













")



")
")




 . Дрейф генов как")
:")

")
, мало изменившаяся с Ордовика (около 400 млн л.н.)")



")

 отбора")




 могут очень долго «вымываться» из популяции, т.к. чем")


 – (второй синтез) Произошел синтез классического дарвинизма, генетики (Ф.")

 синтеза (1980 – н.в.)")




")


")

 biology
biologySimilar presentations:










Биогеографические доказательства эволюции
1.
7. Биогеографическиедоказательства эволюции
Географическое распространение животных и
растений соответствует их эволюционной истории
Если два вида недавно произошли от одной популяции,
то они как правило обитают недалеко от ареала исходной
популяции, а значит недалеко друг от друга. Т.о, с точки
зрения эволюционной теории географическое
распределение видов должно быть совместимо с
филогенетическим деревом. Если не принимать во
внимание теорию эволюции, то разумно предположить,
что виды живут в наиболее подходящих для них
условиях. Теория эволюции же предсказывает, что
должно быть много благоприятных для вида мест, в
которых представители вида, тем не менее, отсутствуют в
связи с наличием географических барьеров.
Так и наблюдается в природе.
2.
Примеры: сумчатые встречаютсяпочти исключительно в Австралии
(раньше они водились и на других
материках, но впоследствии
вымерли вытесненные
плацентарными конкурентами). Но
условия Австралии вполне
подходят для плацентарных - так
завезенные кролики и собаки
заселили весь континент.
Двоякодышащие рыбы и
бескилевые птицы (страусы, киви)
встречаются только на юге Южной
Америки, Африки и Австралии
(расхождение Гондваны в
мезозое).
Условия обитания в пустынях
Африки, Америки и Австралии
очень похожи, и растения из одной
пустыни хорошо растут в другой.
Тем не менее, кактусы были
обнаружены только в Америке.
3.
Островная биогеографияПримеры: в Новой Зеландии до появления
человека не было змей, и даже млекопитающих,
но было много эндемичных древних птиц и
растений. На Гавайских островах тоже живет
множество эндемичных (нигде больше не
встречающихся) птиц, растений и насекомых, но
на них полностью отсутствуют местные
пресноводные рыбы, амфибии, рептилии и
наземные млекопитающие. Сейчас завезенные
людьми млекопитающие заполнили эти о-ва и
вытесняют местную фауну. На Галапагоссах
есть эндемичные птицы, случайно завезенные
туда игуана и черепахи, но нет млекопитающих,
амфибий и пресноводных рыб. Исключение –
только летучие мыши.
какапо
Орел Хааста и моа
киви
4.
При этом на тех же островах местные эндемичные видыдали мощную близкородственную радиацию (пример –
вьюрки на Галапагоссах).
Или почему предполагаемый разумный дизайнер создал
эндемичные виды летучих мышей на островах, но не
создал там других млекопитающих? Объясняется это
просто: наземные млекопитающие практически
неспособны пересекать широкие проливы, а летучие
мыши умеют летать.
Еще одним свидетельством в пользу эволюции является
высокое сходство островных флор и фаун с флорами и
фаунами ближайших массивов суши. Например,
животный и растительный мир Галапагосских островов,
несмотря на все своеобразие, явно связан тесными
родственными связями с флорой и фауной ближайшего
материка - Южной Америки.
Островная теория МакАртура (1967).
5.
Остров Пасхи (Рапа Нуи) – самое отдаленное ипотому беднейшее биологическими видами
место на планете.
6.
Соответствие резкости биогеографических границмежду фаунами и геологической истории
Пример: «линия Уоллеса»: в Индонезии (малые Зондские о-ва) есть четкая
граница между азиатской и австралийской флорой и фауной по мелким овам Бали и Ламбок, которые не соединялись в поздней геологической
истории. На западе Индонезии 100 пресноводных видов рыб (на востоке 3),
живут фазаны, дрозды, дятлы, райские мухоловки, сорокопуты и дронго. И
ни одно из этих животных не встречается на востоке. Зато на востоке
водятся казуары, райские птицы, попугаи-лори и какаду, которых
невозможно найти на западе, но они есть в Австралии.
7.
Там где по суше возник переход - фауны переходятодна в другую в соответствии с временем
образования этого перехода (Берингийская суша,
Панамский перешеек). Например в Ю.Америке очень
своеобразная флора и фауна (броненосцы, ламы,
муравьеды), но нет сейчас местных копытных, т.к.
они давно вымерли, а новые не смогли туда снова
проникнуть с севера. Причем чем южнее – тем она
своеобразнее (это коррелирует с геологическим
временем изоляции) (см. Дж. Симпсон «Великолепная
изоляция» 1980)
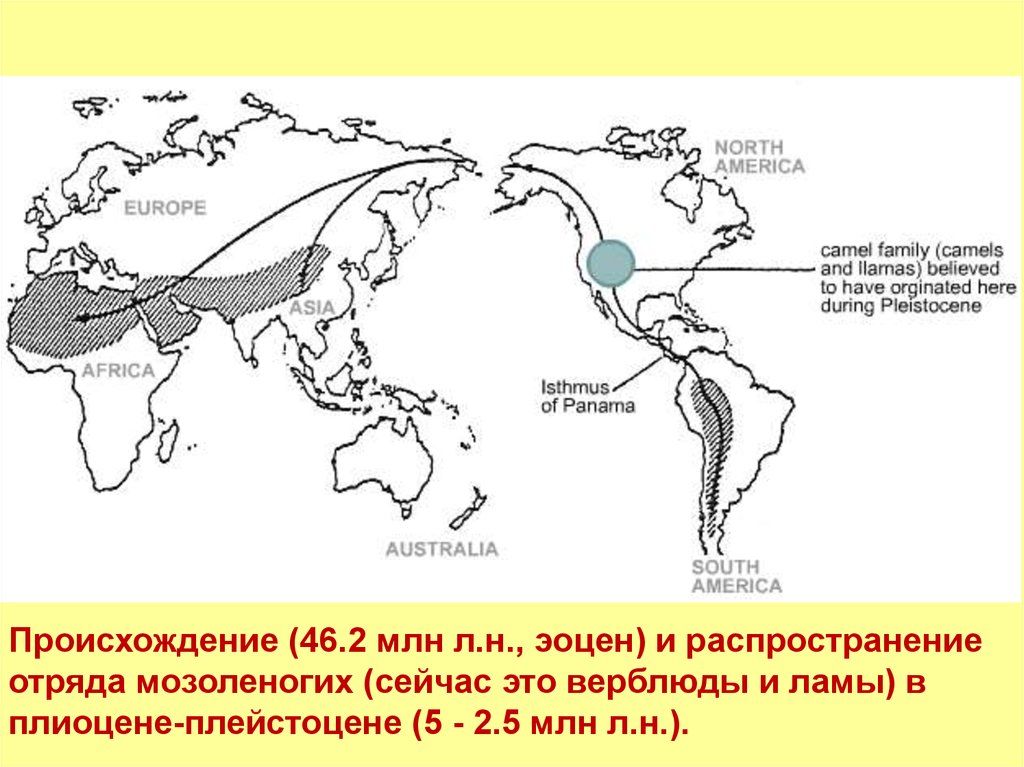
Верблюды и другие мозоленогие имеют
североамериканское происхождение, а дальше
группа распространялась с одного конца в Южную
Америку, с другого - в Старый Свет через
берингийский перешеек. На месте своего
возникновения они вымерли, что подтверждает
набор ископаемых.
8.
Происхождение (46.2 млн л.н., эоцен) и распространениеотряда мозоленогих (сейчас это верблюды и ламы) в
плиоцене-плейстоцене (5 - 2.5 млн л.н.).
9.
За последние 3 млн леттерритория Берингии
поднималась и уходила
под воду, по крайней
мере, 6 раз. Каждый раз,
когда два материка
соединялись,
из Старого света в Новый
и обратно
происходила миграция
животных. Последний раз
это произошло 15-18 и 1011 тыс лет назад.
Панамский перешеек
возник 20 млн л.н. До
этого Ю.Америка более 40
млн лет оставалась о-вом
после распада Гондваны.
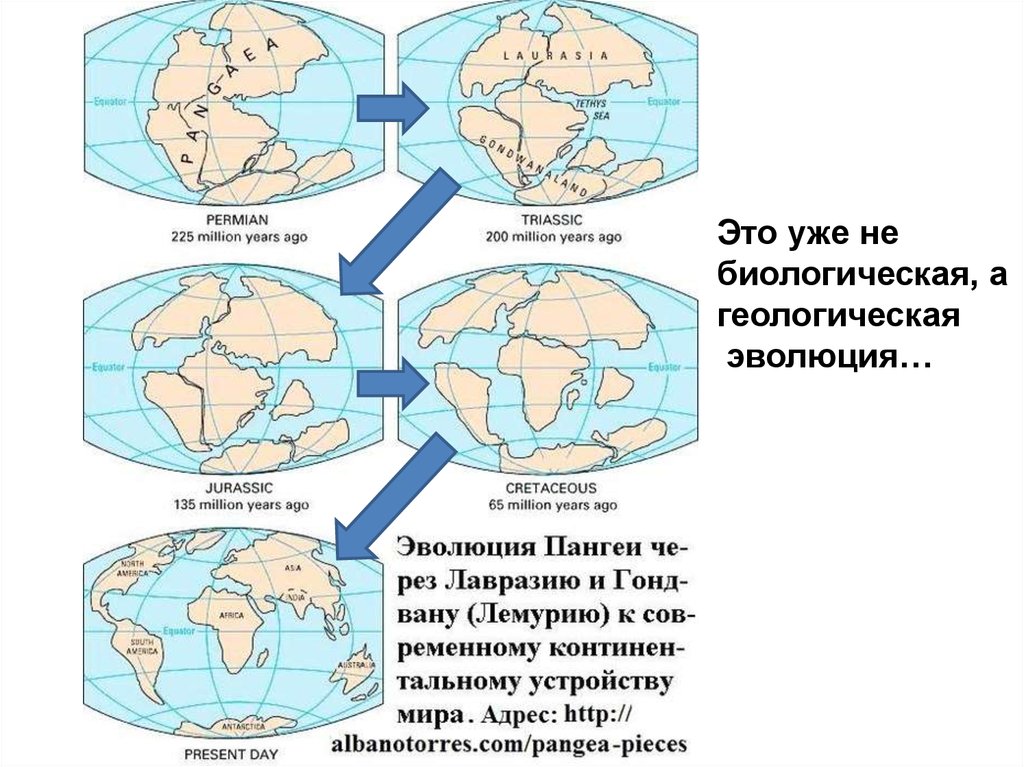
10.
Это уже небиологическая, а
геологическая
эволюция…
11.
Заселение изолированных территорийПопав на большой остров с бедной фауной (или, для
водных животных - в большое озеро), виды-вселенцы с
большой вероятностью испытают адаптивную
радиацию - быстрое видообразование, в ходе которого
они займут ряд свободных экологических ниш. Кроме
галапагосских вьюрков, хорошие примеры таких
быстрых адаптивных радиаций - это рачки-бокоплавы
озера Байкал (зачем Создателю нужно было создавать
специально для озера Байкал 250 нигде больше не
встречающихся видов рачков-бокоплавов, пусть
антиэволюционисты придумывают сами), рыбыцихлиды Великих Африканских озер (Малави,
Виктория), мушки дрозофилы и птицы нектарницы
Гавайских островов и др.
12.
Эти виды равноногих рачков(бокоплавов) – эндемики о.Байкал
13.
Эндемичные рыбки-цихлиды озера Малави (слева) имеют большесиневатые оттенки, тогда как из озера Виктория (справа) –
желтоватые (коррелирует с большей мутностью воды).
14.
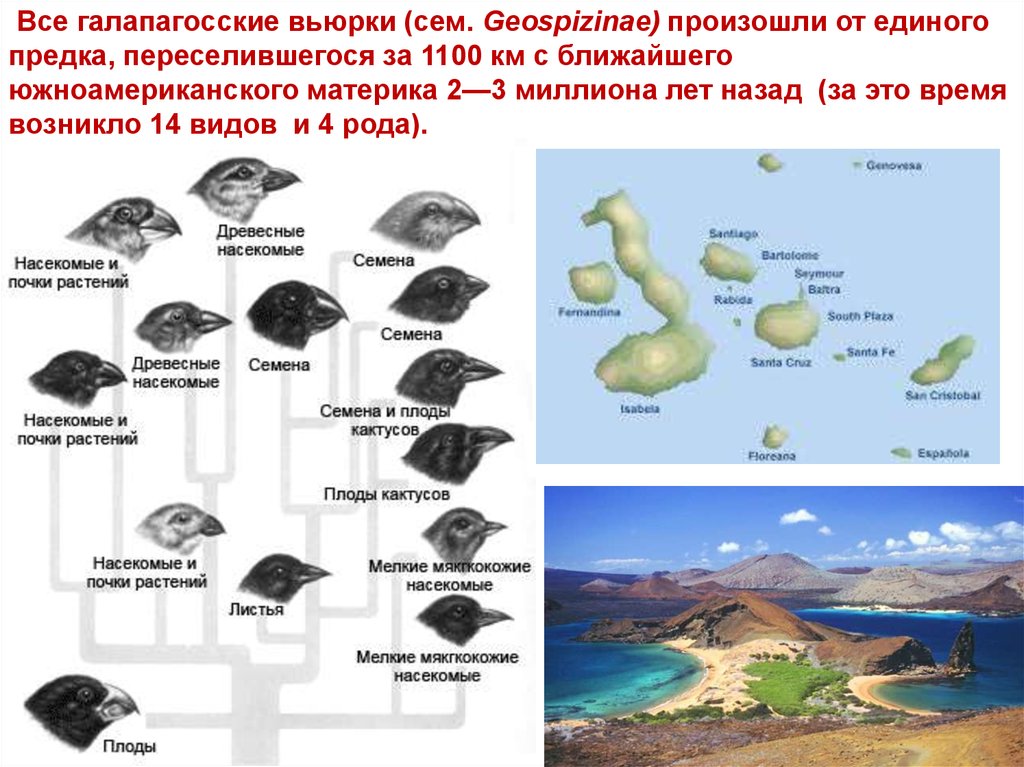
Все галапагосские вьюрки (сем. Geospizinae) произошли от единогопредка, переселившегося за 1100 км с ближайшего
южноамериканского материка 2—3 миллиона лет назад (за это время
возникло 14 видов и 4 рода).
15.
Параллельная эволюция наразобщенных массивах суши
Трудно объяснить иначе, как с точки
зрения эволюции, почему на разных
континентах существуют внешне
сходные формы, но совершенно разные
по ряду других, более глубоких
признаков, больше роднящих их с
внешне непохожими видами на том же
континенте.
16.
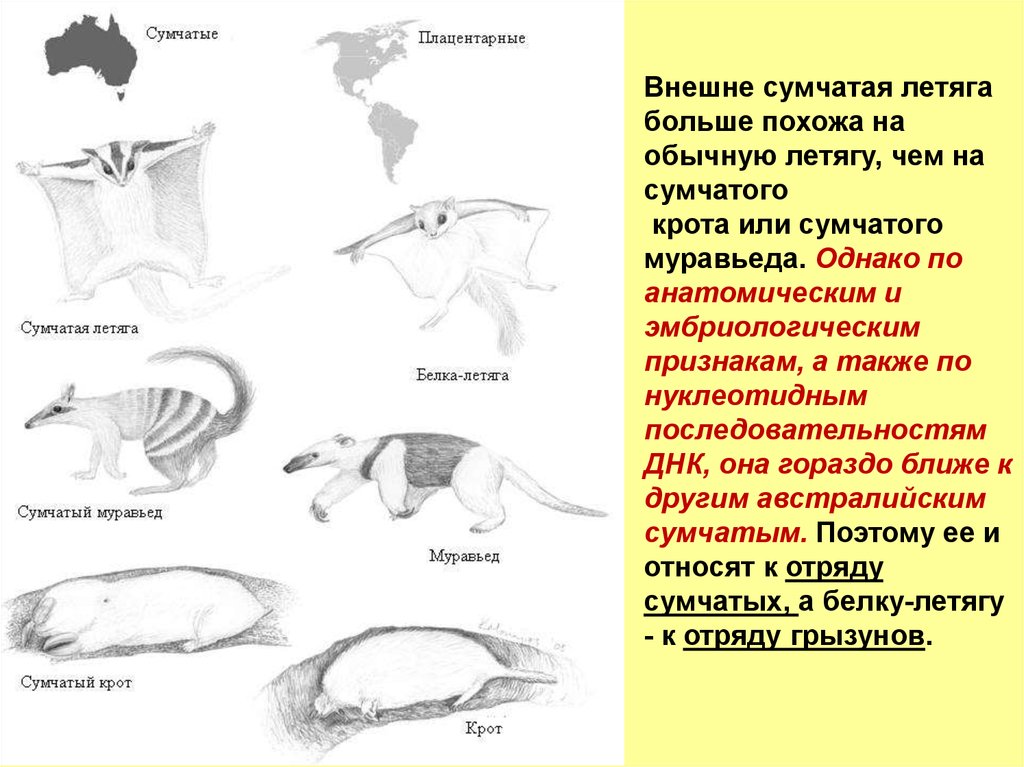
Внешне сумчатая летягабольше похожа на
обычную летягу, чем на
сумчатого
крота или сумчатого
муравьеда. Однако по
анатомическим и
эмбриологическим
признакам, а также по
нуклеотидным
последовательностям
ДНК, она гораздо ближе к
другим австралийским
сумчатым. Поэтому ее и
относят к отряду
сумчатых, а белку-летягу
- к отряду грызунов.
17.
Распространение ископаемых видовсогласуется с эволюционным деревом и
палеогеографическими реконструкциями
Древнейшие окаменелости сумчатых найдены в Китае, их
возраст около 125 млн лет. 40 миллионов лет назад сумчатые
уже были распространены в Южной Америке, но в Австралии,
где они сейчас доминируют, сумчатые появились только
около 30 млн лет назад. Теория эволюции предсказывает, что
австралийские сумчатые произошли от американских.
Согласно теории тектоники плит 30—40 млн лет назад Южная
Америка и Австралия еще оставались частью Гондваны,
крупного континента в южном полушарии, а между ними
находилась будущая Антарктида. На основании двух теорий
исследователи предсказали, что сумчатые мигрировали из
Южной Америки в Австралию через Антарктиду 30—40
миллионов лет назад. Это подтвердилось находками в 1982г.
останков сумчатых возрастом 35-40 млн лет на
антарктических о-вах.
18.
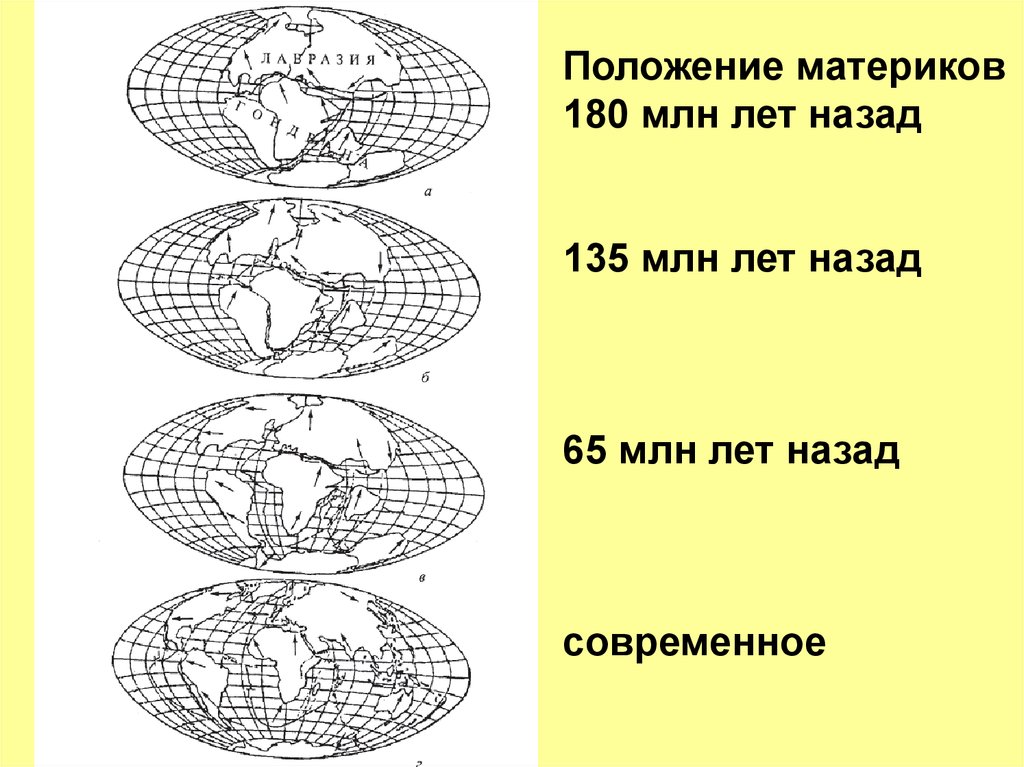
Положение материков не было неизменным. ЮжнаяАмерика с Австралией были до эоцена (30 млн лн),
соединены через Антарктиду, которая в это время еще
не была обмерзшая. Сумчатые проникли из Северной
Америки в Южную Америку, и через Антарктиду — в
Австралию.
Из Старого Света сумчатые так и не проникли в
Австралию по причине длительной разобщенности в
этом районе суши (линия Уоллеса). Поэтому заселение
Австралии через север, через Зондский архипелаг, для
сумчатых был невозможен.
Сходство крайней южной фауны и флоры Америки,
Африки и Австралии с Новой Зеландией – объясняется
тем, что у них был один источник – Антарктида, которая
была 40 млн лет назад вполне приятным местом по
климату.
См. К.Ю. Еськов «Удивительная палеонтология. История Земли и жизни на
ней»
19.
Положение материков180 млн лет назад
135 млн лет назад
65 млн лет назад
современное
20.
21.
22. Распространение остатков вымершего в Перми древнего папортника Glossopteris, и направление движения ледников (стрелки)
Распространениеостатков вымершего в
Перми древнего
папортника
Glossopteris, и
направление
движения ледников
(стрелки) выглядят
совершенно
естественно на
палеогеографической
реконструкции, но не
на современной карте
мира (Coyne, 2009).
23.
Ближайшие живые родственникисовременных людей — гориллы и шимпанзе
— обитают в Африке. Исходя из этого, в 1872
г. Дарвин предположил, что и древних
предков человека следует искать в Африке.
Многие ученые последовали совету Дарвина,
и начиная с 1920-х годов в Африке было
найдено множество промежуточных форм
между человеком и человекообразными
обезьянами, равно как и наиболее древние
предковые формы гоминин (старше 2 млн л).
Если бы ископаемых австралопитеков
обнаружили, например, в Австралии, а не в
Африке, то теорию эволюции пришлось бы
пересматривать.
24.
Основные видыотбора,
факторы и формы
видообразования
(эволюции)
25.
Естественный отбор единственная известнаяпричина адаптаций, но не
единственная причина эволюции.
К числу ее неадаптивных
(случайных) причин
относятся мутации и
генетический дрейф
(в том числе, поток генов).
26. Эффект основателя (поток генов)
27. Смешение популяций (поток генов)
28. Центральный принцип эволюции
«Репликация цифровых носителей информации(таких как ДНК, РНК и белки) неизбежно
подвержена ошибкам (их снижение требует
затрат энергии), что автоматически ведет к
эволюции путем естественного отбора или
дрейфа генов» (если только ошибки не
превышают критического порога 1-10 на геном
за цикл репликации).
Uegene Kunin “The logic of chance: the nature and origin of biological
evolution” 2014 (Евгений Кунин «Логика случая»)
29.
Дрейф генов – случайные изменения частотвстречаемости аллелей генов в популяции
в результате реализации в потомстве
разных гамет, вымирания, миграции, и
других причин.
При «слепом» выборе (20 раз по 1 шарику с возвратом его обратно в банку)
из каждой предыдущей банки и заполнении следующей банки на основе
этого выбора (красный - красным, синий – синим) - в конце останется один
цвет. Этот пример - аналог случайного смещения частоты аллелей.
30.
Эволюция – это и есть изменениечастот аллелей (встречаемости
генетических вариантов) в
популяции. Есть два основных
механизма этого изменения –
генетический дрейф и
естественный отбор.
Остановить их нельзя,
поэтому и эволюция вечна.
31.
Без очищающего отбора накапливаютсяслабовредные мутации просто потому что
мутагенез неостановим, а большинство
ненейтральных мутаций - вредно, а не полезно.
Клонированием с запретом размножения дрейф генов можно остановить, но нельзя
остановить накопление вредных изменений. Тут
нужен очищающий отбор, что стало проблемой
современного человечества, где он ослаблен.
Но все равно - репродуктивный успех у таких
особей с грузом слабовредных мутаций - ниже !
32. Дрейф генов – новый источник случайности в эволюции на стадии закрепления мутаций (они - первый ее источник) . Дрейф генов как
Дрейф генов – новый источник случайности вэволюции на стадии закрепления мутаций (они первый ее источник) .
Дрейф генов как бы отбирает часть ответственности у естественного
отбора действуя, как против, так и вместе с ним. Следствие из теоремы
Фишера: «средняя приспособленность популяции, управляемой ест. отбором,
не может падать (если она не вымирает)».
Адаптивный ландшафт
Сьюэлла Райта (1931) - одного
из основателей
популяционной генетики.
Согласно дарвинизму и
синтетической теории
эволюции
приспособленность
популяции в ходе эволюции
не может падать. Согласно
теории дрейфа генов – иногда
может.
Uegene Kunin “The logic of chance: the nature and origin of biological evolution”
2014
33. Теорема Рональда Фишера (1930):
«Интенсивность отбора (скоростьэволюции путем отбора) пропорциональна
величине генетической вариации
приспособленности эволюционирующей
популяции, которая, в свою очередь,
пропорциональна эффективному размеру
популяции». В больших популяциях (пунктир на рис)
даже «слегка» благоприятные мутации быстрее
закрепляются, а мутации, даже «слегка» вредные, быстрее устраняются.
Р. Фишер, С.Райт и Дж Холдейн основали неодарвинизм, или
синтетическую теорию эволюции. («Генетическая теория
естественного отбора» Фишер, 1930, и др.). Синтез дарвинизма и
34. Сьюэлл Райт:
Но в малой популяции (сплошная линия нарисунке) решающую роль играет дрейф генов,
с помощью которого случайным образом
часто закрепляются нейтральные и даже
слабовредные (но не летальные) мутации.
Очевидно, с помощью генетического дрейфа
эволюционирующая популяция может
избежать однонаправленного подъема по
«адаптивному ландшафту» и может даже
недолго спускаться (временно терять
приспособленность), если эта «долина» узкая.
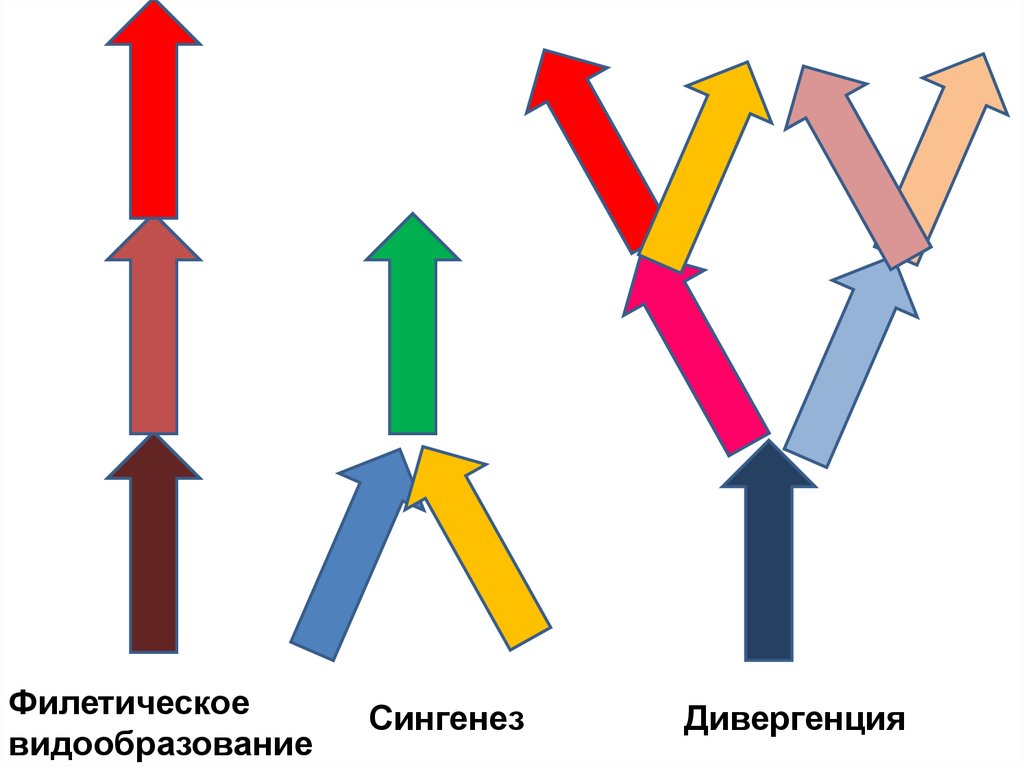
35. Формы эволюции (видообразования)
Филетическоевидообразование
(1 вид-предок –
1 вид-потомок)
Дивергентное видообразование
(кладогенез, идиоадаптация,
адаптивная радиация видов)
1 вид-предок – много видовпотомков)
Разные механизмы изоляции
Стасигенез
(«застой»)
Анагенез
(быстрое
прогрессивное
изменение)
Сингенез,
симбиогенез
Симпатрия
и парапатрия
(В. в пределах
родительского
ареала – эколог-ое,
хромосомное и др)
Разные пути
приспособления
к одним условиям)
Аллопатрия
(В. в результате
географ.
изоляции)
36. Мечехвосты – группа членистоногих (родичи паукообразных), мало изменившаяся с Ордовика (около 400 млн л.н.)
37.
Филетическоевидообразование
Сингенез
Дивергенция
38. Три механизма дивергентного видообразования
39.
Движущий отбор – смещение признака принаправленном изменении среды
-частый вид отбора (пример: промышленный
меланизм насекомых)
Дизруптивный (разрывающий) отбор – отбор
против средней нормы в пользу крайних вариантов
(пример: выгода более ранних и более поздних сроков
размножения у растений, полиморфизм окраски,
комплексы близкородственных видов в озерах) –
следствие устойчивой гетерогенности среды.
Как его вариант: половой отбор.
Стабилизирующий (очищающий) отбор – отбор на
среднее значение признака (очень много примеров).
Действует после двух первых видов отбора при
постоянстве среды.
Балансирующий отбор.
40. Формы естественного отбора
41. Примеры дивергенции (формы изоляции:симпатрия и аллопатрия)
42. Реконструкция адаптивной радиации вьюрков со смещением признаков
43. Факторы абиотической среды– как факторы естественного (движущего) отбора
44. Потомки следуют в направлении родительских изменений
45. Движущий отбор
46. Стабилизирующий отбор
47. Искусственный отбор как модель дизруптивного естественного отбора
48. Редкие слабовредные или слабоположительные рецессивные аллели (мутации) могут очень долго «вымываться» из популяции, т.к. чем
они реже, темслабее на них действует
отбор. Далее они либо
исчезают в результате
дрейфа генов, либо
подхватываются
отбором, если
становятся выгодными
или сохраняются в
изолированной группе.
49. Аномалия, связанная с наличием 6-го пальца у секты амишей в США Амиши, основанные группой из 200 человек (сейчас их более 250
000) – важный модельныйобъект эволюционногенетических исследований.
Одна из самых быстрорастущих
популяций человека.
50.
Основные этапы эволюциисамой теории эволюции:
от
классического дарвинизма
(первый синтез – открытие естественного отбора)
к
синтетической теории эволюции
(синтез дарвинизма с генетикой;
автор термина: Джулиан Хаксли,1942)
И
современному синтезу («эво-дево»)
51. Синтетическая теория эволюции (1920-е – 1980-е) – (второй синтез) Произошел синтез классического дарвинизма, генетики (Ф.
Добржанский)палеонтологии (Дж.Симпсон) и экологии (Э. Майр).
1. Материал эволюции – мутации (в осн. точечные).
2. Основной движущий фактор Э. – естественный отбор случайных
мелких мутаций.
3. Наименьшая эволюционирующая единица - популяция, а не особь.
4. Эволюция носит дивергентный характер (один вид ≠ два предка).
5. Э. носит постепенный (градуалистический) характер.
6. Вид состоит из подвидов и популяций.
7. Поток генов возможен лишь внутри вида (репродуктивная
обособленность вида).
8. Репродуктивная обособленность – критерий вида только для форм с
половым размножением. За рамками СТЭ остаются прокариоты и
низшие эукариоты.
9. Реальный таксон (не сборный) имеет монофилетическое
происхождение
10. Эволюция непредсказуема и имеет ненаправленный характер.
52.
Основные постулаты синтетической теорииэволюции (1920-е – 1980-е) – (второй синтез)
1. Центр внимания СТЭ – приспособленность.
Все признаки имеют приспособительное значение
(сейчас пересмотрено).
2. В природе нет целеполагающего начала, а у Э. нет
цели.
3. Потребность в адаптации и сама адаптация
возникают синхронно.
4. Средством Э. является «отбор редких полезных
изменений и элиминация вредных»
5. Изменчивость непрерывна, неограниченна и
случайна (пересмотрено).
6. У микро- и макроэволюции одни закономерности
(пересмотрено)
53. Предпосылки современного (третьего) синтеза (1980 – н.в.)
• Необратимость эволюции в силувероятностных причин (хотя обратимость
отдельных признаков возможна, известны
прямые и обратные мутации)
• Чем больше специализация, тем более
ограничен выбор возможных путей
дальнейшего исторического развития
54. Неравномерность темпов эволюции
• Относительная независимость темповпреобразования разных структур организма
• Степень адаптивности одного итого же признака
или органа может меняться в зависимости от
степени функциональной нагрузки, которую от
несет
На разных этапах исторического развития вида
один и тот же признак может менять свою
селективную ценность от позитивной до
негативной, проходя через точку нулевой
приспособленности
55.
Представление о запретных ходах эволюции, иэволюция на основе закономерностей, а не
случайностей (истоки: «гомологические ряды
растений» Н.И. Вавилова, «номогенез» Л.С. Берга,
А.А.Любищев, С.В.Мейен) .Существуют запретные
состояния, к которым не могут привести никакие
мутации .
Примеры. У млекопитающих цвет волос может меняться от черного через
рыжий и желтый к бесцветному. Запрет на синий и зеленый – в цепи
реакций образования пигмента нет возможности возникновения зеленого
цвета (ленивцы и симбиотические водоросли на их шерсти)
Эволюционные ограничения могут распространяться на таксоны
высокого ранга: представители класса амфибий живут в диапазоне от
тропиков до тайги, от пустынь до поземных вод. Но ни один вид никогда не
жил в морях. Это невозможно в силу их кожно-легочного дыхания и типа
водно-солевого обмена.
56. О чем сейчас не спорят эволюционисты
• 1. Жизнь на земле развивается около 3.8 – 4млрд. лет (от прокариот к эукариотам).
• 2. В ходе эволюции идут приспособительные
процессы, но не все признаки могут быть
адаптивными. Эволюционный процесс идет
непрерывно, но неясно насколько равномерно.
• 3. Процесс выработки адаптаций идет при
участии естественного отбора.
57. О чем сейчас не спорят эволюционисты
• 4. Элементарной единицей эволюцииявляется популяция особей.
• 5. Материалом для эволюции (через отбор
или дрейф генов, или за счет иных
стохастических процессов) служат мутации
и их комбинация (макро- или микромутации –
это предмет спора).
• 6. Важнейшее событие в эволюции –
видообразование (но пока неясно, как идет
этот процесс – постепенно или
скачкообразно).
58. Современное состояние проблемы (третий синтез)
1. В силе постулат о популяции как о единицеэволюции.
2. Естественный отбор – движущая сила
эволюции, но не единственная. Дарвиновская и
«недарвиновская» эволюция взаимно не
противоречивы.
3. Эволюция не всегда носит дивергентный
характер, может иметь место симгенез,
симбиогенез, и горизонтальный перенос
генетического материала.
4. Возможна парафилия наряду с монофилией.
59. Современное состояние проблемы
5. Эволюция может идти постепенно и скачкаминапр. путем полиплоидии, за счет внезапных
хромосомных перестроек, через дупликации генов
(автор: Сусуму Оно), и прерывистое равновесие
(автор: Стивен Гулд)
6. Вид является целостной, генетически замкнутой
системой, но есть дополнение (описаны случаи
просачивания потока генов через неабсолютные
барьеры изолирующих механизмов).
7. Макроэволюция может идти как через
микроэволюцию, так и своими путями.
60. Современное состояние проблемы
8. Представление о политипическом виде остается в силе (видсостоит из подвидов и популяций).
9. Репродуктивный критерий для определения вида сегодня
недостаточен, пока не предложено общего определения вида
для бесполых форм и форм с половым размножением.
10. Случайный характер мутационной изменчивости не
противоречит возможности существования
«канализованности» путей эволюции, возникшей в ходе
истории развития данного вида. Изменчивость не безгранична.
11. Несмотря на множество ее факторов и случайного в целом
характера, эволюция отчасти может быть предсказуема с
учетом информации о ее конкретных запретах, знания
прошлой истории, генотипического окружения и известного
направления влияния среды.
61. Две модели эволюции: градуализм и прерывистое равновесие (С.Гулд, 1972)
Эволюция идет не толькопутем отбора постепенных
изменений, но и за счет
фиксации внезапных, резких
перестроек морфогенеза
(мутации регуляторных генов
в ходе эмбриогенеза).
Также - хромосомное
видообразование
(робертсоновские веера). У
домовых мышей (это один вид)
на территории Альп и
Аппенинского полуострова
число хромосом варьирует от
40 до 22. Между вариантами
наблюдается репродуктивная
изоляция разного уровня. Также
у человека - 2-ая хромосома.
62.
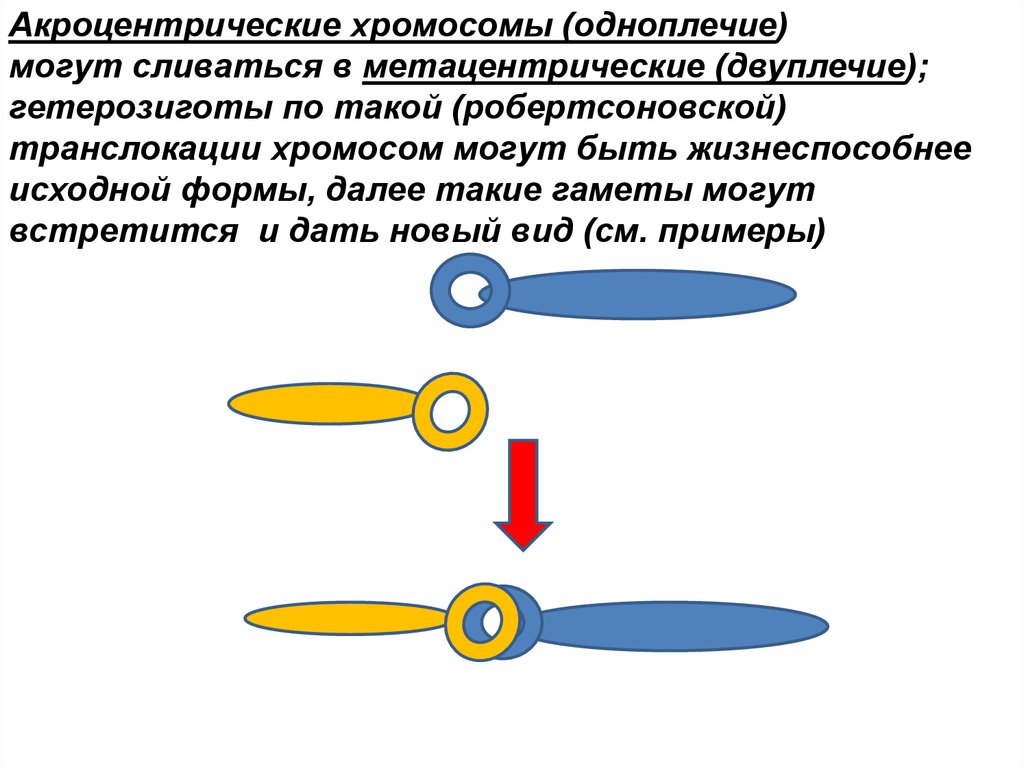
Акроцентрические хромосомы (одноплечие)могут сливаться в метацентрические (двуплечие);
гетерозиготы по такой (робертсоновской)
транслокации хромосом могут быть жизнеспособнее
исходной формы, далее такие гаметы могут
встретится и дать новый вид (см. примеры)
63.
Не исключено, что отсутствиепереходных форм, которое обычно
со времен Ч. Дарвина списывают на
неполноту палеонтологической
летописи – на самом деле
отражение специфики
эволюционного процесса –
скачкообразности изменений
(наличия «прерывистого равновесия» - смены
быстрого кладогенеза периодами
длительного стасигенеза).