
, относятся к рецепторам растяжения, но")
 иннервируют интрафузальные мышечные веретёна. 2 типа волокон: – Аγ-волокна с")











")













")



 рефлексы спинного мозга")


























, участвующих в регуляции двигательной активности:")























 – развивается децеребрационная регидность, хотя имеются")







")















 и тормозящих (-) зон РФ и ее связи с корой и подкоркой")





































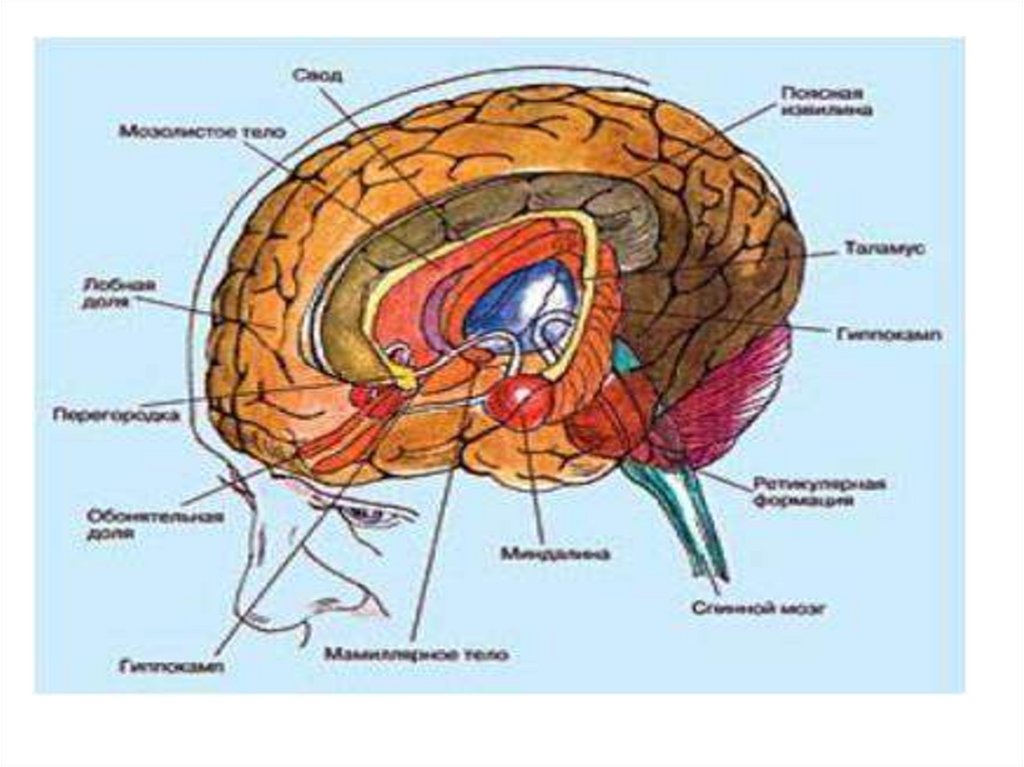
 – темно-серая область, старый")
 – к нему относится клочково-узелковая доля, имеет наиболее выраженные связи с")





 – состоит из параллельных волокон, разветвлений дендритов")
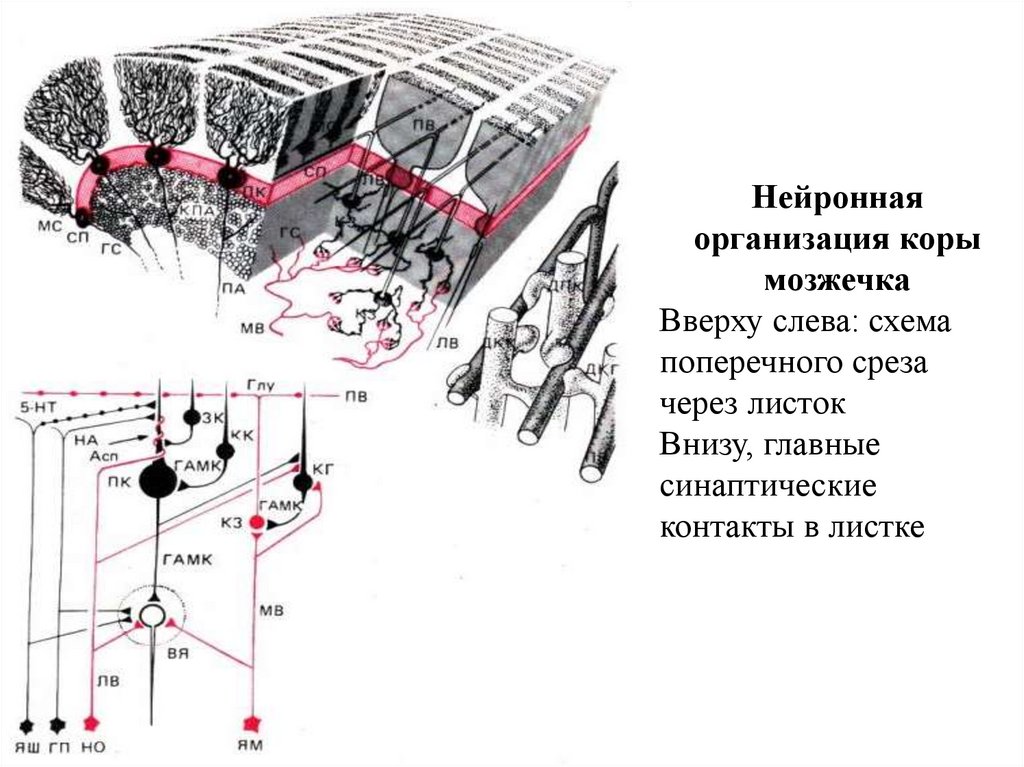

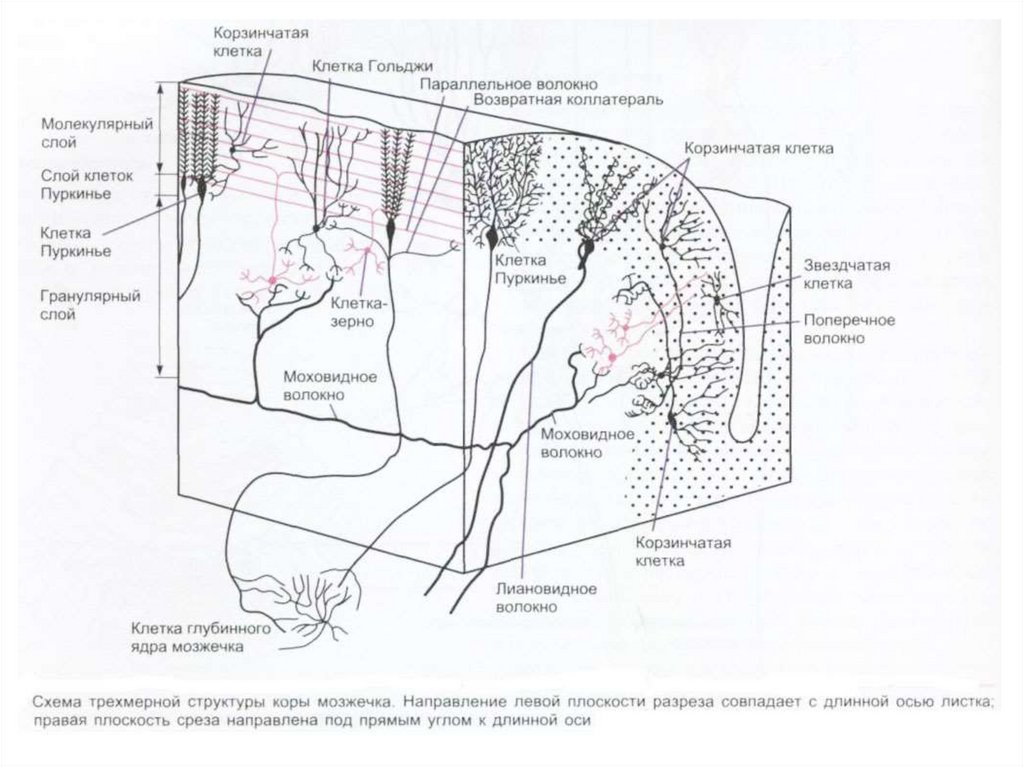
 коры мозжечка, сферическая форма (60×30")


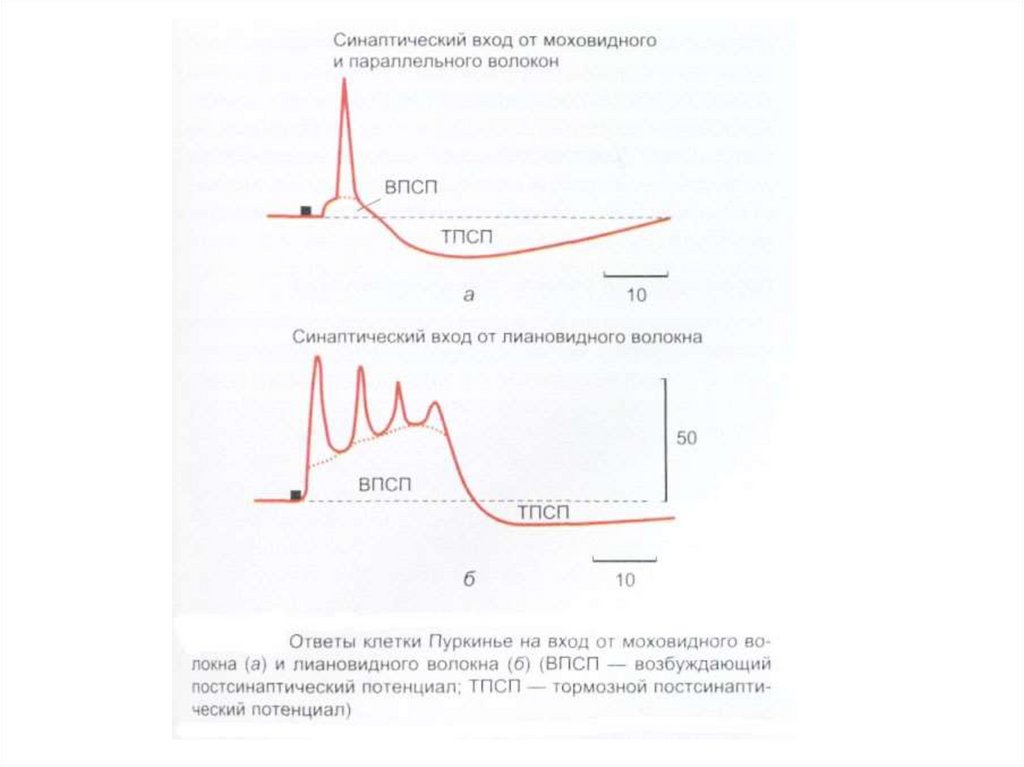

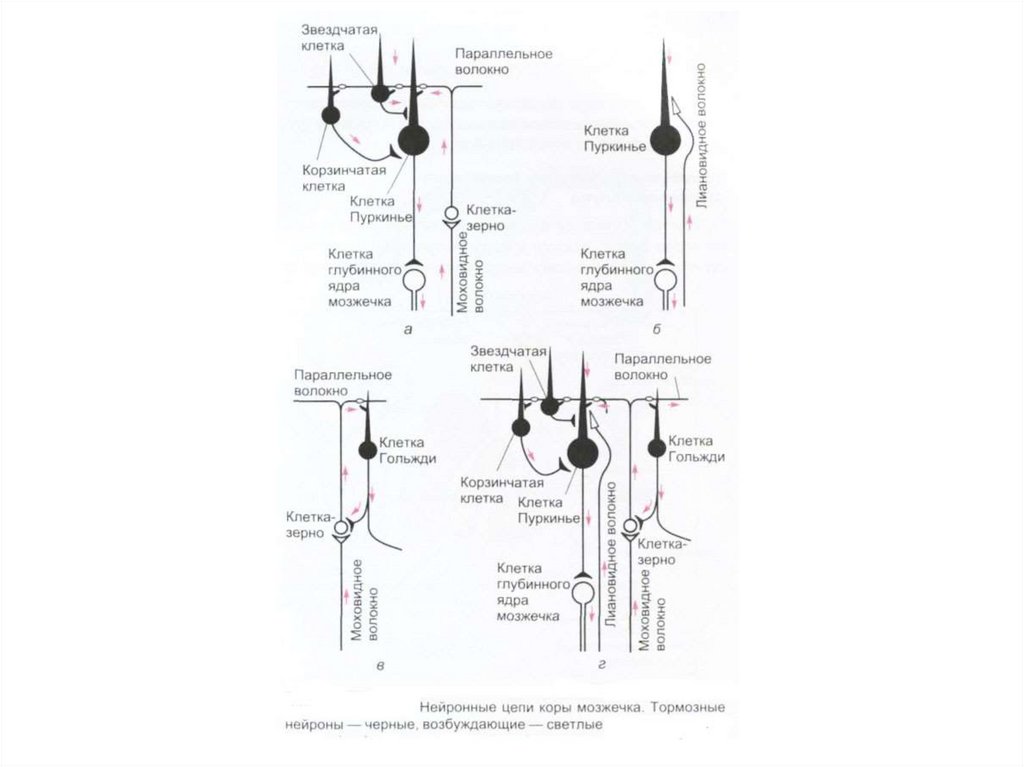





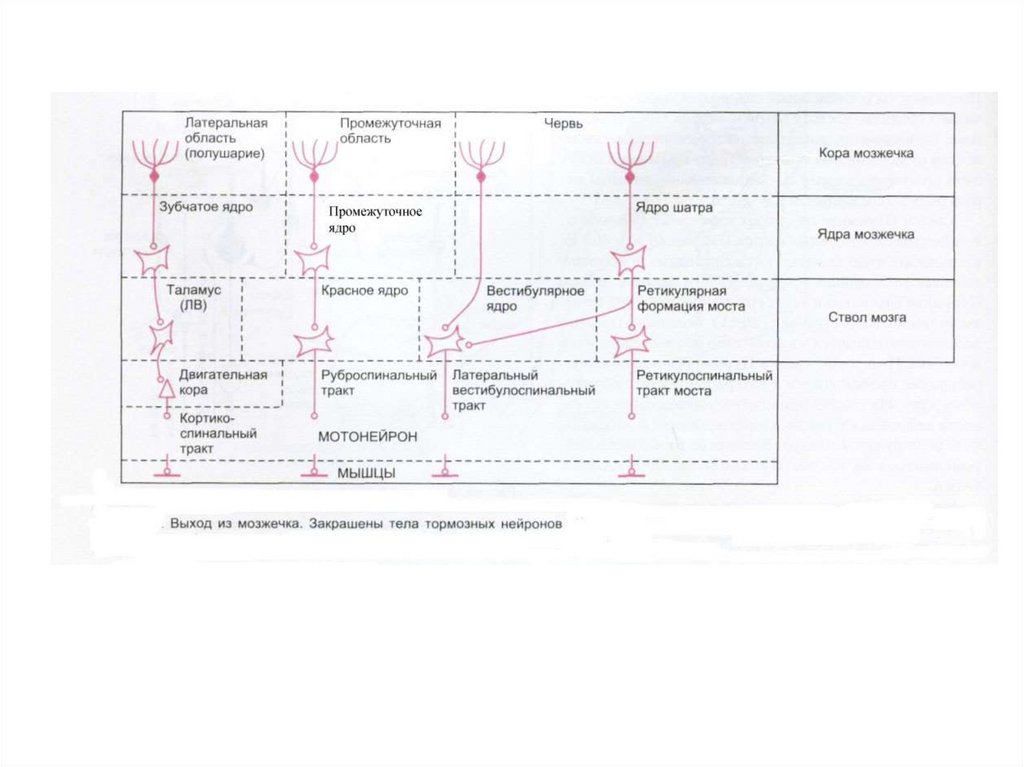





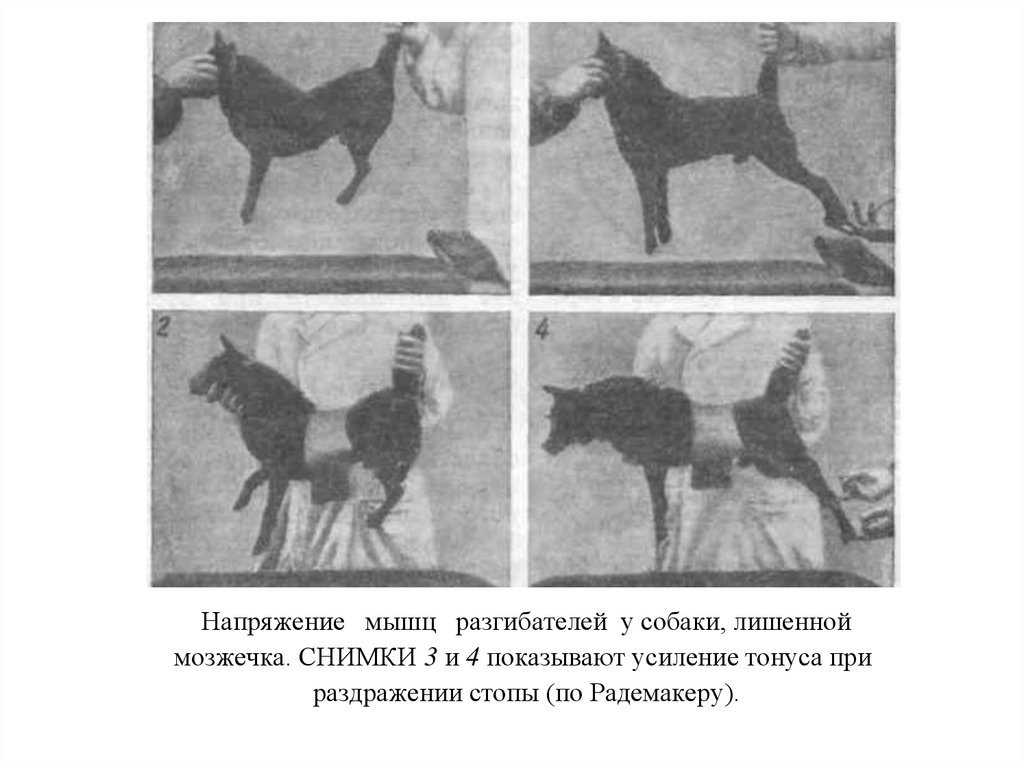

 – в 1891г. показал, что основным комплексом двигательных нарушений мозжечкового происхождения")



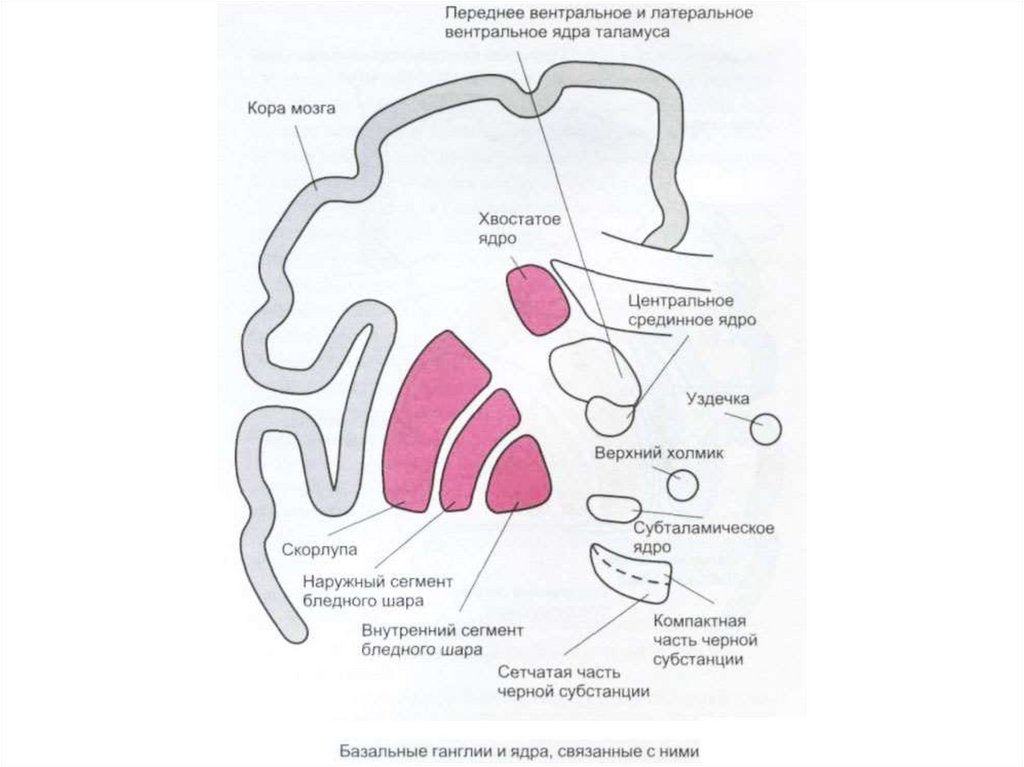

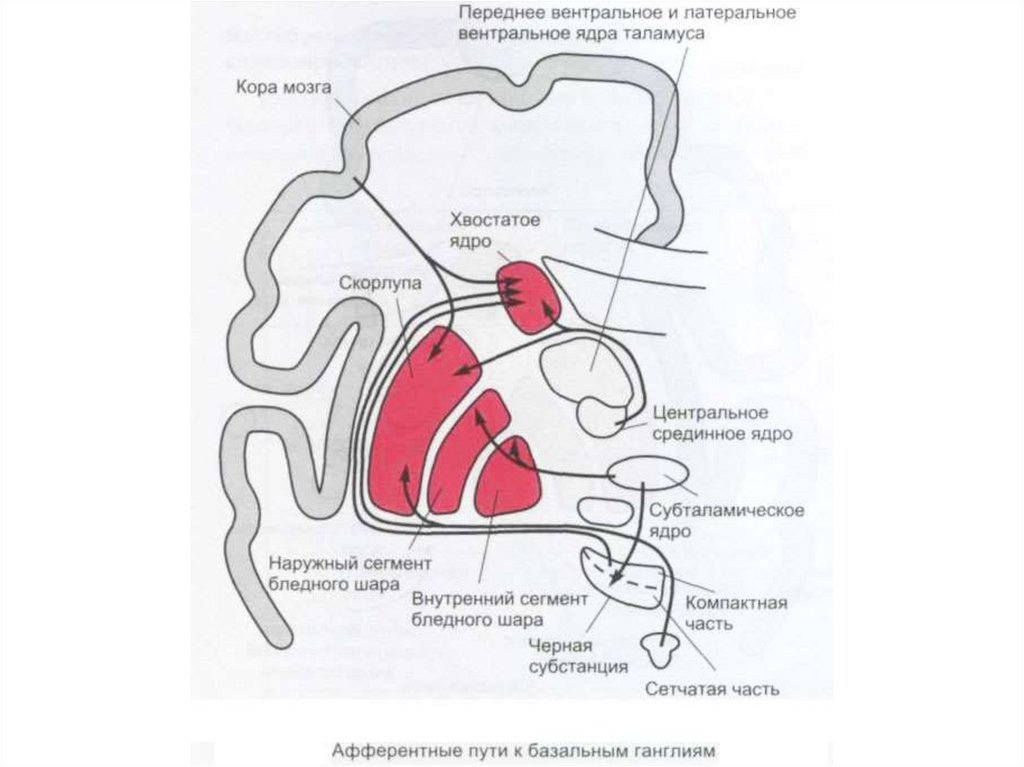









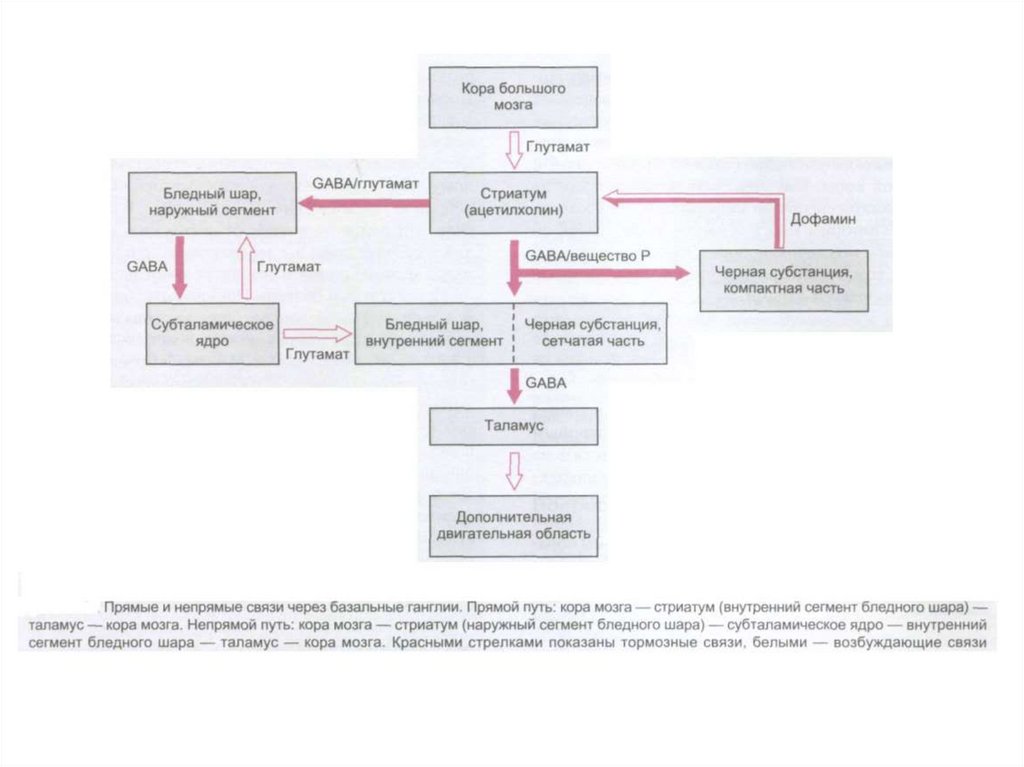









")



















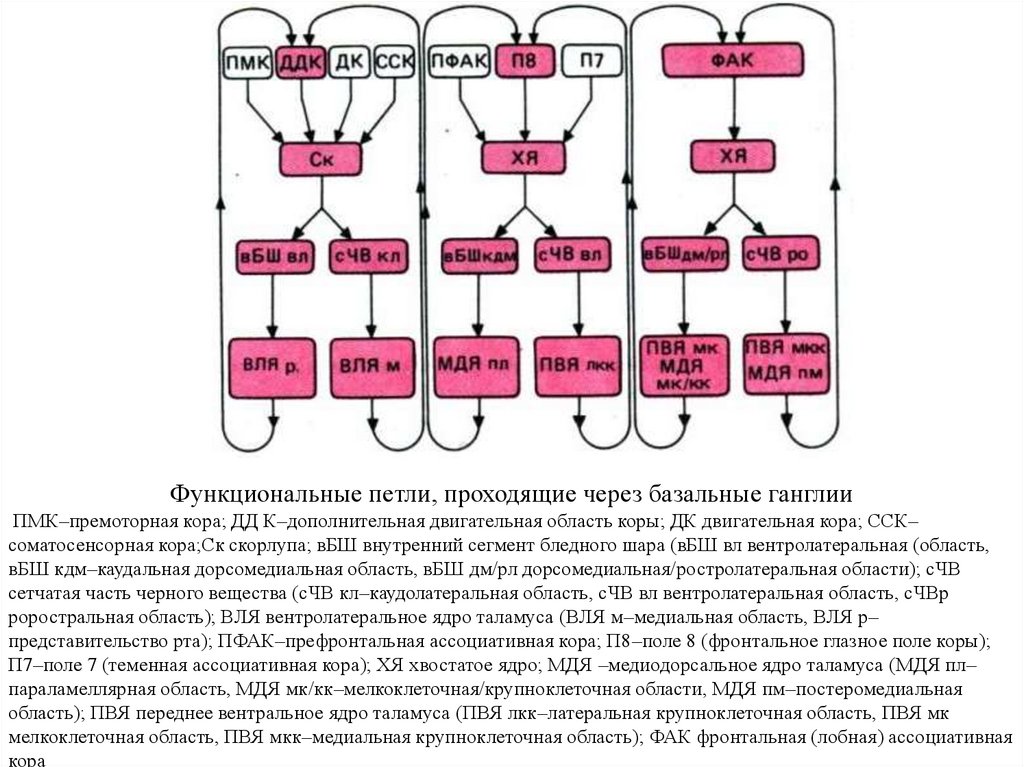
 функций")



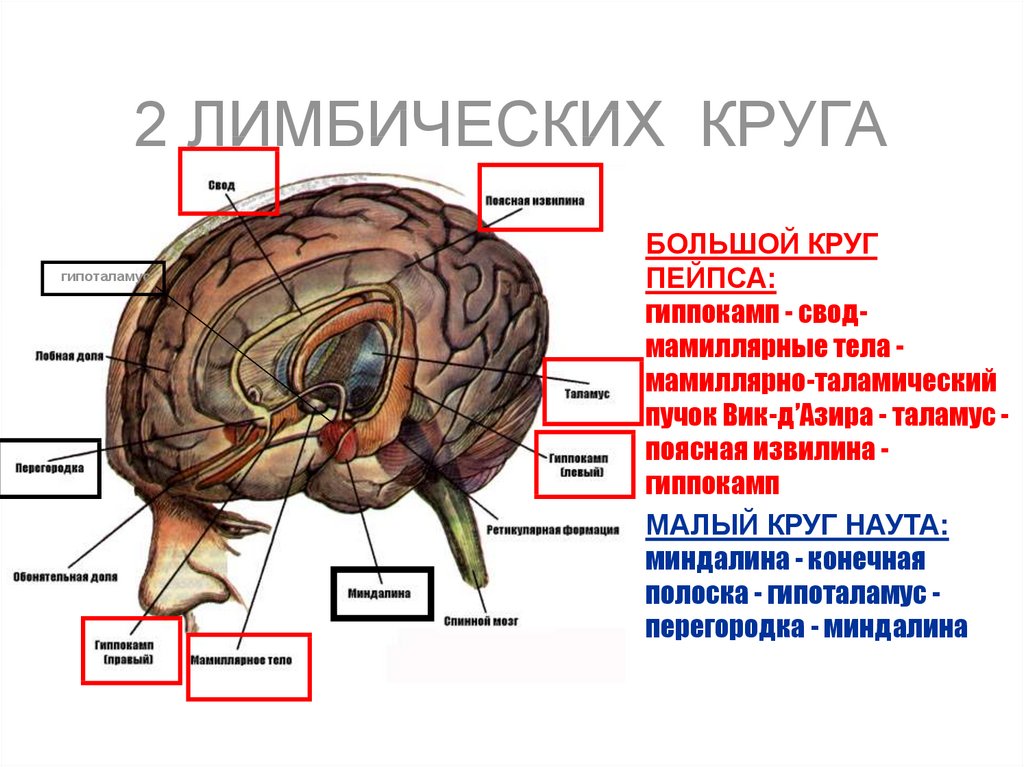
")









 функций")
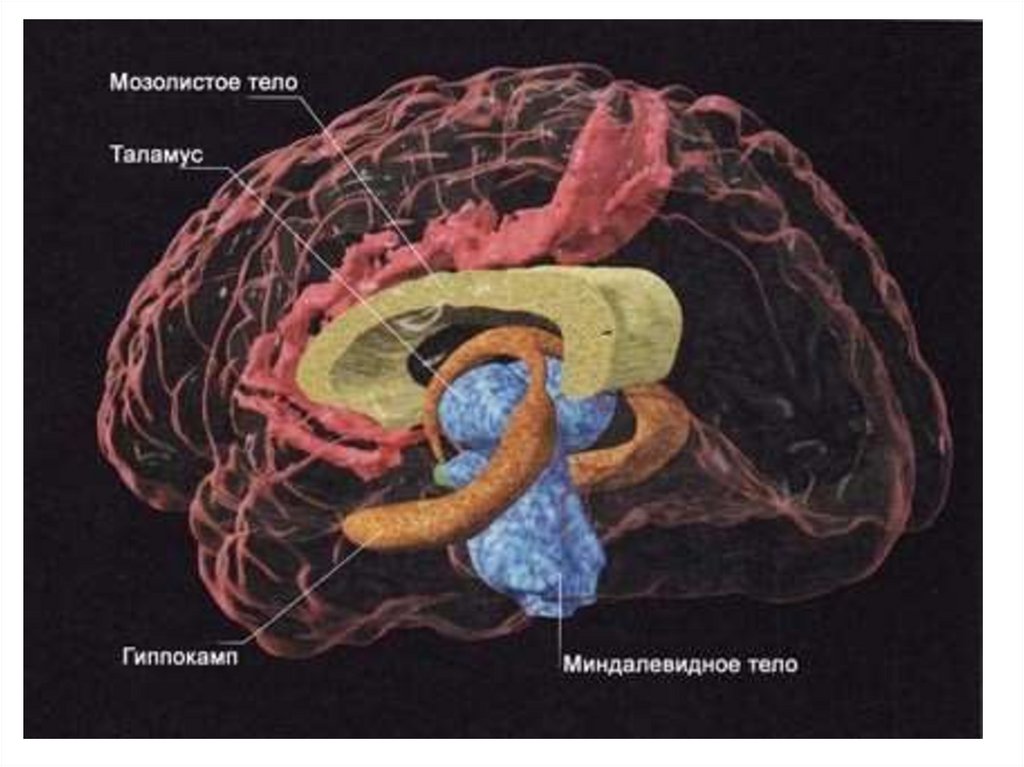
")










; вид сбоку. В них расположены первичная")












")

")
")
















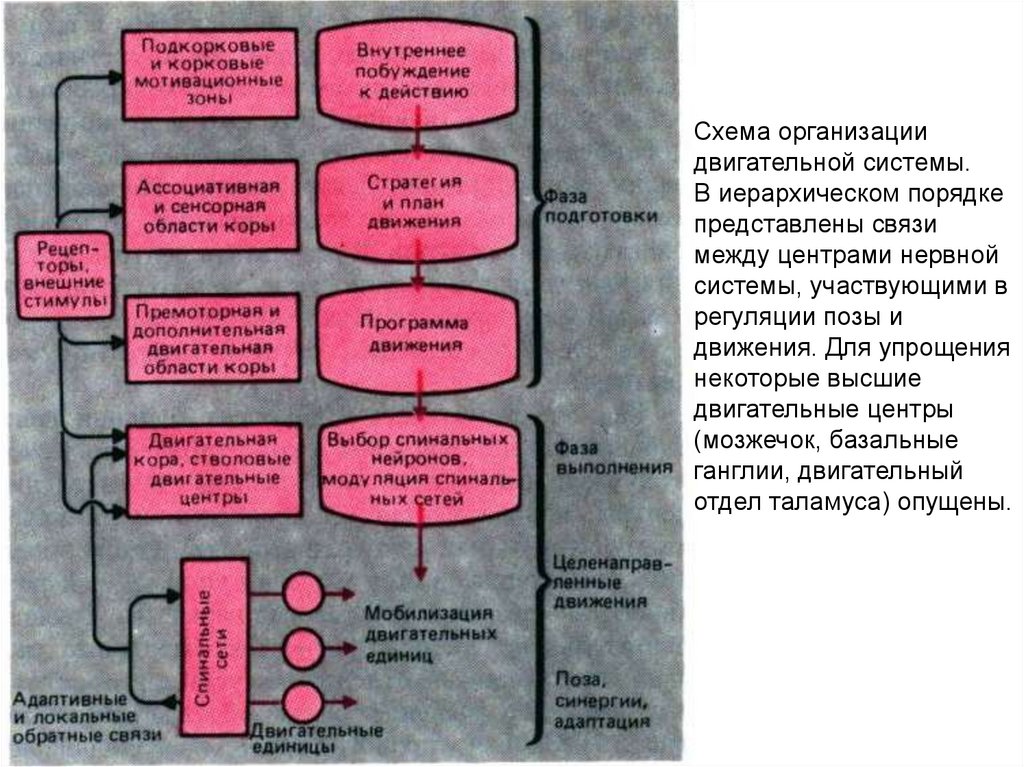
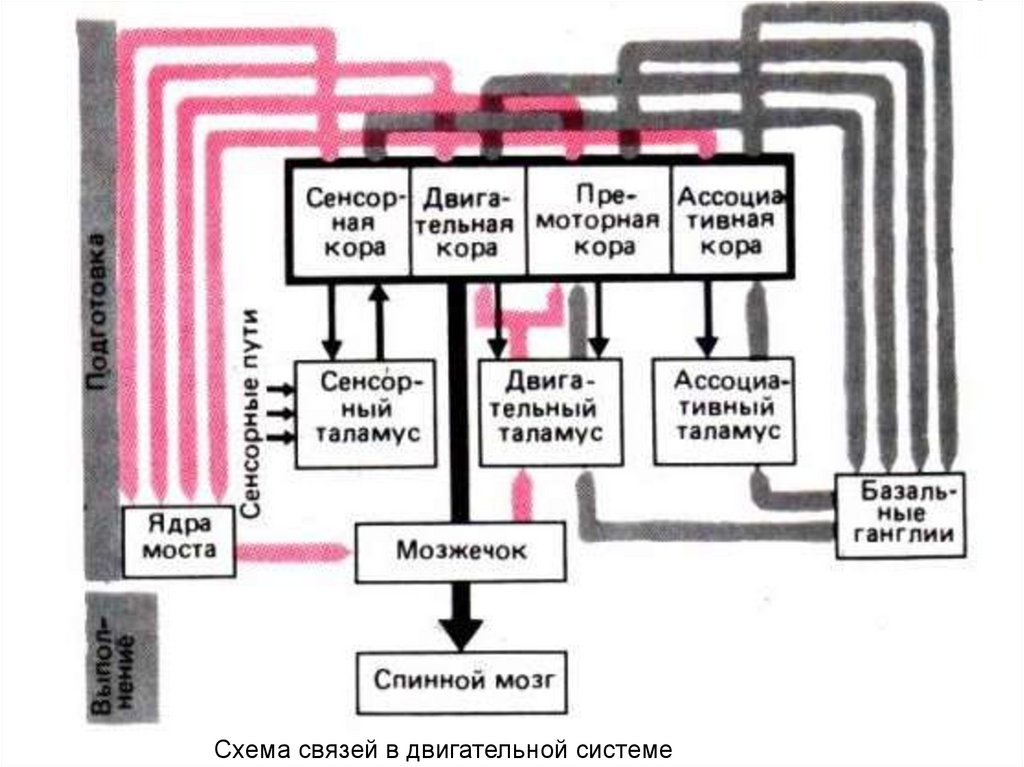

 medicine
medicineSimilar presentations:


")







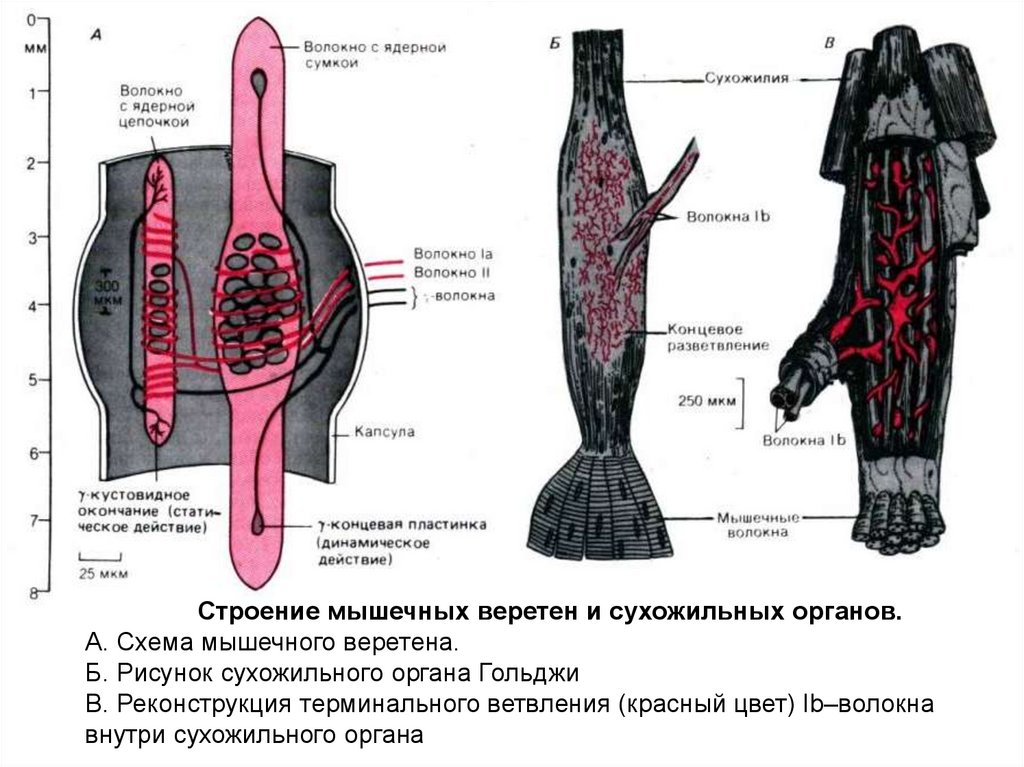
Строение мышечных веретен и сухожильных органов
1.
Строение мышечных веретен и сухожильных органов.А. Схема мышечного веретена.
Б. Рисунок сухожильного органа Гольджи
В. Реконструкция терминального ветвления (красный цвет) Ib–волокна
внутри сухожильного органа
2. Аференты Iа – аферентные волокна первичных мышечных веретён от центральных частей, являются датчиками изменения длины мышцы, то
есть своеобразныерецепторы растяжения. Посылают
информацию в ЦНС о том, что
мышца расслаблена и удлинена.
Импульсы по Iа-волокнам
возбуждают нейроны своего центра
и тормозят нейроны центра
антагониста.
Аференты II – вторичные
афференты от окончаний мышечных
веретён, служат датчиками
статической длины, работают по
всей конечности.
Iа и II – являются датчиками
изменения длины мышцы в
целом.
3. Аференты Ib – афференты от рецепторов сухожильно-мышечного соединения (аппарат Гольджи), относятся к рецепторам растяжения, но
Аференты Ib – афференты отрецепторов сухожильномышечного соединения (аппарат
Гольджи), относятся к
рецепторам растяжения, но
регистрируют напряжение,
развиваемое мышцей, то есть
посылают информацию в ЦНС о
том, что мышца сокращена, а
следовательно напряжено и
сухожилие.
Импульсы от них тормозят
нейроны своего центра и
возбуждают нейроны центраантагониста.
4. γ-мотонейроны – эфферентные γ-волкна (Аγ-волокна) иннервируют интрафузальные мышечные веретёна. 2 типа волокон: – Аγ-волокна с
концевойпластинкой, иннервируют
волокна с ядерной
сумкой.
– Аγ-волокна с кустиковым
окончанием, иннервируют
волокна с ядерной
цепочкой.
5.
6.
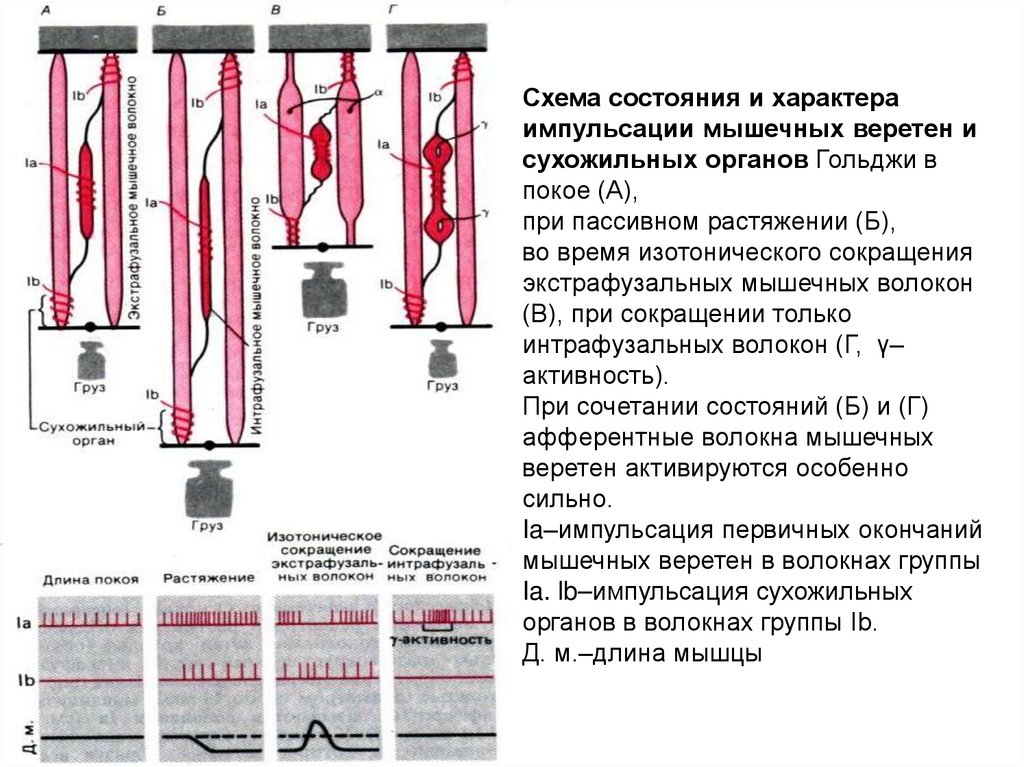
Схема состояния и характераимпульсации мышечных веретен и
сухожильных органов Гольджи в
покое (А),
при пассивном растяжении (Б),
во время изотонического сокращения
экстрафузальных мышечных волокон
(В), при сокращении только
интрафузальных волокон (Г, γ–
активность).
При сочетании состояний (Б) и (Г)
афферентные волокна мышечных
веретен активируются особенно
сильно.
Ia–импульсация первичных окончаний
мышечных веретен в волокнах группы
Ia. lb–импульсация сухожильных
органов в волокнах группы Ib.
Д. м.–длина мышцы
7.
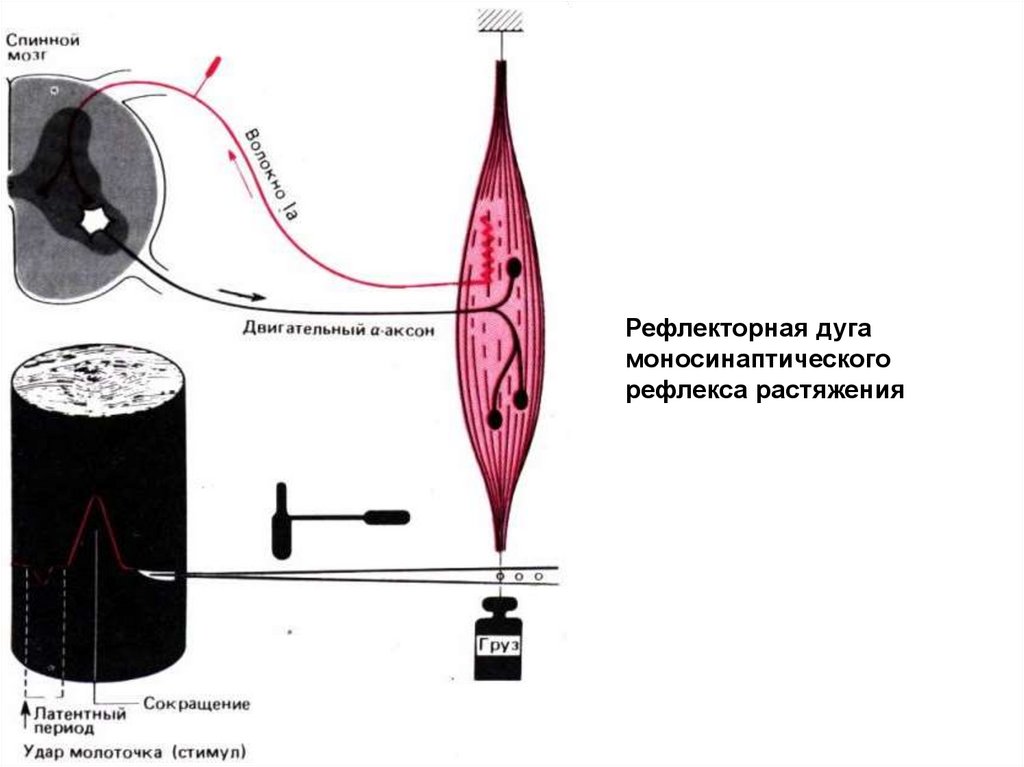
Рефлекторная дугамоносинаптического
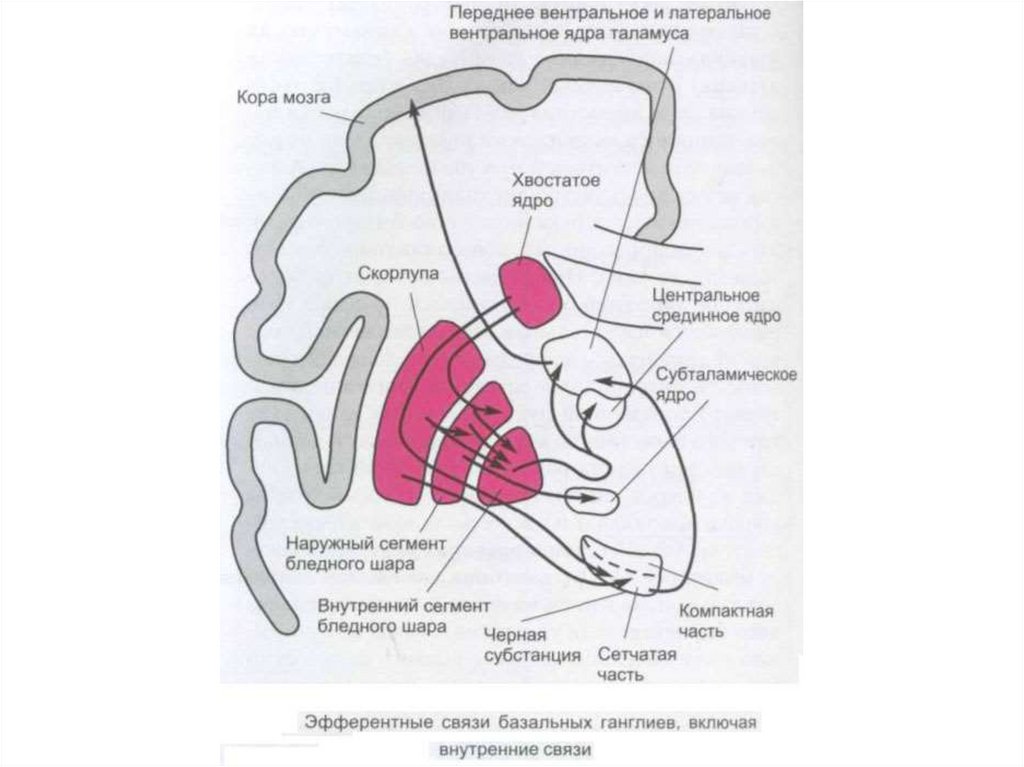
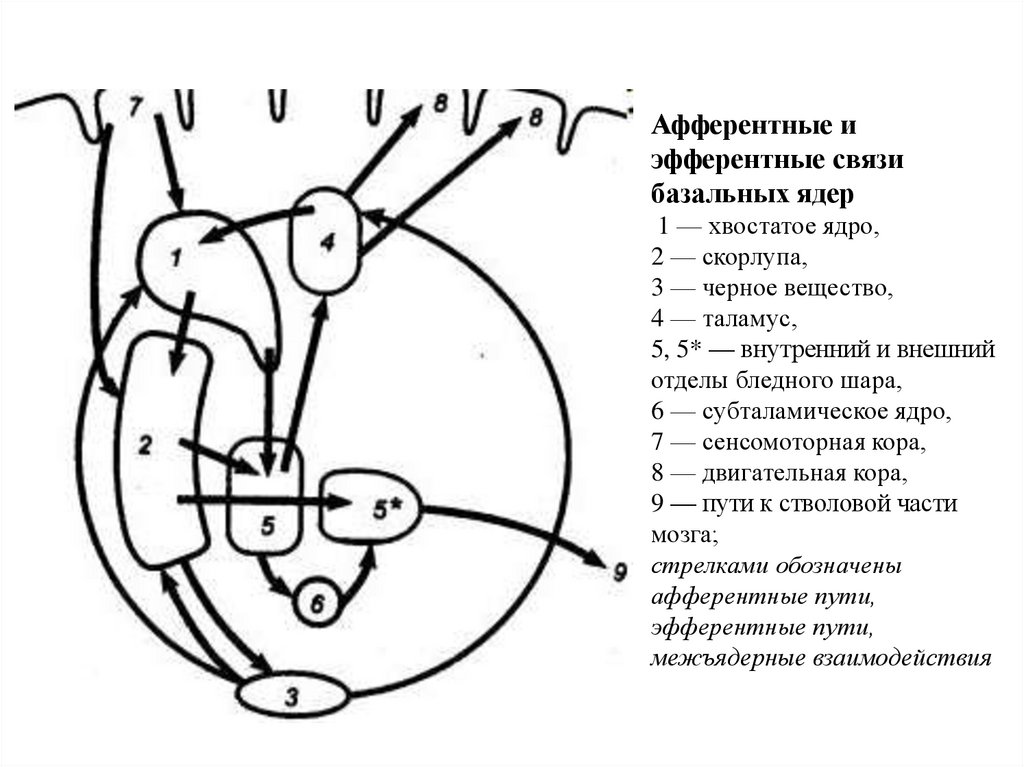
рефлекса растяжения
8. Облегчение Т–рефлексов.
Облегчение Т–рефлексов.Если коленный и другие Т–рефлексы нижней
конечности ослаблены, их можно усилить, если
больной, сцепив пальцы рук перед грудью,
попытается разорвать такой «замок» или сожмет
ладонь
другого
человека
(прием
Ендрассика). Развиваемое при этом усилие
облегчает активацию мотонейронов поясничного
отдела спинного мозга.
Аналогичным образом, Т–рефлексы облегчаются под
влиянием слабой фоновой активности мотонейронов
(т. е. при слабом напряжении иннервируемых ими
мышц).
9.
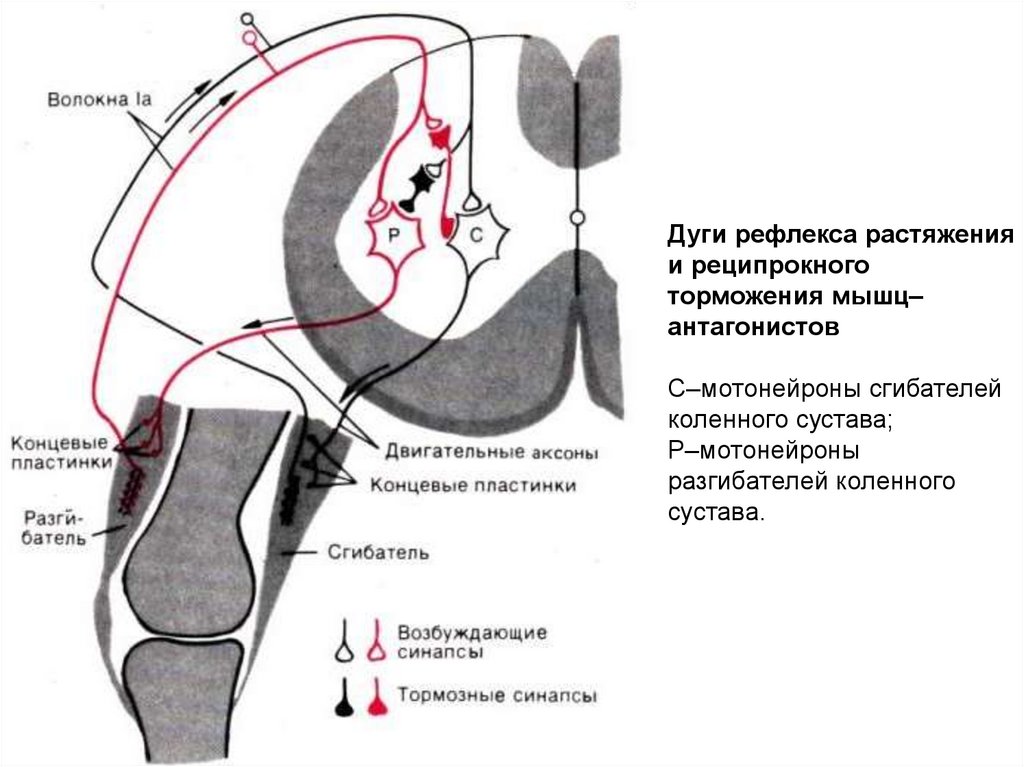
Дуги рефлекса растяженияи реципрокного
торможения мышц–
антагонистов
С–мотонейроны сгибателей
коленного сустава;
Р–мотонейроны
разгибателей коленного
сустава.
10.
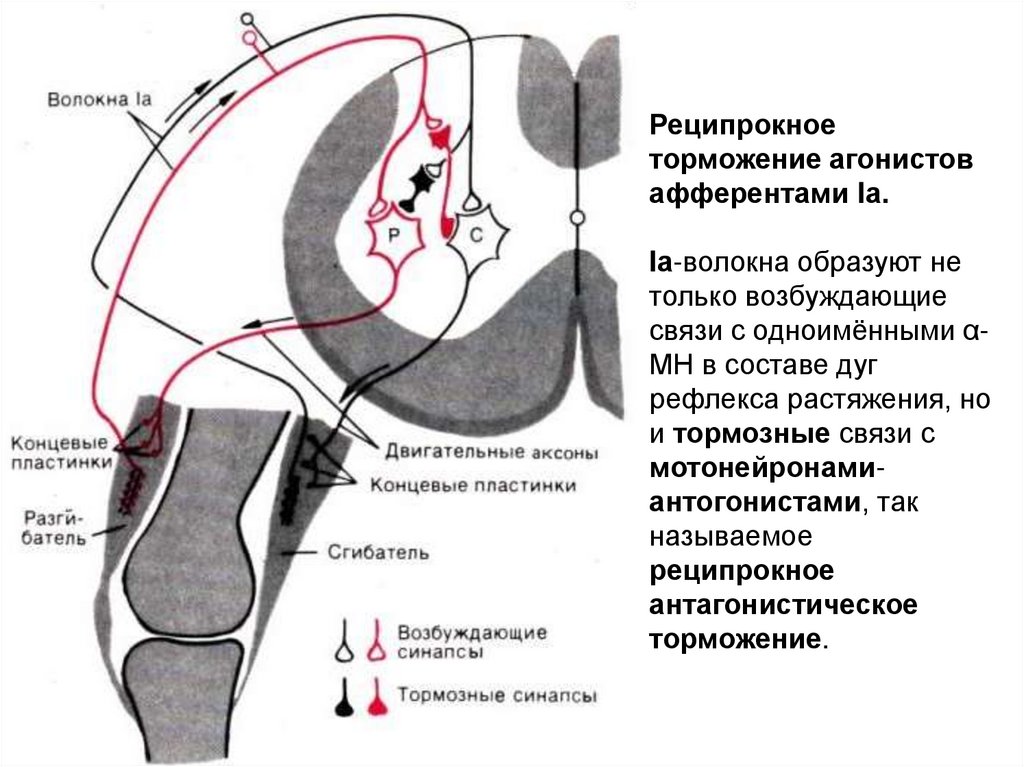
Реципрокноеторможение агонистов
афферентами Iа.
Iа-волокна образуют не
только возбуждающие
связи с одноимёнными αМН в составе дуг
рефлекса растяжения, но
и тормозные связи с
мотонейронамиантогонистами, так
называемое
реципрокное
антагонистическое
торможение.
11.
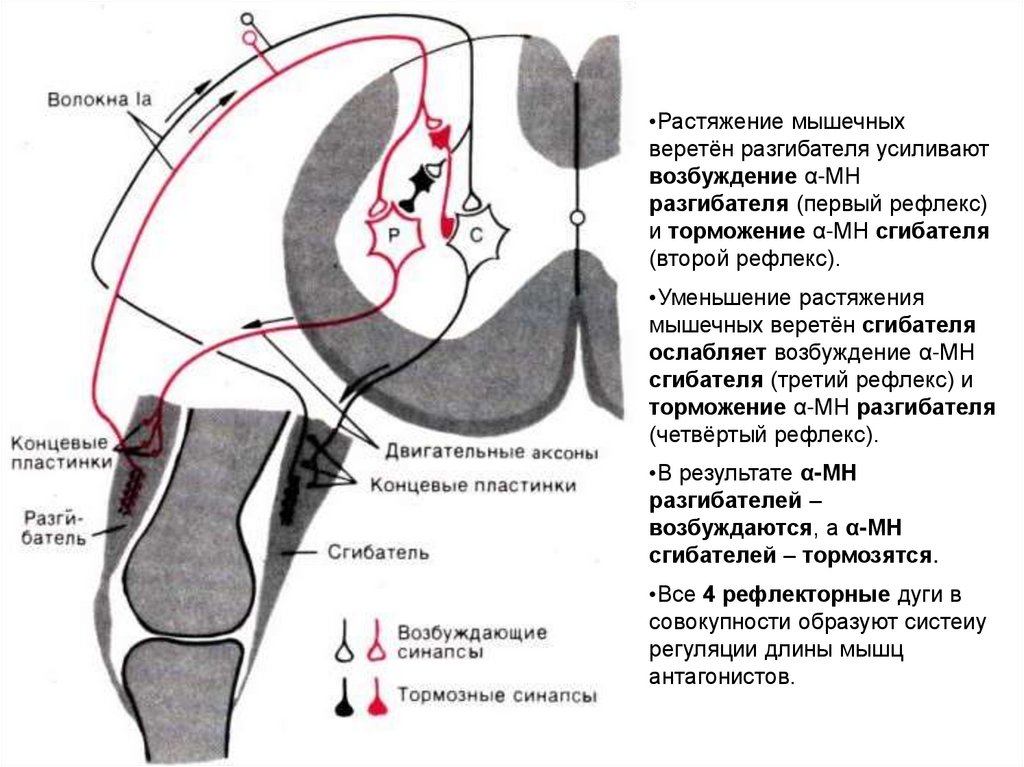
•Растяжение мышечныхверетён разгибателя усиливают
возбуждение α-МН
разгибателя (первый рефлекс)
и торможение α-МН сгибателя
(второй рефлекс).
•Уменьшение растяжения
мышечных веретён сгибателя
ослабляет возбуждение α-МН
сгибателя (третий рефлекс) и
торможение α-МН разгибателя
(четвёртый рефлекс).
•В результате α-МН
разгибателей –
возбуждаются, а α-МН
сгибателей – тормозятся.
•Все 4 рефлекторные дуги в
совокупности образуют систеиу
регуляции длины мышц
антагонистов.
12.
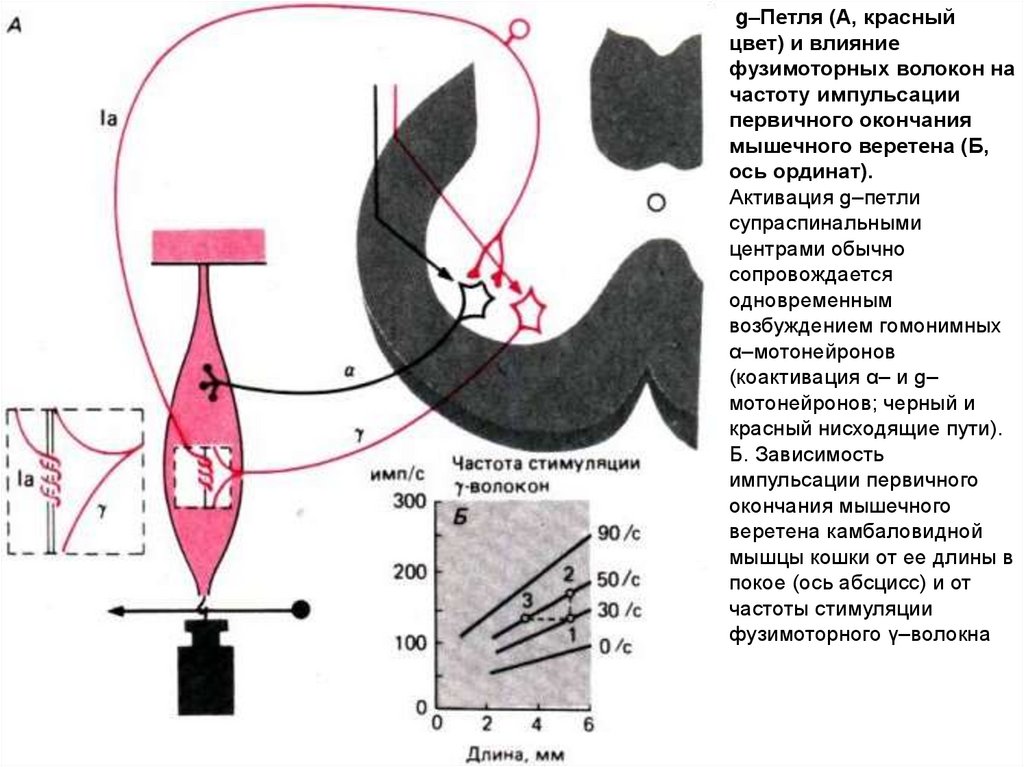
g–Петля (А, красныйцвет) и влияние
фузимоторных волокон на
частоту импульсации
первичного окончания
мышечного веретена (Б,
ось ординат).
Активация g–петли
супраспинальными
центрами обычно
сопровождается
одновременным
возбуждением гомонимных
α–мотонейронов
(коактивация α– и g–
мотонейронов; черный и
красный нисходящие пути).
Б. Зависимость
импульсации первичного
окончания мышечного
веретена камбаловидной
мышцы кошки от ее длины в
покое (ось абсцисс) и от
частоты стимуляции
фузимоторного γ–волокна
13. γ-петля
Мышечные веретёна могут вызвать сокращениеэкстрафузальных волокон в 2-х случаях:
1. При их растяжении.
2. При их активации со стороны γ-МН, приводящей к
сокращению веретён и растяжению их центральных
участков.
Второй механизм получил название γ-петли со стороны
супраспинальных структур, которые активируют
одновременно и α и γ-мотонейроны.
При этом, активация γ-МН со стороны супраспинальных
структур вызывает сокращение интрафузальных веретён и
увеличение Iа-импульсации на α-МН, а это в свою очередь
приводит к сокращению экстрфузальных волокон до
оптимальной длинны, то есть наблюдается коактивация α
и γ-мотонейронов при возбуждении.
14.
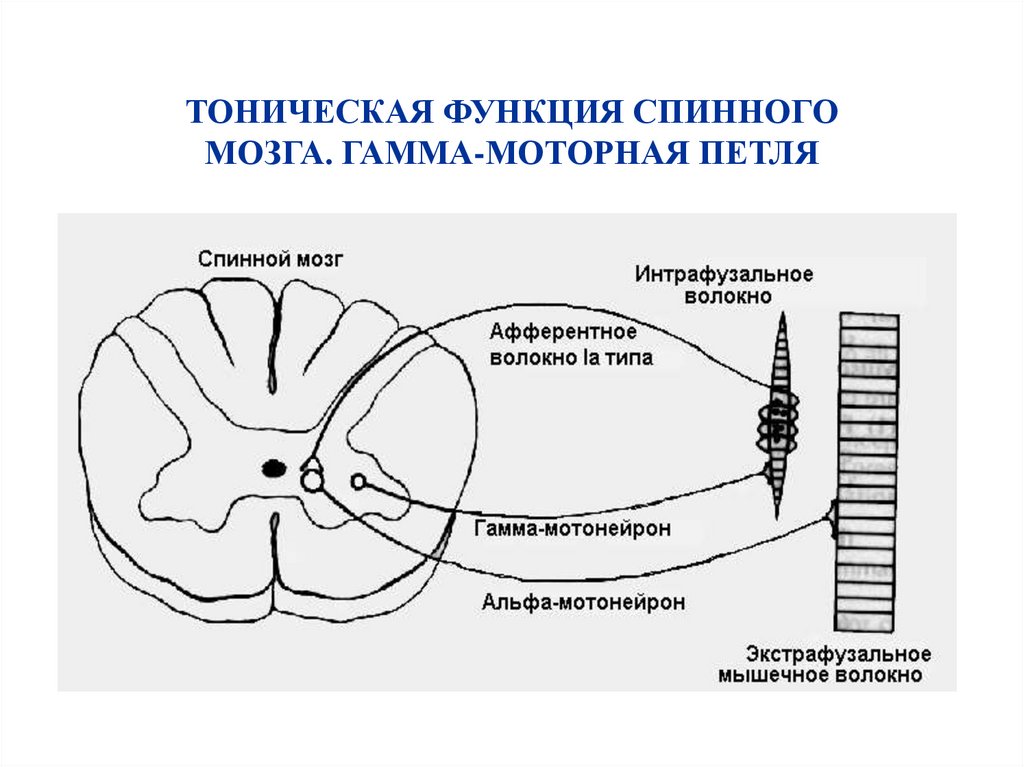
ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГОМОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
15. Коактивация альфа– и гамма–мотонейронов при движениях.
Коактивация альфа– и гамма–мотонейронов при движениях.
α–
и
γ–мотонейроны
возбуждаются
одновременно;
однако
начало
импульсации
афферентов
мышечных
веретен отстает от вспышки электромиографически
регистрируемой активности из–за относительно низкой скорости проведения по у–волокнам и латентного
периода сокращения интрафузальных волокон.
Из этого следует, что главное назначение γ–иннервации, возможно, в том, чтобы предотвратить во время
сокращения экстрафузальных волокон расслабление мышечных веретен, а следовательно, сохранить их
адекватную рецепторную функцию и, таким образом, стабилизирующий эффект рефлекса растяжения
даже во время движения.
Кроме того, усиление активности мышечных веретен при возбуждении γ–мотонейронов способствует развитию
начавшегося движения.
Одновременно, чувствительность рецептора регулирующей системы (первичных окончаний мышечных
веретен) поддерживается в адекватном диапазоне. Следовательно, γ–петля представляет собой
сервомеханизм для оптимизации движений.
16. Выводы по γ-петле
• Главное назначение γ-иннервации состоит втом, чтобы предотвратить во время
сокращения экстрафузальных волокон –
расслабление интрафузальных веретён, а
следовательно сохранить их адекватную
рецепторную функцию во время движения.
• Кроме этого, увеличение активности
мышечных веретён при возбуждении γМНспособствует развитию и поддержанию
начавшегося движения.
17. Функции сухожильных органов.
Рефлекторная дуга сухожильных органов служит для поддержания постоянстванапряжения мышцы.
У каждой мышцы две регуляторные системы обратной связи: регуляции
длины с мышечными веретенами в качестве рецепторов и регуляции
напряжения, рецепторами в которой служат сухожильные органы.
Влияние системы регуляции длины в принципе ограничивается одной мышцей и ее
антагонистом, тогда как регуляция напряжения с участием афферентов Ib
относится к мышечному тонусу всей конечности.
18.
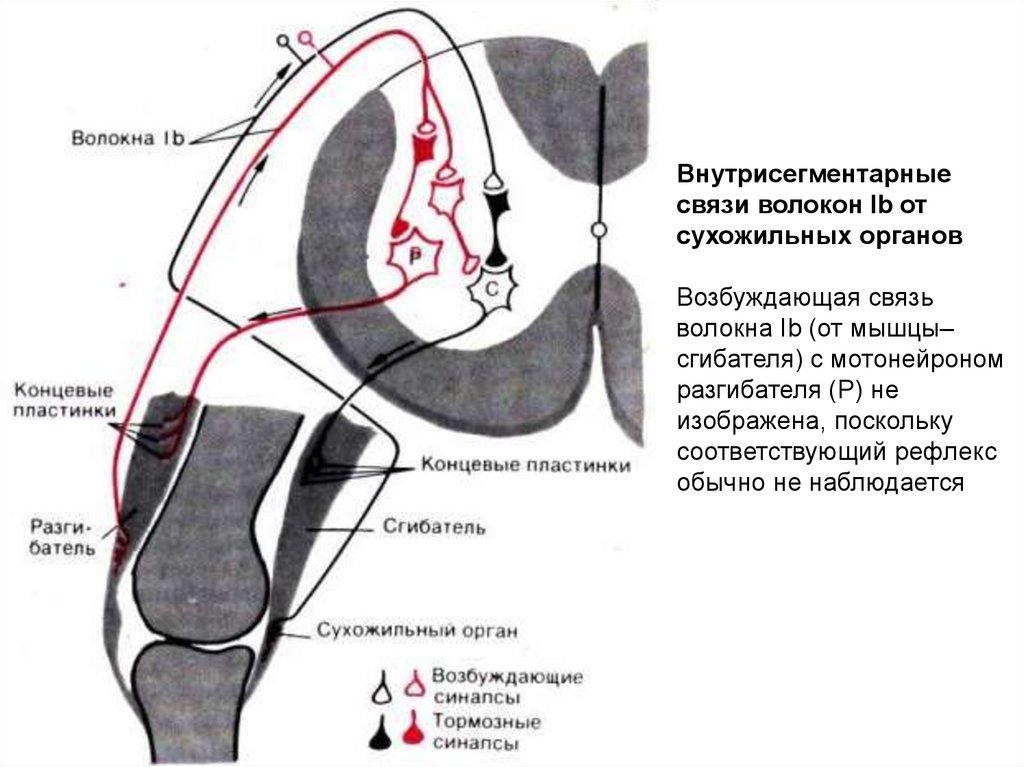
Внутрисегментарныесвязи волокон Ib от
сухожильных органов
Возбуждающая связь
волокна Ib (от мышцы–
сгибателя) с мотонейроном
разгибателя (Р) не
изображена, поскольку
соответствующий рефлекс
обычно не наблюдается
19.
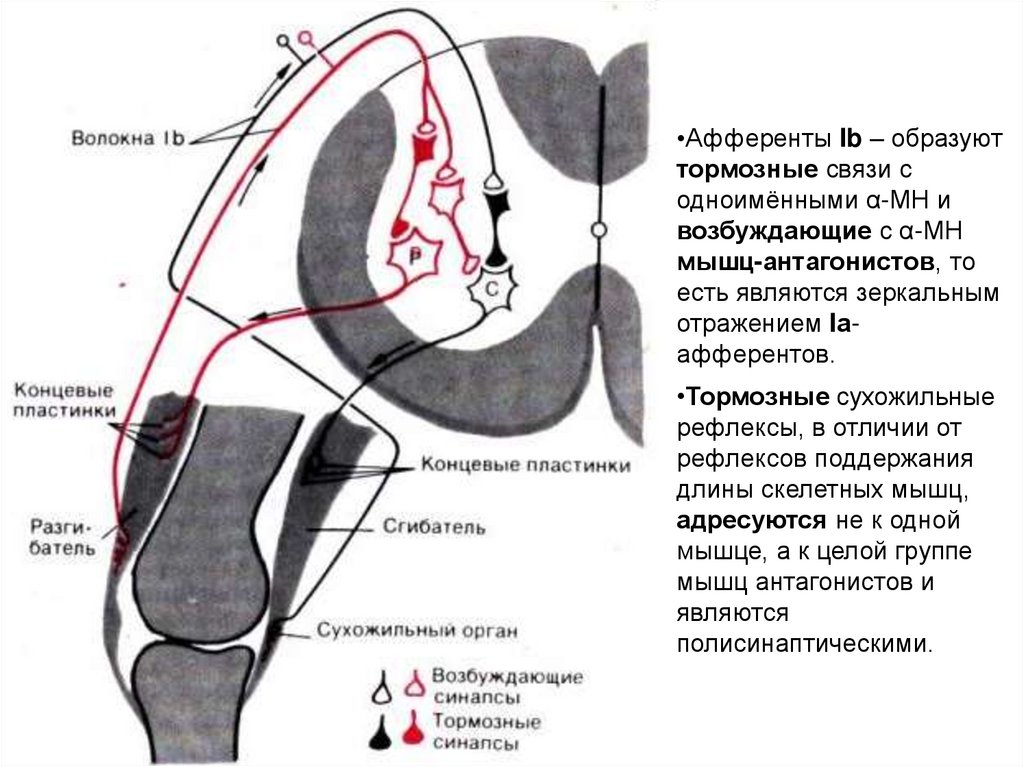
•Афференты Ib – образуюттормозные связи с
одноимёнными α-МН и
возбуждающие с α-МН
мышц-антагонистов, то
есть являются зеркальным
отражением Iаафферентов.
•Тормозные сухожильные
рефлексы, в отличии от
рефлексов поддержания
длины скелетных мышц,
адресуются не к одной
мышце, а к целой группе
мышц антагонистов и
являются
полисинаптическими.
20. Сгибательные рефлексы
Осуществляются при раздражении кожных рецепторов, рецепторовсуставов, рецепторов давления и боли скелетных мышц.
Например, оборонительный сгибательный рефлекс. Этот рефлекс намного
сложнее миототического – здесь сочетается процесс координации
фазного движения с процессом сохранения позы (тонический рефлекс).
21.
22.
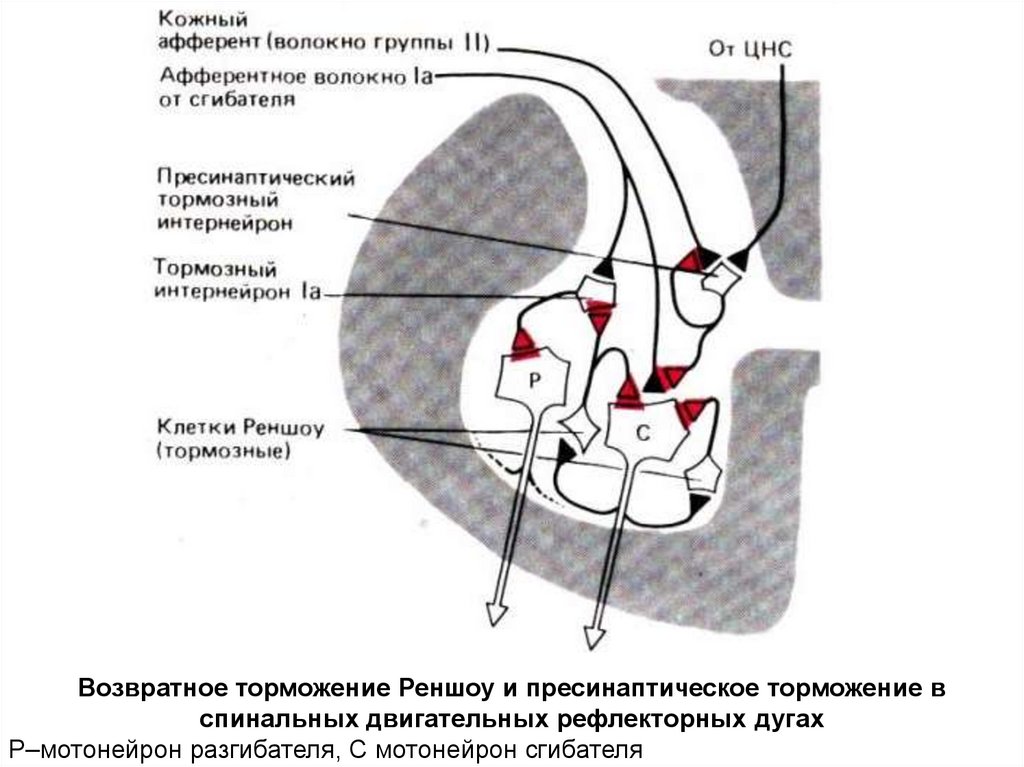
Возвратное торможение Реншоу и пресинаптическое торможение вспинальных двигательных рефлекторных дугах
Р–мотонейрон разгибателя, С мотонейрон сгибателя
23. Таким образом: на уровне СМ в деятельности каждой мышцы участвуют как минимум 2 системы обратной связи: - Система регуляции
длины смышечными веретёнами в качестве
рецепторов.
- Система регуляции напряжения,
рецепторами в которой служат
сухожильные органы.
24. Постуральные рефлексы
(от английск. postur—поза, положение),термин, предложенный Шеррингтоном
для
обозначения
рефлексов,
обеспечивающих
сохранение
определенного положения.
Постуральные
(позотонические,
статические)
рефлексы
обеспечивают
поддержание
в
пространстве
определённого
положения всего тела или его части
(например, конечности).
Познотонические рефлексы возникают с
проприорецепторов
мышц
шеи,
рецепторов фасции шеи животного.
Переключение с этих рецепторов
осуществляется на уровне шейного
отдела спинного мозга и приводит к
изменению
тонуса
мышц
при
изменении положения головы и шеи (
Магнус и Клейн).
Наиболее наглядно это проявляется на
декапитированой
лягушке.
У
млекопитающих
этот
вид
деятельности переходит к структурам
ствола мозга.
25.
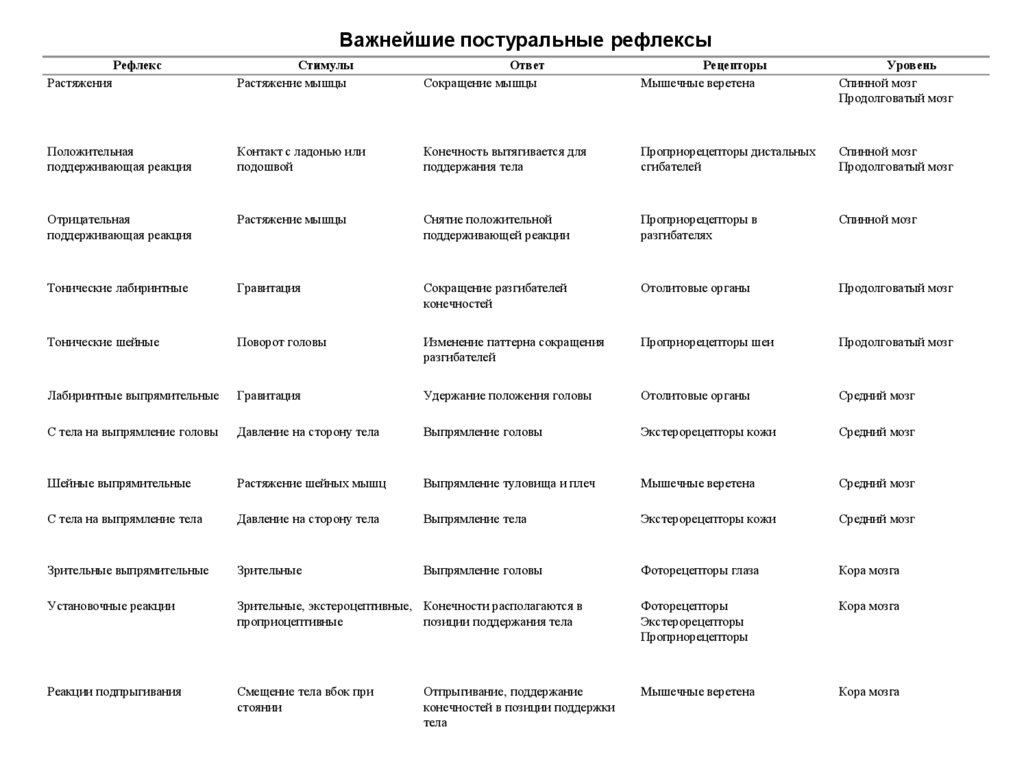
Важнейшие постуральные рефлексыРастяжения
Рефлекс
Стимулы
Растяжение мышцы
Ответ
Сокращение мышцы
Рецепторы
Мышечные веретена
Уровень
Спинной мозг
Продолговатый мозг
Положительная
поддерживающая реакция
Контакт с ладонью или
подошвой
Конечность вытягивается для
поддержания тела
Проприорецепторы дистальных
сгибателей
Спинной мозг
Продолговатый мозг
Отрицательная
поддерживающая реакция
Растяжение мышцы
Снятие положительной
поддерживающей реакции
Проприорецепторы в
разгибателях
Спинной мозг
Тонические лабиринтные
Гравитация
Сокращение разгибателей
конечностей
Отолитовые органы
Продолговатый мозг
Тонические шейные
Поворот головы
Изменение паттерна сокращения
разгибателей
Проприорецепторы шеи
Продолговатый мозг
Лабиринтные выпрямительные
Гравитация
Удержание положения головы
Отолитовые органы
Средний мозг
С тела на выпрямление головы
Давление на сторону тела
Выпрямление головы
Экстерорецепторы кожи
Средний мозг
Шейные выпрямительные
Растяжение шейных мышц
Выпрямление туловища и плеч
Мышечные веретена
Средний мозг
С тела на выпрямление тела
Давление на сторону тела
Выпрямление тела
Экстерорецепторы кожи
Средний мозг
Зрительные выпрямительные
Зрительные
Выпрямление головы
Фоторецепторы глаза
Кора мозга
Установочные реакции
Зрительные, экстероцептивные, Конечности располагаются в
проприоцептивные
позиции поддержания тела
Фоторецепторы
Экстерорецепторы
Проприорецепторы
Кора мозга
Реакции подпрыгивания
Смещение тела вбок при
стоянии
Мышечные веретена
Кора мозга
Отпрыгивание, поддержание
конечностей в позиции поддержки
тела
26.
Постуральные рефлексы, приведённые в таблице, показывают,что на уровне спинного, продолговатого и среднего мозга
осуществляются сложнокоординированные реакции позы и
движений.
Автоматизм этих реакций разгружает более высокие уровни
регуляции мышечной активности от управления стандартными
положениями частей тела по отношению друг к другу.
27. Спинальная локомоция (автоматизмы)
Основные характеристики локомоции, т.е. перемещения человека или животного в окружающейсреде при помощи координированных движений конечностей, запрограммированы на
уровне спинного мозга .
Болевое раздражение какой–либо конечности спинального животного вызывает рефлекторные
движения всех четырех; если же такая стимуляция продолжается достаточно долго, могут
возникнуть ритмичные сгибательные и разгибательные движения не подвергающихся
раздражению конечностей.
Если такое животное поставить на тредбан, то при некоторых условиях оно будет совершать
координированные шагательные движения, весьма сходные с естественными. Их
выполнение обеспечит изолированный спинной мозг в отсутствие обратной афферентации
от рецепторов, активирующихся во время локомоции.
Активность центров координируется проприоспинальными системами и трактами,
пересекающими спинной мозг в пределах отдельных сегментов.
У более примитивных животных спинальные локомоции могут проявляться - в беге без головы.
28.
Предполагают, что у человекаспинальные локомоторные центры.
тоже
есть
По–видимому, их активация при раздражении кожи
проявляется в виде шагательного рефлекса
новорожденного ( сохраняется до 2-х месяцев).
Однако, по мере созревания центральной нервной
системы
супраспинальные
отделы,
очевидно,
настолько подчиняют себе такие центры, что у
взрослого человека они утрачивают способность к
самостоятельной активности.
Возможно,
именно
поэтому
у
больных
с параплегией пока еще не удавалось добиться
координированной локомоции.
29. СОСАТЕЛЬНЫЙ РЕФЛЕКС
30. Шагательный автоматизм
Поднимите малыша под мышки ( стопы должны лишьприкасаться к поверхности ). Затем слегка наклоните его
вперед: ребёнок сделает несколько шагов. Некоторые
перебирают ногами так, словно поднимаются по лестнице.
Реакция сохраняется до 8 месяцев. Скрещивание ног или
отсутствие рефлекса автоматической ходьбы говорят о
проблемах в поясничном отделе.
31. Автоматизм Бауэра
Когда ребенок лежит на животе, подставьте свою ладонь емупод стопы. Он оттолкнется ножками и продвинется вперед.
Иногда ему достаточно небольшой опоры ( например, одного
лишь вашего пальца ), чтобы набрать скорость. Он может
упереться и в прутья кровати, и в бортик пеленального
столика. Так что не оставляйте малыша без присмотра.
32. Рефлекс Бабинского
Грудничок разводит пальчики на ногах веером лишь тогда,когда вы касаетесь внешней стороны стопы или пяточки. Этот
автоматизм проходит к 5-му месяцу.
33. Рефлекс Моро
Положите малыша на спинку и хлопните ладонями по кроватирядом с его головой. Он раскроет кулачки, разведет руки в
стороны и резко обнимет себя ими. Ножки пи этом либо
вытянет, либо прижмет к животу. Проходит автоматизм к 4-му
месяцу.
34. РЕФЛЕКС ГАЛАНТА
Проведите пальцем вдоль позвоночника с правой стороны ( нарасстоянии сантиметра от него ). Малыш выгнется дугой в
ответ на ваше прикосновение и подожмет правую ножку. Так
он будет реагировать до полугода, а затем автоматизм
постепенно исчезнет.
35. Хватательный автоматизм Робинсона.
Угасает этот рефлекс к 4-му месяцу. А на смену ему приходитсознательное хватание и удерживание предметов
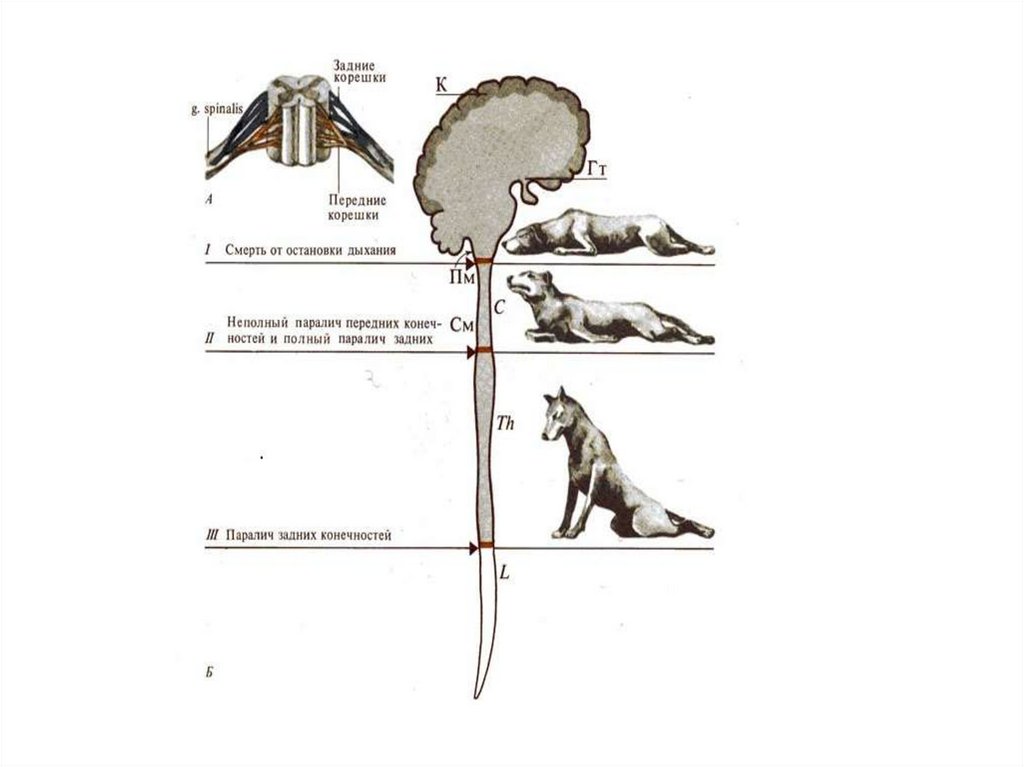
36. Полная параплегия возникает обычно при разрыве спинного мозга в грудном отделе– от Т2 до Т12 и сопровождается:
Cопровождается:1)мгновенным и окончательным параличом всех произвольных движений мышц, иннервируемых от сегментов,
расположенных каудальнее места повреждения;
2) полной и окончательной потерей осознаваемой чувствительности областей тела, соответствующих этим
сегментам;
3) временной полной арефлексией, т.е. временным исчезновением всех двигательных и вегетативных
рефлексов в тех же областях тела.
В
последующие
недели
рефлексы восстанавливаются.
и
месяцы
двигательные
Выделяют следующие четыре стадии:
(I)полная арефлексия (обычно 4–6 недель);
(II)небольших рефлекторных движений пальцев ног, в первую очередь–большого (от двух недель до нескольких
месяцев);
(III)постепенное усиление сгибательных рефлексов–сначала большого пальца ноги (рефлекс Бабинского) и
голеностопного сустава, затем колена и бедра. Генерализованные сгибательные рефлексы в некоторых случаях
сопровождаются перекрестными разгибательными. Стопа, особенно подошва, представляет самую чувствительную
рефлексогенную зону для таких сложных реакций; даже ее слабое тактильное раздражение способно вызвать
генерализованный сгибательный рефлекс;
(IV)в хронической стадии (через шесть и более месяцев) обычно продолжают преобладать сгибательные рефлексы,
хотя могут усиливаться и разгибательные, иногда переходящие в длительные разгибательные спазмы.
37.
38.
СПИНАЛЬНЫЙ ШОКРазвивается в результате прерывания супраспинапьных
влияний на мотонейроны спинного мозга при его
травмах.
Развиваются
обратимые
угнетения.
Различают 4 стадии спинального шока:
1 - полная арефлексия (4-6 недель) — мгновенно и
окончательно
выключаются
все
произвольные
движения мышц, иннервируемых из сегментов,
лежащих ниже места повреждения.
- полный и окончательный потерей осозноваемой
чувствительности в областях тела, соответствующих
этим сегментам.
временной
арефлексии,
т.е.
временным
исчезновением рефлексов, дуги которых проходят
ниже места травмы.
39.
• 2. Период появления небольших рефлекторных движенийпальцев ног и особенно в первую очередь большого пальца (от
2 недель до несколько месяцев)
• 3. Стадия постепенного усиления сгибательных рефлексов. В
эту стадию сначала появляются рефлекторные реакции
большого пальца ноги (рефлекс Бабинского) и стопы, а в
дальнейшем рефлекторное сгибание колена и бедра. Иногда
сопровождаются перекрестными расгибательными рефлексами.
• 4. Хроническая стадия (спустя шесть и более месяцев)
характеризуется преобладанием сгибательных рефлексов,
однако могут наблюдать и выраженные расгибательные
рефлексы, переходящие в длительные расгибательные спазмы
(спинальное стояние).
Отклонения от подобной клинической картины, особенно наличие
расгибательных рефлексов и повешенного мышечного тонуса, обычно
служит симптомом неполного перерыва СМ.
40. Механизм гипертонуса мышц после спинального шока
• После исчезновения спинального шока, тонусмышц, ниже повреждения резко повышается, а
сгибательные и разгибательные рефлексы нижних
конечностей усиливаются:
• Причинами гипертонуса яв-ся:
- спонтанная активность мышечных рецепторов, что
обеспечивает обильную импульсацию от них к αмотонейронам, а их возбуждение приводит к
сокращению мышц.
- спонтанная активность γ-мотонейронов приводит к
сокращению мышечных веретен и как следствие к их
возбуждению, которое по Ia – волокнам поступает к
α-мотонейронам, вызывая их возбуждение, что
приводит к постоянному (тоническому) сокращению
мышц и развивается гипертонус у спинального
организма.
41.
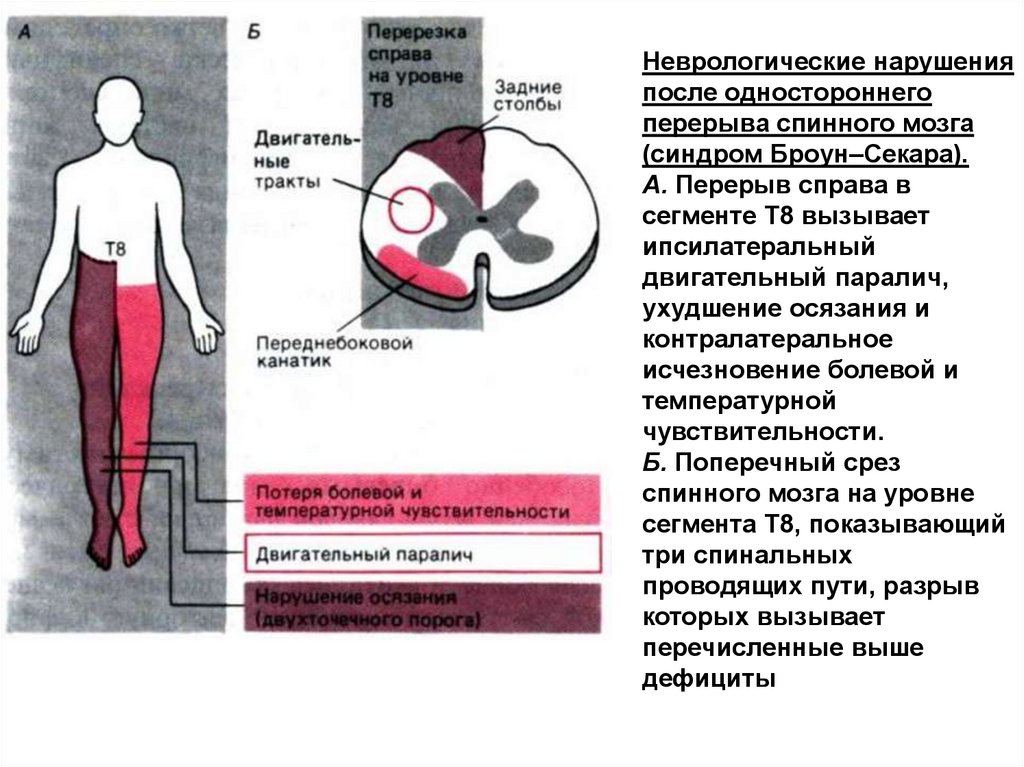
СИНДРОМ БРОУН-СЕКАРА Одностороннийперерыв
спинного
мозга,
характеризуется на стороне поражения двигательным
параличем и потерей осязания, а на противоположной потерей болевой и температурной чувствительности.
Причины:
блокада
нисходящих
путей
(пирамидный
тракт) — паралич произвольных движений
повреждение
заднего
столба
(потеря
осязания)
повреждение
переднебокового
канатика
—
потеря
болевой,
температурной
чувствительной на противоположной стороне.
42. Зоны поражения у человека при повреждении спинного мозга на уровне грудных сегментов (синдром Броун-Секара)
43.
Неврологические нарушенияпосле одностороннего
перерыва спинного мозга
(синдром Броун–Секара).
А. Перерыв справа в
сегменте Т8 вызывает
ипсилатеральный
двигательный паралич,
ухудшение осязания и
контралатеральное
исчезновение болевой и
температурной
чувствительности.
Б. Поперечный срез
спинного мозга на уровне
сегмента Т8, показывающий
три спинальных
проводящих пути, разрыв
которых вызывает
перечисленные выше
дефициты
44.
45.
46.
47. Вегетативные (интероцептивные ) рефлексы спинного мозга
48. Висцеромоторные рефлексы Проявляются в двигательных реакциях мышц грудной клетки и брюшной стенки, разгибателей спины при
раздражении рецептороввнутренних органов.
Рефлексы вегетативной нервной
системы
Обеспечивают реакцию внутренних органов, сосудистой
системы на раздражение висцеральных, мышечных кожных
рецепторов
(Зоны Геда, рефлекс Гольца и др.)
49. рефлекс Гольца рефлекс Данини-Ашнера
рефлекс Гольцарефлекс Данини-Ашнера
раздражение - удар в эпигастральную
область, реакция— рефлекторная остановка сердца
50. Зоны Захарьина-Геда
51.
52.
ПАТОЛОГИЧЕСКИЕ РАЗГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ СПИННОГОМОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ ВЛИЯНИЙ У
ЧЕЛОВЕКА
-
-
-
РЕФЛЕКС БАБИНСКОГО - при раздражении подошвы стопы
штриховым движением от пятки до носка происходит
разгибание большого пальца в сторону тыла.
В норме этот рефлекс имеется у детей примерно до двух лет.
Потом
тормозиться
и
выявляется
при
нарушении
супраспинальных рефлексов.
РЕФЛЕКС ОПЕНГЕЙМА - при надавливании на поверхность
голени
в
области ТИБИА от колена до голеностопного сустава происходит тыльное расгибание большого пальца.
РЕФЛЕКС ГОРДОНА - при сдавливании икраножной мышцы
происходит тыльное расгибание большого пальца
РЕФЛЕКС ШЕФЕРА - то же самое, при сдавливании ахиллового
сухожилия.
53.
ПАТОЛОГИЧЕСКИЕ СГИБАТЕЛЬНЫЕ РЕФЛЕКСЫСПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ
ВЛИЯНИЙ У ЧЕЛОВЕКА
• ХВАТАТЕЛЬНЫЙ РЕФЛЕКС ЯНУШКЕВИЧА - возникает при
касании пальцев руки или стопы каким либо предметом и
проявляется в непроизвольном захватавании этого предмета. У
детей этот рефлекс проявляется в возрасте до 2 - 4 месяцев,
при этом удается приподнять ребенка над опорой —
хватательный рефлекс РОБИНСОНА
• РЕФЛЕКС РОССОЛИНО - сгибание пальцев стопы, в том числе
и большого при быстром касательном ударе по подушечкам
пальцев стопы
• РЕФЛЕКС БЕХТЕРЕВА - МЕНДЕЛЯ - подошвенное сгибание
большого пальца при ударе молоточком по тылу стопы
• РЕФЛЕКС ЖУКОВСКОГО - то же самое при ударе по
подошвенной поверхности под пальцами стопы
• РЕФЛЕКС БЕХТЕРЕВА - то же самое при ударе по пятке
54.
ПАТОЛОГИЧЕСКИЕ РЕФЛЕКСЫ, ВЫЗЫВАЕМЫЕ ПРИНАНЕСЕНИИ РАЗДРАЖЕНИЯ НА ВЕРХНИЕ КОНЕЧНОСТИ:
• РЕФЛЕКС РОССОЛИНО - сгибание
пальцев кисти в ответ на быстрое
касание к пальцам
• РЕФЛЕКС ЖУКОВСКОГО - то же самое
при ударе молоточком по
ладонной поверхости кисти
• РЕФЛЕКС БЕХТЕРЕВА - то же самое
при ударе по тылу кисти
55.
56. Физиология ЦНС физиология ствола мозга
57. ФИЗИОЛОГИЯ СТВОЛА МОЗГА
58.
• Продолговатый мозг• Варолиев мост
• Средний мозг
59. Функции продолговатого мозга
•Рефлекторная•Проводниковая
•Тоническая
60. СРЕЗ ПРОДОЛГОВАТОГО МОЗГА
61.
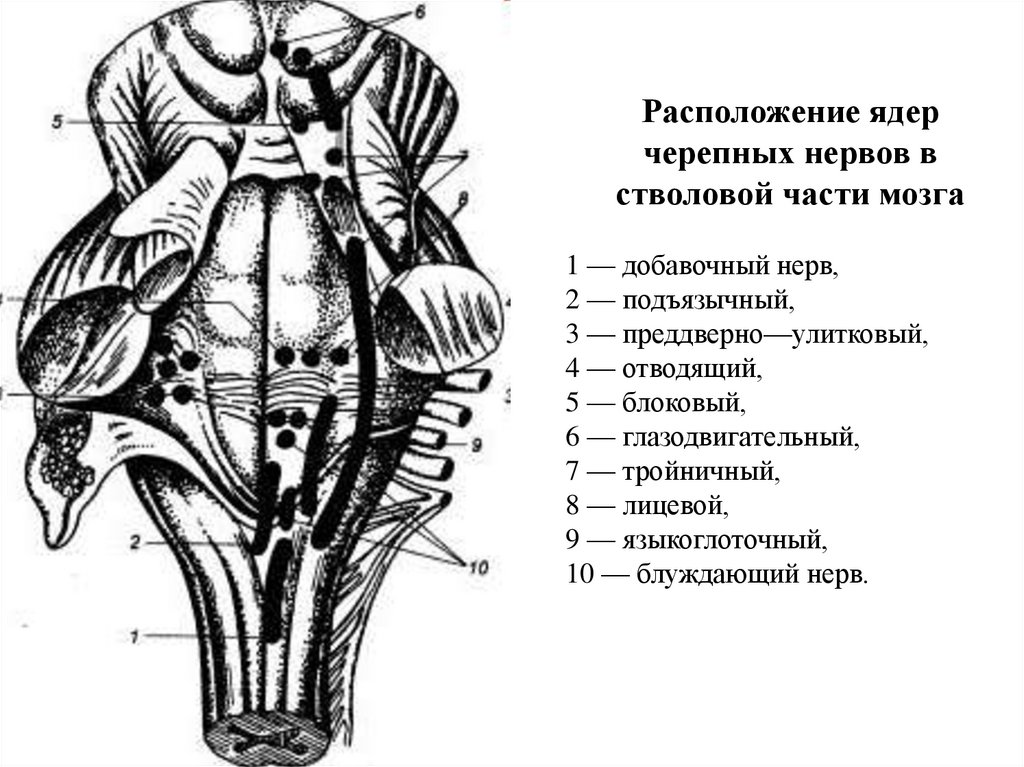
Расположение ядерчерепных нервов в
стволовой части мозга
1 — добавочный нерв,
2 — подъязычный,
3 — преддверно—улитковый,
4 — отводящий,
5 — блоковый,
6 — глазодвигательный,
7 — тройничный,
8 — лицевой,
9 — языкоглоточный,
10 — блуждающий нерв.
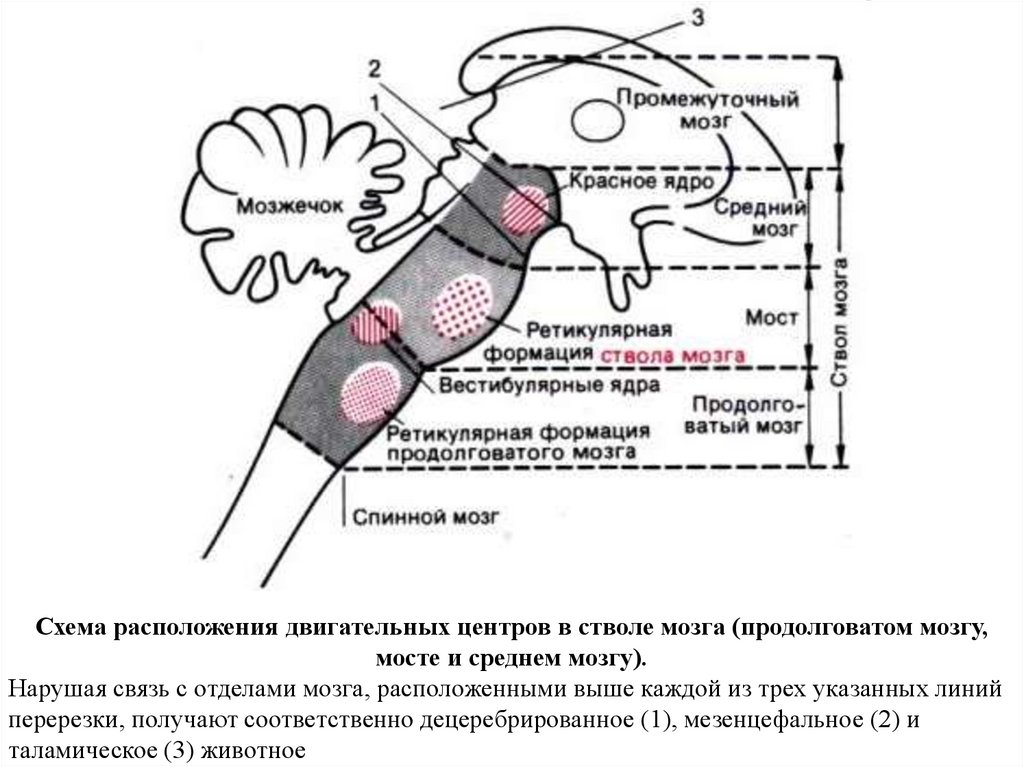
62.
Схема расположения двигательных центров в стволе мозга (продолговатом мозгу,мосте и среднем мозгу).
Нарушая связь с отделами мозга, расположенными выше каждой из трех указанных линий
перерезки, получают соответственно децеребрированное (1), мезенцефальное (2) и
таламическое (3) животное
63. Ядра продолговатого мозга
1. ЯДРА ЧЕРЕПН0М03Г0ВЫх НЕРВОВXII пара - ПОДЪЯЗЫЧНОГО НЕРВА - n.
hypoglossus - двигательные ядра
XI пара - ДОБАВОЧНОГО НЕРВА - n. accessorius
двигательные ядра
X пара - БЛУЖДАЮЩЕГО НЕРВА n. vagus:
• 1) вегетативное ядро
• 2) чувствительнее ядро одиночного пучка
• 3) обоюдное ЯДРО - двигательное глотки и
гортани
64. Ядра продолговатого мозга - 2
2. ЯДРА ЧЁРЁПНОМОЗГОВЫХ НЕРВОВ:IX пара - языкоглоточного НЕРВА:
• 1) двигательное ядро - рот и глотка
• 2) чувствительное ядро - вкус задней трети языка
• 3) вегетативное ядро - слюнные железы
На границе с мостом:
YIII пара - ВЕСТИБУЛОКОХЛЕАРНОГО НЕРВА
• 1) кохлеарные ядра
• 2) вестибулярные ядра - медиальное Швальбе,
латеральное Дейтерса, верхнее Бехтерева, ядро
Роллера
65. Ядра продолговатого мозга - 3
ПЕРЕКЛЮЧАЮЩИЕ ЯДРА:• Голля и Бурдаха – к таламусу
• Ретикулярной формации - от коры и
подкорковых ганглиев к спинному мозгу
• Оливарные ядра - от коры, подкорковых
ядер и мозжечка к спинному мозгу и от
спинного мозга к мозжечку, таламусу и
коре; от слуховым ядер в средний мозг
и четверохолмие.
66. Центры продолговатого мозга
1) Дыхательный7) Мигания
2) Сердечно-сосудистый
8) Рвоты
3) Слюноотделения
9) Сосания
4) Слезоотделения
10) Жевания
5) Кашля
11) Глотания
6) Чихания
12) Рефлексов поддержания позы
67. РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА
Жизненно-важные рефлексы
Защитные рефлексы
Рефлексы пищевого поведения
Рефлексы поддержания позы
Вегетативные рефлексы
Вестибуло-вегетативные рефлексы
68. Постуральные рефлексы
СТАТИЧЕСКИЕ - от рецепторов преддверия• рефлексы положения
• рефлексы выпрямления (установочные]
СТАТОКИНЕТИЧЕСКИЕ - от рецепторов
полукружных каналов
• рефлексы прямолинейного ускорения
• рефлексы углового ускорения
69. ВЕСТИБУЛЯРНЫЕ ЯДРА ПРОДОЛГОВАТОГО МОЗГА
• ВЕРХНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядроБехтерева
• НИЖНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Роллера
(нисходящие влияния)
• МЕДИАЛЬНОЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро
Швальбе
• ЛАТЕРАЛЬНОЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро
Дейтерса (начинается вестибулоспинальный путь)
• Также
проходит
транзитом
в
мозжечёк,
непрерываясь
в
продолговатом
мозге,
вестибулоцеребеллярный путь от афферентных
нейронов вестибулярного ганглия
70. ОСНОВНЫЕ ПРОВОДЯЩИЕ ПУТИ ОТ ВЕСТИБУЛЯРНЫХ ЯДЕР
ВЕСТИБУЛОСПИНАЛЬНЫЙ ПУТЬ — от ядра Дейтерса передает
информацию на альфа - и гамма - мотонейроны СМ, активируя мышцы
разгибатели и тормозя - сгибатели.
ВЕСТИБУЛООКУЛЯРНЫИ ПУТЬ — передает информацию на мышцы
глаза при движении, благодаря этому на сетчатке глаза сохраняется
объект наблюдения, несмотря на всевозможные перемещения.
ВЕСТИБУЛОМОЗЖЕЧКОВЫЙ ПУТЬ — передает информацию к
мозжечку и характеризует о положении тела в пространстве.
Совместно с вестибулослинальным трактом обеспечивает регуляцию
мышечного тонуса и других перемещений тела.
ВЕСТИБУЛОТАЛАМИЧЕСКИЙ ПУТЬ - обеспечивает передачу
информации к:
1.Специфическим ядрам таламуса по Лемнисковому пути, а от них к
сенсорной зоне коры, расположенной в прецентральной извилине в
области проекции лица.
2.Колатерали идут к ретикулярной формации, а от нее к
неспецифическим ядрам таламуса, от которых диффузно ко многим
участкам коры, вызываю их активацию.
71. ОБЩАЯ СХЕМА ОБРАБОТКИ И ПЕРЕДАЧИ ИНФОРМАЦИИ ВЕСТИБУЛЯРНЫМИ ЯДРАМИ
• 1. Отвестибулярного
ганглия
информация
передается в верхнее вестибулярное ядро (ядро
Бехтерева), где происходит ее обработка.
• 2. Из ядра Бехтерева, переработанная информация
оценки необходимости изменения позы, посылается к
латеральному вестибулярному ядру (ядро Дейтерса) и
медиальному вестибулярному ядру (ядро Швальбе)
• 3. От медиального (ядро Швальбе) и латерального
ядер
(ядро
Дейтерса)
информация
по
вестибулоспинальному тракту поступает к передним
рогам соответствующим сегментам СМ, обеспечивая
иннервацию
альфа
—
мотонейронов
мышц,
которые изменяют позу в данный момент времени.
72.
• VIIIпара
ЧМН
имеет
две
веточки:
вестибулярная
и
слуховая
—
являющихся
чувствительными (афферентными).
• Слуховая
ветвь
образована
афферентными
волокнами от кортиевого органа улитки и передает
информацию в нижнее двухолмие.
• Вестибулярная ветвь — в значительной мере
образована
афферентными
волокнами
идущими от
полукружных
каналов
вестибулярного анализатора, заканчивающихся в
вестибулярном ганглии, а от него к вестибулярным
ядрам продолговатого мозга.
73. ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ ОТ ВЕСТИБУЛЯРНОГО АНАЛИЗАТОРА
Информацияот
вестибулярного
анализатора (VIII пара ЧМН) поступает:
• от волосковых клеток макулы (расположенных
в маточке и мешочках — линейное ускорение,
а также наклон головы вперед или назад)
• от
волосковых
клеток
гребешков
(расположенных
в
расширенной
части
полукружных каналов — угловое ускорение
(ускорение Кориолиса), а также начало и конец
движения)
передается
на
дендриты
аффентных нейронов вестибулярного ганглия,
а
от
него
к
вестибулярным
ядрам
продолговатого мозга.
74. ЯДРО ДЕЙТЕРСА
находится под контролем мозжечка (ядро шатра),определяя состояние активности его активности и
контролирует
тонус
мышц
сгибателей
и
разгибателей.
Патология
мозжечка
проявляется
примерно
сходными
симптомами,
что
и
патология
вестибулярного аппарата и вестибулярных ядер
продолговатого мозга.
От ядра Дейтерса по вестибулоспинальному пути
информация поступает к альфа-мотонейронам СМ,
вызывая
возбуждение
альфа-мотонейронов
разгибателей и одновременно торможению альфамотонейронов мышц сгибателей.
Благодаря этому меняется тонус мышц верхних и
нижних конечностей — и сохраняется равновесие
75.
От вестибулярных ядер продолговатого мозгаинформация передается к медиальному продольному
пучку, который начинается от ядра Даршкевича и
промежуточного ядра среднего мозга и ведет в
сторону спинного мозга, соединяя в единый
функциональный ансамбль все ядра нервов,
участвующих в регуляции активности мышц глазного
яблока (III пара) глазодвигательных, IV пара (
блоковый), VI пара (отводящий) ЧМН.
Благодаря этому, движение обоих глаз происходит
синхронно и в норме у большинства людей
отсутствует косоглазие.
С участием продольного медиального пучка
осуществляется
явление
НИСТАГМА глазных яблок — т.е. движения глаз,
которые
способствуют
поддержанию взора на точке фиксации — на
сетчатку,
изображение
падает
примерно на одно и тоже место, не смотря на то, что
человек совершает движения.
76. При патологии вестибулярного аппарата или вестибулярных ядер и вестибулярного тракта, возникают такие симптомы, как
• головокружение (субъективное осущениевращение предметов окружающего мира,
которое возникает вследствие
• «НЕ ВЕРНОЙ ИНФОРМАЦИИ, ПОСТУПАЮЩЕЙ В
АССОЦИАТИВНЫЕ УЧАСТКИ КОРЫ»),
• нарушения
равновесия
(атаксия
статическая
и
статокинетическая)
возникающий спонтанно НИСТАГМ глаз:
- горизотальный
- вертикальный
- или ротационный
77. ЯДРА РЕТИКУЛЯРНОЙ ФОРМАЦИИ ЗАДНЕГО МОЗГА (ПМ И МОСТ), участвующих в регуляции двигательной активности:
• 1. Нейроны продолговатого мозга —активируют альфа-мотонейроны сгибателей и
тормозят альфа-мотонейроны разгибателей, т.е.
ведут себя как нейроны Красного ядра.
• 2. Нейроны ретикулярной формации моста
активируют — альфа-мотонейроны разгибателей
и тормозят альфа-мотонейроны сгибателей, т.е.
действуют как нейроны вестибулярных ядер (ядро
Дейтерса)
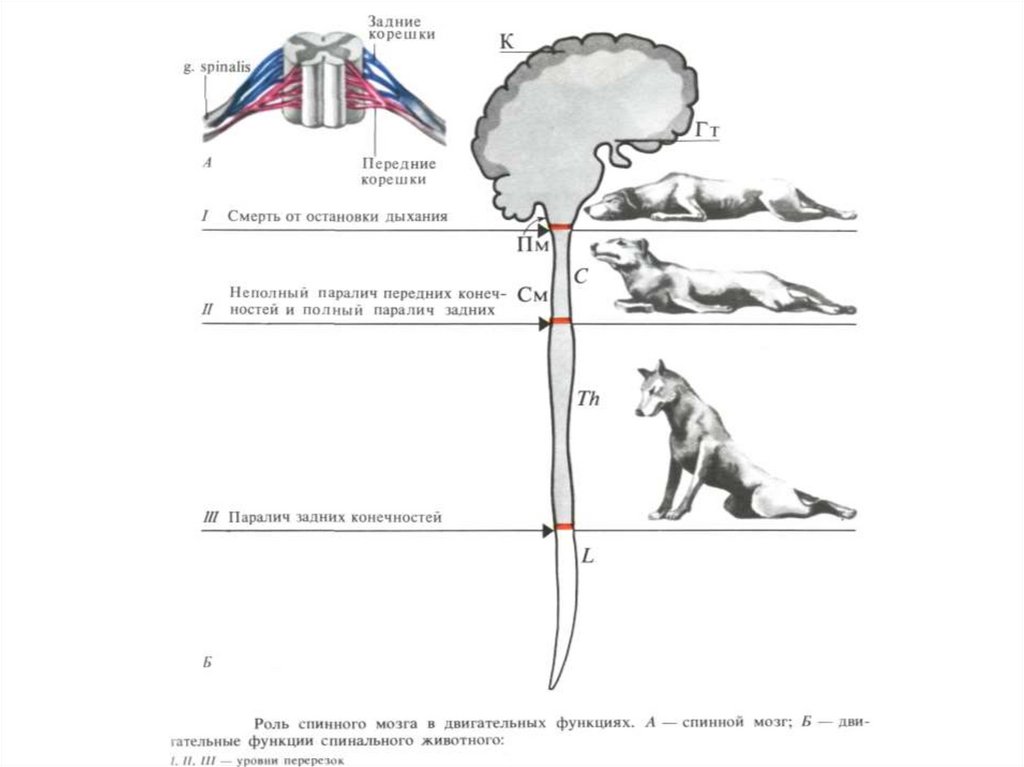
78. Роль продолговатого мозга в двигательных функциях
79. ФИЗИОЛОГИЯ СРЕДНЕГО И ПРОМЕЖУТОЧНОГО МОЗГА
80. ПРОДОЛГОВАТЫЙ МОЗГ, МОСТ И СРЕДНИЙ МОЗГ
81. Средний мозг
82. ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ
83. Основные ядра среднего мозга
• Ядра черепномозговых нервов:• --- III пара - глазодвигательного нерва
--- IV пара - блокового нерва
--- Ядро Даркшевича - продольный пучок среднего мозга,
связывающий ядра глазодвигательного, блокового и отводящего
нерва в единую систему
• Непарное вегетативное ядро Якубовича-Эдингера - через
цилиарный ганглий к мышцам радужки и ресничного тела
• Ядра тектальной области: верхнее или переднее двухолмие зрительные рефлексы; нижнее или заднее двухолмие - слуховые
рефлексы - четверохолмие
• Черная субстанция
• Красные ядра
84. ДВИГАТЕЛЬНЫЕ ЯДРА СТВОЛА МОЗГА
Можно условно разделить на двакласса по их влиянию на мышцы:
• флексорные системы — повышающие
активность сгибателей — нейроны
Красного
ядра
и
ретикулярной
формации ПМ.
• экстензорные системы — повышающие
активность разгибателей — нейроны
вестибулярных ядер и ретикулярной
формации моста
85. Стволовые рефлексы тонуса мышц, позы и равновесия тела.
• Осуществляетсяс
вестибулярных,
проприоцентивных,
тактильных и зрительных рецепторов через двигательные
центры ствола, включая центры ретикулярной формации (РФ).
• Эти эффекторные влияния из моторных центров ствола
распространяются на мускулатуру конечностей и туловища по
четырем нисходящим путям:
1. вестибулярному
2. медиальному ретикулоспинальному
[1 и 2 – состовляют медиальную нисходящую экстензорную
(разгибательную) систему]
3. руброспинальному
4. латеральному ретикулоспинальному
[3 и 4 составляют латеральную нисходящую флексорную
(сгибательную) систему].
86. Медиальная разгибательная система
Вестибулоспинальный путь – от латеральныхвестибулярных
ядер,
которые
получают
афферентную
импульсацию
от
рецепторов
отолитового аппарата и ампул полукружных
каналов.
Волокна
этого
пути
возбуждают
спинальные α- и γ- мотонейроны мышцразгибателей конечностей, туловища, шеи и
одновременно, через тормозные интернейроны
тормозят сгибатели.
Медиальный ретиколоспинальный путь – от
ретикулярных
ядер
моста.
Через
них
осуществляется стимуляция α- и γ- мотонейронов
мышц-разгибателей осевой мускулатуры тела
(туловища и проксимальных отделов конечностей), и
торможение сгибателей.
87. Латеральная сгибательная система
Руброспинальный путь – от красного ядра покрышкимозга, возбуждает α- и γ- мотонейроны сгибателей
и реципрокно тормозят мотонейроны разгибателей.
Латеральный ретикулоспинальный путь – от
гигантоклеточного
ретикулярного
ядра
продолговатого мозга, его волокна возбуждают α- и
γ- мотонейроны мышц-сгибателей и реципрокно
тормозят мышцы-разбибатели.
Вместе с тем раздражение нейронов вентральной
части
РФ
продолговатого
мозга,
вызывает
подавление
всех
спинальных
рефлексов
(сгибательных и разгибательных) и ликвидацию
децеребрационной
ригидности.
Поэтому
механизму
осуществляется
центральное
торможение рефлексов СМ, открытое И.М.
Сеченовым (1862г.)
88. КРАСНОЕ ЯДРО
Расположено в области среднего мозга, егонейроны получают информацию из моторной
зоны
коры больших
полушарий,
промежуточных ядер мозжечка
(пробковидного и шаровидного) и от нейронов
черной
субстанции,
т.
о.
имеют
всю
информацию о положении тела в прострастве,
о состоянии мышечной системы и кожи. По
рубраспинальному
тракту передает информацию
в спинальные центры, контролирующие мускулатуру
верхних
и
нижних
конечностей.
89. НЕЙРОНЫ КРАСНОГО ЯДРА -
НЕЙРОНЫ КРАСНОГО ЯДРАЧерез руброспинальный тракт оказывает активирующее
влияние на альфа — мотонейрона мышц сгибателей и
тормозят активность
альфа-мотонейронов
разгибателей,т.о.
уравновешивают
активность
вестибулярного ядра Дейтерса, а совместно с ним
участвуют в регуляции позы.
Отделение красного ядра от нижележащих структур
(перерезка выше моста) ведет к повышению тонуса мышц
разгибателей
(экстензоров)
и
развитию
ДЕЦЕРЕБРАЦИОННОЙ ригидности -в этих условиях
разрушение
ядра
Дейтерса
или
перерезка
вестибулоспинального тракта, ведет к исчезновению
ригидности.
В
развитии
ДЕЦЕРЕБРАЦИОННОЙ
ригидности
участвует гамма - петля, т. к. ДЕАФФЕРЕНТАЦИЯ
конечности устраняет децеребрационную ригидность.
90.
Децеребрационная ригидность.а — кошка с выключенными большими полушариями (путем
перевязки сосудов); б — то же при одновременном выключении
мозжечка (по Дэвису и Поллоку, из И. С. Беритова).
91. ЧЕРНАЯ СУБСТАНЦИЯ
Получает обильную информацию от базальныхганглиев и участвует в координации движений и
эмоций.
При разрушении развивается болезнь
ПАРКИНСОНА -нарушается координация тонких
движений пальцев рук, мышечной ригидности,
тремор.
92. Статические рефлексы
Их ролью яв-ся сохранение естественной позы человека и животных- Это тонические рефлексы в покое (человек стоит и сидит)
- В том числе и при поворотах и наклонах головы и движения рук, а
сидя – и ног, вызывающих смещение центра тяжести.
Осуществляются с участием:
- Сегментарных и надсегментарных (шейных тонических рефлексов СМ.)
- Рефлексов ствола мозга, возникающих с вестибулорецепторов
Р. Магнус:
- для выполнения роли шейных рефлексов разрушал у животного
вестибулярный аппарат, чтобы устранить вестибулярные рефлексы.
- Для выяснения роли вестибулярных рефлексов, на шею животного,
расположенную на одной линии с туловищем, накладывали гипсовую
повязку, чтобы прекратить шейные надсегментарные рефлексы.
Вестибулярные рефлексы выявляются при поворачивании животного
через ось висков.
- При горизонтальном расположении животного спиной и затылком
вниз поворот головы от 0 до 45 градусов выше горизонтали приводит к
максимальному повышению тонуса мышц-разгибателей.
- При дальнейшем повороте, если оно располагается спиной и затылком
вверх по горизонтале и до -45 градусов ниже горизонтали, то тонус
разгибателей становиться минимальным.
У человека, в положении стоя (голова вверх, шея наклонена несколько
вперед, руки вдоль туловища) шейные и лабиринтные рефлексы
усиливают друг друга, и происходит повышение тонуса разгибателей
нижних конечностей и сгибателей верхних конечностей.
93.
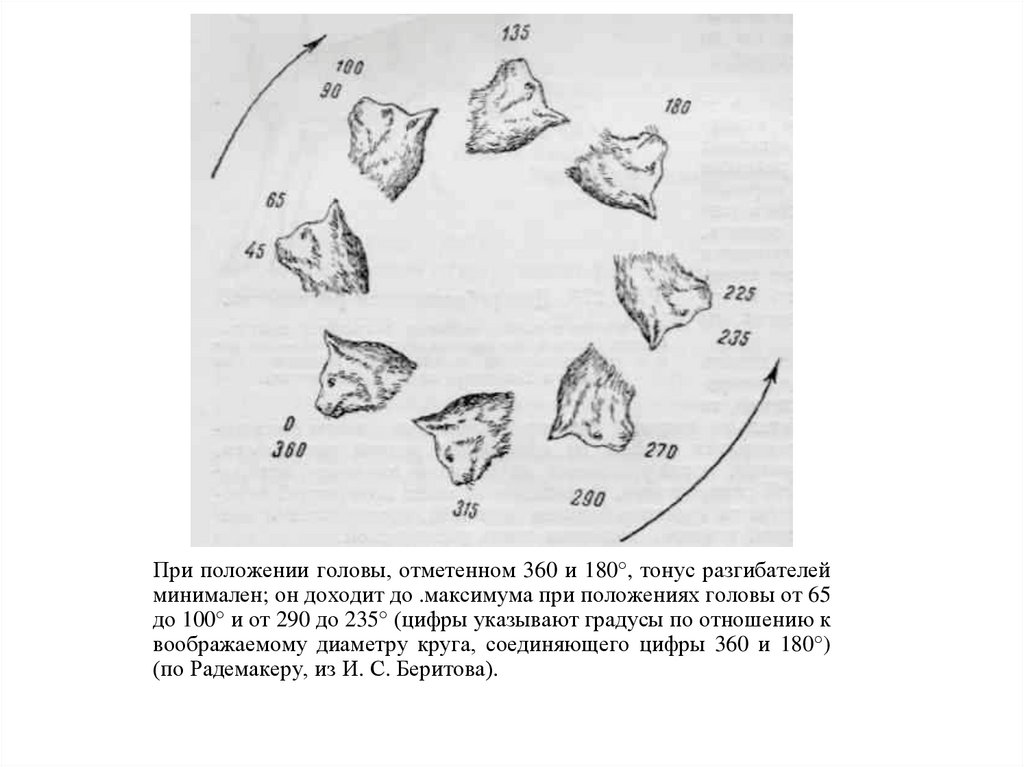
Положения головы, вызывающие минимум (а)и максимум (б) экстензорного тонуса (по Магнусу).
94.
При положении головы, отметенном 360 и 180°, тонус разгибателейминимален; он доходит до .максимума при положениях головы от 65
до 100° и от 290 до 235° (цифры указывают градусы по отношению к
воображаемому диаметру круга, соединяющего цифры 360 и 180°)
(по Радемакеру, из И. С. Беритова).
95. Статокинетические рефлексы
Их роль – восстановление нарушенной позы
Это рефлекторное перераспределение мышечного тонуса, возникающее с:
Кожных
Проприо – (шея)
Вестибулорецепторов
При нарушении позы (организм находится в горизонтальном положении).
К этим рефлексам относятся выпрямительные (установочные) рефлексы - выпрямление
головы и туловища
Выпрямление головы запускается с:
Вестибулярных и кожных рецепторов
Выпрямление туловища запускается с:
Проприорецепторов шеи и кожных рецепторов.
Т.е. имется две рефлесогенных зоны, поэтому выключение одной из зон (или рецепторов
кожи или вестибулярного аппарата) у мезэнцефального организма не нарушает
выпрямительные рефлексы. Выключение обеих названных рефлексогенных зон приводит к
потере выпрямительных рефлексов.
Это надсегментарные рефлексы ствола мозга и спинного мозга. Особо важную роль играют
Красные ядра – при их разрушении нарушенная поза не восстанавливается.
У человека и обезьян выпрямительные рефлексы осуществляются при обязательном участии
КБП – ее выключение приводит к потере этих рефлексов.
Р. Магнус – относил эти рефлексы – к статическим, однако они имеют две состовляющие –
статическую (нет перемещения тела в пространстве или вращения вокруг своей оси на
месте) и кинетическую – животное поднимается и занимает естественную позу.
96.
Поза лежащей на левом боку обезьяны, у которой кора мозга удалена при сохранениизрительных бугров и части стриопаллидарной системы. Если животное переложить на
правую сторону, то разогнуты будут правые конечности, а согнуты левые. В сгибании кисти
правой руки ясно виден «хватательный» рефлекс (по Фултоиу)
97. Кинетические рефлексы
Рефлекторное перераспределение мышечноготонуса при ходьбе и беге, вращательном движении на
месте, при изменении скорости движения в транспорте
и лифте
Возникают с:
• Проприорецепторов
• Вестибулорецепторов
Назначение этих рецепторов – сохранение
естественной позы при движении, а сокращения мышц
направлены
на
преодоление
сил
гравитации,
изменяющихся в следствии ускорения или замедления
движения
Важную роль играет система зрения – в ходе
вращения наблюдается поворот головы и нистагм глаз
(пилообразные движения глаз)
98. Рефлексы вращения – 1
Возникают с ампул полукружных каналов и включают вперераспределение тонуса мускулатуру тела и глазные мышцы.
Важным для практической медицины является нистагм – непроизвольные
ритмические двухфазные движения глаз, который имеет две фазы:
• При ускорении вращения сначала происходит медленное отклонение
глазных яблок в сторону, противоположную вращению
(сохранение
имеющегося поля зрения)
• После этого они быстро отклоняются обратно – по ходу вращения (захват
нового поля зрения)
Медленный компонент вызывается с рецепторов полукружных
каналов
Быстрый компонент связан с влиянием центров зрительной (поле
19) и лобной (поле 8) коры, действующих через верхнее двухолмие,
моторные центры III, IV, VI нервов, и ретикулярную формацию ствола (при
наркозе он исчезает). Во врачебной практике направление нистагма
определяют по быстрому компоненту.
При замедлении вращения, напротив, медленное движение глаз
направлено в сторону вращения, и быстрое – против направления вращения.
Вид нистагма (горизонтальный, вертикальный, ротационный) –
зависит от полукружного канала, находящегося в плоскости вращения и
служит для оценки функционального состояния вестибулярного аппарата.
У здорового человека глазной нистагм может возникать и без
вращения: при длительной фиксации взгляда на близкорасположенном
предмете, а также при крайнем отведении глазных яблок (более 45 градусов)
99. Рефлексы вращения – 2
В реализации рефлексов вращательного движения, кромезрительного, участвуют волосковые клетки полукружных
каналов вестибулярного аппарата:
- При вращении – эндолимфа по инерции «отстает» от движения
костных стенок канала и в результате усиливается ее
давление на тот конец канала, который противоположен
направлению вращения.
- При остановке ее давление возрастает в области другого конца
канала.
• Этот сигнал от вестибулярного анализатора идет через
вестибулярные ядра на медиальный продольный пучок к
ядрам III, IV и VI пары ЧМН и приводит к изменению положения
в пространстве глазных яблок – что сохраняет ориентацию в
пространстве.
• Одновременно – через вестибулярный тракт меняется тонус
мышц конечностей и туловища, что сохраняет равновесие.
100.
Лифтные рефлексывозникают при подъеме вверх –
повышается тонус сгибателей, и животное
приседает, а при опускании вниз –
возрастает тонус разгибателей и животное
как бы приподнимается.
Кроме зрительных рецепторов, в этом
рефлексе
участвуют
волосковые
клетки
преддверия вестибулярного аппарата, т.к.
рефлексы сохраняются и при закрытых глазах
– подъем в самолете.
При подъеме тела – отолиты более
обычного давят на волосковые клетки, а в
конце подъема – их давление снижается.
101.
Локомоторная функция ствола мозга• лежит в основе совокупности согласованных
движений, с помощью которых организм
перемещается в пространстве (ходьба).
• Большая роль отводится клинообразному
ретикулярному ядру в задненижнем отделе
среднего
мозга,
т.н.
локомоторную
область
среднего
мозга,
имеющую
эффекторный выход к ядрам моста.
• Так
же
в
эту
систему
входит
субмамиллярная зона гипоталамуса. У
человека локомоция без участия КБП –
невозможна.
102.
103. Бульбарный организм (перерезка между мостом и средним мозгом) – развивается децеребрационная регидность, хотя имеются
ЖИВОТНЫЕ С ПЕРЕЗКОЙ НА РАЗЛИЧНЫХ УРОВНЯХ ЦНСБульбарный организм (перерезка между
мостом и средним мозгом) – развивается
децеребрационная регидность, хотя имеются
статические позно-тонические рефлексы
шейные, лабиринтные), но отсутствуют
выпрямительные рефлексы, локомоция,
произвольные движения.
Мезэнцефальный организм – имеется
нормальный мышечный тонус, но у него
отсутствуют произвольные движения.
104.
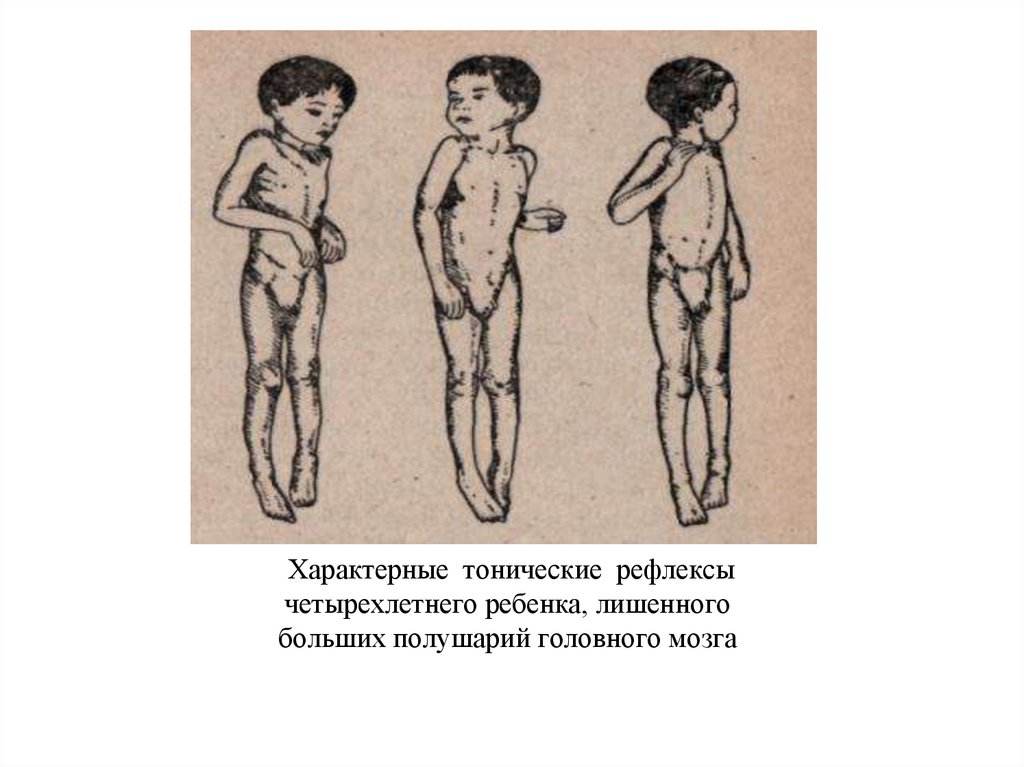
105.
Характерные тонические рефлексычетырехлетнего ребенка, лишенного
больших полушарий головного мозга
106.
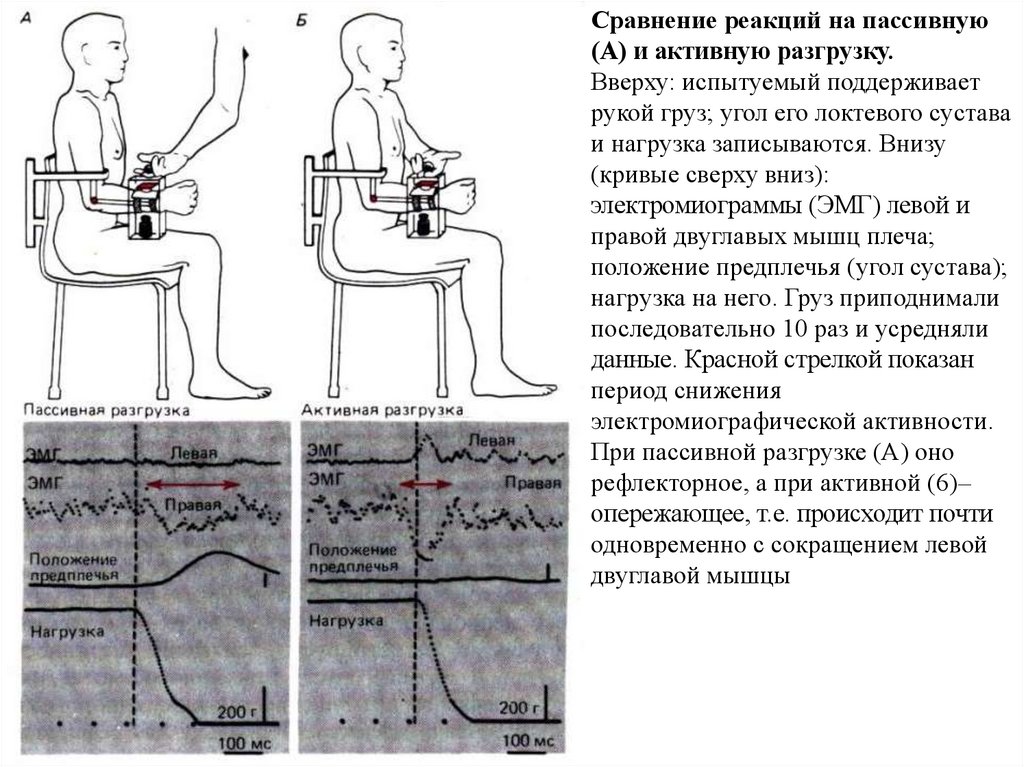
Сравнение реакций на пассивную(А) и активную разгрузку.
Вверху: испытуемый поддерживает
рукой груз; угол его локтевого сустава
и нагрузка записываются. Внизу
(кривые сверху вниз):
электромиограммы (ЭМГ) левой и
правой двуглавых мышц плеча;
положение предплечья (угол сустава);
нагрузка на него. Груз приподнимали
последовательно 10 раз и усредняли
данные. Красной стрелкой показан
период снижения
электромиографической активности.
При пассивной разгрузке (А) оно
рефлекторное, а при активной (6)–
опережающее, т.е. происходит почти
одновременно с сокращением левой
двуглавой мышцы
107. Позно-тонические рефлексы у человека
ярко выражены в первое полугодие жизни, так у грудных детей:• Лабиринтный тонический рефлекс – у ребенка лежащего на
спине, повышен тонус разгибателей шеи, спины, ног. Если
ребенок лежит на животе, то повышен тонус сгибателей шеи,
спины, конечностей.
• Симметричный шейный тонический рефлекс – при пассивном
сгибании головы ребенка, лежащего на спине, происходит
повышение тонуса сгибателей рук и повышение тонуса
разгибателей ног. При разгибании головы наблюдаются
противоположные процессы.
• Ассиметричный шейный тонический рефлекс – у ребенка
лежащего на спине голову поворачивают в сторону так, чтобы
подбородок касался плеча, при этом снижается тонус сгибателей
конечностей в сторону поворота и повышается тонус
разгибателей. На противоположной стороне, на руке и ноге,
возрастает тонус сгибателей.
Все эти рефлексы тормозятся у детей ко 2-3 месяцу, если они
сохраняются в возрасте одного года, это может свидетельствовать
о замедлении развития моторных центров головы.
108. В онтогенезе
преобладает тонус мышц сгибателей (спинальный)- Определяющий адекватную позу у плода, т.е. максимальную
сгибательную активность в суставах позвоночника и конечностей.
- У новорожденного наличие примитивных сгибательных
рефлексов (рефлекс Моро – симметрические обхватывающие
движения ручек при действии различных стимулов)
Появление разгибательного тонуса – обусловлено более поздним
созреванием структур продолговатого мозга.
- У детей, только на второй месяц появляется рефлекс Ландау –
ребенок лежа на животе, приподнимает голову, опираясь на
предплечья, а позже и на кисти.
- на 5-6 месяц, лежа на животе, ребенок разгибает ноги и
приподнимает таз, что создает готовность к ползанью.
Так постепенно флексорная поза, сменяется – экстензорной, при
этом увеличивается пространство видимое глазами и
обогащающая жизненный двигательный опыт ребенка.
109. Сторожевой и ориентировочный рефлексы
осуществляются с участием четвертохолмия, вответ на зрительный сигнал (световой) или на
звук происходит активация соответствующих
рецепторов, сигнал достигает первичных
центров (зрительные – передних или верхних
бугров, слуховые – задние или нижние бугры),
переключаются на тектоспинальный тракт –
возникают
ориентировочные
рефлексы:
поворот головы в сторону, откуда поступает
сигнал
и
одновременно
происходит
перераспределение тонуса мышц, в том числе
за
счет
изменении
положения
головы,
благодаря которому сохраняется равновесие и
возникают условия для совершения рефлекса
«бег или борьба».
110. Роль ЧМН ствола
• С участием ядер V, IX, X, XI и XII паросуществляется произвольные движения
типа жевания (V пара) и глотания
(последовательное
возбуждение
αмотонейронов
языкоглоточного,
вагуса,
добавочного
и
подъязычного
нервов).
Повреждение – нарушаются акты жевания и
глотания.
• С участием нейронов IX и X пары идет
управление голосовыми связками, а с
помощью XII пары – языком. Повреждение
этих структур – нарушается фонация и
артикуляция (речь).
111. Проводниковая функция ствола мозга
Восходящие (афферентные) пути являются частьюпроводникового отдела анализаторов передающих
информацию в сенсорную зону КБП.
Выделяют:
• Специфическую
• Неспецифическую
Специфическая – составляет
лемнискоталамический путь, в котором выделяют:
• Медиальную петлю
• Латеральную петлю
• Функционально-проводящие пути (т.к. не входит в
состав лемнисков) зрительного анализатора –
переключаются в латеральном коленчатом теле
таламуса.
112. Медиальная петля
Преимущественно образуется из аксонов нейронов ядер:1)Тонкого (Голля) ядра
2) Клиновидного (Бурдаха) ядра
• (1+2 - проводят информацию от рецепторов
конечностей, туловища и шеи, тактильную и
висцеральную чувствительность)
3)Тергеминоталамический путь (от ядер V нерва)
проводит проприоцептивную и тактильную
чувствительность от головы
4) Одиночного пути на подходе к таламусу – проводят
вкусовую и висцеральную чувствительность
5) Бокового спиноталамического пути (небольшая
часть) – проводят острую (быструю) локализованную
боль. Волокна этой петли – переключаются
преимущественно в вентральных задних
специфических ядрах таламуса.
113. Латеральная петля (слуховая петля)
Сформирована аксонами он нейронов ядер:• Трапецевидного тела
• Верхней оливы моста
Входящих в проводниковый
отдел
слухового
анализатора,
переключается
в
медиальном
коленчатом теле таламуса.
Функционально – обеспечивает быструю передачу
информации с четким различением свойств
раздражителя, с формированием в КБП первичного
ответа вызванных потенциалов соответствующего
анализатора
114.
Неспецифическая(экстралемнисковая)
–
преимущественно волокна:
• Бокового спиноталомического пути
• Спиноретикулярного пути (переключаются в ядрах РФ)
• Часть волокон тригеминоталомического пути
Проводят температурную и болевую чувствительность.
Переключаются в неспецифических (интраламинарных и
ретикулярном) ядрах таламуса, прое5цируются в различные
области КБП, особенно в лобную орбитальную кору.
Функционально – обеспечивают медленную передачу
информации нечеткой локализацией раздражителя и плохим
анализом его свойств, с формированием в КБП вторичного
ответа вызванных потенциалов.
Неспецифическая система – получает коллатерали от
специфической системы, что обеспечивает тесную связь этих
двух афферентных систем.
115.
К афферентной (проводящей) системе ствола такжеотносятся пути передающие сенсорную информацию
в кору мозжечка:
• Дорсальный спиномозжечковый путь – от
рецепторов мышц и связок
• Вентральный спиномозжечковый путь – от
рецепторов сухожилий, кожи и внутренних органов
• Вестибуломозжечковый путь – от вестибулярных
рецепторов
Из коры мозжечка – к его зубчатым и
промежуточным ядрам, далее по
церебеллоталамическому пути к двигательным
вентральным ядрам таламуса и после
переключения в них в соматосенсорную, моторную
и премоторные зоны КБП.
116. Нисходящие проводниковые пути ствола мозга
Функционально объединяются:• Пирамидный (двигательный) путь – от клеток Беца КБП прецентральной
извилины – переключаются на мотонейроны передних рогов спинного
мозга (кортико-спинальный) путь или двигательных ядер ЧМН (кортикобульбарный путь) – обеспечивают произвольные сокращения мышц
конечностей, туловища, шеи и головы.
Экстрапирамидные – кортико-рубральный и кортико-ретикулярный –
оканчиваются на моторных центрах ствола – от которых идут руброспинальный и ретикулоспинальный пути. Основная функция –
произвольная регуляция мышечного тонуса, позы и равновесия.
Кортико-мостомозжечковый путь – в мозжечок поступает информация
от двигательной и других областей КБП – обработанная в коре мозжечка
и его ядрах информация поступает на моторные ядра ствола (красное,
вестибулярные, ретикулярные), что включает мозжечок в функции
экстрапирамидной системы.
Тектоспинальный путь – от ядер четвертохолмия – обеспечивает
двигательные реакции организма в ориентировочных, зрительных и
слуховых рефлексах.
117. Нейрогуморальная функция ствола мозга
осуществляется за счет выработки нейронаминейромодуляторов, которые действуют как «регуляторы
нейромедиаторов»
• Норадренергическая система ствола мозга – медиатор
норадреналин, имеется в двух ядрах:
1. Голубом пятне среднего мозга
2. Вентральное ядро ретикулярной формации продолговатого
мозга
Главной структурой этой системы является голубое пятно,
содержащее более 50 % всех норадренергических нейронов
ствола, имеющих обширные афферентные и эфферентные
связи со всеми отделами ЦНС.
Учувствуют в контроле проведения болевой и неболевой
чувствительности, активности моторных систем, в регуляции
деятельности висцеральных систем, эмоционального
состояния тоски и страха, механизмов памяти и внимания, фаз
цикла «сон – бодрствование», индуцируя фазу быстрого сна, в
формировании (через гипоталамус) центрального звена
стрессформирующей системы, участвуя в развитии общего
адаптационного синдрома.
118.
Серотонинергическая система ствола мозга• образована скоплением серотонинергических
нейронов, образующих ядра шва, центральное серое
вещество и ретикулярные магноцеллюлярные ядра.
• участвует в контроле проведения болевой
чувствительности, в регуляции цикла «сон –
бодрствование», потребления пищи и воды,
тормозит сексуальное и агрессивное поведение.
Дофаминергическая система ствола мозга
• Представлена - черным веществом среднего мозга,
в функциональном плане относится к
стриопаллидарной системе. Нейроны черного
вещества (медиатор дофамин) оказывают влияние на
полосатое тело приводя к двигательным
расстройствам типа Паркинсонизм.
119. Интегративная функция ствола мозга
Осуществляется его ретикулярной формацией (РФ),нейроны которой расположены диффузно или в виде ядер
в центральных отделах ствола мозга:
• Нейроны РФ имеют длинные, мало ветвящиеся дендриты
и
хорошо
ветвящиеся
аксоны,
которые
могут
устанавливать контакты с 25 тыс. других нейронов
• Нейроны РФ принимают информацию от различных
сенсорных путей, они полисенсорные, имеют большие
рецепторные поля
• Имеют длительный латентный период на периферическую
стимуляцию, за счет многочисленных синапсов
• Фоновоактивны (5-10 Гц), высоковозбудимы и лабильны
• Чувствительны к веществам крови – адреналину, СО2 и
лекарственным препаратам – барбитураты и аминазин
120. Ретикулярная формация ствола мозга
121.
В 1946 г. американский нейрофизиологХ.Мегоун обнаружил, что РФ ствола мозга имеет
непосредственное отношение к регуляции не
только
вегетативной,
но
и
соматической
рефлекторной деятельности.
122.
Х.Мегоунпоказал,
что
раздражение
вентрокаудальной части гигантоклеточного ядра
продолговатого мозга вызывает ослабление в
одинаковой мере всех спинальных двигательных
рефлексов: и сгибательных, и разгибательных.
Позже было обнаружено, что раздражение более
дорсальных и оральных ее участков вызывало,
наоборот, диффузное облегчающее действие на
спинальную рефлекторную деятельность.
123.
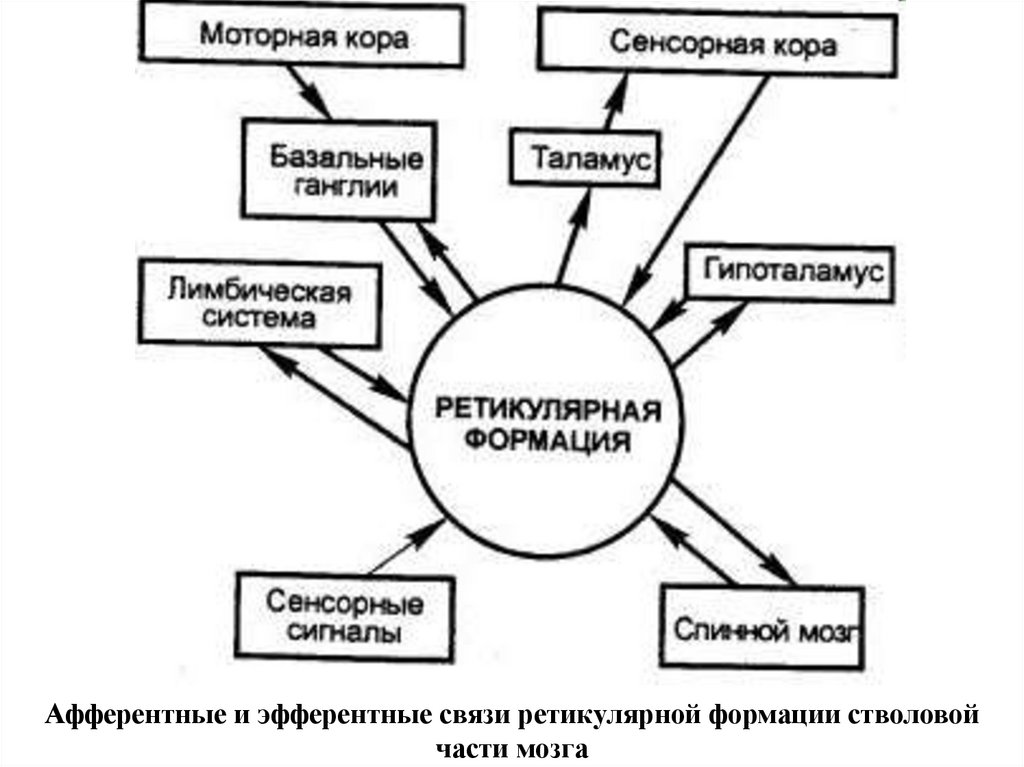
Афферентные и эфферентные связи ретикулярной формации стволовойчасти мозга
124. Основные системы ретикулярной формации
АФФЕРЕНТНЫЕ СИСТЕМЫ:
от спинного мозга, от мозжечка, от четверохолмия, от коры,
от чувствительных ядер черепномозговых нервов
ЭФФЕРЕНТНЫЕ СИСТЕМЫ:
Восходящая активирующая система неспецифическое тонизирование через синапсы на
дентритах нейронов I и II слоев коры
• Нисходящие ретикулоспинальные системы: облегчающая
тормозящая
125. Афферентные входы РФ
Преимущественно образованы от 3 источников:• От температурных и болевых рецепторов
по волокнам спиноретикулярного пути и
тройничного нерва.
• От моторных и сенсорных областей КБП по
кортикоретикулярным путям импульсация
идет в ядра, дающие начало
ретикулоспинальным путям, а так же
проецируется в мозжечок
• От ядер мозжечка по
мозжечковоретикулярным путям
126. Эфферентные выходы РФ проецируются:
• В спинной мозг по латеральному имедиальному ретикулоспинальным путям.
• К неспецифическим ядрам таламуса,
заднему гипоталамусу, полосатому телу по
восходящим путям, которые начинаются в
ядрах РФ продолговатого мозга и моста.
• К мозжечку, по путям, которые начинаются в
латеральном и парамедиальном
ретикулярных ядрах и в ядре покрышки
моста.
127. Интеграция соматических функций:
• Проявляется в координирующем влиянии РФ, посредствомкоторой отдельные рефлексы ствола мозга объединяются в
сложные двигательные акты.
• Имеет специализированные ядра, участвующие в
осуществлении сложных форм двигательного поведения:
жевания, глотания, содружественного движения глаз в
горизонтальном и вертикальном направлениях, локомоции, пения
и др.(генератор ритма жевания)
• Осуществляют контроль передачи сенсорной информации на
различных уровнях ЦНС, так:
• На уровне первичных афферентов нейронов V (тройничного)
ядра передача информации может облегчатся или тормозиться
при раздражении ретикулярного гигантоклеточного ядра
продолговатого мозга
• В ядрах задних столбов СМ сенсорный поток контролируются
тормозными влияниями из РФ продолговатого мозга.
• Восходящая РФ контролирует передачу информации в сенсорных
специфических ядрах таламуса, определяя объем и качество
передаваемой в головной мозг информации
128.
В 1949 г. Х.Мегоун и итальянский нейрофизиологДж.Моруцци показали, что раздражение РФ
эффективно влияет и на функции высших структур
мозга, в частности КБП, определяя переход ее в
активное (бодрствующее) или неактивное (сонное)
cостояние.
129. Восходящие влияния РФ на большой мозг
Может быть активирующим и тормозным,реализуется 5-17% нейронов РФ,
формирующими ретиколоталамические пути,
а после переключения в таламусе сенсорный
поток проектируется в различные области
коры.
- кроме таламуса восходящее влияния идут в
задний гипоталамус, полосатое тело,
прозрачную перегородку.
Выделяют две части
- ростральную – образуемую нейронами РФ
среднего мозга
- каудальную – образуемую нейронами РФ
продолговатого мозга и моста
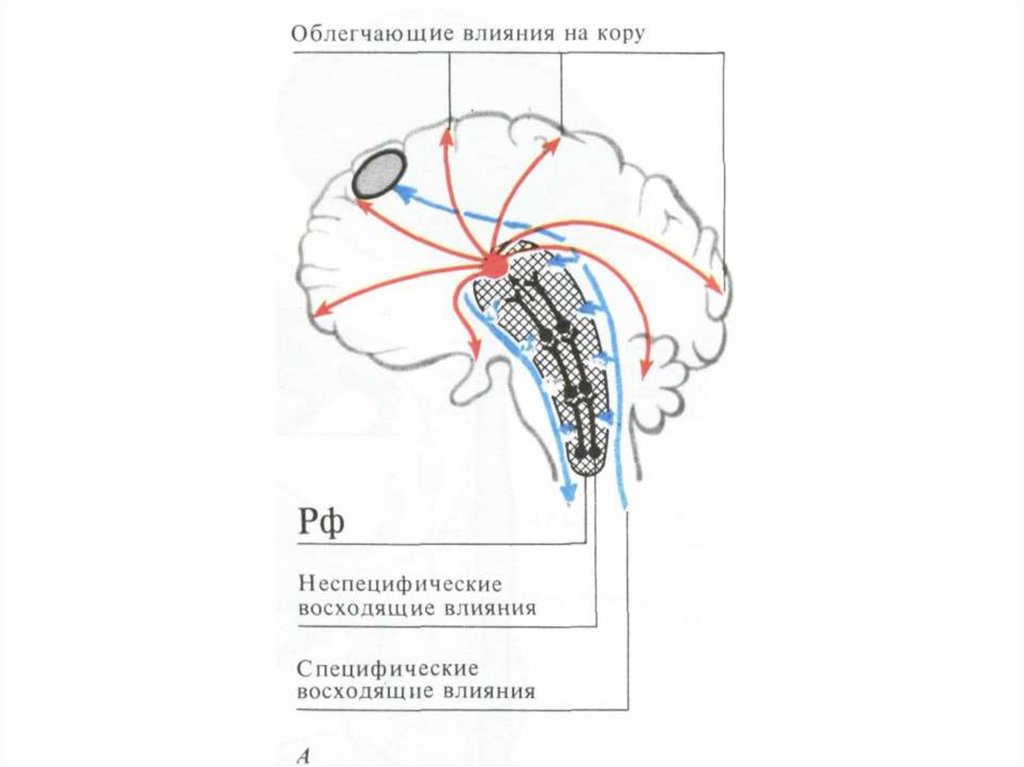
130. Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи с корой и подкоркой
ОБЛЕГЧАЮЩИЕ ПУТИ:5 – спиноретикулярный,
6 -вестибулоспинальный
ТОРМОЗЯЩИЕ ПУТИ:
1 – кортикоретикулярный,
2- каудатоспинальный,
3 – мозжечковретикулярный,
4 -ретикулоспинальный
131. Схема восходящих влияний РФ на кору мозга
Восходящаяактивирующая
система РФ
Первичный ответ специфической проекционной зоны
Вторичный ответ в зонах неспецифической стимуляции
132. Активирующее влияние на КБП
оказывает ростральная часть РФОпыт Бренера (1935г.) – перерезка ствола мозга
между верхним и нижним холмиками
четверохолмия вызывает снопоподобное
состояние – нарушен контакт с внешним миром,
отсутствие реакции на свет и звук («спящий мозг»)
Х.Мегун и Дж.Моруцци (1949 г.) – раздражение РФ у
спящих животных вызывало их пробуждение,
появление ориентировочной реакции и появление
высокочастотного β – ритма ЭЭГ (реакция
десинхронизации), что доказывает генералзованное
активирующее действие РФ на КБП.
Важнейшая функция активирующей возходящей РФ
яв-ся регуляция цикла «сон - бодрствование» и
уровня сознания.
133.
Бельгийский нейрофизиолог Ф.Бремер (1935),проводя перерезку головного мозга на различных
уровнях и наблюдая за функциями отделенных от
остальной ЦНС участков головного мозга, обратил
внимание на то, что имеется чрезвычайно
существенное различие между животным, у
которого
перерезка
проведена
по
межколликулярному
уровню
(т.е.
между
передними и задними холмами четверохолмия), и
животным, у которого линия разреза прошла между
продолговатым и спинным мозгом.
134.
Первый препарат был назван Бремером cerveauisolee (изолированный передний мозг), а второй
encephale isolee (изолированный головной мозг).
Межколликулярная перерезка оставляет ниже места
разреза почти всю ретикулярную формацию;
перерезка же ниже продолговатого мозга оставляет
в связи с высшими отделами мозга все стволовые
структуры.
135.
У животного с ceryeau isolee остаютсясвязанными с корой головного мозга такие
важнейшие
афферентные
системы,
как
обонятельная и зрительная. Однако такое животное
не реагирует на световые и обонятельные
раздражения; глаза у него закрыты, и оно
фактически не вступает в контакт с внешним миром.
136.
У животного же с encephale isolee реакцииполностью сохраняются; такое животное открывает
глаза на свет, реагирует на запахи, т.е. ведет себя
как бодрствующее, а не как спящее животное.
137.
Однако дальнейшие исследования показали, чтодля поддержания бодрствующего состояния КБП
важно не просто поступление к ней импульсации по
афферентным системам. Если сделать перерезку
ствола мозга так; чтобы не повредить основные
афферентные
системы
(например,
систему
медиальной петли), но перерезать восходящие связи
ретикулярной формации, то животное тем не менее
впадает в сонное состояние, конечный мозг
перестает активно функционировать.
138.
У человека в спокойном дремотном состояниитакие колебания имеют частоту 8-10 в секунду и
являются довольно регулярными (альфа-ритм).
Во время активности регулярные колебания
сменяются сразу же значительно меньшими по
амплитуде и более высокочастотными колебаниями
(бета-ритм).
139.
Когда правильные колебания большой амплитудысменяются низковольтными, частыми колебаниями,
то это, очевидно, говорит о том, что клеточные
элементы коры начинают функционировать менее
синхронно, поэтому такой тип активности
называется реакцией десинхронизации. Переход
от спокойного, неактивного состояния коры к
активному связан в электрическом отношении с
переходом от синхротзированной активности ее
клеток к десинхронизированной.
140.
В 1940 г. американскими исследователямиА.Форбсом и Б.Мориссоном, которые исследовали
вызванную электрическую активность коры при
различных афферентных влияниях. При радражении
какой-либо
афферентной
системы
в
соответствующей
проекционной
зоне
коры
обнаруживается электрический ответ, указывающий
на приход к этой области афферентной волны, этот
ответ получил название первичного ответа.
141.
Кроме этого локального ответа, афферентноераздражение вызывает длиннолатентный ответ,
возникающий в обширных областях коры мозга.
Этот ответ Форбс и Мориссон назвали вторичным
ответом.
142.
То обстоятельство, что вторичные ответывозникают со скрытым периодом, значительно
превышающим скрытый период первичного ответа,
ясно говорит о том, что они связаны с поступлением
афферентной волны в кору не по прямым, а по
каким-то окольным связям, через дополнительные
синаптические переключения.
143.
Позже,когда
было
применено
прямое
раздражение РФ, было показано, что оно может
вызвать ответ такого же типа. Это позволяет
заключить, что вторичный ответ является
электрическим проявлением поступления в КБП
афферентное волны через ретикулокортикальные
связи.
144.
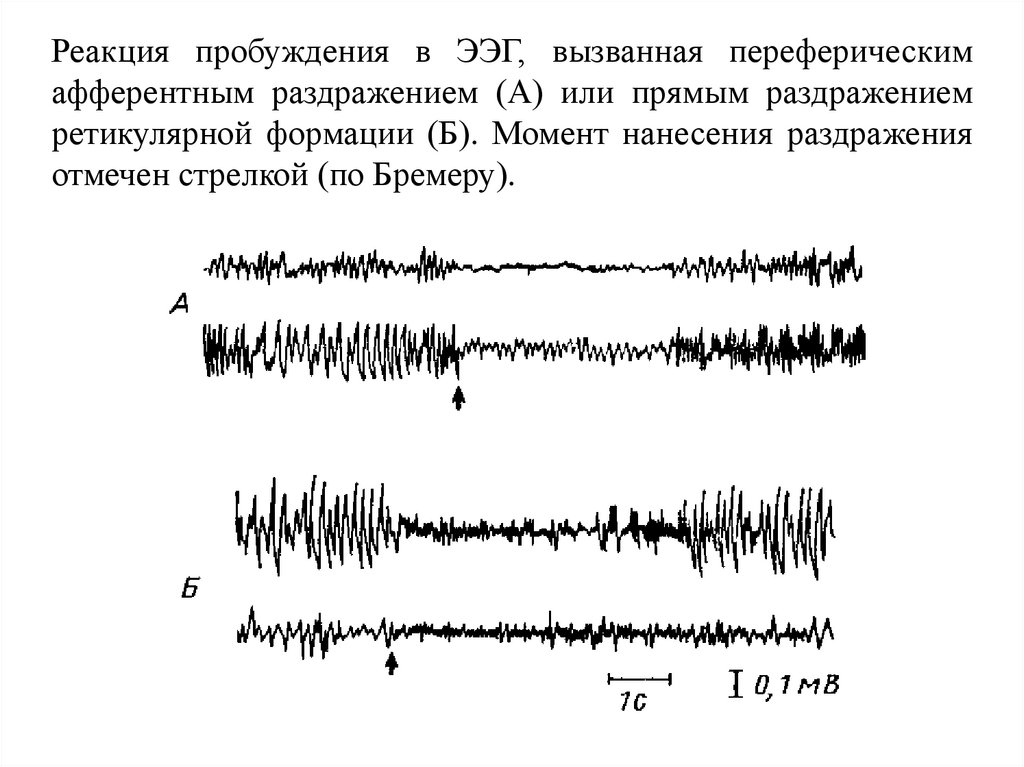
Реакция пробуждения в ЭЭГ, вызванная переферическимафферентным раздражением (А) или прямым раздражением
ретикулярной формации (Б). Момент нанесения раздражения
отмечен стрелкой (по Бремеру).
145.
Швейцарский физиолог В. Гесс (1929) впервыепоказал, что в стволе мозга можно найти точки, при
раздражении которых у животного наступает сон.
Гесс назвал эти точки центрами сна.
146.
Позже Дж.Моруцци (1941) также обнаружил,что, раздражая некоторые участки РФ заднего
мозга, можно у животных вызвать в коре вместо
десинхронизации - синхронизацию электрических
колебаний и соответственно перевести животное из
бодрствующего состояния в пассивное, сонное.
147. Тормозные влияния РФ на КБП
оказывает её каудальный отдел, стимуляцияего приводит к формированию на корковых
нейронах ТПСП и появляется
синхронизирующий эффект на ЭЭГ.
Тормозное влияние РФ вызывает
синхронизацию ЭЭГ в фазе «медленного»
сна и атонию мышц во время «быстрого»
сна.
У бодрствующего животного – угнетает
локомоторную функцию
148. Нисходящее влияния РФ на СМ
- облегчающие активность моторных системСМ, берут начало от всех отделов РФ.
Пути, идущие от моста – тормозят активность
мотонейронов мышц-сгибателей и
активируют мотонейроны разгибателей.
Пути, идущие от продолговатого мозга –
тормозят разгибатели и активируют
сгибатели.
Раздражение РФ – вызывает тремор и
повышение тонуса мышц, которое
сохраняется длительное время
(последействие).
149.
Аксоны нейронов гигантоклеточного ядра, т.е.основного ядра ретикулярной формации, очень
часто делятся Т-образно, и один из отростков идет
вниз, образуя ретикулоспинальньй путь, а второй
вверх, направляясь в верхние отделы головного
мозга. Создается такое впечатление, что и
восходящие, и нисходящие функции РФ могут быть
связаны с деятельностью одних и тех же нейронов.
По функциональным свойствам ретикулярные
структуры, создающие восходящие влияния, также
имеют
много
общего
со
структурами,
обеспечивающими нисходящие влияния.
150. Схема нисходящих влияний РФ на мотонейроны
РФРетикуло-спинальные пути
Альфа-мотонейрон
Активация
Торможение
Гамма-мотонейрон
151.
152.
Ретикулярное торможение устойчиво кдействию
стрихнина.
Стрихнин
является
специфическим
ядом,
устраняющим
постсинаптическое торможение мотонейронов,
вызванное
импульсацией
из
первичных
афферентов и связанное с выделением медиатора
глицина.
Это говорит о том, что ретикулярные
тормозящие эффекты создаются действием на
спинальные клетки другого медиатора.
153.
Восходящиевлияния
носят
тонический
характер, они легко изменяются гуморальными
факторами
и
высокочувствительны
к
фармакологическим веществам. Снотворное и
наркотическое действие барбитуратов основано,
именно на блокировании в первую очередь
восходящих влияний ретикулярной формации.
Особенно активными оказываются соединения
барбитуровой кислоты (аминозин), которые даже в
небольших концентрациях, не действующих на
спинальные нейроны или на нейроны КБП,
полностью прекращают активность ретикулярных
нейронов.
154.
Ретикулярныеструктуры,
регулирующие
вегетативные функции, отличаются высокой
химической чувствительностью; влияния РФ на
двигательные центры также легко изменяются под
влиянием таких химических факторов, как уровень
СО2 в крови и содержание в ней ФАВ
(адреналина).
155.
Промежуточный мозгСостоит из таламической области и
гипоталамуса.
Таламическая область – включает таламус,
коленчатые тела и эпиталамус (эпифиз)
Таламус (зрительный бугор)
1. Парный ядерный комплекс (до 60 ядер)
2. Ядра подразделяются на: релейные
(специфические), ассоциативные,
неспецифические
3. Выполняет 3 функции: переключающую,
интегративную, модулирующую.
156. Основные ядра таламуса
Специфические ядра• переключающие ассоциативные
моторные
Неспецифические ядра
• срединные ядра, надколенное ядро,
пограничное ядро, парафасцикулярное
ядро, ретикулярное ядро (проекция к
полосатому телу и V -VI слоям всех
областей коры больших полушарий)
157.
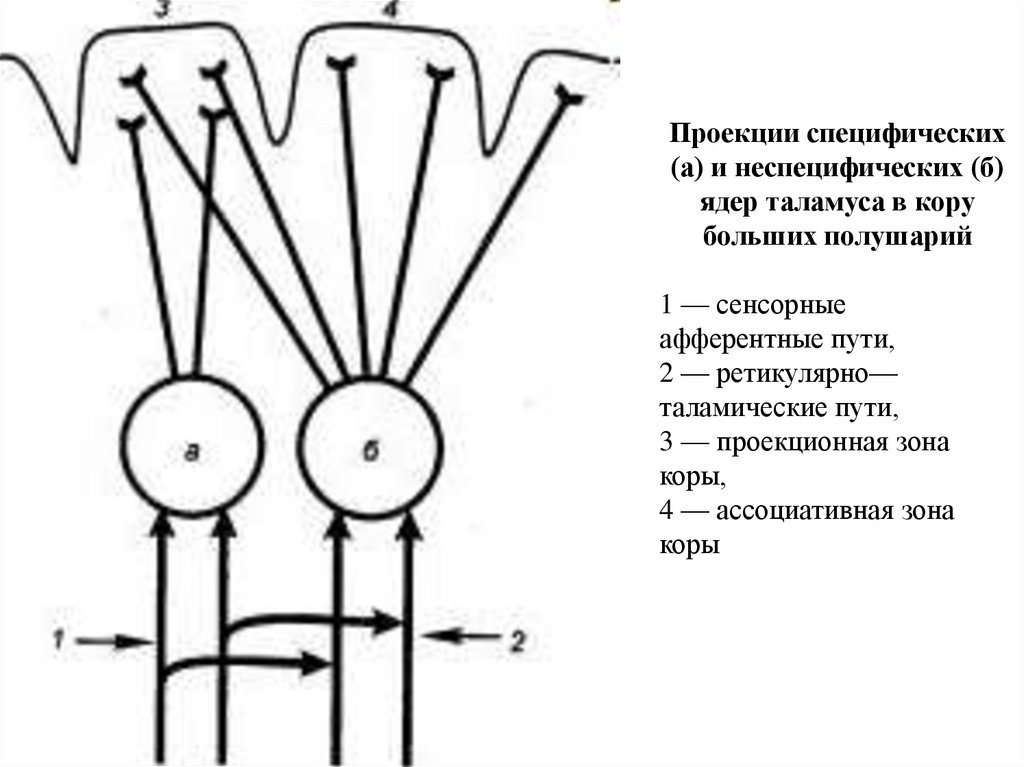
Проекции специфических(а) и неспецифических (б)
ядер таламуса в кору
больших полушарий
1 — сенсорные
афферентные пути,
2 — ретикулярно—
таламические пути,
3 — проекционная зона
коры,
4 — ассоциативная зона
коры
158.
Релейные ядра (специфические)
Сенсорные – переключают афферентную
импульсацию в сенсорные зоны КБП,
перекодируют и обрабатывают информацию
Несенсорные – переключают в КБП
несенсорную импульсацию поступающую из
разных отделов головного мозга
Сенсорные ядра
Выделяют 3 основных структуры:
Вентральные задние ядра (вентробазальный
комплекс)
Латеральное коленчатое тело
Медиальное коленчатое тело
159.
Специфические ядра таламусаПЕРЕКЛЮЧАЮЩИЕ
АССОЦИАТИВНЫЕ
МОТОРНЫЕ
Вентробазальный
комплекс (тактильная,
проприоцептивная,
температурная
болевая, вкусовая информация
в соматосенсорную тону
кору больших полушарий)
Внутреннее коленчатое
ядро тело (слуховая
информация в слуховые
зоны коры Наружное
коленчатое тело
(зрительная информация в
зрительные зоны коры)
Медиодорсальное
ядро (Проекция в лобные доли)
Подушка
(Проекция в теменную и височную
кору)
Заднелатеральное
(Проекция в теменную кору)
переднее ядро (Проекция в
лимбнчвскую кору)
Переднецентральное и
вентролатеральное
ядра
(Переключение)
от мозжечка и базальных
ганглиев в моторную
коры
больших
полушарий)
160. Ассоциативные системы таламуса
• ТАЛАМОПАРИЕТАЛЬНАЯ: ОТ ЗАДНЕЛАТЕРАЛЬНОГОЯДРА И ПОДУШКИ ТАЛАМУСА К ТЕМЕННОЙ И ВИСОЧНОЙ КОРЕ
• ТАЛАМОФРОНТАЛЬНАЯ: ОТ МЕДИОДОРСАЛЬНОГО И
ПЕРЕДНЕГО ЯДЕР ТАЛАМУСА К ФРОНТАЛЬНОЙ И ЛИМБИЧЕСКОЙ КОРЕ
• Механизмы работы:
• 1) Мультисенсорная конвергенция
• 2) Пластическая перестройка при гетеромодальных сенсорных воздействиях: избирательное привыкание, сенситизация
и эстраполяция
• 3) Кратковременное хранение следов интеграции в виде
реверберации импульсов
161. Основные функции ассоциативных систем таламуса
• ТАЛАМОПАРИЕТАЛЬНАЯ СИСТЕМА• 1) Центральный аппарат анализа и синтеза обстановочной
афферентации, запуска ориентационных движений глаз и
туловища
• 2) Один из центральных аппаратов «схемы тела» и сенсорного контроля текущей двигательной активности
• 3) Аппарат формирования полимодальных образов
• ТАЛАМОФРОНТАЛЬНАЯ СИСТЕМА
• Корковый модулятор лимбической системы, программирование целенаправленных поведенческих актов на основе
опыта и мотивации
162.
Вентральные задние ядраявляются главными реле переключения соматосенсорноафферентной
системы в составе волокон медиальной петли и примыкающей к ней
волокнам других афферентных путей. Передается в соматосенсорную
кору постцентральной извилины (поля 1-3 по Бродману), где
формируются соответствующие ощущения.
В них переключаются:
Тактильная
Проприоцептивная
Вкусовая
Висцеральная
Частично температурная и болевая
В ядрах имеется топографическая проекция периферии, при этом
функционально более тонко организованные части тела (язык, лицо)
имеют большую зону представительства.
Стимуляция ядер вызывает парастезии (ложные ощущения) в разных
частях тела и нарушение «схемы тела» (искаженное восприятие частей
тела в пространстве и относительно друг друга)
Особенностью сенсорных ядер является точность передачи
информации с минимальным искажением в КБП, по принципу «точка в
точку»
163.
Латеральное коленчатое тело (наружное)Реле для переключения зрительной
импульсации в затылочную кору (поле 17), где
используется для формирования зрительных
ощущений, далее в лобные и теменные доли
– ассоциативные зоны коры, где формируется
ответ на вопрос «Что такое», т.е.
окончательное формирование образа.
Медиальное коленчатое тело (внутреннее)
• Реле для переключения слуховой и,
возможно, вестибулярной импульсации в
верхнюю часть височной коры (поля 41 и 42)
164. Несенсорные релейные ядра -
Несенсорные релейные ядра Передние и вентральныеПередние ядра
- Афферентные – поступают из мамиллярных тел гипоталамуса
- Эффекторные – проектируются в лимбическую кору, а от туда в
гиппокамп и опять к гипоталамусу, формируя нейронный круг, лежащий
в основе возникновения эмоций (так называемые «лимбические» ядра
таламуса)
Вентральные ядра
Участвуют в регуляции движения, выполняя моторную функцию. В них
переключается информация от
- базальных ганглиев
- зубчатого ядра мозжечка
- красного ядра
которая затем проецируется в моторную и промоторную кору (поля 4
и 6), обеспечивая выполнение сложных двигательных программ,
образованных в мозжечке и базальных ганглиях.
Специфические ядра получают также нисходящие корковые волокна
из своих проекционных зон, что лежит в основе кольцевых
взаиморегулирующих отношений между корой и таламусом.
165. Ассоциативные ядра таламуса
Относится:- подушка
- медиодорсальное ядро
- латеральные ядра
их функция интегрирующая, в объединении деятельности таламических ядер и
различных зон ассоциативной КБП.
Подушка
- получает входы от коленчатых тел и неспецифических ядер
- эфференты – в височно – теменно – затылочные зоны КБП, которые учавствуют в
гностических (узнавание предметов, явлений), речевых и зрительных функций
(интеграция слова и зрительного образа), а также в восприятии «схемы тела».
Медиодорсальное ядро
- получает информацию от гипоталамуса, амигдалы, гипокампа, таламических ядер,
центрального серого в-ва ствола.
- эфференты – на ассоциативную лобную и лимбическую кору.
Участвует в формировании эмоциональной и поведенческой двигательной
активности и в запоминании.
Разрушение – устраняет страх, тревогу, напряженность, страдание от боли, но
возникает лобный синдром – снижение инициативы, безразличие и гипокинезия.
Латеральные ядра
Получают зрительную и слуховую импульсацию от коленчатых тел и от
вентрикулярного ядра (соматосенсорная импульсация), далее она передается в
ассоциативную теменную кору и используется в функции гнозиса, праксиса,
«схемы тела» (т.н. таламотеменная система)
166.
ФИЗИОЛОГИЯ МОЗЖЕЧКА167. Масса мозжечка взрослого человека 120 – 160 г. У новорожденного ≈ 20 г., в 3 месяца она удваивается, в 5 месяцев –
увеличиваетсяв 3 раза, в конце 9-го месяца – в 4 раза, а к 6
годам его масса достигает нижней границы
взрослого
человека
≈
120
г.
Выраженное
увеличение
мозжечка
отмечается между 5-м и 11-м месяцами жизни
– когда ребенок учится сидеть и ходить.
Площадь
поверхности
мозжечка
≈ 975 – 1500 см2.
168.
МозжечокСоставляет в среднем 10 % от массы головного мозга
Включает в себя более 50 % всех нейронов в ЦНС
В эволюционном плане выделяют три образования:
Древний мозжечок – клочок, узелок, часть червя (язычок)
Старый мозжечок – центральная долька, вершина, пирамида, язычок
червя и четырехугольная долька полушарий
Новый мозжечок – полушарии и части червя (ската, листка и бугра),
появляются у животных передвигающихся с помощью конечностей.
169.
170. СТРОЕНИЕ МОЗЖЕЧКА
171.
172.
Схема областей коры мозжечкаДольки обозначены римскими цифрами I – IX, а
соответствующие части полушарий п II – п IX
173. Анатомическое и функциональное подразделение мозжечка на: древний (архицеребеллюм) – темно-серая область, старый
(палеоцеребеллюм) – заштрихованоновый (неоцеребеллюм) – серая область.
174. Древний мозжечок (вестибулоцеребеллюм) – к нему относится клочково-узелковая доля, имеет наиболее выраженные связи с
вестибулярной системой (окончания вестибуломозжечковыхпутей, обозначены крупными черными кружочками
Старый мозжечок (спиноцеребеллюм) – состоит из участков червя, пирамиды, языка и
околоклочкового отдела – получает информацию от проприоцептивных систем мышц,
сухожилий, надкостницы, оболочек суставов по спиномозжечковым путям (светлые кружочки)
Новый мозжечок (понтоцеребеллюм) – включает в себя кору полушарий и участки червя, он
получает информацию от КБП (лобно-мосто-мозжечковый путь), от зрительных и слуховых
рецептирующих систем (мостомозжечковые пути – красные кружочки)
175. Медиальный мозжечок – функционально включает в себя вестибуло- и спиноцеребеллюм, дает проекции к ядру шатра и
проприорецептивным ядрам(шаровидному и пробковидному), от которых информация идет к стволовым и
двигательным центрам
Латеральный мозжечок – относят понтоцеребеллюм, дает проекцию в зубчатые ядра,
от которых к двигательным ядрам таламуса,, от них в двигательные области КБП.
Поступают волокна от ядра моста – мшистые, от нижней оливы – лазающие
176.
3 пары ножек мозжечкаСредние – соединяют его с варлиевым мостом (самые сильные)
Верхние – соединяют его с четвертохолмием
Нижние (веревчатые тела) – с продолговатым мозгом
Афферентные входы
По нижним ножкам – афферентные волокна заднего спинно-мозжечкового
пучка (пучки Голля, Бурдаха, Флексига) и вестибулярного аппарата,
определяют положение тела в пространстве и положение мышц и суставов
По средним ножкам – афферентные волокна из варолиева моста, а в него
из лобных и теменных областей КБП
По верхним ножкам – от переднего четвертохолмия и переднего спинномозжечкового пучка
Эфферентные выходы
По верхним ножкам – от зубчатого, пробковидного и шарообразного ядер,
от них в красное ядро, зрительные бугры и подбугорковую область. От
красного ядра – руброспинальный пучок, от зрительных бугров – в кору
больших полушарий
По нижним ножкам – от ядра покрышки в продолговатый мозг
177. Фазы последствия удаления мозжечка
• 1. Фаза раздражения — связана с повреждением мозжечка,• У человека — длиться несколько суток и характеризуется
параличом двигательных систем.
• 2. Фаза выпадения функций — характеризуется резким
нарушением координированности, пластичности движений и их
точности. Сопровождается потерей способности к выполнению
сложных двигательных актов (катание на коньках, выполнение
спортивных движений и т.д.) Длиться у человека до нескольких
лет.
• 3. Стадия компенсации — наступает через 3 — 5 лет и
сопровождается постепенным исчезновением симптомов
мозжечковой недостаточности второй фазы. Декортикация
устраняет эту стадию, так разрушение лобных долей ведет к
возникновению мозжечковых изменений (лобно — мозжечковый
путь).
178. Симптомы фазы выпадения функций:
1. атония — снижение тонуса мышц
2. aстения — повышенная утомляемость при выполнении движений (причина
нарушения трофики мышц)
З. астезия — нарушение способности к слитному тетаническому сокращению (качанию
головь, туловища, конечностей, затруднение стояния или сидения)
4. атаксия — нарушение точности и координированности движений. Тремор действия
может переходить в размашистые колебания конечностей, а поставленная цель не
достигается. Наблюдается неуверенная походка, с широко расставленными ногами и
избыточными движениями, при которых больного как бы «бросает» из «стороны в
сторону» («пьяная походка»). Больной не может выполнить пальце-носовую,
указательную или пяточно-коленную пробы.
5. асинергия — неспособность сесть из положения лежа в положение сидя без помощи
рук.
6. дистония — непроизвольное повышение или понижение тонуса мышц.
7. дисметрия — расстройство равномерности движения выражающееся в излишнем,
либо недостаточном движении. Больной пытается взять предмет со стола и проносит
руку (гиперметрия), либо не доносит до предмета (гипометрия)
8. адиадохокинез — расстройства при работе антагонистических групп мышц
(невозможным становится быстрое сгибание и разгибание пальцев, вращение ладоней
вниз-вверх), неспособность выполнения быстрой последовательности движений.
9. деэквилибраиия —- нарушение равновесия (выявляется пробой Ронберта)
10. дизартрия — расстройства речевой моторики (скандированная речь)
11. абазия - потеря способности ходить, в основном при заболеваниях нервной системы,
связанная с расстройствами равновесия тела или с двигательными нарушениями нижних
конечностей. Больной может совершать соответствующие действия, в необходимом
объёме и с достаточной силой, лёжа.
179. СОМАТИЧЕСКАЯ ПРОЕКЦИЯ В КОРЕ ЧЕРВЯ И ПОЛУШАРИЯХ МОЗЖЕЧКА
180.
181. Кора мозжечка содержит 3 слоя Молекулярный (самый верхний слой коры) – состоит из параллельных волокон, разветвлений дендритов
и аксонов 2-3 слоев. В нижней части этого слоярасположены:
1) Корзинчатые клетки;
2) Звездчатые клетки;
3) клетки Лугаро;
которые обеспечивают взаимодействие с клетками Пуркинье.
Средний слой – (ганглиозный) или слой грушевидных клеток Пуркинье,
выстроенных в один ряд и имеющих самую мощную в ЦНС дендритную систему, на
которой может оканчиваться до 60 тысяч синапсов и которые выполняют задачу
сбора, обработки и передачи информации. Аксоны клеток Пуркинье образуют
единственный выход из коры мозжечка на его ядра и другие структуры ЦНС.
Клетки Пуркинье –это тормозные нейроны, и их активация ведет к торможению
клеток ядер мозжечка. Кроме дендритов клеток Пуркинье в верхних участках слоя
проходят параллельные волокна, представленные вставочными нейронами.
Гранулярный или зернистый слой – состоит из клеток-зёрен и клеток Гольджи.
Клетки-зёрна – являются возбуждающими в отличии от остальных 5 видов клеток,
которые выполняют роль тормозных структур. Число этих клеток достигает 10
млрд, а их аксоны поднимаются вверх и Т-образно делятся на поверхности коры,
образуя дорожки контактов с клетками Пуркинье.
Волокна клеток Гольджи – направляются в молекулярный слой.
182.
Нейроннаяорганизация коры
мозжечка
Вверху слева: схема
поперечного среза
через листок
Внизу, главные
синаптические
контакты в листке
183.
184.
185. Чешский анатом Ян Пуркинье – 1837 г. описал грушевидные нейроны (neuronum piriforme) коры мозжечка, сферическая форма (60×30
мкм), располагаются в один слой,каждый аксон иннервируется около 1000
нейронов ядер мозжечка, всего их около
26 млн.
186.
Клетки Пуркинье коры мозжечкаОбразуют средний слой коры, являются главной функциональной
единицей, обладают резко выраженной интегративной функцией
На одной клетке может быть до 100 тыс. синапсов
Общая площадь дендритов и шипиков клеток Пуркинье в коре
мозжечка ≈ 4 кв.м.
Являются единственными эфферентными нейронами коры
непосредственно
связывают
ее
с
внутримозжечковыми
вестибулярными ядрами
Являются тормозными нейронами (медиатор ГАМК)
и
и
187. Клетки Пуркинье достигают окончательного развития к 8 годам, поэтому дети не умеют рассчитывать движения, выглядят неуклюжими и
неловкими, при письме – выходят «каракули».Тренировки ускоряют созревания клеток
Пуркинье: самый развитый мозжечок имеют
гимнасты,
фигуристы
и
балерины.
Клетки Пуркинье – очень чувствительны к
алкоголю, что приводит к сбою в работе
мозжечка, определяя траекторию движения и
согласованность работы ног и рук.
188.
189. КЛЕТОЧНЫЕ ЭЛЕМЕНТЫ КОРЫ МОЗЖЕЧКА
190.
191. Мелкие рецептивные поля шести клеток Пуркинье, активность которых регистрировалась двумя микроэлектродами, введенными в дольки
Мелкиерецептивные
поля шести
клеток
Пуркинье,
активность
которых
регистрировалас
ь двумя
микроэлектрода
ми, введенными
в дольки V и ПVI
192. СВЯЗИ КОРЫ МОЗЖЕЧКА
• АФФЕРЕНТНЫЕ СВЯЗИ- МОХОВИДНЫЕ ВОЛОКНА: от
• 1) Вестибулярных ядер - вестибулоцеребеллярные
тракты
• 2) Спинного мозга - спиноцеребеллярные тракты
• 3) Ретикулярной формации - ретикулоцеребеллярные
тракты
• 4) Коры больших полушарий - кортикоцеребеллярные
тракты
- ЛИАНОВИДНЫЕ ВОЛОКНА: от
• Нижней оливы - клетки Пуркинье (1 волокно-1 клетка)
• ЭФФЕРЕНТНЫЕ СВЯЗИ - К ПОДКОРКОВЫМ ЯДРАМ
193.
Лиановидные волокна (лазающие)Почти все являются аксонами нейронов нижней оливы продолговатого мозга, к которым
поступает информация от мышечных и кожных рецепторов и от двигательных зон КБП
Каждое волокно устанавливает синапс с дендритами, как правило, одной клетки
Пуркинье (медиатор аспартат)
Число синапсов может достигать трехсот (300), что объясняет сильное возбуждающее
действие этого афферентного входа (большой ВПСП, на котором формируется 3-5 ПД)
Через корзинчатые и звездчатые клетки оказывают слабое тормозное действие на
клетки Пуркинье.
Моховидные волокна (мшистые)
По ним поступает афферентная информация от:
-КБП (преимущественно из ассоциативных зон)
-Проприорецепторов
-Вестибулярных рецепторов
-Ретикулярной формации
Минуя нейроны нижней оливы волокна образуют возбуждающие синапсы на клеткахзернах, а через них возбуждающее влияние (медиатор глютамат) на корзинчатые и
звездчатые клетки, которое вызывает торможение клеток Пуркинье
Активность клеток-зерен регулируется через тормозные клетки Гольджи, по типу
возвратного торможения (медиатор ГАМК)
Т.о. моховидные волокна – снижают тормозное влияние клеток Пуркинье на ядра
мозжечка, а лиановидные - усиливают это тормозное влияние
194.
Три пары ядер мозжечкаРасполагаются в белом веществе:
•Шатра (фастигиальное)
•Промежуточные (пробковидное и шаровидное)
•Зубчатые
Клетки Пуркинье коры мозжечка проецируются:
-из древнего мозжечка – на ядра шатра и далее на ядро Дейтерса
(повышение тонуса разгибателей и снижение сгибателей) и РФ моста мозга
(повышение тонуса разгибателей, снижение – сгибателей)
- из старого мозжечка – на промежуточные ядра и далее к красному ядру и
РФ продолговатого мозга (вызывая повышение тонуса сгибателей и тормозя
разгибателей)
-из нового мозжечка – на зубчатые ядра и далее в таламус, а оттуда в
двигательную кору
Нейроны этих ядер имеют высокую фоновую активность (50-100 имп/сек),
находятся под тормозящим влиянием клеток Пуркинье и возбуждающим
влиянием афферентных входов в мозжечок, которые по коллатералям
поступают в мозжечок
Эти три пары ядер образуют главные эфферентные выходы мозжечка на
стволовые и корковые моторные центры
195. Связи ядер мозжечка
• АФФЕРЕНТНЫЕ СВЯЗИ ВСЕХ ЯДЕР - ОТ КОРЫ МОЗЖЕЧКА- ЗУБЧАТЫЕ ЯДРА: от коры полушарий
- ВСТАВОЧНЫЕ ЯДРА (ПРОБКОВОЕ И ШАРОВИДНОЕ): от средней
части коры
- ЯДРО ШАТРА: от коры червя
• ЭФФЕРЕНТНЫЕ СВЯЗИ ЯДЕР:
- ЗУБЧАТЫЕ ЯДРА: к моторным ядрам таламуса и затем к
двигательной зоне коры больших полушарий
- ВСТАВОЧНЫЕ ЯДРА: к красным ядрам
- ЯДРО ШАТРА: к ретикулярной формации и вестибуляр-ному
ядру Дейтерса
196.
197. ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
• РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА, РАВНОВЕСИЯ,ПОДДЕРЖИВАЮЩИХ ДВИЖЕНИЙ.
• КОРРЕКЦИЯ МЕДЛЕННЫХ ЦЕЛЕНАПРАВ-ЛЕННЫХ ДВИЖЕНИЙ
И ИХ КООРДИНАЦИЯ С РЕФЛЕКСАМИ ПОДДЕРЖАНИЯ ПОЗЫ
• ПРАВИЛЬНОЕ ВЫПОЛНЕНИЕ БЫСТРЫХ
ЦЕЛЕНАПРАВЛЕННЫХ ДВИЖЕНИЙ ПО КОМАНДАМ КБП В
СТРУКТУРЕ ОБЩЕЙ ПРОГРАММЫ ДВИЖЕНИЙ
• Т.О. МОЗЖЕЧОК НЕ ТОЛЬКО МОДУЛИРУЕТ УЖЕ НАЧАТЫЕ
ДВИГАТЕЛЬНЫЕ АКТЫ, НО И ОБЕСПЕЧИВАЕТ СОЗДАНИЕ И
ХРАНЕНИЕ ИНДИВИДУАЛЬНЫХ ПРОГРАММ БЫСТРЫХ,
ВЫСОКОКООРДИНИРОВАННЫХ ДВИЖЕНИЙ.
198.
Двигательная функция мозжечка1. Регуляция равновесия и формирования позы
- осуществляются преимущественно древним мозжечком и частично старым мозжечком, входящим
в медиальную червячную зону.
Получая и обрабатывая информацию от вестибулярных рецепторов, проприорецепторов аппарата
движения, кожных, зрительных и слуховых рецепторов – он способен оценить состояние мышц и
положения тела в пространстве, и через ядра Шатра и вестибулярные ядра изменить позу тела и
сохранить равновесие, нарушение которого является наиболее характерным симптомом
поражения древнего мозжечка.
2. Координация выполняемых движений
- осуществляется старым и новым мозжечком, входящим в околочервячную зону (промежуточная
зона), за счёт анализа информации, поступающей от проприорецепторов и от моторной зоны
КБП, в которых заложена “программа движения”.
Это достигается с помощью анализа информации о “программе” и выполнении движения через
промежуточные ядра, имеющие выходы на красное ядро и моторную кору. Преимущественно
регулируется амплитуда, направление и темп движения – симптомом нарушения функции
промежуточной зоны является нарушение координации движения (атаксия).
3. Коррекция быстрых движений
- которая не может быть осуществлена посредством импульсации от проприорецепторов мышц
Осуществляется следующим образом:
А. Мозжечок получает копию эфферентной программы движения от моторной зоны КБП до
начала движения
Б. Кора мозжечка, которая уже имеет программу этого движения в ходе предыдущего
обучения в фило – и онтогенезе, сравнивает её и исправляет возможные ошибки путём
изменения эфферентной программы движения и посылает эту информацию в КБП для реализации
её выполнения
К таким движениям можно отнести многие быстрые спортивные движения, игра на музыкальных
инструментах и др.
199.
4. Программирование целенаправленных движений-осуществляется корой нового мозжечка, которая получает информацию из
ассоциативных зон КБП через ядра моста. Эта информация характеризует
замысел движения, который инициируется лимбической системой и
ассоциативной КБП.
-в коре нового мозжечка, в базальных ядрах и в моторной КБП эта информация
перерабатывается в программу движения, которая через зубчатое ядро
мозжечка и вестибулярные ядра таламуса поступает в премоторную КБП.
-в промежуточной коре осуществляется дальнейшая обработка информации, и
через пирамидную и экстрапирамидную системы реализуется в сложный
целенаправленный двигательный акт.
Эфференты мозжечка
не имеют прямого выхода на мотонейроны СМ, а действуют на них через
корково-стволовые двигательные центры, что обуславливает высокую степень
пластичности КБП в компенсации нарушенных функций мозжечка. Так,
известны случаи врождённого отсутствия мозжечка, при которых у человека не
определяются симптомы нарушения движения.
200.
Упрощенная функциональная схема медиальных структур мозжечка201.
Главные связи медиальной (слева) и латеральной (справа) частей мозжечка Проекциипервой идут преимущественно в двигательные центры ствола мозга, а второй через
вентролатеральный таламус (Т) в двигательную КБП.
Ш – ядро шатра, ВЯ – вставочное ядро, ЗЯ – зубчатое ядро, КЯ – красное ядро, РФ –
ретикулярная формация, ЯД- ядро Дейтерса
202.
Напряжение мышц разгибателей у собаки, лишенноймозжечка. СНИМКИ 3 и 4 показывают усиление тонуса при
раздражении стопы (по Радемакеру).
203. Характер движений после удаления мозжечка
У животных с удаленным или разрушенным мозжечкомнаблюдается: атония, астазия, астения, атаксия, абазия
204. Луиджи Лучиани (1840—1919) – в 1891г. показал, что основным комплексом двигательных нарушений мозжечкового происхождения
является атаксия,включающая такие симптомы, как атония, астазия и
астения (триада Лучиани).
205. ЦИТАТА
• «Как скульптор избирательно удаляетрезцом все лишнее из первоначально
бес-форменного камня, так и
мозжечок, подавляя торможением
лишние возбуждения, добивается
четкой формы двигательной
реакции»
Экклз, 1969
206. ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА
• ТРИАДА ЛЮЧИАНИ: атония,астазия,
астения
• ТРИАДА ШАРКО:
нистагм,
тремор,
скандированная речь
• АТАКСИЯ (пьяная походка)
• ДИСМЕТРИЯ (избыточность)
• ДИЗАРТРИЯ
• ДИЗЭКВИЛИБРАЦИЯ
• АДИАДОХОКИНЕЗ
207. БАЗАЛЬНЫЕ ГАНГЛИИ
208.
209. Базальные ганглии
Совокупность парных образованийрасположенных в конечном мозге, в
основании больших полушарий
• Полосатое тело – хвостатое ядро и скорлупа
• Бледный шар (pallidum)
• Субталамическое ядро
Возбуждающая афферентная импульсация
от 3 основных источников:
• Всех областей коры прямо и через таламус
• Неспецифических интраламинарных ядер
таламуса
• Черного вещества
210.
211. Эфферентные связи базальных ганглиев
Имеют 3 главных выхода:1. от полосатого тела тормозящие пути идут к бледному шару
напрямую и с участием субталамического ядра – от
бледного шара начинается самый главный путь, идущий
преимущественно в таламус (в его двигательные
вентральные ядра) и от них возбуждающий путь в
двигательную КБП
2. часть эфферентных волокон из бледного шара и
полосатого тела идет к центрам ствола мозга
(ретикулярная формация, красное ядро и далее в спинной
мозг), а также через нижнюю оливу в мозжечок
3. от полосатого тела тормозящие пути идут к черной
субстанции и после переключения – к ядрам таламуса
В целом – базальные ганглии, эта структура является
промежуточным звеном, своеобразной станцией
переключения со сложной переработкой
информации, связывающей преимущественно,
ассоциативную, и частично, сенсорную кору с
двигательной КБП
212.
213.
Афферентные иэфферентные связи
базальных ядер
1 — хвостатое ядро,
2 — скорлупа,
3 — черное вещество,
4 — таламус,
5, 5* — внутренний и внешний
отделы бледного шара,
6 — субталамическое ядро,
7 — сенсомоторная кора,
8 — двигательная кора,
9 — пути к стволовой части
мозга;
стрелками обозначены
афферентные пути,
эфферентные пути,
межъядерные взаимодействия
214.
Главные связи базальных ганглиев.Возбуждающие влияния показаны красным, тормозные–черным. БШ – бледный
шар (БШн–наружный сегмент, БШв–внутренний сегмент); ЧВ черное вещество (Кч
компактная часть, Сч сетчатая часть); СТЯ субталамическое ядро; КСМ крыша
среднего мозга; ПВЯ переднее вентральное ядро таламуса; ВЛЯ–
вентролатеральное ядро таламуса; ВМЯ вентромедиальное ядро таламуса;
ЦМПФЯ центральное медиальное и парафасцикулярное ядра таламуса; ДЯШ
дорсальное ядро шва.
215. Роль БГ в регуляции в ДА
Отношение между полосатым телом и черным веществом• Имеются двухсторонние связи, за счет чего возникает реверберация,
необходимая для нормального межъядерного баланса в стриопаллидарной
системе, направленной на саморегуляцию системы и предупреждение
гиперактивности ее отдельных звеньев
• Нейроны полосатого тела связывают тормозящее влияние (медиатор
ГАМК) на черную субстанцию
• Нейроны
черного
вещества
(медиатор
дофамин)
оказывают
модулирующее влияние (возбуждающее и тормозное) на полосатое тело
• Дофаминэргическим путям в базальных ганглиях придается большее
значение в связи с двигательными расстройствами типа Паркинсонизм,
когда падает резко концентрация дофамина в обоих ядрах полосатого
тела – хвостатом ядре и скорлупе, иннервируемых нейронами черного
вещества
• Черное вещество оказывает тормозное действие на нейроны таламуса
(медиатор ГАМК) и получает возбуждающие афференты – от
субталамического ядра
216. Влияние полосатого тела
Роль БГ в регуляции в ДАВлияние полосатого тела
•В эксперименте показано тормозное влияние (медиатор АЦХ)
хвостатого ядра на двигательную кору, при этом вытормаживая
ненужные в данных условиях рефлексы
•Стриатум – участвует в поворотах головы и туловища вдоль
продольной оси при ходьбе по кругу, которые входят в структуру
ориентировочного поведения
•Поражение стриатума (его хвостатого ядра) – приводит к
появлению несинхронных избыточных движений (гиперкинезы,
хорея и атетоз), организм как бы не может справиться со своей
мускулатурой, число бесцельных движений в пространстве
увеличивается в 5-7 раз
217. Функции бледного шара
Роль БГ в регуляции в ДАФункции бледного шара
• Получает из полосатого тела преимущественно тормозное и
частично возбуждающее влияние
• Бледный шар оказывает модулирующее влияние на
двигательную кору, мозжечок, ретикулярную формацию, красное
ядро
• При его стимуляции – преобладают элементарные двигательные
реакции в виде сокращения мышц конечностей, шеи и лица
• При его разрушении – снижается двигательная активность –
возникает адинамия
218.
Активность нейрона внутреннего сегмента бледного шара обезьяныА. Спонтанная, до начала движения.
Б, В. Во время условного рефлекса с выполнением специфических движений.
Видно, что активность коррелирует с движениями рук (Б), но не ног (В)
219.
Активность нейронакомпактной части
черного вещества
обезьяны коррелирует с
саккадами глаз.
Вертикальной линией
отмечен момент появления
светового пятна. Обезьяна
обучена саккадически
переводить взгляд на эту
новую точку фиксации.
Примерно через 100 мс
после появления светового
пятна и за 200 мс до
начала саккады нейронная
активность почти
полностью подавляется
(перерыв точечной записи
и гистограммы; плато на
кумулятивной кривой). Это
ведет к растормаживанию
(активации) нейронов–
мишеней в таламусе и
крыше среднего мозга
220. Роль базальных ганглиев в регуляции двигательной активности
При циркуляции возбуждения в стриопаллидарной системе(базальные ганглии → таламус → различные зоны КБП →
базальные ганглии) она:
• Создает программы целенаправленных движений с
учетом доминирующей мотивации
• В моторном обеспечении суточных биоритмов
• Регулирует движения глаз
• Вращательные движения
• Различные параметры движения (амплитуду, силу,
направление)
Кроме этого сриопаллидарная система включается:
• В регуляцию цикла «сон – бодрствование»
• В формирование условных рефлексов
• В сложные формы восприятия (например – осмысление
текста)
• В организацию эмоционально-мотивационной сферы
• Адекватного приспособительного поведения
221. ФУНКЦИИ БАЗАЛЬНЫХ ГАНГЛИЕВ
• 1. Центры координации сочетанныхдвигательных актов
• 2. Центры сложных безусловных
рефлексов и инстинктов
• 3. Центры контроля координации
тонуса мышц и произвольных
движений
• 4. Центры торможения агрессивных
реакций
• 5. Участие в механизмах сна
222. Формирование программы действия с участие БГ
ДВИГАТЕЛЬНАЯКОРА
ТАЛАМУС
АССОЦИАТИВНАЯ
КОРА
БЛЕДНЫЙ ШАР
СТВОЛ МОЗГА
СУБТАЛАМИЧЕСКОЕ
ЯДРО
ЧЕРНАЯ СУБСТАНЦИЯ
ПОЛОСАТОЕ
ТЕЛО
223. ЭКСТРАПИРАМИДНАЯ СИСТЕМА
224.
225.
Функциональные петли, проходящие через базальные ганглииПМК–премоторная кора; ДД К–дополнительная двигательная область коры; ДК двигательная кора; ССК–
соматосенсорная кора;Ск скорлупа; вБШ внутренний сегмент бледного шара (вБШ вл вентролатеральная (область,
вБШ кдм–каудальная дорсомедиальная область, вБШ дм/рл дорсомедиальная/ростролатеральная области); сЧВ
сетчатая часть черного вещества (сЧВ кл–каудолатеральная область, сЧВ вл вентролатеральная область, сЧВр
роростральная область); ВЛЯ вентролатеральное ядро таламуса (ВЛЯ м–медиальная область, ВЛЯ р–
представительство рта); ПФАК–префронтальная ассоциативная кора; П8–поле 8 (фронтальное глазное поле коры);
П7–поле 7 (теменная ассоциативная кора); ХЯ хвостатое ядро; МДЯ –медиодорсальное ядро таламуса (МДЯ пл–
параламеллярная область, МДЯ мк/кк–мелкоклеточная/крупноклеточная области, МДЯ пм–постеромедиальная
область); ПВЯ переднее вентральное ядро таламуса (ПВЯ лкк–латеральная крупноклеточная область, ПВЯ мк
мелкоклеточная область, ПВЯ мкк–медиальная крупноклеточная область); ФАК фронтальная (лобная) ассоциативная
кора
226. ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
• Поражения хвостатого ядра:• гиперкинезы- атетозы и хорея (пляска
святого Витта)
• Поражения паллидум:
• обеднение двигательной
• активности при повышенном
пластическом тонусе и треморе
(болезнь Паркинсона)
227. ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
Базальные ядра совместно с корой больших полушарий контролируют дваважных показателя — амплитуду движений и скорость изменений движения.
Поэтому нарушения движений, связанные с заболеваниями базальных ядер,
подразделяют на гиперкинетические и гипокинетические
Гиперкинезы:
Атетозы - патологические непроизвольные движения, выражающиеся
медленной тонической судорогой мышц конечностей, лица, туловища
Хорея - синдром, характеризующийся беспорядочными, отрывистыми,
нерегулярными движениями, сходными с нормальными мимическими
движениями и жестами, но различные с ними по амплитуде и интенсивности,
то есть более вычурные и гротескные, часто напоминающие танец
(хореический гиперкинез, ранее «виттова пляска» или «пляска святого Вита»)
Гемибаллизм – при поражении подбугорного ядра и его связи с бледным шаром
(вариант экстрапирамидного гиперкинеза, характеризующийся быстрыми
размашистыми движениями в большом объеме на одной стороне тела,
напоминающие толкание ядра или бросание мяча, возможны элементы
ротаторного движения туловища)
228. ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
Болезнь Хантингтона- при потери ГАМК–ергических и
холинергических нейронов полосатого тела (характерна хорея —
беспорядочные, неконтролируемые движения, в начале может
проявляется в виде беспокойства, небольших непроизвольных
или незавершённых движений, нарушения координации и
замедления скачкообразных движений глаз)
Болезнь Паркинсона - при поражении черной субстанции
(характерны двигательные нарушения: тремор, гипокинезия,
мышечная ригидность, а также вегетативные и психические
расстройства — результат снижения тормозящего влияния
бледного шара на полосатое тело.
229. ЛИМБИЧЕСКАЯ СИСТЕМА
230. Лимбическая система
Функциональное объединение различных структур конечного,промежуточного и среднего мозга, обеспечивающее
эмоционально-мотивационные компоненты поведения и
интеграцию висцеральных функций организма
Структурно-функционально включают:
1. Корковые структуры:
– Обонятельную извилину
– Гиппокамп (амнонов рог)
– Парагиппокамповую извилину
– Зубчатую извилину
– Поясную извилину
2. Подкорковые структуры:
– Амигдала
– Ядра перегородки
– Ограду
– Гипоталамус
– Передние ядра таламуса
231. Афферентные входы
• От различных областей головного мозга• Через гипоталамус от ретикулярной формации
ствола (один из главных источников ее возбуждения)
• От обонятельных рецепторов по волокнам
обонятельного нерва
Эфферентные выходы
• Через таламус (особенно его мамиллярные тела), на
вегетативные и соматические центры спинного мозга
и ствола
• В КБП – особенно ассоциативные зоны, а через нее в
регуляцию высших психических функций
232. Лимбическая система
233. ОСНОВНЫЕ ФУНКЦИИ ЛИМБИКИ
1. Организация вегетативно-соматическихкомпонентов эмоций
2. Организация кратковременной и
долговременной памяти
3. Участие в формировании ориентировочноисследовательской деятельности
4. Организация простейшей мотивационноинформационной коммуникации (речи)
5. Участие в механизмах сна
6. Центр обонятельной сенсорной системы
234. Кольцевые нейрональные связи
Дают возможность длительной циркуляции(реверберации) возбуждения, что создает условия для
сохранения единого функционального состояния
структур замкнутого круга и навязывание этого
состояния другим структурам мозга
Круг Пейпеца (1937) – от гиппокампа → мамилярные
тела гипоталамуса → передние ядра таламуса → в
поясную извилину → парагиппокамповая извилина →
гиппокамп – этот круг лежит в основе формирования
эмоций, обучения, и памяти
Другой круг – от амигдалы → мамилярные тела
гипоталамуса → лимбической области среднего мозга
→ обратно к амигдале – лежит в основе формирования
агрессивно-оборонительных, пищевых и сексуальных
реакций
235.
2 ЛИМБИЧЕСКИХ КРУГАгипоталамус
• БОЛЬШОЙ КРУГ
ПЕЙПСА:
гиппокамп - сводмамиллярные тела мамиллярно-таламический
пучок Вик-д’Азира - таламус поясная извилина гиппокамп
• МАЛЫЙ КРУГ НАУТА:
миндалина - конечная
полоска - гипоталамус перегородка - миндалина
236. Функциональная структура лимбики по МакЛину (1970)
• 1. Нижний отдел - миндалина игиппокамп - центры эмоций и
поведения для выжива-ния и
самосохранения
• 2. Верхний отдел - поясная извилина и
височная кора - центры
общительности и сексуальности
• 3. Средний отдел - гипоталамус и
поясная извилина - центры
биосоциальных инстинктов
237. Функции лимбической системы
После обработки информации отвнешней среды и внутренних
органах запускает вегетативные,
соматические и поведенческие
реакции, обеспечивая адекватное
приспособление организма к
внешней среде и сохранение
гомеостаза
238. Физиология гиппокампа
239.
240. Физиология гиппокампа
Гиппокамп (hippocampus) является основной структурой лимбическойсистемы. Морфологически гиппокамп представлен стереотипно
повторяющимися модулями, связанными между собой и с другими
структурами.
Модульное
строение
обусловливает
способность
гиппокампа
генерировать высокоамплитудную ритмическую активность. Связь
модулей создает условие циркулирования активности в гиппокампе при
обучении. Многочисленные связи гиппокампа со структурами как
лимбической системы, так и других отделов мозга определяют его
многофункциональность.
Выраженными и специфическими являются электрические процессы в
гиппокампе. Активность здесь чаще всего характеризуется быстрыми
бета-ритмами (14—30 в секунду) и медленными тета-ритмами (4—7 в
секунду).
Если в новой коре ослабить десинхронизацию на новое раздражение,
то в гиппокампе затрудняется возникновение тета-ритма. Раздражение
ретикулярной формации ствола мозга усиливает выраженность тетаритма в гиппокампе и высокочастотных ритмов в новой коре.
241. Физиология гиппокампа
Значение тета-ритма заключается в том, что он отражает реакцию гиппокампа, а темсамым — его участие в ориентировочном рефлексе, реакциях настороженности,
повышения внимания, в динамике обучения, при высоком уровне эмоционального
напряжения — страхе, агрессии, голоде, жажде.
Нейроны гиппокампа отличаются выраженной фоновой активностью. В ответ на
сенсорное раздражение реагирует до 60% нейронов гиппокампа. Особенность
строения
гиппокампа,
взаимосвязанные
модули
обусловливают
цикл
генерирования возбуждения в нем, что выражается в длительной реакции (до 12 с)
нейронов на однократный короткий стимул.
Повреждение гиппокампа у человека нарушает память на события, близкие к моменту
повреждения (ретроантероградная амнезия).
Нарушаются:
1. запоминание,
2. обработка новой информации,
3. различие пространственных сигналов.
Повреждение гиппокампа ведет к снижению эмоциональности, инициативности,
замедлению скорости основных нервных процессов, повышаются пороги вызова
эмоциональных реакций.
242. Физиология миндалевидного тела
243.
244. Физиология миндалевидного тела
Миндалевидное тело, миндалина — подкорковая структура лимбическойрасположенная в глубине височной доли мозга. Ядра миндалины полисенсорны.
системы,
Функции миндалины связаны:
1. с обеспечением оборонительного поведения,
2. вегетативными,
3. двигательными,
4. эмоциональными реакциями,
5. мотивацией условнорефлекторного поведения.
Миндалины реагируют многими своими ядрами:
1. на зрительные,
2. слуховые,
3. интероцептивные,
4. обонятельные,
5. кожные раздражения.
Реакция ядра на внешние раздражения длится, как правило, до 85 мс, т. е. значительно меньше,
чем реакция на подобные же раздражения новой коры.
Раздражение ядер миндалевидного тела создает выраженный парасимпатический эффект на
деятельность:
1. сердечно-сосудистой,
2. дыхательной систем,
3. приводит к понижению (редко к повышению) кровяного давления,
4. урежению сердечного ритма,
5. нарушению проведения возбуждения по проводящей системе сердца,
6. возникновению аритмий и экстрасистолий.
245. Физиология миндалевидного тела
При искусственной активации миндалины появляются реакции:1.принюхивания,
2.облизывания,
3.жевания, глотания,
4.саливации,
5.изменения перистальтики тонкой кишки
Повреждение миндалины у животных снижает:
1.адекватную подготовку автономной нервной системы к организации и реализации
поведенческих реакций,
2.приводит к гиперсексуальности,
3.исчезновению страха,
4.успокоению,
5.неспособности к ярости и агрессии.
Животные
становятся
доверчивыми.
Например,
обезьяны
с
поврежденной миндалиной спокойно подходят к гадюке, вызывавшей ранее у них
ужас, бегство. Видимо, в случае повреждения миндалины исчезают некоторые
врожденные безусловные рефлексы, реализующие память об опасности.
246. Влияние миндалин на иерархические отношения в стае
До операцииПосле удаления миндалин у Дейва
247. Физиология гипоталамуса
Гипоталамус (hypothalamus) или подбугорье — отдел головного мозга,расположенный ниже таламуса, или «зрительных бугров», за что и получил
своё название.
248. Основные структуры гипоталамуса
249. ФУНКЦИИ ГИПОТАЛАМУСА
1. Высший центр регуляции вегетативной нервнойсистемы
2. Высший центр регуляции эндокринных функций
3. Регуляция мотиваций пищевого поведения
4. Высший трофический центр
5. Вегетативное обеспечение и реализация эмоций
6. Половые, оборонительные, агрессивные
мотивации
7. Участие в терморегуляции
8. Участие в регуляции цикла «сон - бодрствование»
250. Поведенческие функции гипоталамуса
Эффекты стимуляции гипоталамусаЛатеральный гипоталамус: жажда, аппетит, увеличение активности
организма, ярость, агрессия.
Вентромедиальное ядро и окружающие его области: чувство
насыщения, снижается аппетит, возникает успокоение.
Перивентрикулярные ядра: страх и боязнь наказания.
Некоторые области переднего и заднего гипоталамуса: усиление
поиска полового партнёра.
Эффекты разрушения гипоталамуса
противоположны эффектам его стимуляции.
Латеральный гипоталамус: потеря аппетита и жажды, пассивность и
малоподвижность.
Вентромедиальная область: неукротимые аппетит и жажда,
жестокость и ярость.
251. Центры поощрения
В различные области мозга обезьянывводили электроды, и обезьяна, нажимая на
рычаг, включала контакты стимулятора. Если
стимуляция
выбранной
области
мозга
приносила животному чувство удовольствия,
то оно нажимало на рычаг снова и снова,
иногда более тысячи раз в час. Более того,
если животному предоставляли на выбор
самую вкусную пищу или стимуляцию, то
животное всё равно выбирало электрическую
стимуляцию.
252. Центры поощрения и наказания
Личные оценки могут характеризоватьощущения как приятные или неприятные
(поощрительные или наказывающие, или как
приносящие удовольствие или вызывающие
неудовольствие). Электрическая стимуляция
некоторых лимбических областей доставляет
удовольствие, раздражение других зон —
боль, страх, защиту, реакции нападения или
избегания. Степень стимуляции этих двух
противоположно
отвечающих
систем
существенно влияет на поведение животных.
253.
• Главные(первичные)
центры
поощрения
располагаются по ходу медиального пучка переднего
мозга и особенно в латеральном и медиальном ядрах
гипоталамуса. Латеральное ядро наиболее реактивно
среди всех центров, так как даже небольшое
усиление стимула может вызвать ярость. Это
положение справедливо для многих областей мозга,
где слабые стимулы вызывали чувство поощрения, а
более сильные — чувство наказания.
• Менее реактивные центры поощрения, которые
можно
назвать
вторичными,
локализуются
в
перегородке,
миндалевидном
теле,
некоторых
областях таламуса, базальных ганглиях и покрышке
среднего мозга
254. Центры наказания
и реакции избегания расположены вцентральном сером веществе, окружающем
сильвиев водопровод, в среднем мозге и в
околожелудочковых зонах гипоталамуса и
таламуса.
Менее
реактивные
центры
наказания находятся в миндалевидном теле и
гиппокампе. Раздражение центров наказания
часто полностью подавляет активность
центров поощрения и удовольствия (т.е.
наказание
и
страх
могут
превышать
удовольствие и поощрение?).
255.
• Яростьвозникает
при
активации
центров наказания. В этом состоянии
даже легкая провокация вызывает
нападение.
• Противоположная
эмоциональная
реакция наблюдается при стимуляции
центров
поощрения
в
виде
безмятежности и смирения.
256. Роль поощрения и наказания в поведении, обучении и памяти
Всё или почти все, что делает человек, имеетотношение к поощрению и наказанию.
Следовательно,
центры
поощрения
и
наказания являются одними из наиболее
важных контролёров нашей физической
активности,
побуждений,
антипатий,
мотиваций. Центры поощрения и наказания
лимбической системы существенно влияют
на отбор получаемой информации. Обычно
около 99% информации удаляется и для
закрепления в памяти остаётся не более 1%.
257.
• Привыкание. Новые сенсорные стимулы почтивсегда возбуждают значительные области коры
больших полушарий. Повторение этих же стимулов
приводит к почти полному затуханию корковых
ответов (если сенсорное научение не вызывает
чувства поощрения).
• Закрепление. Если стимулы вызывают поощрение
или наказание выше индифферентного уровня, то
корковые ответы прогрессивно становятся всё более
и более интенсивными во время повторной
стимуляции и ответ вновь усиливается. Животное
прочно запоминает только то, что поощряется или
наказывается,
и
полностью
игнорирует
индифферентные сенсорные стимулы.
258. Регуляция вегетативных (висцеральных) функций
• Осуществляется преимущественно черезгипоталамус, куда информация поступает от
различных участков лимбической системы –
особенно миндалин
• Происходит изменение секреции гормонов
аденогипофиза (АКТГ и гонадотропинов)
• Частота сердечных сокращений
• Моторики и секреции желудка и кишечника
259. Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль играют гиппокамп и
связанные с ним задние зоны лобной коры. Их основная
деятельность необходима для консолидации памяти – перехода
кратковременной в долговременную память.
Повреждение гиппокампа у человека вызывает резкое нарушение
усвоения новой информации, образования промежуточной и
долговременной памяти
Сенсорная функция лимбической системы
Находится
корковый
отдел
обонятельного
анализатора
(парагиппокампальная извилина, ее крючок, гиппокамп). Его главный
эфферентный выход через свод, мамиллярные тела, передние ядра
гипоталамуса на другие структуры лимбической системы, что ведет к
резко выраженному компоненту в обонятельном восприятии
(ароматерапия).
260. ЭМОЦИОНАЛЬНЫЙ МОЗГ
261. ЦИТАТА
• «Являетсяли
эмоция
продуктом
волшебства
или
физиологическим
процессом,
который
зависит
от
анатомического механизма?
Я
думаю что эмоции - настолько важная
функция, что, каков бы ни был их
механизм,
он
должен
иметь
морфологическую основу»
Пейпс, 1937
262. Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь,эмоции, являются субъективным компонентом мотиваций, которые
запускают и реализуют поведение, направленное на удовлетворение
возникших потребностей
В структуре эмоций выделяют:
• Собственно эмоциональные переживания
• Периферические (вегетативные и соматические) проявления
Важную роль при формировании эмоций играют поясная извилина и
амигдала
Амигдала – ее раздражение вызывает у человека отрицательные
эмоции: страх, гнев, ярость
Удаление амигдалы – снижает агрессивность, повышает
тревожность, неуверенность в себе. Кроме этого – она участвует в
процессе сравнения конкурирующих эмоций, выделяя
доминирующую эмоцию (и мотивацию) и как следствие, влияет на
выбор поведения
Поясная извилина – играет роль главного интегратора различных
систем мозга, участвующих в формировании эмоций
263.
Эмоция—
специфическое
состояние
психической сферы, обусловленная рефлекторной
реакцией организма на внешние и внутренние
раздражения,
характеризующееся
ярко
выраженной
субъективной
окраской
и
включающие
практически
все
виды
чувствительности.
264.
Эмоциональное возбуждение как результатопределенной
мотивационной
деятельности
теснейшим образом связано с удовлетворением
трех основных потребностей человека: пищевой,
защитной и половой.
Мотивационное возбуждение, ассоциируемое с
разными эмоциональными состояниями (жажда,
голод, страх), мобилизует организм к быстрому и
оптимальному удовлетворению потребности.
Удовлетворенная потребность реализуется в
положительной эмоции,
265.
Удовлетворенная потребность вызываетэмоциональное переживание положительного
характера
и
определяет
направление
поведенческой деятельности.
Положительные эмоции, закрепляясь в
памяти, выполняют важную роль в механизмах
формирования целенаправленной деятельности
организма.
266.
Эмоции, проявляются при недостатке точныхсведений
и
путей
достижения
жизненных
потребностей. Такое представление о природе эмоции
позволяет формировать ее информационную природу
в следующей форме (П. В. Симонов):
Э=П (Н—С),
где: Э — эмоция (определенная количественная характеристика эмоционального
состояния организма, обычно выражаемая важными функциональными параметрами
физиологических систем организма, например частота сердечных сокращений, артериальное
давление, уровень адреналина в организме и т.д.);
П— жизненно важная потребность организма (пищевые, оборонительные, половые
рефлексы), направленная на выживание индивида и продолжение рода, у человека
дополнительно еще определяемая социальными мотивами;
Н — информация, необходимая для достижения цели, удовлетворения данной потребности;
С — информация, которой владеет организм и которая может быть использована для
организации целенаправленных действий.
267.
По Г. И. Косицкому, оценка величиныэмоционального напряжения определяется по
формуле:
СН = Ц (Ин∙Вн∙Эн— Ис∙Вс∙Эс),
где: СН — состояние напряжения,
Ц— цель,
Ин,Вн,Эн — необходимые информация, время и энергия,
Ис, Дс, Эс — существующие у организма информация, время и энергия.
268.
Первая стадия напряжения (CHI) —состояние внимания, мобилизация активности,
повышение работоспособности. Эта стадия имеет
тренирующее
значение,
повышая
функциональные возможности организма.
Вторая
стадия
напряжения
(CHII)
характеризуется максимальным увеличением
энергетических ресурсов организма, повышением
артериального давления, увеличением частоты
сердцебиений, дыхания. Возникает стеническая
отрицательная эмоциональная реакция, имеющая
внешнее выражение в форме ярости, гнева.
269.
Первая стадия напряжения (CHI) —состояние внимания, мобилизация активности,
повышение работоспособности. Эта стадия имеет
тренирующее
значение,
повышая
функциональные возможности организма.
Вторая стадия напряжения (CHII) —
характеризуется максимальным увеличением
энергетических ресурсов организма, повышением
артериального давления, увеличением частоты
сердцебиений, дыхания. Возникает стеническая
отрицательная эмоциональная реакция, имеющая
внешнее выражение в форме ярости, гнева.
270.
Третья стадия (СНШ) — астеническаяотрицательная
реакция,
характеризующаяся
истощением ресурсов организма и находящая
свое психологическое выражение в состоянии
ужаса, страха, тоски.
Четвертая стадия (CHIV) — стадия невроза.
Отрицательная эмоция возникает в случае,
когда Н>С и, напротив, положительная эмоция
ожидается, когда H < С.
271.
В теории функциональной системы по П. К.Анохину нейрофизиологическая природа эмоций
связывается с представлениями о функциональной
организации приспособительных действий животных
и человека на основе понятия об «акцепторе
действия».
Сигналом к организации и функционированию
нервного аппарата отрицательных эмоций служит
факт рассогласования «акцептора действия» —
афферентной модели ожидаемых результатов с
афферентацией
о
реальных
результатах
приспособительного акта.
272. Регуляция вегетативных (висцеральных) функций
• Осуществляется преимущественно черезгипоталамус, куда информация поступает от
различных участков лимбической системы –
особенно миндалин
• Происходит изменение секреции гормонов
аденогипофиза (АКТГ и гонадотропинов)
• Частота сердечных сокращений
• Моторики и секреции желудка и кишечника
273. Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь,эмоции, являются субъективным компонентом мотиваций, которые
запускают и реализуют поведение, направленное на удовлетворение
возникших потребностей
В структуре эмоций выделяют:
• Собственно эмоциональные переживания
• Периферические (вегетативные и соматические) проявления
Важную роль при формировании эмоций играют поясная извилина и
амигдала
Амигдала – ее раздражение вызывает у человека отрицательные
эмоции: страх, гнев, ярость
Удаление амигдалы – снижает агрессивность, повышает
тревожность, неуверенность в себе. Кроме этого – она участвует в
процессе сравнения конкурирующих эмоций, выделяя
доминирующую эмоцию (и мотивацию) и как следствие, влияет на
выбор поведения
Поясная извилина – играет роль главного интегратора различных
систем мозга, участвующих в формировании эмоций
274. Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль играют гиппокамп и
связанные с ним задние зоны лобной коры. Их основная
деятельность необходима для консолидации памяти – перехода
кратковременной в долговременную память.
Повреждение гиппокампа у человека вызывает резкое нарушение
усвоения новой информации, образования промежуточной и
долговременной памяти
Сенсорная функция лимбической системы
Находится корковый отдел обонятельного анализатора
(парагиппокампальная извилина, ее крючок, гиппокамп). Его главный
эфферентный выход через свод, мамиллярные тела, передние ядра
гипоталамуса на другие структуры лимбической системы, что ведет к
резко выраженному компоненту в обонятельном восприятии
(ароматерапия).
275.
ФИЗИОЛОГИЯКОРЫ БОЛЬШИХ
ПОЛУШАРИЙ ГОЛОВНОГО
МОЗГА
276.
277. Вызванные потенциалы в коре
Вторичный ответПервичный ответ
278.
279. Расположение нейронов и нервных волокон в коре больших полушарий. Номера слоёв коры I - VI
280. Слои коры больших полушарий
• 1 слой - верхний молекулярный - ветвления дендритовпирамидных нейронов, редкие горизонтальные нейроны и
клетки-зерна, волокна неспецифических ядер таламуса
• 2 слой - наружный зернистый - звездчатые клетки, пути
реализующие циркуляцию импульсов, волокна неспецифических ядер таламуса
• 3 слой - наружный пирамидный - малые пирамидные клетки
и корково-корковые связи различных извилин коры
• 4 слой - внутренний зернистый - звездчатые клетки, окончание
специфических таламокортикальных путей
• 5 слой - внутренний пирамидный - крупные пирамидные
клетки Беца - выходные нейроны кортико - мозговых путей
• 6 слой - полиморфных клеток - кортикоталамические пути
281. КБП
В I – IV слоях коры происходит восприятие иобработка поступающей информации
В II и III слоях коры осуществляется кортикокортикальные ассоциативные связи
В V и VI слоях коры формируются
эфферентные
пути.
Клетки
Беца
расположенные в двигательной коре образуют
эфферентные
кортико-спинальные
и
кортико-бульбарный
двигательные
пути
(пирамидные). Веретенообразные клетки VI
слоя формируют кортико-таламические пути
282. Функциональная организация коры
• Функциональная единица коры - вертикальнаяколонка диаметром около 500 мкм - макромодуль
Колонка - зона распределения разветвлений
одного восходящего афферентного
таламокортикального волокна
Каждая колонка содержит до 1000
нейронных ансамблей - микромодули
Возбуждение одной колонки тормозит
соседние колонки
283. Нейронные колонки
В коре имеются функциональные объединениянейронов,
расположенные
в
цилиндрике
диаметром 0,5-1 мм, включая все слои коры и
содержащем
несколько
сотен
нейронов.
Колончатый принцип конструкции коры впервые
был описан Лоренто де Но: Lorento de no R.
Studies on structure of the cerebral cortex. J.
Physiol. Neurol., 1933, v. 45, P. 381-392.
Они обнаружены в моторной коре и различных
зонах сенсорной коры. Соседние колонки могут
частично
перекрываться,
а
также
взаимодействовать друг с другом по принципу
латерального торможения и осуществлять
саморегуляцию
по
типу
возвратного
торможения.
284.
Колончатая организациякорковых нейронов.
А. Сагиттальный разрез
через постцентральную
извилину.
Б. Рецептивные поля
(ладонь обезьяны) пяти
нейронов.
В. Колонка
функциональный элемент
коры. То, что нейроны во
всех колонках обладают
сходными функциями,
отчасти объяснимо
анатомическими
соображениями.
285.
Сильно упрощенная схема нейронов, их сетей, афферентных иэфферентных связей в различных слоях коры большого мозга
А — локализация и форма двух основных типов корковых нейронов; В
— корково—корковые входы и выходы (ассоциативные а
комиссуральные волокна); В — таламо—корковые (неспецифичные и
специфичные) и корково—таламические связи; Г — синаптические
входы пирамидной клетки, аксон которой идет к субталамическим
отделам (стволу мозга и спинному мозгу); Д — обобщенная схема
корковых контуров.
286. ФУНКЦИОНАЛЬНЫЕ ЗОНЫ КОРЫ
• СЕНСОРНЫЕ ( зрительные, слуховые,кожные и др.)
• МОТОРНЫЕ ( первичные, вторичные,
комплексные)
• АССОЦИАТИВНЫЕ ( лобные, теменные,
височные) - полисенсорность,
пластичность, длительность хранения
следов
287. Четыре основные доли коры головного мозга (лобная, височная, теменная и затылочная); вид сбоку. В них расположены первичная
двигательная исенсорная области, двигательные и сенсорные области более высокого
порядка (второго, третьего и т.д.) и ассоциативная (неспецифичная) кора
288. Локализация функций в КБП
Как современная нейробиологическая концепция локализациифункций
базируется
на
принципе
МНОГОФУНКЦИОНАЛЬНОСТИ
корковых
полей,
т.е.
возможность данной корковой структуре включаться в
обеспечение различных форм деятельности, реализуя при
этом основную, генетические присущую ей функцию:
• Степень мультифункциональности неодинакова: в полях
ассоциативной коры она выше, чем во вторичных сенсорных
полях, а в корковых структура выше, чем в стволовых.
• В основе лежит многоканальность поступления в кору
афферентного возбуждения.
• Перекрытия афферентных возбуждений, особенно на
таламическом и корковом уровне.
• Модулирующие влияния различных структур (например,
неспецифического таламуса или базальных ганглиев) на
корковые функции.
• Взаимодействие корково-подкорковых и межкорковых путей
обработки информации.
289. Цитоархитектоническая карта коры головного мозга человека по Бродману.
290. Основные зоны коры мозга
291. Локализационная карта Клейста. Наружная поверхность мозга
292. Локализационная карта Клейста. Внутренняя поверхность мозга
293. Сенсорные зоны КБП
В этих зонах проецируется информацияот рецепторов всех рецептивных полей
(понятия – проекционная кора, корковые
отделы анализаторов).
Расположены:
•Теменная
•Височная
•Затылочная
Афферентная импульсация поступает от
специфических сенсорных систем
294. В сенсорных зонах коры различают
Первичныесенсорные
поля
(проекционные или центральные поля)
• Их структурная организация завершается
к моменту рождения ребенка
• Состоят
преимущественно
из
моносенсорных цепей и формируют
ощущение качества раздражителя
• Имеется
четкое
пространственное
(топографическое)
представительство
частей тела и их рецепторных полей
295. Представительство вторичных сенсорных зон
Их развитие происходит в первые месяцы послерождения, что связано с образованием многочисленных
внутри- и межанализаторных связей в ответ на
действие различных раздражителей,
К ним относится постцентральная извилина теменной
доли и ее соответствующая часть на медиальной
поверхности
полушарий
(поля
1-3)
–
поля
соматосенсорной области (S1) при этом, поле 3 – как
первичное сенсорное поле, а 1-2 – как вторичные поля.
В этих областях имеются проекции:
• Кожной чувствительности противоположной стороны
тела от: тактильных, болевых, и температурных
рецепторов
• Проприорецептивной чувствительности от: мышечных,
суставных и сухожильных рецепторов
• Интероцептивной чувствительности
296. Проекции чувствительности в коре постцентральной извилины
Представлены:• Головы
и
верхних
отделов
туловища
–
в
нижнелатеральных участках
• Нижней половины тела и ног – в верхнемедиальных зонах
• Нижней части голени и стоп – в коре парацентральной
дольки на медиальной поверхности
Предположительно,
что
в
зоне
тактильной
чувствительности языка расположена проекция вкусовой
чувствительности
Кроме SI выделяют SII-соматосенсорную область, которая
расположена в стенке боковой борозды на границе ее
пересечения с центральной бороздой, в которой
локализация поверхности тела:
• Менее четкая
• Поступает информация не только противоположной
стороны, но и от «своей» стороны
• Возможна моторная координация двух сторон тела
297. Проекции частей тела в соматосенсорной зоне коры больших полушарий
298. Слуховая сенсорная кора
Расположена в верхней височной извилине и представлена:• Первичным проекционным полем
• Вторичной слуховой корой
Первичное проекционное поле
Локализуется в коре поперечных височных извилин (поле
41, в глубине боковой борозды), где имеется четкая
топическая проекция различных участков кортиева органа
на звуковые ощущения:
• По громкости
• По тону
• Другим качествам
Вторичная слуховая кора
Расположена в верхней височной извилине (поле 42), где
происходит более сложная обработка звуковой и речевой
информации.
Предположительно, рядом со слуховой зоной находится
вестибулярная проекционная зона.
299. Зрительная сенсорная кора
представлена в затылочной доли (поля 17 и 18).Первичная проекционная зона (поле 17) зрительного
анализатора имеет топическое представительство
рецепторов сетчатки, особенно желтого пятна
(основная проекция в поле 17), что обеспечивает
высокую остроту зрения.
В этой зоне формируется ощущение:
• - яркости
• - контрастности
• - цвета
• - детальный анализ формы неподвижных объектов
• - элементарный анализ движения предметов
300. Вторичная зрительная область (поля 18 и 19)
в ней осуществляется тонкий анализ образов
- цвета
- движения предметов
- формируется зрительное внимание
- перемещение взгляда
- узнавание знакомой обстановки и знакомых лиц
Нейроны этой зоны полисенсорны и отвечают не только на
световые но и на тактильные и слуховые раздражители.
Раздражение этих полей вызывает:
• - зрительные галлюцинации
• - навязчивые ощущения
• - движения глаз
301. Ассоциативные области коры
• - включают участки новой КБП, которые расположенырядом с сенсорными и двигательными, но не выполняют
непосредственно чувствительных или двигательных
функций.
• - филогенетически наиболее молодая часть, у человека
составляет 70 – 75 % неокортекса.
• - нейроны ассоциативных зон полисенсорны, они отвечают
с почти одинаковым порогом на несколько раздражителей:
зрительные, слуховые, кожные и др.
• - полисенсорность нейронов создается как за счет
кортико-кортикальных связей с разными проекционными
зонами, так и за счет афферентных входов от
ассоциативных ядер таламуса, в которых уже произошла
обработка информации.
• - в результате этого ассоциативная кора становится
мощным аппаратом конвергенции различных сенсорных
возбуждений, как от внешней, так и от внутренней среды
организма.
302. Лобная ассоциативная кора (поля 9 – 14)
имеет основной источник информации от ассоциативного медиодорсальногоядра таламуса
- главной функцией этих областей является формирование программ
целенаправленного поведения, особенно в новой для человека обстановке.
Реализация этой функции основывается на других функциях таламолобной
системы:
1. Формирование доминирующей мотивации – которая обеспечивает
направление поведения человека. Эта функция основана на двухсторонних связях
лобной коры с лимбической системой и ролью последней в регуляции эмоций
человека.
2. Обеспечение вероятностного прогнозирования, что выражается изменением
поведения в ответ на изменения обстановки окружающей среды и
доминирующей мотивации.
3. Самоконтроль действий путем постоянного сравнения результата действия с
исходными намерениями.
Лоботомия (пересекаются пути между лобной долей и таламусом) приводит к
развитию «эмоциональной тупости», отсутствие мотивации, твердых намерений
и планов. Люди становятся грубыми, нетактичными, ненадежными.
303. Теменная ассоциативная кора (5, 7, 39, 40)
- получает афферентные вещи от задней группы ассоциативныхядер таламуса
- эфферентные выходы идут на ядра таламуса и гипоталамуса,
моторную кору и ядра экстрапирамидной системы.
Основными функциями таламотеменной системы является:
- гнозис
- формирование «схемы тела»
- праксис
Под гнозисом – понимают функцию разных видов узнавания:
- формы
- величины
- значения предметов
- оценка их пространственных ощущений
- понимание речи и др.
Вариантом гностической функции является формирование в сознании
трехмерной модели тела («схемы тела»).
Под праксисом – понимают целенаправленное действие, которые
обеспечивают хранение и реализацию программ двигательных
автоматизированных актов (рукопожатие, причесывание, зажигание
спички и т.д.).
304. ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ
• 1. Управление врожденнымиповеденческими реакциями при
помощи накопленного опыта
• 2. Согласование внешних и
внутренних мотиваций поведения
• 3. Разработка стратегии поведения и
программы действия
• 4. Мыслительные особенности
личности
305. Двигательные области коры
Выделяют первичную , вторичную и дополнительнуюобласти
- Пирамидные нейроны V слоя этих областей иннервируют
α- и γ- мотонейроны двигательных центров как
непосредственно (моносинаптически), так и через
интернейроны двигательных центров (полисинаптически)
- Пирамидные нейроны двигательной колонки получают
информацию от мышечных, суставных и тактильных
рецепторов через нейроны верхних слоев и формируют
двигательные команды, реализуемые стволовыми и
спинальными центрами, иннервирующими мышцы
одного сустава или одну мышцу.
- Соседние колонки в функциональном плане
перекрываются и являются организаторами движения.
306. Первичная моторная зона коры
- Прецентральная извилина (поле 4)- Нейроны управляют функцией мотонейронов мышц
лица, туловища и конечностей
- Имеется четкая топографическая проекция мышц тела
- Двигательные реакции на раздражения представлены
элементарными сокращениями мышц противоположной
стороны тела (для мышц головы сокращение может
быть билатеральным)
- Поражение пирамидных путей, ведет к нарушению
двигательной активности, особенно сильно тонкие
движения пальцев рук противоположной стороны,
прекращаются произвольные движения нижней части
лица и языка на противополжной стороне.
307. Вторичная и дополнительная двигательная кора
расположена на латеральной поверхности полушарий,впереди прецентральной извилины, и на медиальной
поверхности.
- участвует в осуществлении высших двигательных функций,
связанных с планированием и координацией произвольных
движений
- премоторная кора получает основную импульсацию от
базальных ганглиев, мозжечка и участвует в
перекодировании
- раздражение вызывает сложные координированные
движения: поворот головы, глаз и туловища в
противоположную сторону, содружественные сокращения
сгибателей или разгибателей на противоположной стороне
- с премоторной корой сопряжены двигательные центры,
связанные с социальными функциями человека: центр
письменной речи в заднем отделе средней лобной извилины;
моторный центр речи Брока, в заднем отделе нижней лобной
извилины (поле 44), обеспечивающий устную речь.
308. Неравномерность представления мускулатуры тела в моторной зоне коры и в гипоталамусе
309. Первичные речевые зоны коры
ЗонаБрока
Зона
Вернике
310. Речевые центры
311. Разделение латеральной коры головного мозга на функциональные поля по Клайсту
312. Локализационная карта Клейста. Внутренняя поверхность мозга
313. Асимметрия полушарий мозга
314. Цитаты
• «Мы говорим левым полушарием»Поль Брока, 1865
• « Сознание локализуется в левом полушарии, а правое
представляет собой простой автомат»
Экклз, 1950
• «Мысль правого полушария - это образ, она служит основой
догадки, интуиции. Эта мысль нерасчлененная, не имея
языкового оформления она скрыта не только для других, но и
для себя. Драма мысли и слова начинается в правом и
заканчивается в левом полушарии. Пройдя этот путь, став
мыслью левого полушария, она становится годной для
сообщения другому и раскрывается для себя. На этом пути
теряется догадка и интуиция, но появляется суждение и
умозаключение, освещенное прожектором осознанности»
Роджер Сперри, 1988
315. Межполушарные различия
• ЛЕВОЕ ПОЛУШАРИЕПРАВОЕ ПОЛУШАРИЕ
• Лучше узнаются стимулы
Словесные
Легко различимые
Знакомые
Несловесные
Трудно различимые
Незнакомые
• Лучше выполняются задачи
На временные отношения
Установление сходства
Идентичность стимулов по
названиям
На пространственные отношения
Установление различий
Идентичность стимулов по
физическим свойствам
• Особенности восприятия
Аналитическое восприятие
Последовательное восприятие
Обобщенное узнавание
Целостное восприятие
Одновременное восприятие
Конкретное узнавание
316. Распознавание лиц левым и правым полушариями больного с расщепленным мозгом
317. Общая схема организации двигательной активности
318. Структура функциональной системы поведения по П.К.Анохину
МОТИВАЦИЯМОТИ ВАЦИЯ
АФФЕРЕНТНЫЙ
СИНТЕЗ
319. ФУНКЦИОНАЛЬНАЯ СИСТЕМА ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ
АФФЕРЕНТНЫЙСИНТЕЗ
ПАМЯТЬ
ЭФФЕРЕНТНЫЙ
СИНТЕЗ
ЭФФЕКТОРНЫЙ
ИНТЕГРАЛ
ОБСТАНОВКА
ЧТО ДЕЛАТЬ?
ПУСКОВОЙ
СИГНАЛ
МОТИВАЦИЯ
ПРИНЯТИЕ
Им -1
РЕШЕНИЯ
КАК ДЕЛАТЬ?
Им -2
Им -3
Им -4
РЕЗУЛЬТАТ
РЕЦЕПТОРЫ
ПРИНЯТИЕ
РЕШЕНИЯ
АКЦЕПТОР РЕЗУЛЬТАТА ДЕ ЙСТВИЯ
320.
Схема организациидвигательной системы.
В иерархическом порядке
представлены связи
между центрами нервной
системы, участвующими в
регуляции позы и
движения. Для упрощения
некоторые высшие
двигательные центры
(мозжечок, базальные
ганглии, двигательный
отдел таламуса) опущены.