66.
Материалы по Гормональной регуляцииобмена углеводов и липидов смотрите в
темы 2.5. «Регуляция углеводного обмена.
Особенности углеводного обмена в органах и
тканях» и
темы 3.4. «Регуляция и патология липидного
обмена»
67.
68.
69.
70.
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯОБМЕННЫХ ПРОЦЕССОВ:
белков,
воды, натрия, калия,
кальция, фосфатов.
71. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНа белков
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯОБМЕНА БЕЛКОВ
72. Гормональная регуляция метаболизма белков
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯМЕТАБОЛИЗМА БЕЛКОВ
Анаболический
эффект
(соматотропина,
инсулина,
глюкокортикоидов,
тестостерона,
эстрогенов,
тироксина в малых
дозах и в детском
возрасте)
катаболическим
эффектам
(глюкокортикоиды,
тироксин в
больших дозах)
динамическое равновесие
синтеза и распада белков
73.
74.
75.
76.
77.
78.
79.
80.
81.
82. Гормональная регуляция белкового обмена (СТГ, инсулин, половые и тиреоидные гормоны, глюкокортикоиды)
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ БЕЛКОВОГООБМЕНА
(СТГ, ИНСУЛИН, ПОЛОВЫЕ И ТИРЕОИДНЫЕ ГОРМОНЫ,
ГЛЮКОКОРТИКОИДЫ)
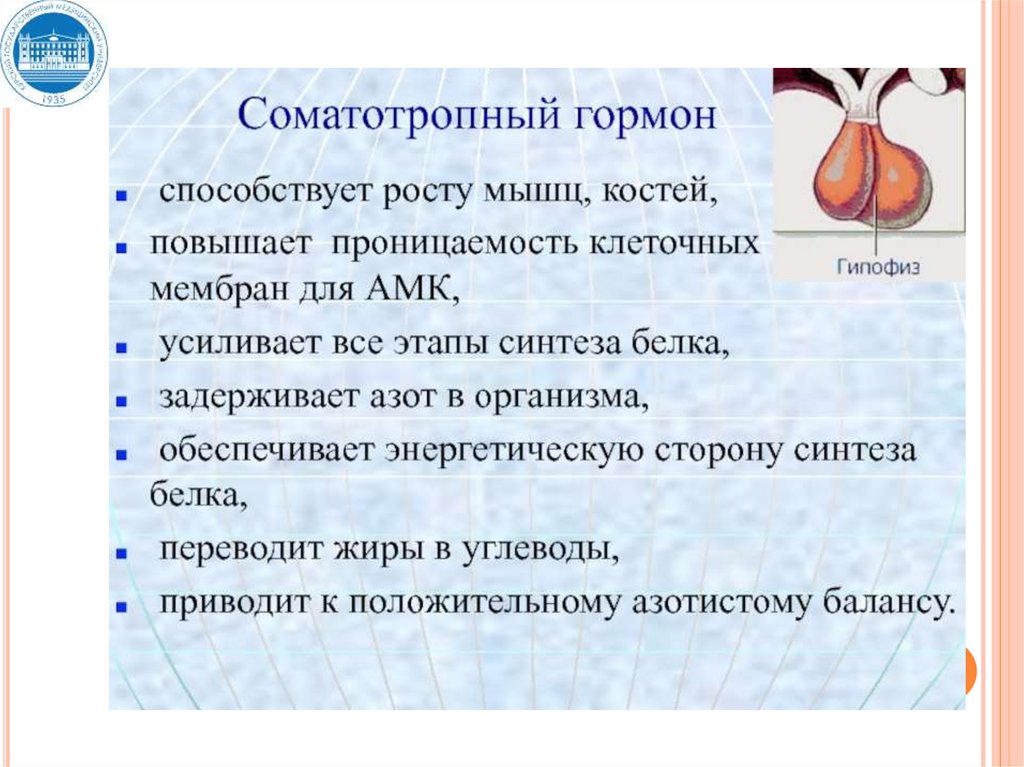
Гормон роста, СТГ – мощный
анаболический гормон
Место синтеза - передняя доля
гипофиза.
СТГ – одноцепочечный пептид,
м.м. 22кД, 191 аминокислота, 2
внутримолекулярных
дисульфидных связи.
СТГ – образуется из неактивного
предшественника – прогормона.
Секреция
СТГ
носит
пульсирующий
характер
с
интервалом в 20-30 мин. Один из
самых
больших
пиков
наблюдается
вскоре
после
засыпания.
Стресс,
физические
упражнения,
гипогликемия,
голодание,
белковая
пища
–
приводит к повышению уровня
СТГ даже у нерастущих взрослых
людей.
Синтез и секреция СТГ
регулируется гипоталамусом:
1. соматолиберином (+);
2. соматостатином (-).
83. Рецепторы для СТГ расположены в плазматической мембране клеток
РЕЦЕПТОРЫ ДЛЯ СТГ РАСПОЛОЖЕНЫ ВПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ КЛЕТОК
1. Гормозависимые ткани:
скелетные мышцы,
хрящи,
кости,
соединительная ткань.
2. Гормончувствительные
ткани:
печень,
жировая ткань,
мозг,
легкие,
поджелудочная железа,
кишечник,
сердце,
почки,
лимфоциты.
84. Действие СТГ на рост скелета и мягких тканей опосредуется соматомединами
ДЕЙСТВИЕ СТГ НА РОСТ СКЕЛЕТА ИМЯГКИХ ТКАНЕЙ ОПОСРЕДУЕТСЯ
СОМАТОМЕДИНАМИ
Синтезируются
различными
Это соединения с
тканями, в основном
инсулиноподобной
активностью и мощным печенью.
Соматомедин С
ростостимулирующим
действием, поэтому они обладает также
регуляторными
называются
свойствами
инсулиноподобные
(активирует
факторы роста
1. ИФР – 1 – соматомедин выработку
соматостатина и
С
ингибирует
2. ИФР – 2 – соматомедин соматолиберина)
А
85. Биологическое действие СТГ
БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ СТГ1. Начальные эффекты
(мышцы, хрящи, кости, соединительная ткань)
увеличение проницаемости клеточных мембран для
аминокислот, глюкозы, Са2+
фосфорилирование белков
изменение активности
ряда ферментов
2. Ранние эффекты
усиление транскрипции (р-РНК, м-РНК, т-РНК).
ускорение трансляции (синтез ферментов и структурных
белков)
3. Поздние эффекты
изменение репликации.
усиление пролиферации, дифференцировки тканей
Конечный физиологический эффект – увеличение
линейного роста, мышечной массы за счет ускорения
дифференцировки и пролиферации клеток
86.
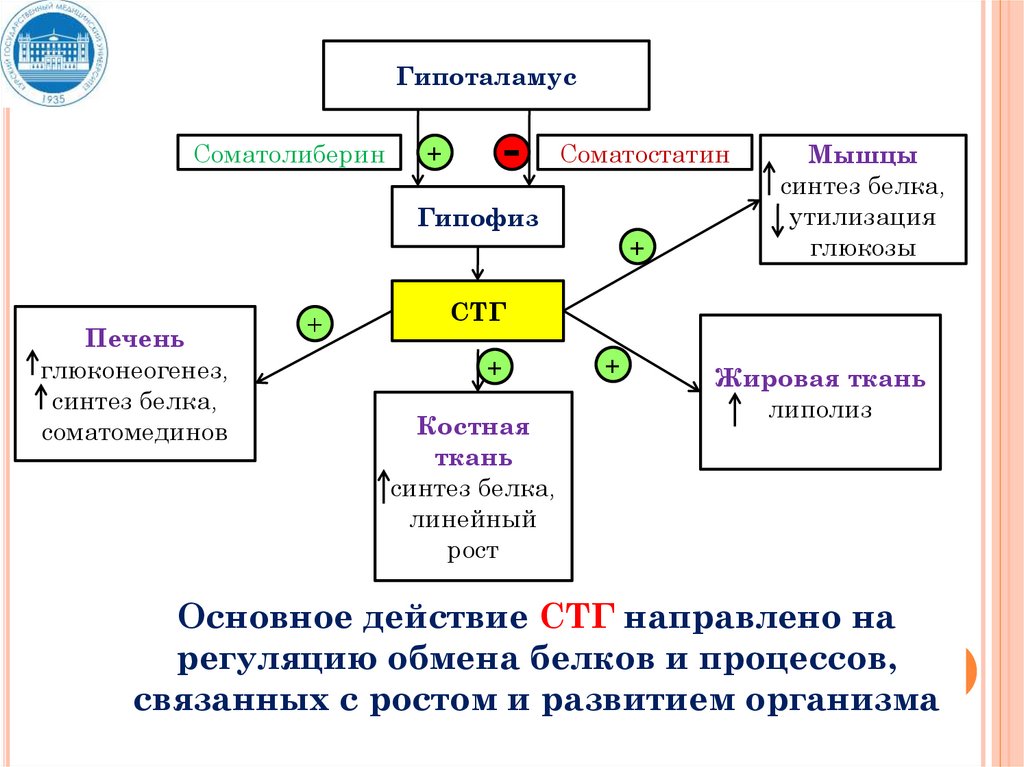
ГипоталамусСоматолиберин
+
-
Соматостатин
Гипофиз
Печень
глюконеогенез,
синтез белка,
соматомединов
+
+
Мышцы
синтез белка,
утилизация
глюкозы
СТГ
+
Костная
ткань
синтез белка,
линейный
рост
+
Жировая ткань
липолиз
Основное действие СТГ направлено на
регуляцию обмена белков и процессов,
связанных с ростом и развитием организма
87.
88. Патология синтеза и секреции СТГ
ПАТОЛОГИЯ СИНТЕЗА И СЕКРЕЦИИСТГ
1. Гипофункция
с
рождения
гипофизарная
карликовость (дефицит СТГ из-за мутации гена
гормона роста). Нарушения роста сочетается с
другими эндокринными нарушениями.
2. Гиперсекреция СТГ у детей - гигантизм (у подростков
с
незакончившимся
процессом
окостенения
эпифизарных хрящей, продолжается рост длинных
костей, происходит их удлинение, увеличение мягких
тканей и органов сравнительно пропорционально).
3. Гиперсекреция
СТГ во взрослом состоянии –
акромегалия (обычно возникает в результате
гормонпродуцирующей
опухоли
гипофиза,
диспропорционально увеличиваются размеры кистей,
стоп, черепа, внутренних органов)
89.
90. Анаболические эффекты инсулина
АНАБОЛИЧЕСКИЕ ЭФФЕКТЫИНСУЛИНА
1. регуляция транспорта
в клетки глюкозы,
аминокислот, ионов.
2. влияние на процессы
репликации и
транскрипции
(регуляция
клеточной
дифференцировки,
пролиферации и
трансформации
клеток).
91. Анаболические эффекты инсулина
АНАБОЛИЧЕСКИЕ ЭФФЕКТЫИНСУЛИНА
1. Начальные эффекты (в
течении
нескольких
секунд-минут,
изменение
трансмембранного
транспорта,
фосфорилирование
и
дефосфорилирование
белков,
активация
и
ингибирование
ферментов)
2. Ранние,
поздние
эффекты
(через
несколько часов – суток,
активация синтеза РНК,
ДНК, белков, усиление
пролиферации,
дифференцировки,
усиление синтеза СТГ)
Инсулин обеспечивает
анаболизм белков
энергией за счет
регуляции углеводного
обмена.
Гормончувствительны
е клетки-мишени для
инсулина (наибольший
анаболический эффект
для обмена белков):
мышцы,
печень,
почки,
соединительная
ткань
92.
93. Йодтиронины
ЙОДТИРОНИНЫ1. Синтез и секреция
йодтиронинов (Т3 и Т4)
регулируется гипоталамогипофизарной системой по
принципу обратной связи.
2. Стимул для секреции
тиреолиберина и тиреостатина
гипоталамуса – изменение
концентрации йодтиронинов в
крови.
3. Тиреолиберин и тиреостатин
контролируют активацию и
ингибирование синтеза ТТГ в
передней доле гипофиза.
4. Синтез и секреция
йодтиронинов регулируется
непосредственно ТТГ.
5. Клетки-мишени для прямого
действия йодтиронинов –
94.
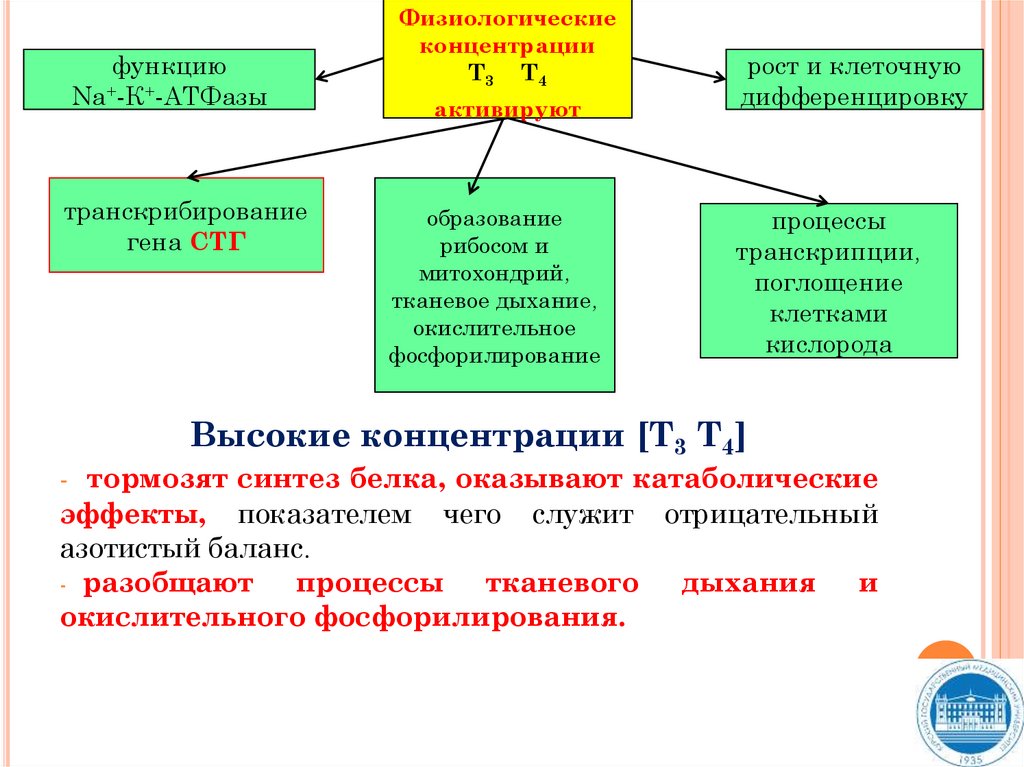
функциюNa+-К+-АТФазы
транскрибирование
гена СТГ
Физиологические
концентрации
Т3 Т4
активируют
образование
рибосом и
митохондрий,
тканевое дыхание,
окислительное
фосфорилирование
рост и клеточную
дифференцировку
процессы
транскрипции,
поглощение
клетками
кислорода
Высокие концентрации [Т3 Т4]
- тормозят синтез белка, оказывают катаболические
эффекты, показателем чего служит отрицательный
азотистый баланс.
- разобщают
процессы тканевого дыхания и
окислительного фосфорилирования.
95.
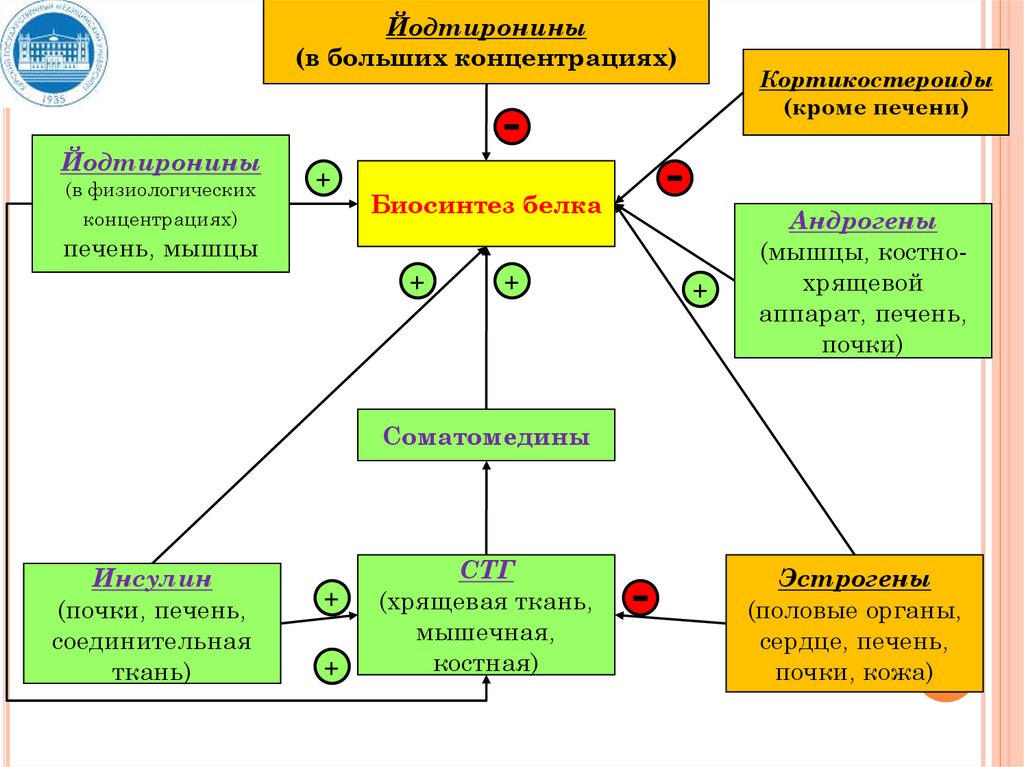
Йодтиронины(в больших концентрациях)
Йодтиронины
(в физиологических
концентрациях)
+
Кортикостероиды
(кроме печени)
-
Биосинтез белка
печень, мышцы
+
+
+
Андрогены
(мышцы, костнохрящевой
аппарат, печень,
почки)
Соматомедины
Инсулин
(почки, печень,
соединительная
ткань)
+
+
СТГ
(хрящевая ткань,
мышечная,
костная)
-
Эстрогены
(половые органы,
сердце, печень,
почки, кожа)
96. Заболевания щитовидной железы
ЗАБОЛЕВАНИЯ ЩИТОВИДНОЙЖЕЛЕЗЫ
1. Гипотиреоз у новорожденных приводит к развитию
кретинизма – тяжелое, необратимое нарушение
умственного развития. Причина – недостаточность
функций щитовидной железы, заболевания гипофиза,
гипоталамуса.
2. Гипотиреоз у взрослых – микседема. Причины те
же, но во взрослом состоянии, в большинстве случаев удаление щитовидной железы. Наблюдается
снижение основного обмена, скорости гликолиза,
мобилизации
гликогена,
жиров,
уменьшение
мышечной массы, теплопродукции, апатия, депрессия.
3. Гипертиреоз
–
повышенная
продукция
йодтиронинов. Отмечается увеличение концентрации
Т3, Т4 в 2-5 раз и развитие тиреотоксикоза.
Признаки: увеличение основного обмена, тахикардия,
снижение массы тела (несмотря на повышенный
аппетит), потливость, увеличение температуры тела,
усиление процессов катаболизма, отрицательный
азотистый баланс.
97. Заболевания щитовидной железы
ЗАБОЛЕВАНИЯ ЩИТОВИДНОЙ ЖЕЛЕЗЫ98.
99. Половые гормоны
ПОЛОВЫЕ ГОРМОНЫАндрогены обладают
выраженным анаболическим
эффектом в печени, почках,
костно-хрящевом аппарате,
покровных тканях, мужских
половых органах, особенно в
период полового созревания
1. Активация транскрипции рРНК
2. Активация трансляции
Эстрогены оказывают более слабое анаболическае действие.
Усиливают синтез белка:
1. В женских половых органах
2. Печени, почках, сердце, коже
3. На костно-хрящевой аппарат оказывают ингибирующее действие,
что способствует остановке роста в период полового созревания
(ингибируются биосинтез белка в мышцах, хрящах)
100.
101.
102. Влияние глюкокортикоидов на обмен белков
ВЛИЯНИЕ ГЛЮКОКОРТИКОИДОВНА ОБМЕН БЕЛКОВ
Проявляется двояко:
1. В печени кортизол в основном оказывает
анаболический эффект (стимулирует синтез белков
и нуклеиновых кислот)
2. В лимфоидной, мышечной и жировой тканях, коже
и костях, кортизол тормозит синтез белков, РНК и
ДНК и усиливает катаболизм РНК и белков.
3. При высокой концентрации (гиперсекреция или
введение как лекарственных препаратов)
глюкокортикоиды подавляют иммунные реакции,
вызывая гибель лимфоцитов и инволюцию
лимфатической ткани, снижая число
циркулирующих лейкоцитов. Подавляют
воспалительную реакцию (индуцируют синтез
липокортинов, которые ингибируют фосфолипазу
А2 снижая т.о. синтез медиаторов воспаления –
простагландинов и лейкотриенов).
Высокая
концентрация
глюкокортикоидов
вызывает
торможение
роста
и
деления
фибробластов, а также синтез коллагена и
фибронектина
Для гиперсекреции глюкокортикоидов типичны:
истончение
кожи,
плохое
заживление
ран,
мышечная слабость и атрофия мышц.
103. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНа воды, натрия, калия
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯОБМЕНА ВОДЫ, НАТРИЯ, КАЛИЯ
104. ПАРАМЕТРЫ ВОДНО-СОЛЕВОГО ГОМЕОСТАЗА
ПАРАМЕТРЫ ВОДНОСОЛЕВОГО ГОМЕОСТАЗАV
внеклеточно
й жидкости
рН
Осмотическое
давление
V
внутриклеточной
жидкости
Изменение этих параметров
может привести к:
Ацидозу
Алкалозу
Отекам
Изменению АД
Дегидратации
105. ГОРМОНЫ, УЧАСТВУЮЩИЕ В РЕГУЛЯЦИИ ВОДНО-СОЛЕВОГО ГОМЕОСТАЗА
РЕГУЛЯЦИИВОДНО-СОЛЕВОГО
ГОМЕОСТАЗА
1.
АДГ
(АНТИДИУРЕТИЧЕС
КИЙ ГОРМОН,
ВАЗОПРЕССИН),
2.
3.
АЛЬДОСТЕР
ОН
Предсердный
натриуретически
й фактор (ПНФ).
106. Регуляция водно-солевого гомеостаза
РЕГУЛЯЦИЯ ВОДНО-СОЛЕВОГОГОМЕОСТАЗА
107. ХАРАКТЕРИСТИКА АДГ
АДГ – пептид, м.м. 1100 Д, содержит 9 АМК,одна дисульфидная связь
Синтез и секреция АДГ
Гипоталамус:
Синтез препрогормона
процессинг
Прогормон (аппарат Гольджи)
нейрофизин
(транспортный
белок)
Транспорт в составе
нейросекреторных гранул
Нейрогипофиз:
хранится зрелый
гормон
108.
109. СТИМУЛ ДЛЯ СЕКРЕЦИИ АДГ
[Na+]2.
осмотического давления
внеклеточной
жидкости
1.
Осморецепторы гипоталамуса
регистрируют :
Р осмотического
нервные
импульсы передаются в нейрогипофиз
высвобождение АДГ.
110. Секреция и механизм действия антидиуретического гормона
СЕКРЕЦИЯ И МЕХАНИЗМ ДЕЙСТВИЯАНТИДИУРЕТИЧЕСКОГО ГОРМОНА
А: 1 - супраоптический нейрон; 2 паравентрикулярный нейрон; 3 - передняя доля
гипофиза; 4 - задняя доля гипофиза; 5 - АДГнейрофизин;
Б: 1 - АДГ связывается с мембранным рецептором V2,
вызывая активацию аденилатциклазы (АЦ) и, как
следствие, образование цАМФ; 2 - цАМФ активирует
протеинкиназу, фосфорилирующую белки; 3 фосфорилированные белки индуцируют
транскрипцию гена белка аквапорина; 4 - аквапорин
встраивается в мембрану клетки почечного канальца
Для АДГ существует два типа рецепторов - V1 и
V2. Рецептор V2 обнаружен только на поверхности
эпителиальных клеток почек. Связывание АДГ с
V2 сопряжено с аденилатциклазной системой и
стимулирует активацию протеинкиназы (ПКА),
которая фосфорилирует белки, стимулирующие
экспрессию гена мембранного белка - аквапорина-2.
Аквапорин-2 перемещается к апикальной мембране,
встраивается в нее и образует водные каналы, через
которые молекулы воды свободно диффундируют в
клетки почечных канальцев, а затем поступают в
интерстициальное пространство. В результате
происходит реабсорбция воды из почечных канальцев.
Рецепторы типа V локализованы в мембранах гладких
мышц. Взаимодействие АДГ с рецептором V1 приводит
111. МЕХАНИЗМ ДЕЙСТВИЯ АДГ
Для АДГ существует 2 типа рецепторов:V 1 и V2
1) V2 – опосредуют главный
физиологический эффект гормона,
локализованы на базолатеральной
мембране клеток собирательных
трубочек и дистальных канальцев
почек.
2) V1 – локализованы в гладкомышечных
клетках сосудов, связывание АДГ с V1
приводит к сокращению
гладкомышечного слоя сосудов.
3) Сродство АДГ к V2 выше, чем к V1.
112. ЭТАПЫ ДЕЙСТВИЯ АДГ НА ЭПИТЕЛИАЛЬНЫЕ КЛЕТКИ ДИСТАЛЬНЫХ КАНАЛЬЦЕВ И СОБИРАТЕЛЬНЫХ ТРУБОЧЕК ПОЧЕК
Н2ОКровь
АЦ
АЦ
R
АТФ
Протеин
киназа
неакт.
цАМФ
+
1
ПКА
акт.
2
Белок-ОН
ДНК
4
А
А
А
Н2О
АДГ
Белок Р
мРНК
3
Аквапорин- 2
Моча
113. НЕСАХАРНЫЙ ДИАБЕТ
Причиной является дефицит АДГ1) генетические дефекты синтеза препро-АДГ в гипоталамусе;
2) дефекты процессинга и транспорта про-АДГ;
3) повреждения гипоталамуса или нейрогипофиза (черепно-мозговая
травма, опухоль, ишемия);
4) нейрогенный несахарный диабет (мутация гена R – V2 к АДГ), в
результате почки не реагируют на АДГ.
Клиника:
гипотоническая полиурия,
выделение большого количества мочи, низкой плотности (до 20 л/сут,
плотность L 1,010).
114. АЛЬДОСТЕРОН
Синтез и секрециянепосредственно
стимулируются:
понижением [Na+] и
повышением [K+] в
плазме крови
2. простагландинами, АКТГ
3. ренин-ангиотензиновой
Наиболее активный
системой
1.
Клетки-мишени:
эпителиальные клетки;
почечных канальцев.
минералокортикоид,
синтезируется в коре
надпочечников
115. МЕХАНИЗМ ДЕЙСТВИЯ АЛЬДОСТЕРОНА
Альдостерон, взаимодействуя с внутриклеточнымирецепторами, стимулирует синтез белков. Эти белки
могут быть:
1 - компонентами натриевых каналов и увеличивать
реабсорбцию Na+ из мочи;
2 - ферментами ЦТК, активность которых
обеспечивает продукцию АТР;
3 - Na+, K+ - АТФазой, насосом, который
поддерживает низкую внутриклеточную
концентрацию ионов натрия и высокую
концентрацию ионов калия
высвобождения ренина.
Этому способствует также снижение импульсации от
барорецепторов предсердий и артерий в результате
уменьшения внутрисосудистого объема жидкости. В
результате увеличивается образование
ангиотензина II и соответственно повышается в
крови концентрация альдостерона, вызывая
задержку ионов натрия. Это служит сигналом для
осморецепторов гипоталамуса и секреции из
нервных окончаний передней доли гипофиза АДГ,
который стимулирует реабсорбцию воды из
собирательных трубочек. Ангиотензин II, оказывая
сильное сосудосуживающее действие, повышает
артериальное давление, а также усиливает жажду.
Поступающая с питьем вода в большей мере, чем это
116. СУММАРНЫЙ БИОЛОГИЧЕСКИЙ ЭФФЕКТ АЛЬДОСТЕРОНА
1) Увеличениереабсорбции ионов
Na+ в канальцах
нефронов
2) Задержка NaCl в
организме и
возрастание
+
117. РОЛЬ СИСТЕМЫ РЕНИН-АНГИОТЕНЗИН-АЛЬДОСТЕРОН В РЕГУЛЯЦИИ ВОДНО-СОЛЕВОГО ОБМЕНА
V крови и межклеточнойжидкости.
АД
Глюкокортикоиды,
эстрогены
Перфузионное давление в
приносящей артерии
клубочка
1
Ангиотензиноген
(α2-глобулин, 400АМК)
Секреция ренина
+
Ангиотензин I
(карбоксипептидилпептидаза)
-
2
Ангиотензин II
Сужение сосудов
Повышение АД
4
Альдостерон
3
Реабсорбция Na+
Экскреция К+
Жажда
Повышение
осмотического
давления
АДГ
Реабсорбция
воды
118. ГИПЕРАЛЬДОСТЕРОНИЗМ
ГИПЕРАЛЬДОСТЕРОНИЗМ
Заболевание вызванное гиперсекрецией
альдостерона надпочечниками.
Причина первичного
гиперальдостеронизма (синдром Конна) –
аденома надпочечников, диффузная
гипертрофия клеток клубочковой зоны,
вырабатывающих альдостерон.
Избыток А приводит к увелечению
реабсорбции Na+ , усиливает секрецию
АДГ и вызывает задержку воды.
Усиливается выведение К+, Mg2+, H+.
Снижен уровень ренина.
Клиника:
гипернатриемия,
гипертония,
гиперволемия,
отеки,
гипокалиемия,
дефицит Mg2+,
метаболический алкалоз.
119. ВТОРИЧНЫЙ ГИПЕРАЛЬДОСТЕРОНИЗМ
ВТОРИЧНЫЙГИПЕРАЛЬДОСТЕРОНИЗ
М
Встречается чаще, чем
первичный.
Причины: сердечная
недостаточность, хронические
заболевания почек, опухоли
секретирующие ренин.
Клиника:
Повышен уровень ренина и
ангиотензина II,что стимулирует
продукцию альдостерона.
Клинические симптомы менее
выражены, чем при первичном
альдостеронизме.
120.
121. ХАРАКТЕРИСТИКА ПРЕДСЕРДНОГО НАТРИУРЕТИЧЕСКОГО ФАКТОРА (ПНФ)
Физиологический антагонист
ангиотензина II.
Этот пептид, 28
АМК, с одним
дисульфидным
мостиком.
Синтез:
кардиомиоциты
предсердий.
122. СТИМУЛЫ ДЛЯ СЕКРЕЦИИ ПРЕДСЕРДНОГО НАТРИУРЕТИЧЕСКОГО ФАКТОРА (ПНФ)
АДОсмолярности
плазмы
Уровня катехоламинов
и глюкокортикоидов в
крови
частоты
сердцебиений
123. ОСНОВНЫЕ КЛЕТКИ-МИШЕНИ ПНФ:
1.почки,2.периферические артерии.
В ПОЧКАХ ПНФ СТИМУЛИРУЕТ:
1. расширение приносящих
артериол.
2. усиление почечного кровотока.
3. увеличение скорости
и экскреции
ионов
Вфильтрации
ПЕРИФЕРИЧЕСКИХ
АРТЕРИОЛАХ
+
Na
ПНФ:
1. снижает тонус гладких мышц.
2. расширяет артериолы.
124. Эффекты действия ПНФ
ЭФФЕКТЫ ДЕЙСТВИЯ ПНФ1 - ингибирует выделение
ренина;
2 - ингибирует секрецию
альдостерона;
3 - ингибирует секрецию АДГ;
4 - вызывает релаксацию
сосудов
связывания ПНФ с
рецептором
гуанилатциклазная
активность рецептора
возрастает и происходит
образование из ГТФ
циклического ГМФ. В
результате действия ПНФ
ингибируется образование и
секреция ренина и
125. Гормональные нарущения водно-солевого обмена
ГОРМОНАЛЬНЫЕ НАРУЩЕНИЯВОДНО-СОЛЕВОГО ОБМЕНА
126.
127. Гормональные нарушения обмена натрия и калия
ГОРМОНАЛЬНЫЕ НАРУШЕНИЯ ОБМЕНАНАТРИЯ И КАЛИЯ
128.
129.
130. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНа кальция и ФОСФАТОВ
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯОБМЕНА КАЛЬЦИЯ И ФОСФАТОВ
131. РОЛЬ ИОНОВ КАЛЬЦИЯ В ОБМЕННЫХ ПРОЦЕССАХ:
1.2.
3.
4.
5.
6.
структурный компонент костной ткани;
играет ключевую роль в мышечном
сокращении;
увеличивает проницаемость мембран
клеток для К+;
влияет на работу ионных насосов;
способствует секреции гормонов;
участвует в каскадном механизме
свертывании крови;
132.
В организме взрослогочеловека содержится до
1,5 кг Ca2+.
Пределы колебаний Ca2+ в
крови: 2,25 – 2,75 ммоль/л
(жесткий контроль
гомеостаза)
Основное депо Ca2+ -
кости (99%)
133.
134.
135. ХАРАКТЕРИСТИКА ПАРАТГОРМОНА
ПТГ – одноцепочечный полипептид, 84АМК. Действие ПТГ:
1. Повышение [Ca2+];
2. Снижение [фосфатов] в плазме крови;
Синтезируется в паращитовидных
железах в виде предшественника –
препрогормона.
Секреция ПТГ регулируется уровнем
ионов Ca2+ в крови.
Гормон секретируется в ответ на
снижение [Ca2+].
Органы мишени: кости, почки.
136. ЭФФЕКТЫ ПАРАТГОРМОНА
ПТГ[Ca2+] в плазме крови
Прямое
воздействие
+
Опосредованное
воздействие
+
+
Кости
(мобилизация
Са2+ и фосфатов
во внеклеточную
жидкость)
Почки ( реабсорбции
Ca2+ в дистальных
канальцах)
Стимулирует синтез
кальцитриола
137. БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ПАРАТГОРМОНА
1 - ПТГ стимулируетмобилизацию кальция из
кости;
2 - ПТГ стимулирует
реабсорбцию ионов кальция в
дистальных канальцах почек;
3 - ПТГ активирует
образование 1,25(OH)2D3 в
почках, что приводит к
стимуляции всасывания Са2+ в
кишечнике в продуктах
питания, но большая часть
витамина, используемого в
синтезе кальцитриола,
образуется в коже из 7дегидрохолестерола в ходе
неферментативной реакции
под действием
ультрафиолетового света.
Образование кальцитриола из
витамина D3 начинается в
печени и заканчивается в
почках.
138.
139. Схема синтеза кальцитриола
СХЕМА СИНТЕЗА КАЛЬЦИТРИОЛАКожа: Провитамин Д3 (7-дегидрохолестерол)
УФ
Витамин Д3 (холекальциферол)
(связывается с белком-транскальциферином,
альбумином и поступает в кровь, далее в печень)
25-гидроксилаза
Печень: образование 25 (ОН) Д3 (кальцидиол)
1α-гидроксилаза
+
паратгормон
Почки: образование 1,25 (ОН)2 Д3 (кальцитриол)
140. Обмен витамина D (D2 – эрго -; D3 – холекальциферол)
ОБМЕН ВИТАМИНА D(D2 – ЭРГО -; D3 – ХОЛЕКАЛЬЦИФЕРОЛ)
In vivo
Пища (молочные
продукты, печень)
Кожа
D2(
Ультрафиолетовое
облучение
Холестерол
3)
Кишечник
Кровь
D2(
3)
Трансферрин
(Транскальциферри
н)
141. Обмен витамина D
ОБМЕН ВИТАМИНА DD2(
3)
Гидроксилирование в
25 положении –
25-окси
D
Печень
(кальцидиол
)
Кровь
25-окси D
(кальцидиол)
Трансферрин
(Транскальциферри
142. Витамин D
ВИТАМИН DПочки
Гидроксилирование
в 24 положении при
повышенном или
нормальном
содержании кальция в
крови
24,25диокси D
25окси
D2(3)
Гидроксилирование
в 1 положенииактивация
паратгормоном
(при снижении уровня в
крови кальция)
1α-гидроксилазы
1,25-диокси D(КАЛЬЦИТРИО
Л)
Клетки-мишени
1.Усиление реабсорбции
кальция в почках (активация
синтеза щелочной фосфатазы и
Са2+-зависимой АТФ-азы)
2.Усиление всасывания из
кишечника кальция путем
активации синтеза
кальцийсвязывающих белков.
1.Усиление реабсорбции кальция в
почках (активация синтеза щелочной
фосфатазы и Са2+-зависимой АТФ-азы)
2.Усиление всасывания из кишечника
кальция путем активации синтеза
кальцийсвязывающих белков.
3.Деминерализация костей
143. ЭФФЕКТЫ КАЛЬЦИТРИОЛА [1,25 (ОН)2 Д3]
Клеткикишечника:
Индуцируют
синтез Ca2+ переносящих
белков, которые
обеспечивают
всасывание Са2+
и фосфатов из
полости
кишечника в
эпителиальную
клетку
В почках:
Стимулирует
реабсорбцию Са2+
и фосфатов
При низкой [Са2+]
способствует
мобилизации Ca2+
из костной ткани
144. ХАРАКТЕРИСТИКА КАЛЬЦИТОНИНА
Кальцитонин – полипептид, 32 АМК.Секретируется:
1. парафолликулярными К – клетками щитовидной
железы
2. С – клетками паращитовидных желез.
Секреция кальцитонина в крови возрастает при
увеличении [Ca2+] и уменьшается при уменьшении
[Ca2+]
145. Эффекты кальцитонина
ЭФФЕКТЫ КАЛЬЦИТОНИНАИнгибирует высвобождение Са2+ из
костной ткани;
2.
Стимулирует экскрецию Са2+ и
фосфатов с мочой;
3.
Снижение содержания кальция в крови.
1.
146.
147. Гормональные нарушения обмена кальция
ГОРМОНАЛЬНЫЕ НАРУШЕНИЯ ОБМЕНАКАЛЬЦИЯ