
















 жирных кислот")






")




 biology
biologySimilar presentations:


")


")




Биосинтез липидов
1. Биосинтез липидов
БИОСИНТЕЗ ЛИПИДОВКафедра химии
к.м.н., доц. Еликов А.В.
2.
Необходим для создания запасных форм. Биосинтезлипидов (липогенез) начинается с биосинтеза жирных кислот.
Синтез жирных кислот
Система синтеза жирных кислот находится в растворимой
цитозольной фракции многих органов и тканей, таких как
печень, почки, мозг, легкие, молочные железы, жировая ткань.
Биосинтез жировых кислот протекает с участием:
НАДФН2
АТФ
Mg2+
HCO3Ацетил-КоА в качестве субстрата и пальмитиновой кислоты в
качестве конечного продукта.
3.
Особенности биосинтеза жирных кислотСинтез жирных кислот не является простым обращением
реакций β-окисления. Наиболее важными особенностями
являются следующие:
Синтез жирных кислот протекает в цитозоле, в отличие от
распада, который протекает в митохондриях;
Промежуточные продукты синтеза жирных кислот ковалентно
связан с сульфгидрильными группами ацилпереносящего
белка (АПБ);
Многие ферменты синтеза жирных кислот у высших
организмов и человека организованы в мультиферментный
комплекс называемый синтетазой жирных кислот;
Непосредственно ацетил-КоА используется только как
затравка;
4.
Растущаяцепь
жирной
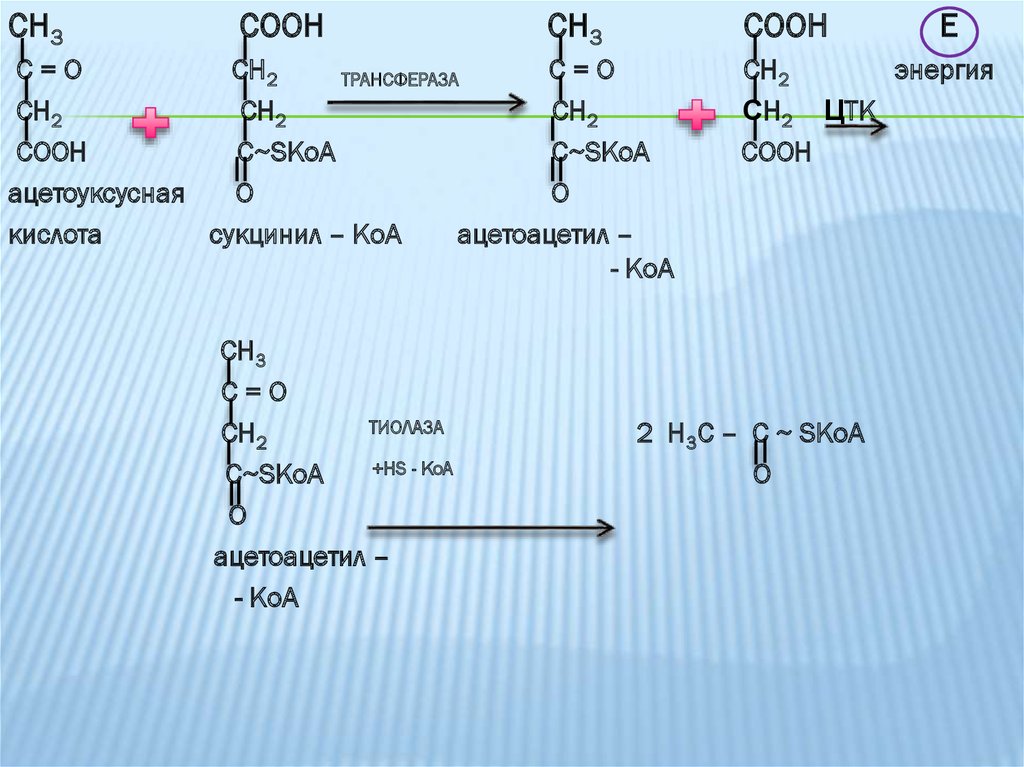
кислоты
удлинятся
путем
последовательного
присоединения
двухуглеродных
компонентов, происходящих из ацетил-КоА. Активированным
донором двухуглеродных компонентов на стадии элонгации
служит
малонил-КоА.
Реакция
элонгации
запускается
высвобождением CO2 ;
Роль восстановителя при синтезе жирных кислот выполняет
НАДФН2;
Синтез жирной кислоты является циклическим процессом,
протекающим на поверхности синтетазы жирных кислот;
Элонгация под действием комплекса синтетазы жирных кислот
останавливается на этапе образования пальмитата C16
Дальнейшая
элонгация
и
введение
двойных
связей
осуществляются другими ферментными системами.
5. Этапы биосинтеза жирных кислот
ЭТАПЫ БИОСИНТЕЗА ЖИРНЫХ КИСЛОТI этап – транспорт ацетил-КоА из митохондрий в
цитоплазму.
Жирные кислоты синтезируются в цитоплазме, а
ацетил-КоА образуется из пирувата в митохондриях.
Мембрана митохондрий не проницаема для ацетилКоА, поэтому транспорт ацетил-КоА через мембрану
обеспечивается специальными механизмами. Роль
карнитина в транспорте ацетил-КоА не велика, т.к. он
переносит только длинноцепочечные жирные кислоты.
Данная проблема решается путем синтеза цитрата:
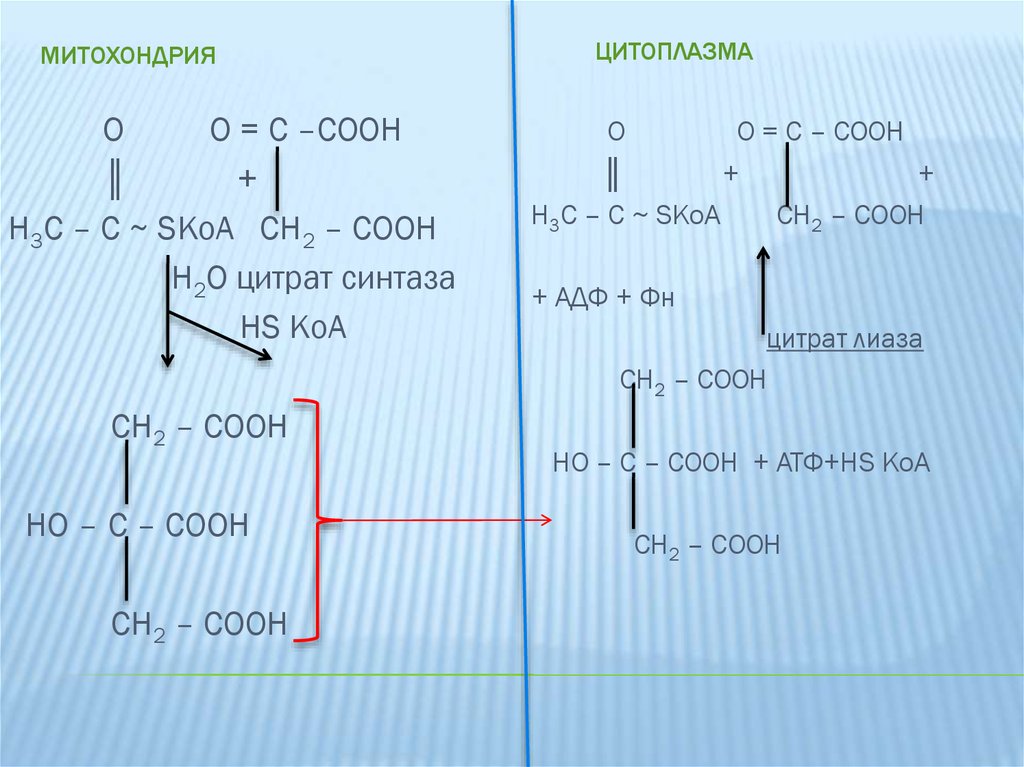
6.
МИТОХОНДРИЯO
O = C –COOH
║
+
H3C – C ~ SKoA CH2 – COOH
H2O цитрат синтаза
HS KoA
CH2 – COOH
HO – C – COOH
CH2 – COOH
ЦИТОПЛАЗМА
O
O = C – COOH
║
+
+
H3C – C ~ SKoA
CH2 – COOH
+ АДФ + Фн
цитрат лиаза
CH2 – COOH
HO – C – COOH + АТФ+HS KoA
CH2 – COOH
7.
Цитрат образуется в митохондриальном матриксепутем конденсации ацетил-КоА и оксалоацетата.
Затем диффундирует в цитоплазму, где расщепляется
цитрат-лиазой.
Цитрат + АТФ + КоА
ацетил-КоА + АДФ + Ф +
оксалоацетат
Таким образом, ацетил-КоА и оксалоацетат
переносятся из митохондрий в цитоплазму с
использованием одной молекулы АТФ.
8.
Источники НАДФН2 для биосинтеза жирных кислотОксалоацетат, образовавшийся в результате переноса
ацетил-КОА в цитоплазму должен быть возвращен
обратно в митохондрию. Данный процесс сопряжен с
генерацией НАДФН2
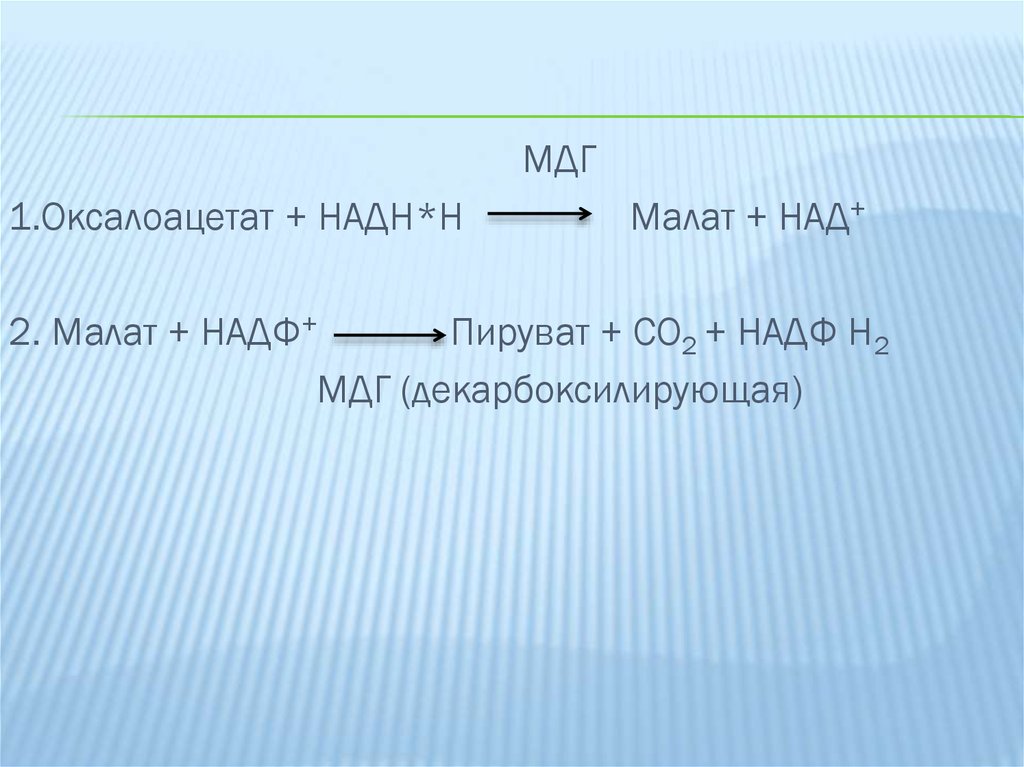
Реакция протекает в 2 этапа:
1. Происходит в цитоплазме и катализируется малатдегидрогеназой;
2. Окислительное декарбоксилирование малата НАДФН2
зависимой малат-дегидрогеназой.
9.
МДГ1.Оксалоацетат + НАДН*Н
Малат + НАД+
2. Малат + НАДФ+
Пируват + СО2 + НАДФ Н2
МДГ (декарбоксилирующая)
10.
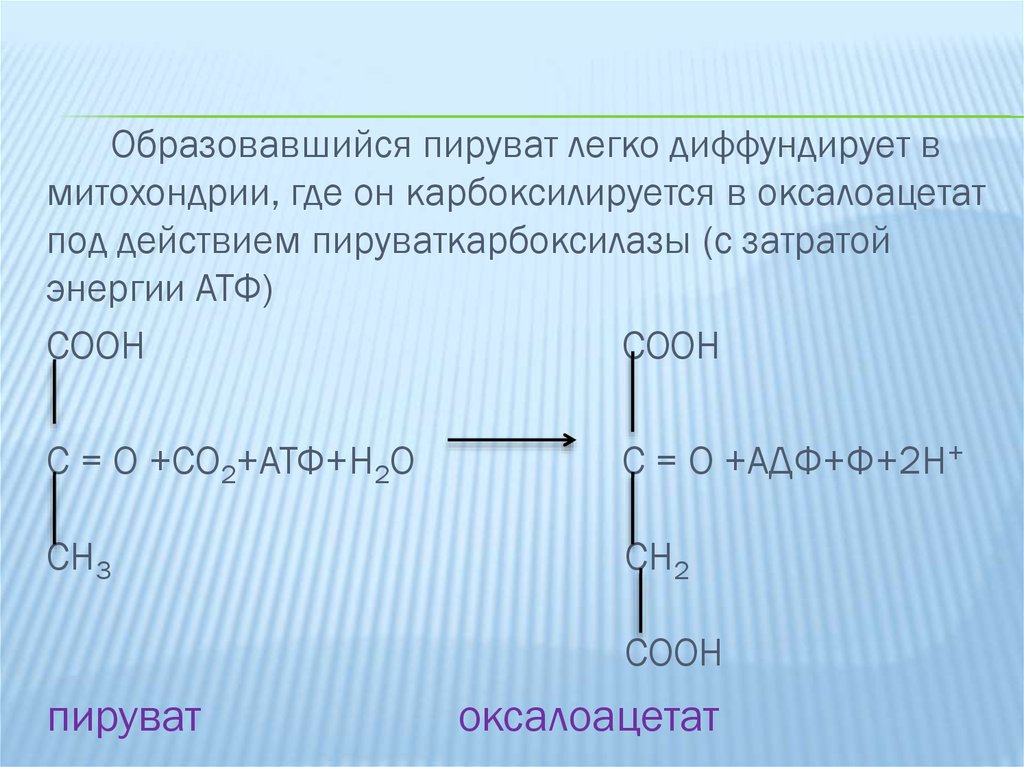
Образовавшийся пируват легко диффундирует вмитохондрии, где он карбоксилируется в оксалоацетат
под действием пируваткарбоксилазы (с затратой
энергии АТФ)
COOH
COOH
C = O +CO2+АТФ+H2O
C = O +АДФ+Ф+2H+
CH3
CH2
COOH
пируват
оксалоацетат
11.
Таким образом, на каждую молекулу ацетил-КоА,которая переходит из митохондрий в цитозоль,
образуется одна молекула НАДФН2.
Следовательно, при переходе 8 молекул ацетилКоА в цитоплазму необходимо для синтеза
пальмитиновой кислоты, образуется 8 НАДФН2. Еще 6
молекул требуемые для этого процесса генерируются
в пентозофосфатном пути.
12.
1.2.
II - этап – образование малонил-КоА.
Является первой реакцией биосинтеза ж.к.
Катализируется ферментом ацетил-КоА-карбоксилазой.
Коферментом является биотин. Реакция заключается в
карбоксилировании ацетил-КоА, источником CO2
является бикарбонат. Реакция протекает в 2 стадии:
Карбоксилирование биотина с участием АТФ;
Перенос карбоксильной группы на ацетил-КоА.
В дальнейшем биосинтезе ж.к. ацетил-КоА
используется как затравка, а непосредственно синтез
идет из малонил-КоА.
13.
H3C│
_
Mg2+
C = O + HCO3 + АТФ E – биотин
SKOA
COOH
│
CH2
│ + АДФ+H3PO4
C=O
SKOA
ацетил - KOA
малонил - KOA
14.
III этап – биосинтез жирных кислот.Строение синтазы жирных кислот представляет из
себя полиферментный комплекс, катализирующий
синтез жирных кислот. Комплекс представляет собой
димер, состоящий из двух идентичных мономеров
1 и 2.
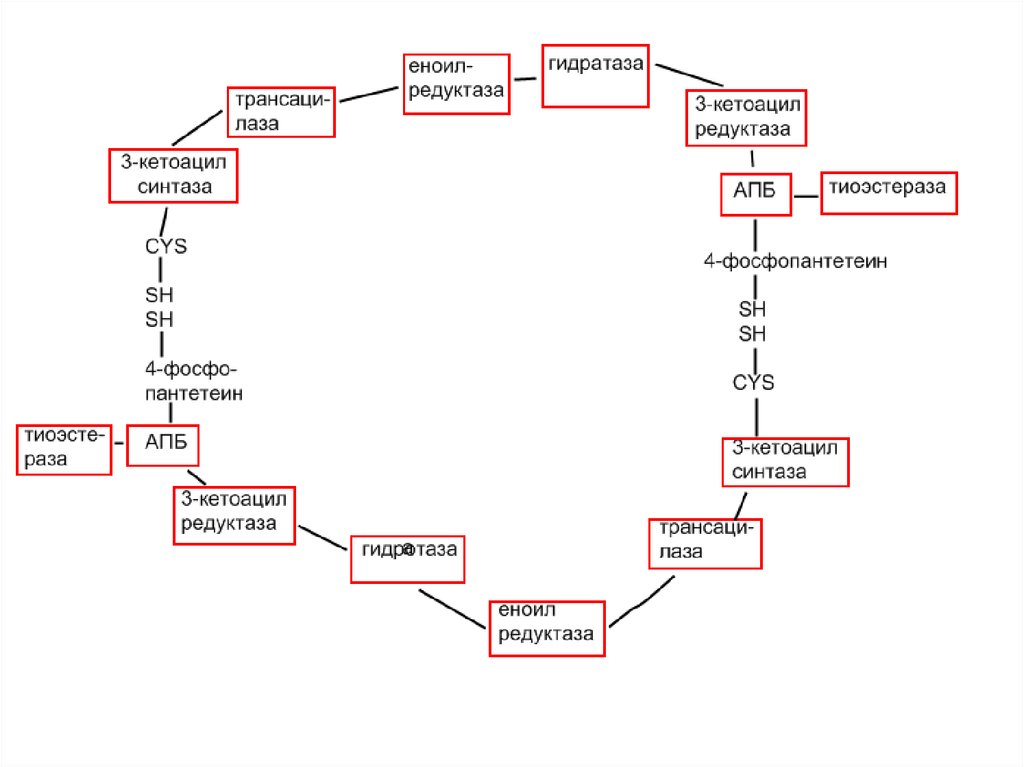
15.
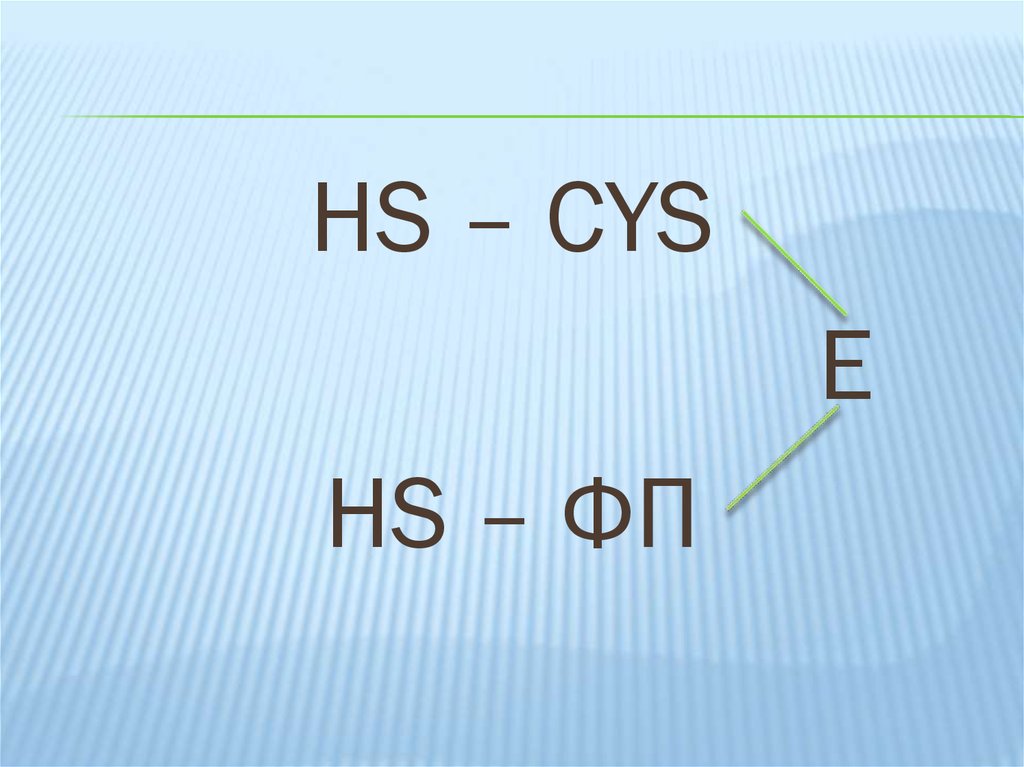
HS – CYS – 1 – ФП – SHHS – ФП – 2 – CYS - SH
16.
Каждый мономер включает 6 индивидуальных ферментови ацилпереносящий белок (АПБ).
Cys-SH-тиоловая группа цистеина.
Сульфгидрильная группа 4-фосфопантетеина одного
мономера расположена в непосредственной близости от SHгруппы остатка цистеина кетоацилсинтетазы, входящий в
состав другого мономера. Это указывает на расположение
мономеров по типу «голова к хвосту». Каждый из мономеров
включает все ферменты, катализирующие биосинтез жирных
кислот. В состав функциональной единицы входят фрагменты
обоих мономеров, при этом половина одного мономера
взаимодействует с комплиментарной половиной другого.
Синтазный
комплекс
одновременно
катализирует
образование 2-х молекул жирных кислот.
17.
HS – CYSЕ
HS – ФП
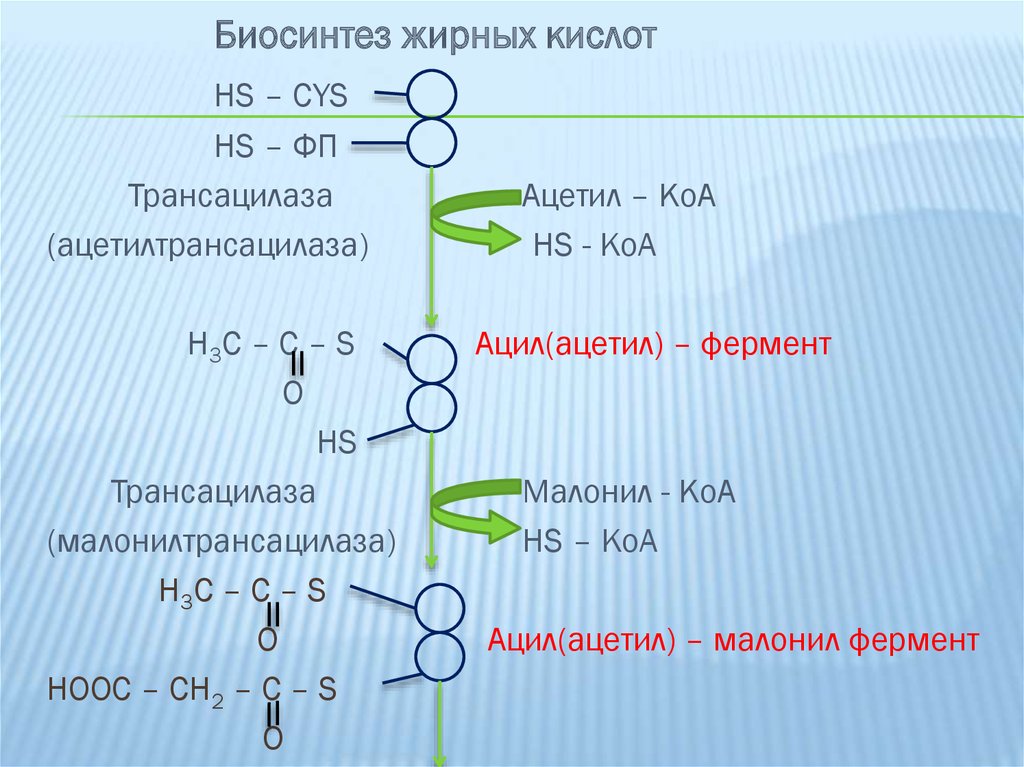
18. Синтез жирных кислот
СИНТЕЗ ЖИРНЫХ КИСЛОТ1-ая
реакция:
ацетил-КоА
под
действием
ацетилтрансферазы переносится на SH-группу
цистеина.
2-ая реакция: малонил-КоА под действием фермента
малонилтрансферазы переносится на SH-группу
фосфопантетина.
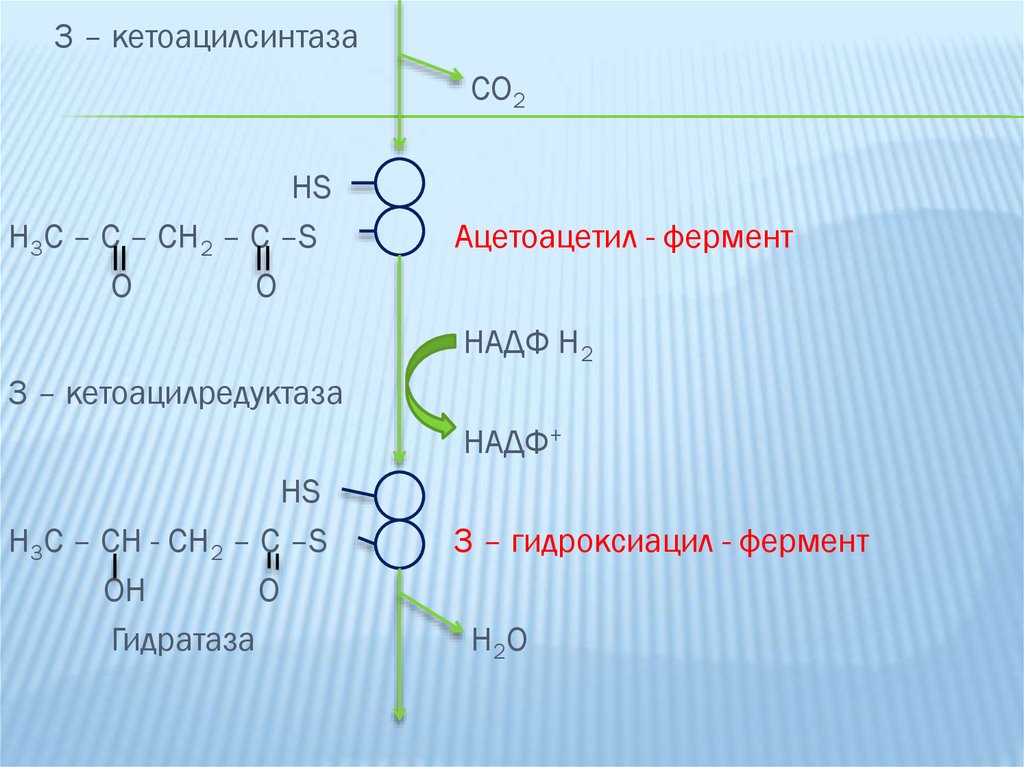
3-ая реакция: перенос ацетильного остатка на
малонинильный с образованием 3-ацетоацетил-Е.
Фермент - 3-кетоацилсинтаза.
4-ая реакция: восстановление 3-кетоацетила-Е до 3гидроксиацетилацил-Е. Фермент- 3-кетоацилредуктаза.
19.
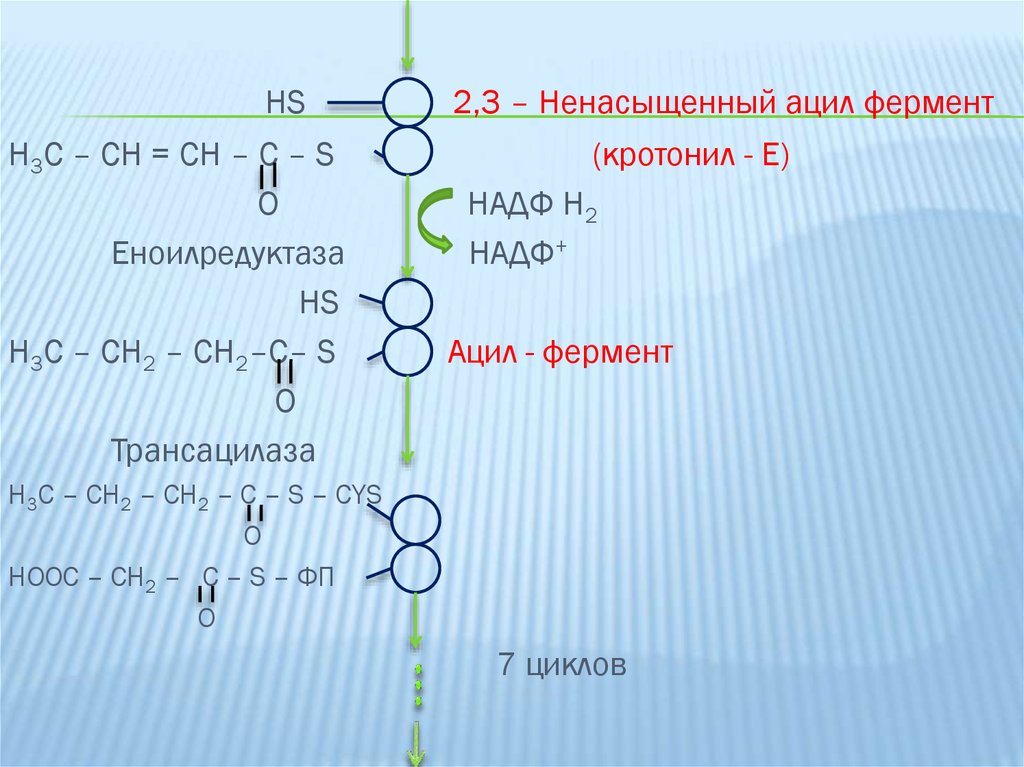
5-ая реакция: заключается в дегидратации 3гидроксиацил-Е ферментом гидратазой, при этомобразуется 2,3-ненасыщенный ацил-Е (кротонил-Е).
6-ая реакция: второе восстановление с участием
НАНДФ2 и фермента еноил-редуктазы, при этом из
кротонил-Е образуется бутирил-Е (ацил-фермент).
Синтезированный бутирил переносится с участием
трансацилазы на SH-группу связанную с остатком
цистеина, где раньше был ацетил-КоА, а на нижнюю
группу связанную с ФП-SH поступает новый
малонильный остаток.
20.
21.
Биосинтез жирных кислотHS – CYS
HS – ФП
Трансацилаза
(ацетилтрансацилаза)
H3C – C – S
O
HS
Трансацилаза
(малонилтрансацилаза)
H 3C – C – S
O
HOOC – CH2 – C – S
O
Ацетил – КоА
HS - KoA
Ацил(ацетил) – фермент
Малонил - КоА
HS – KoA
Ацил(ацетил) – малонил фермент
22.
3 – кетоацилсинтазаCO2
HS
H3C – C – CH2 – C –S
O
O
Ацетоацетил - фермент
НАДФ Н2
3 – кетоацилредуктаза
НАДФ+
HS
H3C – CH - CH2 – C –S
OH
O
Гидратаза
3 – гидроксиацил - фермент
H2O
23.
HSH3C – CH = CH – C – S
О
Еноилредуктаза
HS
H3C – CH2 – CH2–C– S
О
Трансацилаза
2,3 – Ненасыщенный ацил фермент
(кротонил - Е)
НАДФ Н2
НАДФ+
Ацил - фермент
H3C – CH2 – CH2 – C – S – CYS
O
HOOC – CH2 – C – S – ФП
О
7 циклов
24.
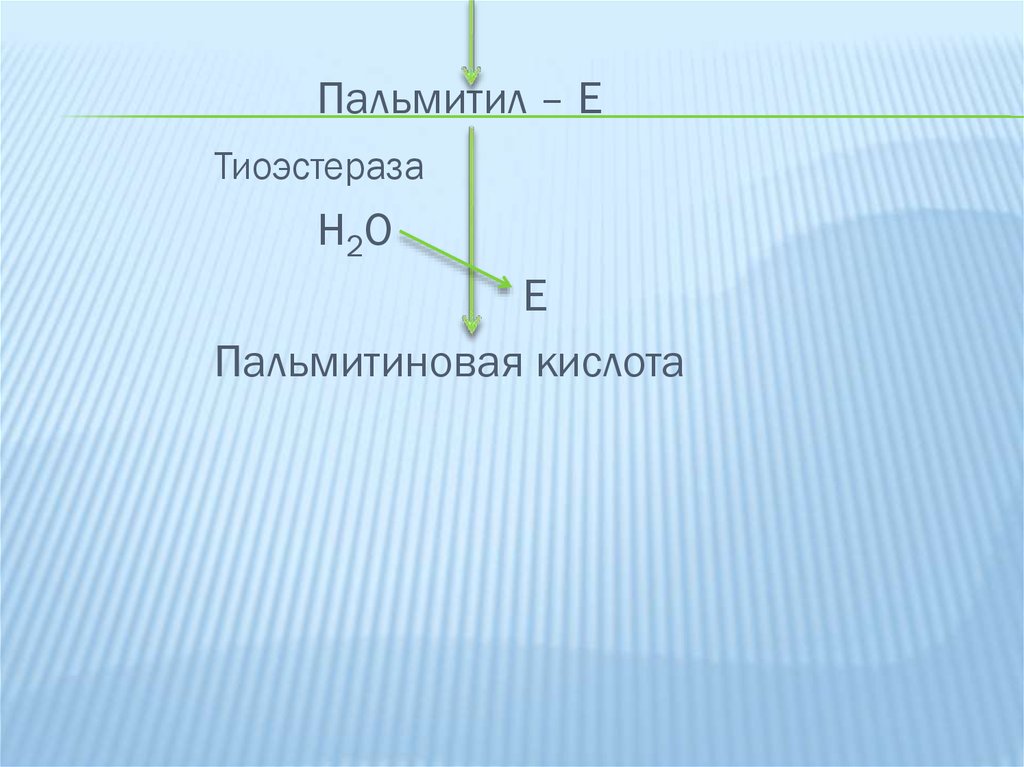
Пальмитил – ЕТиоэстераза
Н2О
Е
Пальмитиновая кислота
25.
Цикл синтеза повторяется.Для синтеза пальмитиновой кислоты требуется 7
таких циклов, соответственно требуется 7 остатков
малонил-КоА и один ацетил-КоА. Образовавшийся
пальмитил-Е
гидролизуется
тиоэстеразой
с
образованием свободной жирной кислоты.
26.
Суммарное уравнение синтеза пальмитиновой кислотыH3C – C(О) ~ SKOA + 7HOOC – CH2 – C(О) ~ SKOA + 14 НАДФ Н2
H3C – (CH2)14 – COOH + 7CO2 + 6H2O + 8 KOA SH + 14 НАДФ+
27.
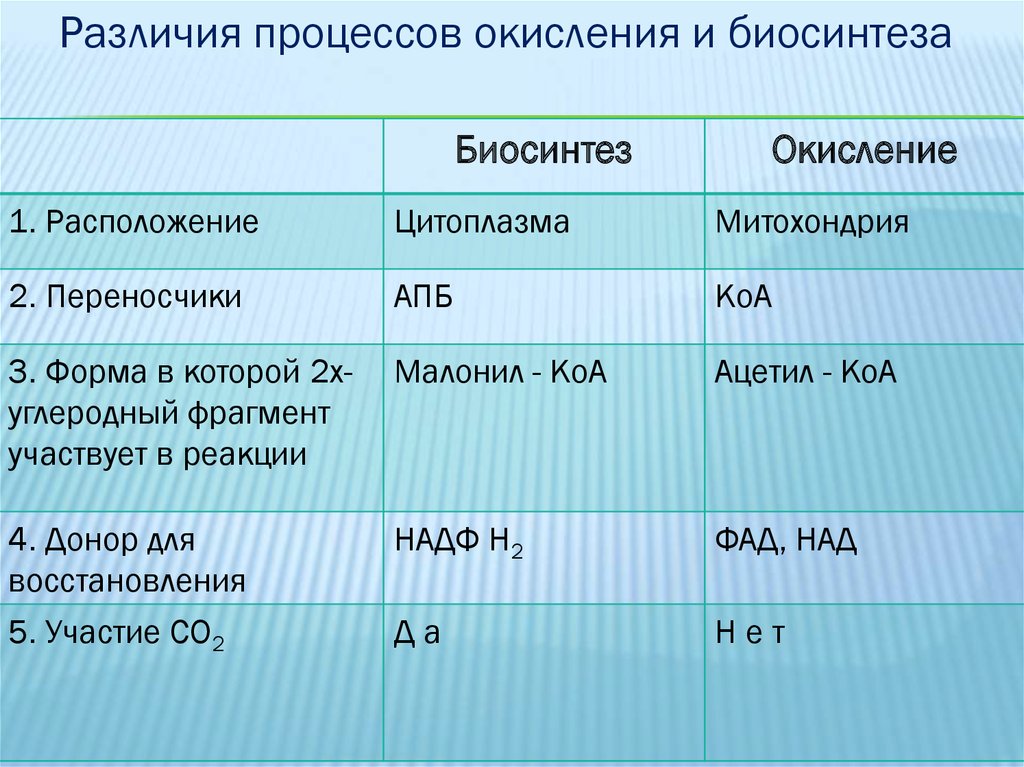
Различия процессов окисления и биосинтезаБиосинтез
Окисление
1. Расположение
Цитоплазма
Митохондрия
2. Переносчики
АПБ
КоА
3. Форма в которой 2хуглеродный фрагмент
участвует в реакции
Малонил - КоА
Ацетил - КоА
4. Донор для
восстановления
НАДФ Н2
ФАД, НАД
5. Участие СО2
Да
Нет
28. Система удлинения (элонгации) жирных кислот
СИСТЕМА УДЛИНЕНИЯ (ЭЛОНГАЦИИ) ЖИРНЫХКИСЛОТ
Данный процесс в основном происходит в микросомах.
Факторы обеспечивающие элонгацию жирных кислот:
Жирные кислоты должны находится в виде ацил-КоА
производных;
Источником двухуглеродных фрагментов C2 является малонилКоА;
В качестве восстановителя используется НАДФН2;
Промежуточными продуктами являются тиоэфиры КоА;
Затравочными молекулами могут служить насыщенные
жирные кислоты (C10 и выше) и ненасыщенные жирные
кислоты.
29.
При голодании элонгация жирных кислотзатормаживается. При образовании миелиновых
оболочек нервных клеток в мозгу резко усиливается
процесс удлинения, в результате образуются C22 C24
- жирные кислоты входящие в состав
сфинголипидов.
30. Синтез ненасыщенных жирных кислот
СИНТЕЗ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТКатализируются микросомальными системами
путем введения двойной связи в КоА-производных
жирных кислот с длинной цепью.
Например,
синтез
олеиновой
кислоты
осуществляется по следующему уравнению:
31.
Стеароил – КоА + НАДН2 + О2(НАДФН2)
Олеоил – КоА + НАД+ + 2Н2О
(НАДФ+)
32.
Таким образом идет синтез из пальмитиновой пальмитиолеиновой кислоты, а из стеариновой –олеиновой.
У
млекопитающих
нет
ферментов,
катализирующие введение связей далее 9-го
углеродного атома. Поэтому полиненасыщенные
жирные кислоты у них синтезироваться не могут и
должны поступать с пищей (линолевая и линоленовая).
Из этих кислот возможен синтез в организме других
полиненасыщенных жирных кислот путем введения
двойных связей до C9 (например, арахидоновая).
33.
Ацетил КоАПальмитиновая кислота
- 2Н
+ С2
Пальмитоолеиновая
кислота (16 : 1)
+ С2
+ С2
Лигноцериновая
кислота (24 : 0)
Стеариновая кислота
(18 : 0)
+ С2
- 2Н
Олеиновая
кислота (18 : 1)
+ С2
+ С2
+ С2
Невроновая
кислота (24 : 1)
34.
Линолевая кислота(поступает с пищей)
18 : 2 (9, 12)
- 2Н
Линоленовая кислота
18 : 3 (6, 9, 12)
+ С2
- 2Н
Арахидоновая кислота
20 : 4 (5, 8, 11, 14)
35. Биосинтез кетоновых тел (ацетоНовых)
БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (АЦЕТОНОВЫХ)К
ацетоновым телами относятся ацетоуксусная
кислота, β- оксимасленая кислота, ацетон.
Кетоновые тела являются нормальным продуктом
обмена. Синтезируются в печени из ацетил-КоА.
36.
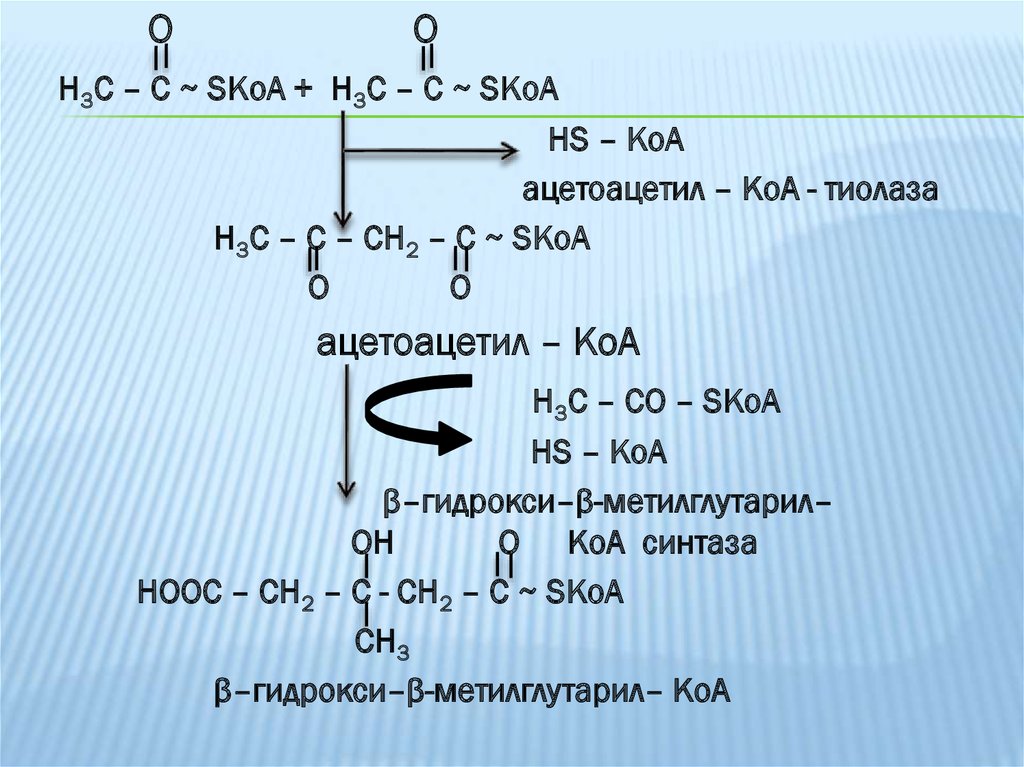
OO
Н3С – С ~ SKоA + Н3С – С ~ SKоA
HS – KoA
ацетоацетил – КоА - тиолаза
Н3С – С – CH2 – C ~ SKoA
O
O
ацетоацетил – КоА
Н3С – CO – SKoA
HS – KoA
β–гидрокси–β-метилглутарил–
OH
O КоА синтаза
HOOC – CH2 – C - CH2 – C ~ SKoA
CH3
β–гидрокси–β-метилглутарил– КоА
37.
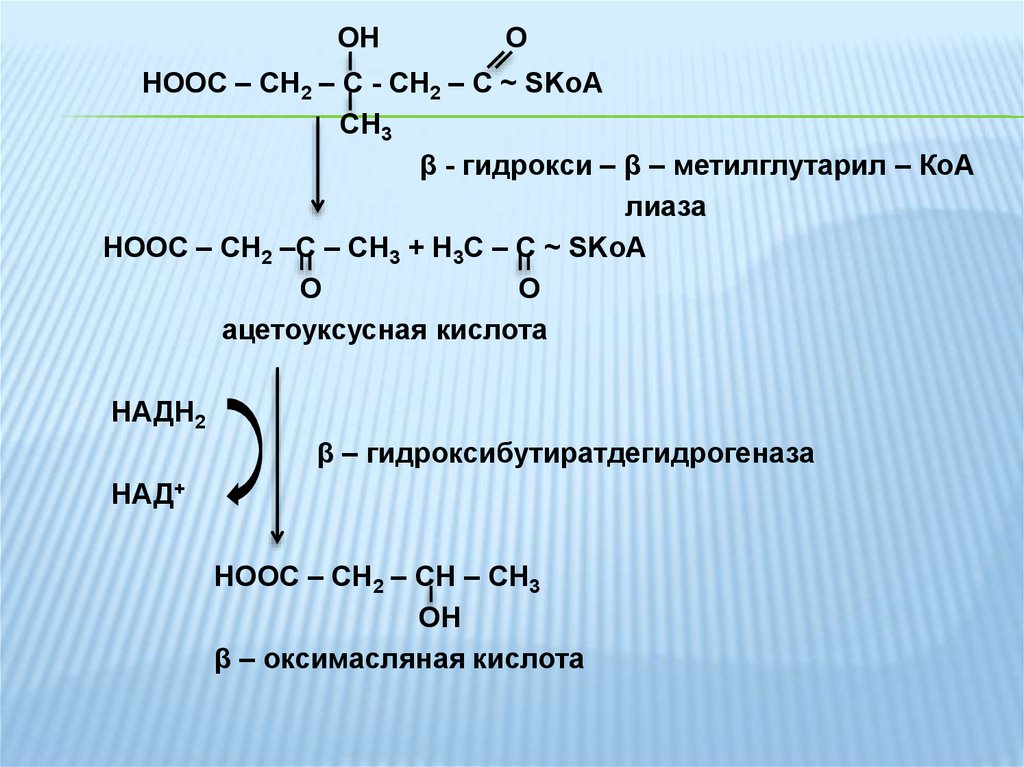
OHO
HOOC – CH2 – C - CH2 – C ~ SKoA
CH3
β - гидрокси – β – метилглутарил – КоА
лиаза
HOOC – CH2 –C – CH3 + Н3С – С ~ SKoA
О
O
ацетоуксусная кислота
НАДН2
β – гидроксибутиратдегидрогеназа
НАД+
HOOC – CH2 – CH – CH3
OH
β – оксимасляная кислота
38.
В организме кетоновые тела выполняют важнуюфункцию
по
поддержанию
энергетического
гомеостаза. Являются поставщиками энергии для
мышц, почек, мозга. Печень не использует кетоновые
тела, т.к. в ней не фермента расщепляющих их и с
током крови они вносятся из печени. Кетоновые тела
является важным источником энергии при голодании.
Ацетон
образуется
при
спонтанном
декарбоксилировании ацетоуксусной кислоты и
энергетического значения не имеет.
Окисление кетоновых тел осуществляется под действием
трансферазы и тиолазы.
39.
СН3СOOH
СН3
COOH
С=О
СН2
С=О
ТРАНСФЕРАЗА
СН2
СН2
СН2
СOOH
C~SKoA
C~SKoA
ацетоуксусная
O
O
кислота
сукцинил – КоА
ацетоацетил –
- КоА
СН3
С=О
ТИОЛАЗА
СН2
+HS - KoA
C~SKoA
О
ацетоацетил –
- КоА
Е
СН2
энергия
СН2 ЦТК
COOH
2 H3C – C ~ SKoA
O
40.
В тех случаях когда скорость образованиикетоновых тел превышает их потребление развиваются
кетозы. Они могут быть физиологическими и
патологическими.
В норме при хорошем питании количество
кетоновых тел в крови не превышает
10 мг/л (0,1 – 0,6 ммоль/л)
В моче 1 мг/сут
Физиологический кетоз возникает при голодании
(связано с исчерпанием запаса легкодоступных
углеводов и вследствие этого увеличения мобилизации
свободных жирных кислот).
41.
Патологический кетоз возникает при тяжелойформе сахарного диабета. Выделение кетоновых тел
с мочой – кетонурия.
Опасность кетоза: ацетоновые тела имеют
кислую реакцию и в больших количествах вызывают
ацидоз.