

































 biology
biologySimilar presentations:










Элементарные факторы эволюции
1. Белорусский государственный педагогический университет имени Максима Танка факультет естествознания кафедра общей биологии
Электронное учебное пособие для самостоятельнойработы студентов по дисциплине
«Эволюционное учение»
Автор:
Политыко П. Ю.
2.
•Факторы эволюции• Схема
• Мутационный процесс
• Изоляция
• Дрейф генов
• Поток генов
• Популяционные волны
•Естественный отбор
• Формы естественного отбора
Движущий отбор
Стабилизирующий отбор
Дизруптивный отбор
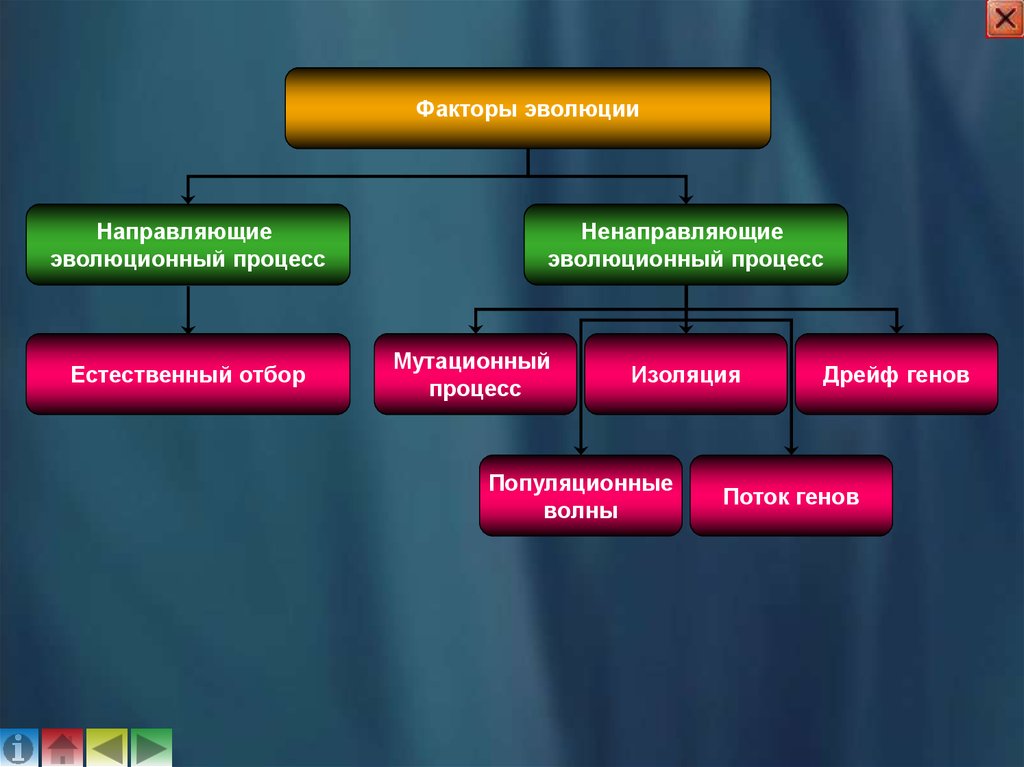
3. Факторы эволюции
Факторы эволюции - движущая сила, вызывающая и закрепляющая изменения в популяциях какэлементарных единицах эволюции. Современные представления о факторах эволюции – результат
развития дарвинизма, генетики и экологии. Чарльз Дарвин в своем классическом труде
«Происхождение видов» решил вопрос о главных движущих силах (факторах) эволюционного
процесса. Он выделил следующие факторы: наследственность, изменчивость и естественный
отбор. Кроме того, Ч. Дарвин указал на важную роль ограничения свободного скрещивания особей
вследствие их изоляции друг от друга, возникшей в процессе эволюционного расхождения видов.
В современном представлении важнейшими факторами эволюционного процесса являются:
мутационный процесс, изоляция, дрейф генов, популяционные волны, поток генов, естественный
отбор.
4.
Факторы эволюцииНаправляющие
эволюционный процесс
Естественный отбор
Ненаправляющие
эволюционный процесс
Мутационный
процесс
Изоляция
Популяционные
волны
Дрейф генов
Поток генов
5. Мутационный процесс
Один из главных факторов эволюции является мутационный процесс. Мутации были открыты вначале XX в. голландским ботаником и генетиком Де Фризом (1848 -1935).
Мутации – стойкие изменения генетического материала (числа хромосом, числа и порядка
расположения генов в хромосоме, структуры генов). Мутационный процесс играет роль главного
поставщика наследственных изменений. Именно он является первичным материалом всех
эволюционных преобразований. Чаще всего мутации рецессивны, находятся в гетерозиготном
состоянии и не проявляются фенотипически. Именно эти мутации и служат генетической основой
эволюции. При скрещивании гетерозиготных особей эти мутации у потомков могут переходить в
гомозиготное состояние. Комбинативная изменчивость ведет к появлению бесконечно большого
разнообразия генотипов и фенотипов.
Отбор из поколения в поколение сохраняет особей с полезными мутациями. Полезные мутации
сохраняются естественным отбором, вредные - накапливаются в популяции в скрытом виде,
создавая резерв изменчивости. Это приводит к изменению генофонда популяции.
Мутации бывают: генные, хромосомные и геномные.
6. Изоляция
Еще Ч. Дарвин указывал, что изоляция – очень важный эволюционный фактор, благодаря которомумежду особями разных популяций не происходит скрещивания, а значит, и обмена генетической
информацией.
В каждой популяции благодаря естественному отбору накапливаются определенные полезные
мутации. Через несколько поколений изолированные популяции, обитающие в разных условиях,
будут различаться по ряду признаков.
В зависимости от природы изолирующих барьеров выделяют два способа изоляции:
пространственный (географический) и биологический. Биологический способ изоляции включает в
себя следующие формы изоляции: экологическую, морфофункциональную, этологическую,
генетическую.
Изоляция закрепляет межпопуляционные различия в частотах встречаемости различных генотипов
и способствует созданию группировок с независимыми генофондами, которые могут стать
самостоятельными видами.
Таким образом, различные типы изоляции, с одной стороны, создают предпосылки к расхождению
популяций и к последующему видообразованию, а с другой – способствуют сохранению
генетической структуры вида.
7. Дрейф генов
Явление генетического дрейфа впервые обнаружили известные ученые-генетики Н. П. Дубинин иД. Д. Ромашов, а также зарубежные ученые С. Райт и Р. Фишер. С. Райт экспериментально
доказал, что в маленьких популяциях частота мутантного аллеля меняется быстро и случайным
образом. Его опыт был прост: в пробирки с кормом он посадил по две самки и два самца мух
дрозофил, гетерозиготных по гену А (их генотип можно записать Аа). В этих искусственно
созданных популяциях концентрация нормального (А) и мутантного (а) аллелей составила 50 %.
Спустя несколько поколений оказалось, что в некоторых популяциях все особи стали
гомозиготными по мутантному аллелю (а), в других популяциях он был вовсе утрачен, и, наконец,
часть популяций содержала как нормальный, так и мутантный аллель. Важно подчеркнуть, что
несмотря на снижение жизнеспособности мутантных особей и, следовательно, вопреки
естественному отбору в некоторых популяциях мутантный аллель полностью вытеснил
нормальный. Это и есть результат случайного процесса – дрейфа генов.
Дрейф генов обычно снижает генетическую изменчивость в популяции, главным образом в
результате утраты редко встречающихся аллелей. Этот механизм эволюционных изменений
особенно эффективен в небольших популяциях.
8. Поток генов
Если изоляция между соседними популяциями неполная, то между ними возникает обмен генами врезультате свободного скрещивания их особей. Этот процесс получил название генного потока.
Поток генов - это изменение частот генов в генофонде популяции под влиянием эмиграции и
иммиграции. Большую роль в осуществлении потока генов играют миграции, кочевки, перелеты,
перенос пыльцы и семян ветром, насекомыми.
Популяция может приобрести новый аллель не в результате мутации, а в результате иммиграции вселения в данную популяцию из соседней носителя нового гена. Генный поток является важным
источником изменчивости. Часть особей -мигрантов одной популяции проникает в другую, и их
гены включаются в генофонд этой популяции. При скрещивании особей разных популяций
генотипы потомства будут отличаться от генотипов обоих родителей. В данном случае происходит
перекомбинация генов на межпопуляционном уровне, т.е. генный поток также является
поставщиком материала для естественного отбора. Именно благодаря потоку генов на обширных
территориях наблюдается фенотипическая однородность особей. Значение этого процесса
отметил еще Дарвин: "Скрещивание играет важную роль в природе, так как поддерживает
однообразие и постоянство признаков у особей одного и того же вида".
Поток генов имеет важнейшее биологическое следствие – объединение всех популяций в единую
видовую систему.
9. Популяционные волны
Популяционные волны, или "волны жизни" - это вспышки численности, периодические илинепериодические значительные изменения числа особей в популяции. С. С. Четвериков назвал
популяционные волны и 1905 г. "волнами жизни". Например, у клестов максимум численности
наблюдается раз в три года. Период колебаний численности у мышевидных грызунов - около 10
лет, у белок - 8-11 лет, бабочки-белянки - 10-12, североамериканского зайца-беляка и рыси на
севере Канады - 9-10, саранчи - около 11 лет. Масштабы колебаний численности у популяций
разных видов обычно разные. Так, численность майского жука на пике волны увеличивается в
миллион раз, сибирского шелкопряда - в 12 млн. раз, зайцев в Канаде - в 10 раз. "Волны жизни"
исчезают за счет появления большого количества хищников, паразитов, возникновения эпизоотии,
изменения абиотических условий… узнать больше>>
Эволюционное значение популяционных волн в
том, что они:
изменяют частоты аллелей (малочисленные
аллели на пике волны могут проявиться
фенотипически, а на спаде - исчезнугь из
генофонда);
на пике волны изолированные популяции
сливаются, растет миграция и панмиксия, растет
гетерогенность генофонда;
популяционные волны изменяют интенсивность
природного отбора и его направление.
10. Естественный отбор – направляющий фактор эволюции
Естественный отбор - основная движущая сила эволюции, и любой вид живых организмов, когдалибо живший на Земле, так или иначе формировался под действием этого фактора. Учение о
естественном отборе – центральная часть эволюционной теории Дарвина. Дарвин пришел к
выводу о существовании естественного отбора в природе на основании собственных наблюдений
за живой природой во время кругосветного путешествия и изучения искусственного отбора.
Понятие естественного отбора относится к фундаментальным понятиям не только эволюционной
теории, но и всей биологии. В современном понимании естественный отбор — это избирательное
(дифференциальное) воспроизведение генотипов, или дифференциальное размножение. Более
приспособленные к данным условиям среды особи оставляют больше потомков, чем менее
приспособленные.
Дифференциальное размножение представляет собой конечный результат многочисленных
процессов: выживания гамет, успеха в оплодотворении, выживаемости зигот, эмбрионов,
рождения, выживаемости в молодом возрасте и в период половой зрелости, стремления к
спариванию, успешности спаривания, плодовитости. Различия в этих процессах есть следствие
различий в признаках и свойствах, различия в генетической программе.
11.
Естественный отбор происходит автоматически. Все живые организмы из поколения впоколение проходят суровую проверку по всем мельчайшим деталям их строения,
функционирования всех их систем в разнообразных условиях. Только те, кто выдержал эту
проверку, оказываются отобранными и дают начало следующему поколению. Дарвин писал:
«Естественный отбор ежедневно и ежечасно расследует по всему свету мельчайшие вариации,
отбрасывая дурные, сохраняя и слагая хорошие, работая неслышно и незаметно, где бы и когда
бы, ни представился к тому случай, над усовершенствованием каждого органического существа по
отношению к условиям его жизни, органическим и неорганическим. Мы ничего не замечаем в этих
медленных переменах в развитии, пока рука времени не отметит истекших веков».
Эффективность естественного отбора зависит от его интенсивности и запаса
наследственной изменчивости, накопленного в популяции. Интенсивность отбора определяется
тем, какая доля особей доживает до половой зрелости и участвует в размножении. Чем меньше эта
доля, тем больше интенсивность отбора. Если, например, из 10 000 особей в каждом поколении
выживают и размножаются только 100 самых крупных, то средний размер особей в этой популяции
растет гораздо быстрее, чем в случае более мягкого отбора, когда, например, половина всех
особей в популяции участвует в размножении.
Интенсивность отбора варьирует в широких пределах в зависимости от вида и экологической
ситуации. Однако она никогда не бывает нулевой. Даже в человеческих популяциях, которые
благодаря достижениям современной цивилизации в сильной степени защищены от действия
отбора, его интенсивность весьма значительна. Около 30% всех беременностей заканчиваются
выкидышами на самых ранних стадиях; 5% новорожденных гибнет при рождении, еще около 3% не
доживает до зрелости; 20% людей, доживших до зрелости, не вступают в брак, и 10% вступивших в
брак не имеют детей. В природных популяциях микроорганизмов, растений и животных
интенсивность естественного отбора гораздо выше.
12.
Однако даже самый интенсивный отбор будет неэффективен, если изменчивость особей впопуляции незначительна или она носит ненаследственный характер. Если все особи в популяции
совершенно одинаковы, то даже в условиях жесткой борьбы за существование все они имеют
равные шансы уцелеть или погибнуть. Жизнь и размножение каждой такой особи зависит от
случая. Из поколения в поколение генетический состав этой популяции будет оставаться
неизменным.
Изменения не будут накапливаться и в том случае, если изменчивость особей в популяции
родителей обусловлена не генетической изменчивостью, а разнообразием средовых условий.
Тогда среднее значение признака и потомков особей с крайне высоким и крайне низким
проявлением признака будет одинаковым. Такую картину мы всегда наблюдаем в так называемых
чистых линиях (гомозиготных по всем генам) растений и животных и в клонах бесполых
организмов. Поэтому отбор в чистых линиях и клонах оказывается неэффективным. Сколько бы
поколений мы ни отбирали представителей чистых линий с наибольшим или наименьшим
проявлением признака, популяционная средняя в последующих поколениях остается неизменной.
В генетически гетерогенных популяциях отбор ведет к постепенному смещению средней из
поколения в поколение.
Важнейшим свойством естественного отбора является его способность накапливать и
совмещать полезные аллели. Отбор действует в каждом поколении, и в каждом поколении он
начинается не с нуля, не с «чистого листа», а того состояния популяции, в которое она была
приведена в результате предыдущего цикла отбора. Если в какой-то популяции из года в год
высокие растения получают преимущество в выживании и размножении, то поколение за
поколением в этой популяции растет частота высоких особей, и увеличивается средних размер
особей. Все новые и новые аллели возникают в популяции за счет мутационного процесса. Каждая
мутация случайна, но ее носители регулярно отбираются, и размножаются в популяции, если она
хоть чуть-чуть повышает приспособленность ее носителей. Если критерием приспособленности в
данной популяции является высота растения, то отбор подхватывает все аллели, которые тем или
иным способом работают на этом признак – увеличивают скорость роста, эффективность
использования солнечного света, воды и других ресурсов.
13.
Формы естественного отбораФОРМА
ОТБОРА
Движущий
Стабилизирующий
Дизруптивный
ДЕЙСТВИЕ
НАПРАВЛЕННОСТЬ
РЕЗУЛЬТАТ
ПРИМЕРЫ
При изменении
условий
существования
организмов
В пользу особей,
имеющих отклонения
от средней нормы
Возникает новая
средняя форма,
более
соответствующая
изменившимся
условиям
Возникновение у насекомых
устойчивости к ядохимикатам;
распространение
темноокрашенных бабочек
березовой пяденицы в условиях
потемнения коры берез от
постоянного задымления
В неизменных,
постоянных
условиях
существования
Против особей с
возникающими
крайними
отклонениями от
средней нормы
выраженности
признака
Сохранение и
укрепление
средней нормы
проявления
признака
Сохранение у
насекомоопыляемых растений
размеров и формы цветка
(цветки должны
соответствовать форме и
величине тела насекомого
опылителя, строению его
хоботка)
В изменяющихся
условиях жизни
В пользу организмов,
имеющих крайние
отклонения от
средней
выраженности
признака
Образование
новых средних
норм вместо
прежней,
переставшей
соответствовать
условиям жизни
При частых сильных ветрах на
океанических островах
сохраняются насекомые с
хорошо развитыми или с
рудиментарными крыльями
14.
Движущий отборДвижущий отбор — форма естественного отбора, когда условия
среды способствуют определённому направлению изменения
какого-либо признака или группы признаков. При этом иные
возможности изменения признака подвергаются
отрицательному отбору. В результате в популяции от поколения
к поколению происходит сдвиг средней величины признака в
определённом направлении. При этом давление движущего
отбора должно отвечать приспособительным возможностям
популяции и скорости мутационных изменений (в ином случае
давление среды может привести к вымиранию).
Современным случаем движущего отбора является
«индустриальный меланизм английских бабочек».
«Индустриальный меланизм» представляет собой резкое
повышение доли меланистических (имеющих тёмную окраску)
особей в тех популяциях бабочек, которые обитают в
промышленных районах. Из-за промышленного воздействия
стволы деревьев значительно потемнели, а также погибли
светлые лишайники, из-за чего светлые бабочки стали лучше
видны для птиц, а тёмные — хуже. В XX веке в ряде районов
доля тёмноокрашенных бабочек достигла 95 %, в то время как
впервые тёмная бабочка (Morfa carbonaria) была отловлена в
1848 году.
Индустриальный меланизм. Темные формы
бабочек незаметны на темных стволах , а
светлые – на светлых.
Светлые и темноокрашенные
бабочки березовой пяденицы
15.
Стабилизирующий отборСтабилизирующий отбор — форма естественного отбора, при котором действие направлено
против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней
выраженностью признака.
Описано множество примеров действия стабилизующего отбора в природе. Например, на первый
взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить
особи с максимальной плодовитостью. Однако наблюдения над природными популяциями птиц
и млекопитающих показывают, что это не так. Чем больше птенцов или детенышей в гнезде,
тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее
приспособленными оказываются особи со средней плодовитостью.
Отбор в пользу средних значений был обнаружен по множеству признаков. У млекопитающих
новорожденные с очень низким и очень высоким весом чаше погибают при рождении или в
первые недели жизни, чем новорожденные со средним весом. Учет размера крыльев у птиц,
погибших после бури, показал, что большинство из них имели слишком маленькие или слишком
большие крылья. И в этом случае наиболее приспособленными оказались средние особи.
16.
Дизруптивный отборПри стабилизирующем отборе преимуществом обладают особи со средним проявлением
признаков, при движущем – одна из крайних форм. Теоретически мыслима еще одна форма отбора
– дизруптивный или разрывающий отбор, когда преимущество приобретают обе крайние формы.
Действием дизруптивного отбора объясняют
образование сезонных рас у некоторых сорных
растений. Было показано, что сроки цветения и
созревания семян у одного из видов таких
растений - погремка лугового- растянуты почти
на все лето, причем большая часть растений
цветет и плодоносит в середине лета. Однако
на сенокосных лугах получают преимущества те
растения, которые успевают отцвести и дать
семена до покоса, и те, которые дают семена в
конце лета, после покоса. В результате
образуются две расы погремка – ранне- и
позднецветущая.
В определенных ситуациях дизруптивный отбор по
признакам, связанным особенностями экологии
(временем размножения, предпочтением разных
видов корма, разных местообитаний) может
приводить к образованию экологически
обособленных рас внутри вида и затем к
видообразованию.
Формы естественного отбора
17.
Эволюционная роль естественного отбораЕстественный отбор определяет становление адаптации.
Естественный отбор контролирует соответствие строения и функции организма среде.
Естественный отбор снижает концентрацию вредных мутаций в генофонде популяции.
Естественный отбор увеличивает концентрацию мутаций, имеющих существенное
приспособительное значение.
Естественный отбор способствует формированию полиморфизма популяции.
Естественный отбор ведет к дивергенции - увеличению разнообразия организмов.
Естественный отбор ведет к биологическому прогрессу.
примеры естественного отбора>>
18.
Наиболее частый тип мутаций – генные (или точечные). Затрагивают изменение одного гена, чащерецессивного, т.е. фенотипически не проявляется. Они играют очень важную роль в эволюционном
процессе. Мутации отдельных генов происходят редко. Мутация гена возникает в среднем в одной
из 100 000 гамет. Но так как количество генов в организме, например, млекопитающих, составляет
около 100 000, то практически каждая особь несет вновь возникшую мутацию.
Генные мутации имеют первостепенное значение для эволюции.
Во – первых, они универсальны, присущи всем живым организмам.
Во – вторых, частота генных мутаций выше, чем частота других типов мутаций.
В – третьих, и это главное, генные мутации вызывают преимущественно мелкие изменения
фенотипа, накопление которых в процессе отбора составляет основной путь микроэволюции.
19.
Хромосомные мутации также играют важную эволюционнуюроль. Они изменяют структуру хромосом. Многие из
хромосомных мутаций летальны. Рецессивные
хромосомные мутации сохраняются в гетерозиготах и
пополняют резерв внутривидовой наследственной
изменчивости.
Благодаря удвоениям генов в процессе эволюции
накапливается генетический материал. Нарастание
сложности организации живого в ходе исторического
развития в значительной степени опиралось на увеличение
количества генетического материала. Достаточно сказать,
что количество ДНК в клетке у высших позвоночных
примерно в 1000 раз больше, чем у бактерий.
Филогенетическое древо глобиновых
генов. Ген глобина в ходе эволюции
несколько раз дуплицировался (отмечено
стрелками). Его добавочные копии затем
приобретали новые свойства и функции.
Из гена бета-глобина общего предка
возникли гены гамма-, дельта-, эпсилонглобинов – белков, которые выполняют
иные функции, чем бета-глобин.
20.
Геномные мутации заключаются в изменении нормального числа хромосом, строго определенногодля каждого вида. Одним из распространенных типов геномных мутаций является полиплоидия,
имеющая большое значение для эволюции растений, в особенности у высших покрытосеменных.
Полиплоидные виды растений часто занимают арктические и альпийские зоны. Считают, что это
связано с их повышенной устойчивостью к неблагоприятным факторам среды.
Слева изображено обычное растение энотеры, справа —
гигантская энотера, обнаруженная Гуго де Фризом в конце 90-х
годов прошлого века. Де Фриз не знал, чему обязан этот экземпляр
энотеры своим большим ростом, множеством стеблей, крупными
цветками. Лишь через 30 лет генетики нашли, что это связано с
увеличением набора хромосом в клетках гигантской энотеры.
Рисунок гигантской энотеры был сделан самим де Фризом.
21.
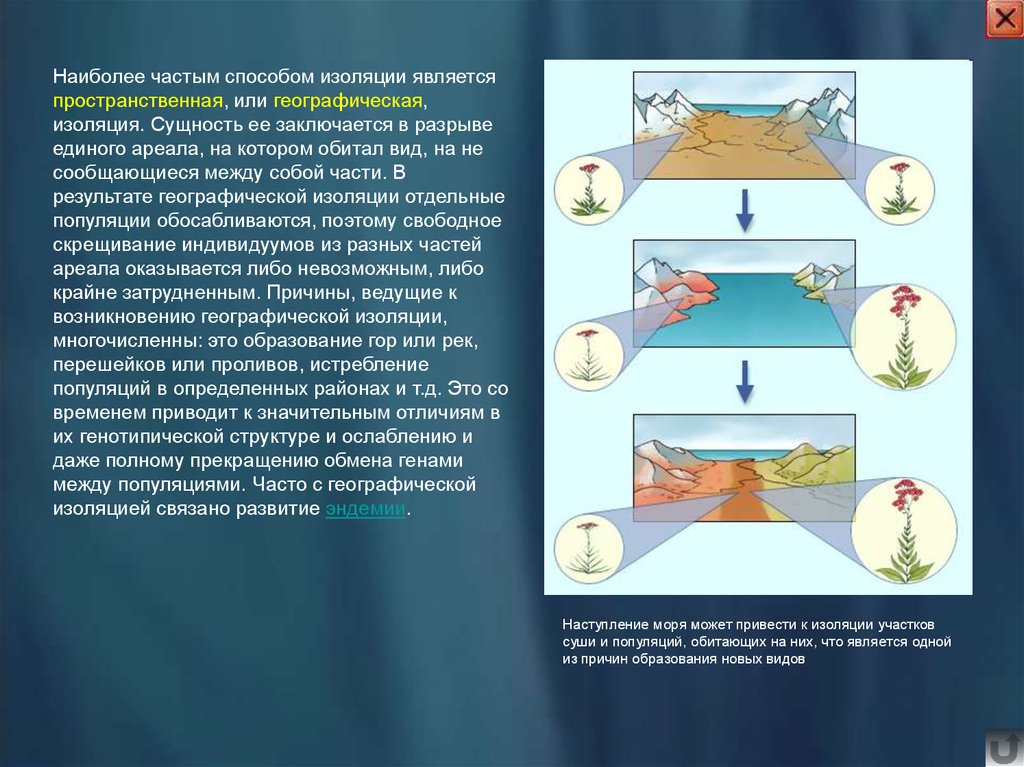
Наиболее частым способом изоляции являетсяпространственная, или географическая,
изоляция. Сущность ее заключается в разрыве
единого ареала, на котором обитал вид, на не
сообщающиеся между собой части. В
результате географической изоляции отдельные
популяции обосабливаются, поэтому свободное
скрещивание индивидуумов из разных частей
ареала оказывается либо невозможным, либо
крайне затрудненным. Причины, ведущие к
возникновению географической изоляции,
многочисленны: это образование гор или рек,
перешейков или проливов, истребление
популяций в определенных районах и т.д. Это со
временем приводит к значительным отличиям в
их генотипической структуре и ослаблению и
даже полному прекращению обмена генами
между популяциями. Часто с географической
изоляцией связано развитие эндемии.
Наступление моря может привести к изоляции участков
суши и популяций, обитающих на них, что является одной
из причин образования новых видов
22.
Биологический способ изоляции включает такие механизмы, которые либо вообще не допускаютскрещивания, либо препятствуют воспроизведению нормального потомства. По своей природе эти
механизмы очень различны, но сходны по биологическому значению как у растений, так и у
животных (за исключением этологической изоляции). Биологический способ включает несколько
форм изоляции: экологическую, морфофункциональную, этологическую, генетическую.
23.
Экологическая форма изоляции определяется условиями существования видов, связанными сместами и сроками размножения. Поэтому экологическую форму изоляции подразделяют на
биотопическую и сезонную.
Биотопическая изоляция возникает в результате расхождения близкородственных видов по разным
местообитаниям. Так, в Молдавии у желтогорлой лесной мыши образовались лесные и степные
популяции. Особи лесных популяций более крупные, питаются семенами древесных пород, а особи
степных популяций - семенами злаков.
Сезонная изоляция определяется различиями во времени полового созревания. Известны пять рас
севанской форели, размножение которых происходит в разные месяцы. Сдвиг в сроках
размножения может быть очень небольшим, но весьма эффективным изолирующим механизмом,
т.к. период оплодотворения у видов ограничен во времени.
Желтогорлая мышь —
Apodemus flavicollis
24.
Морфофункциональная форма изоляции обусловлена особенностями строения ифункционирования органов размножения. В ходе эволюции выработались два основных способа
такой изоляции: морфологический и физиологический.
Морфологическая изоляция заключается в несоответствии строения органов размножения,
которое препятствует оплодотворению. Морфологическая половая несовместимость не допускает,
например, копуляцию между двумя видами наземных улиток (Сepaea nemoralis и Cepaea hortensis).
Физиологическая изоляция связана с нарушениями процессов размножения (гибель гамет или их
неспособность к оплодотворению при попадании к особям других видов ). Так, если на рыльце
пестика попадает пыльца чужого вида, она не прорастает или недоразвиваются пыльцевые трубки
и мужские гаметы не достигают яйцеклетки.
25.
Этологическая форма изоляции определяется особенностями поведения, допускающимиспаривание только с особями своего вида. Она свойственна только животным и основана на
генетически запрограммированных сложных ритуалах опознания брачного партнера, например,
знаменитый тетеревиный ток. Сигналами видового распознавания являются различные
раздражители (зрительные, звуковые, обонятельные). Так, зарождающаяся этологическая
изоляция установлена у двух рас тритона, обитающего на разных островах Японии. Хотя их
гибриды вполне жизнеспособны, в случаях, когда спаривание все же происходит, самка и самец
предпочитают партнеров своей расы.
Тетеревиный ток
26.
Генетическая форма изоляции заключается в несовместимости половых продуктов самца и самки,например, по числу и строению хромосом. Так, 15-хромосомная раса австралийского кузнечика
Moraba scuria в результате транслокаций превратилась в 17-хромосомную расу, которая
распространилась в небольшой изолированный популяции. Генетическая изоляция проявляется на
разных стадиях онтогенеза, начиная с гибели зигот и эмбрионов до образования потомства с
пониженной жизнеспособностью или стерильного.
27.
Эндемики – виды, роды, семейства растений или животных, распространение которых ограниченонебольшой географической областью, т.е. это специфическая для данной области флора или
фауна. Например, наиболее раннее отделение Австралии от южного материка Гондваны (более
120 млн. лет) привело к самостоятельному развитию ряда животных. Не испытывая давления со
стороны хищников, которые отсутствуют в Австралии, здесь сохранились однопроходные
млекопитающие – первозвери: утконос и ехидна; сумчатые: кенгуру, коала.
Коала
28.
Ч. Дарвин описывает явление, которое вполне можно объяснить дрейфом генов. "Кролики,одичавшие на острове Порто-Санто, близ о. Мадейры, заслуживают более полного описания.
В 1418 или 1419 г. у Гонзалеса Зарко на корабле случайно оказалась беременная крольчиха,
которая родила во время путешествия. Все детеныши были выпущены на остров. Кролики
уменьшились почти на три дюйма в длину и почти вдвое в весе тела. По окраске кролик с
Порто-Санто значительно отличается от обыкновенного. Они необычайно дики и проворны.
По своим привычкам они более ночные животные. Производят от 4 до 6 детенышей в помете.
Не удалось спарить с самками других пород". Примером воздействия дрейфа генов могут быть
кошки о. Вознесения. Более 100 лет назад на острове появились крысы. Они расплодились в
таком количестве, что английский комендант решил избавиться от них с помощью кошек. По
его просьбе привезли кошек. Но они сбежали в отдаленные уголки острова и стали уничтожать
не крыс, а домашнюю птицу и диких цесарок.
Другой комендант, чтобы избавиться от кошек, завез собак. Собаки не прижились — они
ранили лапы об острые кромки шлака. Кошки со временем стали свирепыми и кровожадными. За
столетие они отрастили себе почти собачьи клыки и стали сторожить дома островитян,
ходить по пятам за хозяином и бросаться на посторонних".
Изоляты людей (кастовые, религиозные или географические) также подвержены дрейфу. Инки —
замкнутая каста правителей в Южной Америке - имели группу крови, не встречавшуюся ни у
древних, ни у современных американцев. Гренландские норманны — прекрасно развитые люди —
за 200 лет изоляции от Европы превратились в чахлых ревматиков и подагриков с
искривленными позвоночниками. А женщины были неспособны рожать. Они вымерли.
29.
Флуктуации численности вызываются самыми различными причинами. И они не всегдаодинаковые для разных видов. Периодические колебания численности популяций, имеющих 10 11-летний период, объясняются периодичностью активности Солнца: количество пятен на Солнце
меняется с периодом в 11 лет. Как "земное эхо солнечных бурь" (А. А. Чижевский) на Земле
меняется погода. В связи с этим изменяется продуктивность фитоценозов - изменяется количество
корма для растительноядных животных, происходят изменения по всем цепям питания экосистем.
Вот свидетельство очевидца. "1955 год называли на целине "годом отчаяния"... За все лето,
начиная с мая, на землю не упало и капли дождя... С утра раскаленное солнце начинало свою
опустошительную работу, медленно плыло в белесом, выцветшем небе, излучая нестерпимый
зной. И так неделя за неделей, месяц за месяцем. Мы знали, что жара и суть в этом краю
никому не в диковинку, но не знали еще зловещей неумолимости стенного календаря, который
раз в 10 лет преподносит особенно жесткие, губительные засухи".
Количество корма есть причина флуктуации у сибирского шелкопряда: он дает вспышку после
сухого теплого лета.
Может вызвать вспышку численности и стечение многих обстоятельств. Например, у берегов
Флориды наблюдаются "красные приливы". Они непериодичны и для их роявления необходимы
такие события: обильные ливни, смывающие с суши микроэлементы (железо, цинк, кобальт - их
концентрация должна совпадать до десятитысячной доли процента), пониженная соленость поды,
определенная температура и безветрие у берега. При таких условиях водоросли динофлагелляты
начинают интенсивно делиться. Теоретически из одной одноклеточной динофлагелляты в
результате 25 последовательных делений могут произойти 33 млн. особей. Вода от них становится
красной.
читать дальше>>
30.
Это явление известно людям давно: "И вся вода в реке превратилась в кровь; и рыба в рекевымерла, и река воссмердела, и Египтяне не могли пить воды из реки" (Библия, Книга Исход).
Динофлагелляты выделяют в воду смертоносный яд, вызывающий паралич, а затем гибель рыбы и
других обитателей моря.
Человек может своей деятельностью вызвать вспышку численности некоторых популяций.
Результатом антропического воздействия есть возрастание численности сосущих насекомых (тлей,
клопов и др.) после обработки полей инсектицидами, которые уничтожают их врагов.
Благодаря человеку кролики и кактус опунция в Австралии, домовые воробьи и непарный
шелкопряд в Северной Америке, колорадский жук и филлоксера в Европе, канадская элодея,
американская норка и ондатра в Евразии дали неимоверные вспышки численности после
попадания на эти новые для них территории, где не было их врагов.
Резкие непериодические колебания численности могут возникать вследствие природных
катастроф. Например, на пожарищах обычны вспышки численности иван-чая и связанного с ним
сообщества насекомых. Многолетняя засуха превращает болото в луг и вызывает рост
численности членов биоценоза луга.
31.
«Красный прилив»Одноклеточная водоросль
Динофлагеллята
32.
Чарльз Дарвин (1809 –1882 гг.) великийученый - биолог, естествоиспытатель, автор
теории эволюции. С помощью теории
естественного отбора объяснил процесс
развития органического мира.
33.
Де Фриз (Де Фрис) Хуго (1848-1935) нидерландский ботаник, один из основателейучения об изменчивости и эволюции. Провел
первые систематические исследования
мутационного процесса. Разработал концепцию
эволюции посредством мутаций (мутационная
теория Де Фриза).
34.
Четвериков Сергей Сергеевич (1880-1959),российский генетик, один из
основоположников эволюционной и
популяционной генетики. Одним из первых
связал закономерности отбора в популяциях
с динамикой эволюционного процесса.
Труды по зоогеографии и энтомологии.
35.
Примеры естественного отбораОдин из сыновей Ч. Дарвина - майор Леонард Дарвин (1850-1943) - задал отцу в конце его жизни вопрос: можно ли и когда ученые смогут
наблюдать процесс естественного отбора в природе. Дарвин ответил: "Если материал собирать правильно, то можно убедиться в
существовании эволюционных явлений, вероятно, в течение какихнибудь 50 лет".
Е. Б. Паультон (Паултон) в 1898 г. сообщил о первых опытах моделирования естественного отбора на бабочке крапивнице. 600 куколок крапивницы
разместили на коре деревьев, на изгороди, стенах построек, желтых листьях крапивы. Если окраска куколки соответствовала цвету фона, птицы
уничтожали не более 57 % куколок. В случае контрастности окраски куколок и фона уничтожалось более 90 % куколок. Так экспериментально была
подтверждена возможность селекционного происхождения покровительственной окраски.
В. Ф. Р. Уэлдон в 1898 г. опубликовал результаты многолетнего исследования популяции крабов в бухте Плимута. После постройки нового большого
мола в Плимутской бухте, загородившего узкий вход в бухту, было замечено, что ширина головогруди у крабов достоверно уменьшилась.
Биометрические измерения показали, что средняя ширина головогруди краба из года в год убывает.
Уэлдон выяснил, что избирательная гибель крабов происходит из-за загрязнения жаберной полости илом, поднимаемым со дна винтами пароходов.
В специальном аквариуме Уэлдон поставил опыт, в котором 248 отловленных крабов жили в условиях постоянного взмучивания ила. Оказалось, что
у 154 погибших крабов жаберные полости были забиты илом. Биометрические исследования показали, что у выживших крабов относительная
ширина головогруди была меньше, чем у погибших.
А. Р. Чеснола в 1904 году провел эксперимент по проверке адаптивного значения покровительственной окраски у богомолов. Он привязывал ниткой
к зеленой траве насекомых разной окраски. Через три недели из 45 коричневых осталось 10, а из 20 зеленых - 20 уцелели. Затем к коричневой траве
было привязано 20 коричневых и 25 зеленых богомолов. На 12-й день оказалось, что все зеленые богомолы были истреблены, а коричневые - живы.
Ж. Тесье поставил опыт по проверке гипотезы Дарвина о роли ветра в качестве фактора естественного отбора, обусловливающего возникновение на
океанических островах бескрылых насекомых. Смешанную популяцию плодовой мушки, состоящую из бескрылых особей (гомозиготы по
рецессивному гену) и крылатых особей, поместили на террасе биостанции в Роскоффе, расположенной на берегу Атлантического океана. Пищи
было в избытке. Бескрылых вначале было 12,5 %. Через 2 месяца под действием ветра изменился фенотипичсский состав популяции - бескрылые
дрозофилы составляли уже 67 %. Затем был поставлен обратный эксперимент. Эту же популяцию перенесли в помещение. Через несколько
поколений большинство популяции составляли крылатые особи. Это было связано с тем, что они были более плодовиты и имели большую
продолжительность жизни, чем бескрылые. Этот эксперимент полностью подтвердил гипотезу Ч. Дарвина. Работа Тесье была опубликована в 1933
году - через 51 год после смерти Ч. Дарвина.
Дж. Гаррисон в 1920 г. опубликовал работу по направлению ЕО на примере бабочки пяденицы. Йоркширский лес до 1800 г. был смешанным. Позже
лес разделился на 2 части полосой вересковой заросли. Буря 1885 г. привела к выпадению в южной части леса сосны. Доминирующим деревом
здесь стала береза. В северном участке леса в процессе естественной сукцессии березу вытеснили хвойные деревья. В 1907 г. при обследовании
соснового леса Гаррисон обнаружил, что среди бабочек только 4 % были светлыми, а остальные 96 % бабочек пяденицы были темные. В березовом
же лесу светлые бабочки составляли 85 %, а темные - 15 %. По остаткам крыльев Гаррисон установил, что в сосновом лесу светлые формы
поедаются примерно в 30 раз чаще, чем темные. Изменение состава деревьев в северном и южном участках леса изменило направление
естественного отбора. Под действием естественного отбора изменился генофонд дочерних популяций, изменился фенотипический состав
популяций.
36.
Источники1.
Георгиевский А.Б. Дарвинизм: учеб. Пособие для студентов биол. и хим. спец. пед. ин-тов. – М.:
Просвещение, 1985. – 271 с.
2.
http://ru.wikipedia.org/wiki/Естественный_отбор
3.
http://www.avifarm.ru
4.
http://evolution2.narod.ru/index.htm
5.
www.nt-creaz.org.ua
6.
http://www.wikiznanie.ru/ru-wz/index.php/Чарлз_Дарвин
7.
http://www.edu.ru/db/portal/spe/progs/540102_pp.04.htm
8.
http://zoolife.com.ua/pageid1037.html
9.
http://macroevolution.narod.ru/factory04.htm
10. http://www.glossary.ru/cgi-bin/gl_sch2.cgi?RUgqyuw:!_iur8.oo
11. http://mylearn.ru/kurs/7
12. http://bio.1september.ru/articlef.php?ID=200304303
13. http://shkola.lv/index.php?mode=lsntheme&themeid=11
14. http://files.school-collection.edu.ru/dlrstore/76e3b8fe-d031-4277-a547-071596df97ef/%5BBIO9_0738%5D_%5BIM_03%5D.swf
15. http://festival.1september.ru/articles/311111
16. http://www.darwin.museum.ru/expos/livenature/
37.
справкаследующий слайд
предыдущий слайд
возврат к содержанию
назад
выйти из программы
В тексте будут встречаться
выделенные слова. Наведите
курсор на такое слово и
нажмите левую кнопку мыши.