






")

")















")



































")

















 обеспечивает изменение структуры ДНК")








")

















 biology
biologySimilar presentations:

. Функции ДНК транскрипция")








Нуклеиновые кислоты: структура и функции
1. План проекта:
• 1. Нуклеиновые кислоты: структура ифункции
• 2. Биосинтез ДНК
• 3. Молекулярные механизмы генных
мутаций. Репарация ДНК
• 4. Биосинтез молекул РНК. Транскрипция в
клетках прокариот и эукариот
• 5. Процессинг РНК в клетках прокариот и
эукариот
2. 1. Нуклеиновые кислоты: структура и функции
• Основные положения молекулярнойбиологии
• Доказательства генетической роли
нуклеиновых кислот
• Пурины и пиримидины
• Химический состав нуклеиновых кислот.
• Структура нуклеиновых кислот
• Модель строения ДНК
• Цепи в ДНК комплементарны и
антипараллельны
• Биологические функции ДНК
• Отличия молекул ДНК и РНК
3. Основные положения молекулярной биологии:
• ДНК - носитель генетической информации,реплицируется по принципу матричного синтеза
• РНК синтезируется на матрице ДНК, копируя
определенный участок (ген)
• Белок синтезируется на матрице РНК,
последовательность аминокислот в белке
определяется последовательностью нуклеотидов
в мРНК
4. Доказательства генетической роли ДНК
• Открытие нуклеиновых кислот –Ф.Мишер, 1869.
• Трансформация бактерий – Ф.Гриффитс,
1928-1931.
• 1944 г. - О. Эйвери, К. Мак-Леод и М. МакКарти доказали, что ДНК является
генетическим материалом бактерий
• 1952 г – А. Херши и М. Чейз доказали, что
ДНК является генетическим
материалом бактериофагов
5. Химический состав нуклеиновых кислот.
• ДНК и РНК – линейные полимеры,состоят из последовательно
расположенных структурных единиц
- мономеров
• мономеры ДНК дезоксирибонуклеотиды
• мономеры РНК - рибонуклеотиды
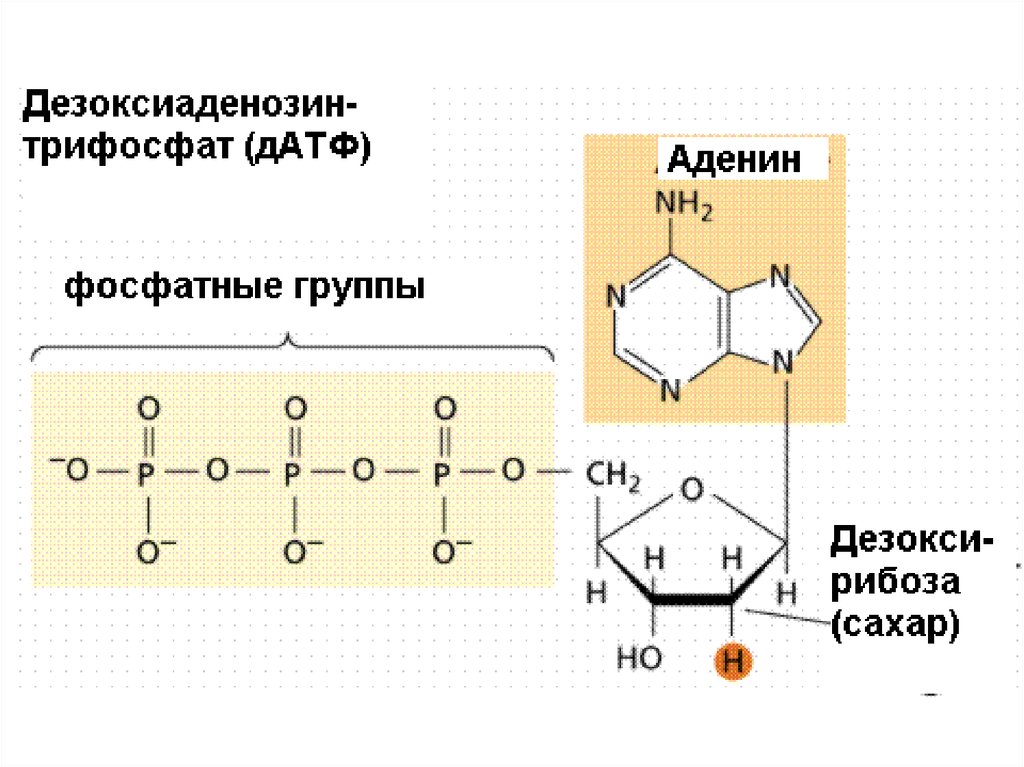
6. Структура нуклеотида
7. Пурины и пиримидины
аденингуанин
8.
9. Первичная структура нуклеиновых кислот (ДНК и РНК)
• определяется последовательностьюнуклеотидов в полинуклеотидной цепи
• Нуклеотиды соединяются с помощью
ковалентных 3’, 5’- фосфодиэфирных
связей
• за направление полинуклеотидной цепи
принято направление от 5’ → к 3’-концу
10. Правила Э.Чаргаффа
• количество пуриновых оснований (A+Г) вмолекуле ДНК всегда равно количеству
пиримидиновых оснований (Т+Ц),
• количество аденина равно количеству
тимина [А=Т, А/Т= 1]; количество гуанина
равно количеству цитозина [Г=Ц, Г/Ц=1];
• соотношение (Г+Ц)/(А+Т)=К, где К коэффициент специфичности, является
постоянным для каждого вида живых
организмов
11. Модель строения ДНК, предложенная Уотсоном и Криком (1953)
12. Цепи в ДНК комплементарны и антипараллельны
• две цепи ДНК закручены в спиральвокруг общей оси
• Цепи комплементарны и
антипараллельны
• основания находятся внутри
молекулы ДНК,
• снаружи находится сахарофосфатный скелет
• диаметр спирали - 2 нм, каждые 10
п.н. составляют один виток спирали
• Расстояние между нуклеотидами –
0,34 нм
• Один виток спирали – 3,4 нм
13. Цепи в ДНК комплементарны и антипараллельны
14. Химические связи, стабилизирующие вторичную структуру ДНК:
• Водородные связи – образуютсямежду комплементарными
основаниями
• Стэкинг-взаимодействия – это
гидрофобные связи, которые
образуются между плоскими
основаниями, которые расположены
друг на другом в одной цепи ДНК
15. Биологические функции ДНК
• Хранение генетической информации• Передача генетической информации
• Реализация генетической информации
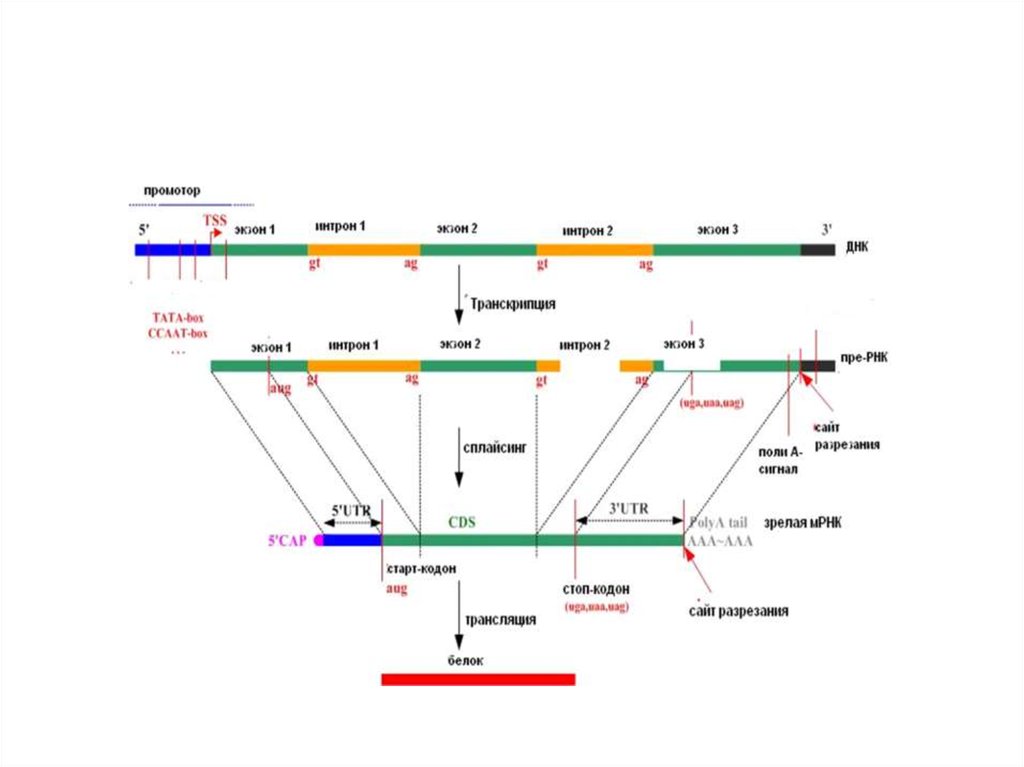
• Изменение генетической информации
16. Отличия молекул ДНК и РНК
17. Отличия в структуре рибозы и дезоксирибозы, урацила и тимина
18. Типы молекул РНК:
мРНК
тРНК,
рРНК
мяРНК
гяРНК
мцРНК
siРНК
XIST-РНК
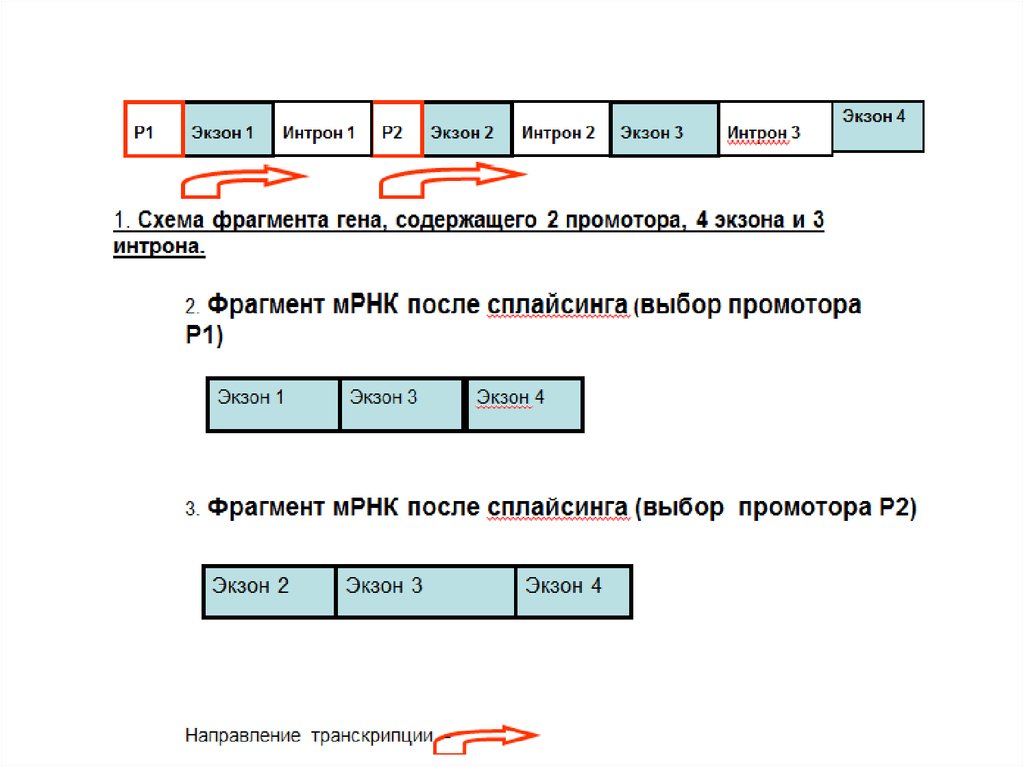
РНК вирусов
19. 2. Биосинтез ДНК
• Полуконсервативный механизм репликации• Синтез новых цепей ДНК катализируют
ДНК-полимеразы
• Реакция полимеризации нуклеотидов
• Этапы синтеза ДНК
• Репликация. Репликон.
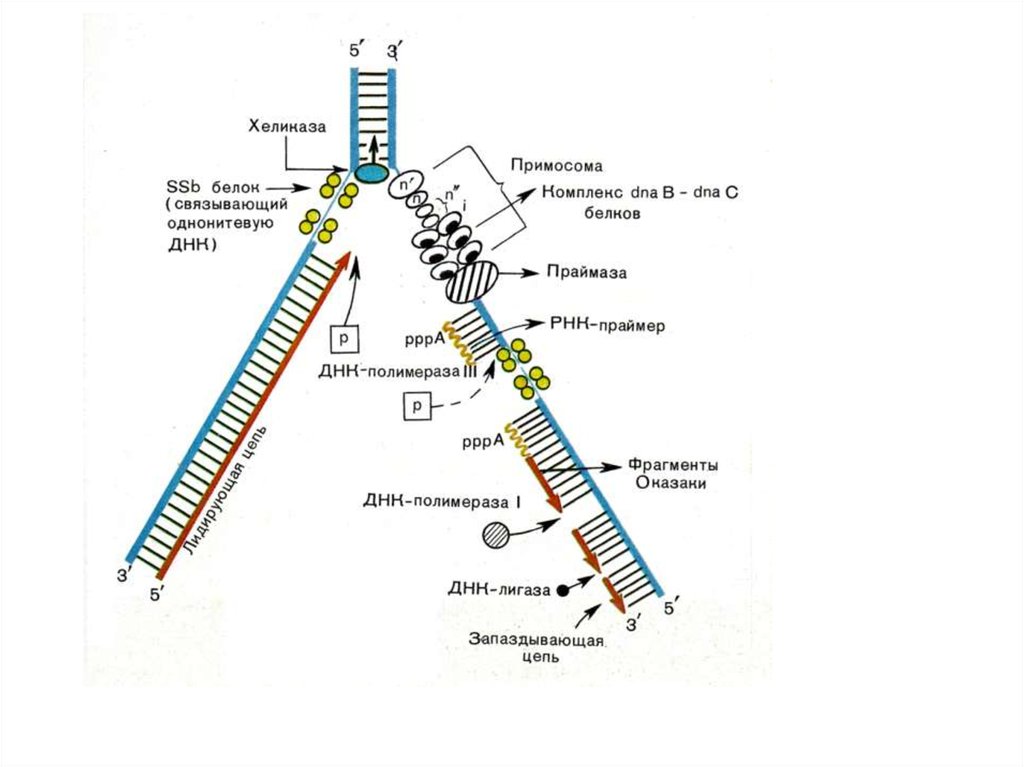
• Синтез фрагмента Оказаки
• Окончание синтеза ДНК
• Синтез теломерных участков ДНК
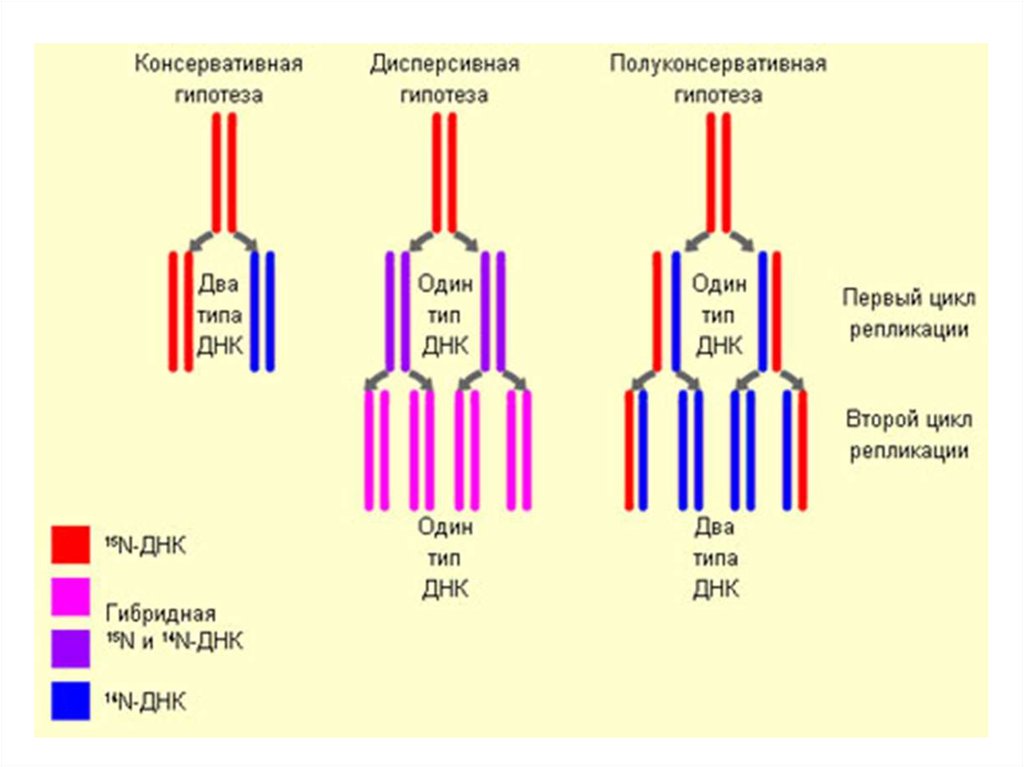
20. Полуконсервативный механизм репликации
• В основе процессарепликации лежит
принцип копирования
материнской цепи ДНК с
образованием 2-х
одинаковых дочерних
молекул ДНК
• Каждая дочерняя
молекула ДНК содержит
одну старую и одну
новую
полинуклеотидную цепь
21.
22. Синтез новых цепей ДНК катализируют ДНК-полимеразы
23. Реакция полимеризации нуклеотидов
• Суммарное уравнение реакции синтезаполинуклеотида:
• (дНМФ)n + дНТФ = (дНМФ)n+1 + ФФ
• ФФ
2Ф
24. ДНК-полимеразы имеют 2 субстрата:
• комплекс матрица-затравка и• дНТФ (дезокси-нуклеозидтрифосфаты):
дАТФ, дГТФ, дЦТФ, ТТФ
• Субстрат – это вещество, на которое
действует фермент
25. Этапы синтеза ДНК
• Инициация – образованиекомплекса матрица – затравка
• Полимеризация – синтез новых
цепей ДНК
• Терминация – окончание
синтеза ДНК
26. Инициация синтеза ДНК
• Origin (Ori) - точка начала репликации – это участокмолекулы ДНК (300 п.н.),со специфической
последовательностью нуклеотидов, который
узнают специальные белки репликации:
• dnaA – у прокариот
• RPA - у эукариот
27. Репликон – фрагмент молекулы ДНК от одной точки начала репликации до другой
• Геном прокариот содержитодин репликон
• Геномы эукариот – сотни и
тысячи репликонов
28. Кольцевая ДНК прокариот имеет одну точку начала репликации (один репликон)
29. Репликоны эукариот
30. Репликация ДНК эукариотической хромосомы
• Показан один из многих репликонов• Репликативные вилки движутся в
противоположных направлениях от точки
начала репликации
31.
32. Образование комплекса матрица-затравка
Образование комплекса матрицазатравка• ДНК-полимеразы не могут
начать инициацию синтеза
новой цепи на матрице
• Праймаза синтезирует РНКпраймер (primer),
• ДНК-полимераза использует 3’конец праймера для синтеза
новой цепи ДНК
• После окончания синтеза ДНК
РНК-праймер удаляется с
помощью экзонуклеаз
33.
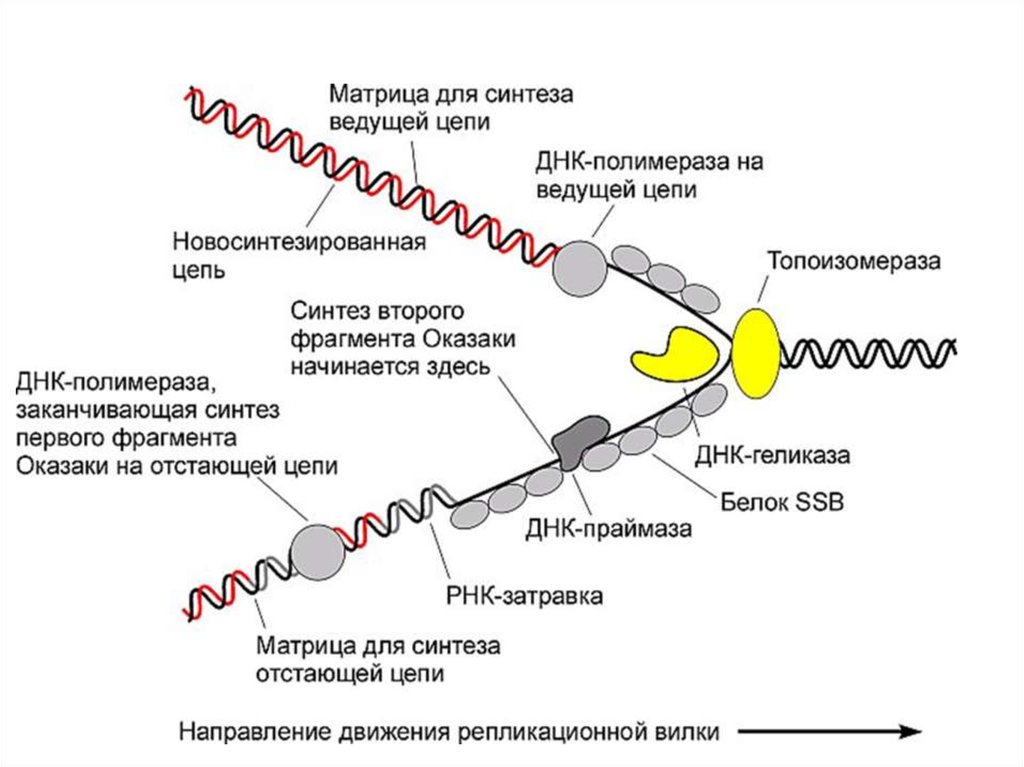
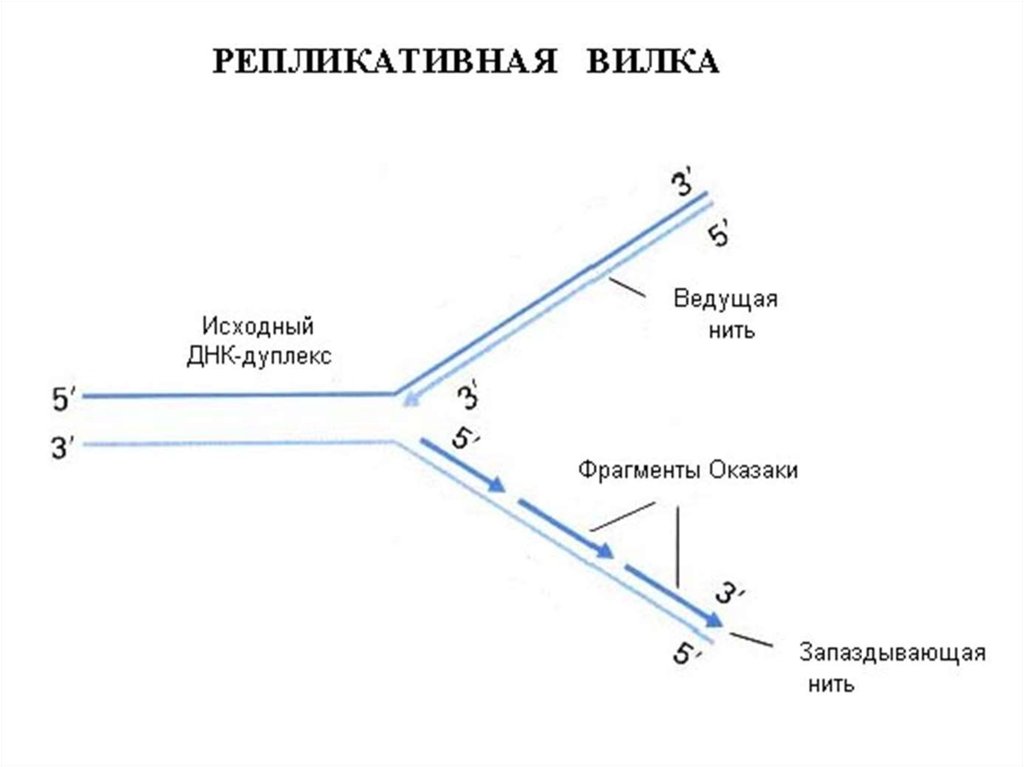
34. Асимметричность репликатавной вилки
35. Синтез фрагментов Оказаки на отстающей цепи
36. ДНК-полимеразы прокариот
• ДНК полимераза I:• Удаляет РНК-праймеры (5’- 3’экзонуклеазная активность)
• Застраивает пробелы цепочкой ДНК
(полимеризная активность)
• Коррегирующая 3’-5’- экзонуклеазная
активность
37. Окончание синтеза ДНК
• РНК-полимераза I (прокариот),(РНКазаН у эукариот) - удаляет РНКпраймеры
• ДНК-лигаза – соединяет
фрагменты Оказаки
• теломераза - синтез теломерных
участков хромосом эукариот
38.
39. Отличия репликации в клетках прокариот и эукариот
40. ДНК- полимеразы эукариот
• Полимеразы α и δ осуществляютинициацию (α) и элонгацию (δ)
синтеза ДНК
• Полимеразы β и ζ – участвуют в
репарации ДНК
• Полимераза γ –синтез мтДНК
• Полимераза ε – возможно синтез
отстающей цепи ДНК
41. Синтез теломерных участков ДНК
42. 3. Молекулярные механизмы генных мутаций. Репарация ДНК
Типы генных мутаций
Мутация со сдвигом рамки считывания
Мутации типа замены оснований
Мутация в гене β-цепи гемоглобина
Факторы, индуцирующие мутации, называются
мутагены
Действие на ДНК ионизирующего излучения
Основные механизмы репарации
Молекулярные механизмы генных мутаций
Нарушения системы репарации ДНК
Болезни экспансии тринуклеотидов
43. Основные понятия:
• Мутации - стойкие изменения структурыДНК по сравнению с геномной ДНК вида
• Генные мутации – изменение
последовательности нуклеотидов в пределах
гена
• Хромосомные мутации - изменение
структуры или количества хромосом в клетке
• Мутон – наименьший участок ДНК,
изменение которого приводит к мутации
44. Типы генных мутаций
45. Мутация со сдвигом рамки считывания
46. Мутации типа замены оснований
транзиции - замена пурина на пурин или пиримидинана пиримидин
трансверсии - замена пурина на пиримидин
47. Эритроциты больного серповидно-клеточной анемией
Эритроциты больного серповидноклеточной анемией48. Мутация в гене β-цепи гемоглобина
49. Факторы, индуцирующие мутации, называются мутагены
Физические мутагены:
УФ-излучение
X-Ray
α-β-γ- излучение
Химические мутагены:
HNO2
нитрозогуанидин
формальдегид
бензапирен
митомицин С
акридиновые красители
химиопрепараты
50. Действие на ДНК ионизирующего излучения
51. УФ-лучи повреждают ДНК за одну пикосекунду
52. Молекулярные механизмы генных мутаций
• Образование пиримидиновых димеров• Включение в ДНК аналогов оснований
• Явление таутомеризации
• Некоторые типы повреждений ДНК: апуринизация - разрыв
гликозидных связей между пурином и д-рибозой, образуется
пробел в цепи ДНК (АP-сайт) – результат: делеция нуклеотида,
Дезаминирование - потеря аминогруппы:цитозин в урацил,
(алкилирование) - присоединение метильной группы к
азотистому основанию –результат:
• гуанин превращается в О6 - метилгуанозин (аналог аденина) пара Г-Ц замещается на Т-А
53. Основные механизмы репарации:
• Репарация - это свойство живых организмоввосстанавливать повреждения ДНК, которые
появились в результате ошибок репликации или
воздействия мутагенных факторов
• Прямое восстановление исходной структуры ДНК.
Репарация без репликации
• Эксцизионная репарация: вырезание
поврежденного основания или вырезание
поврежденного участка ДНК
• Репарация неспаренных оснований
• SOS-индуцированная репарация
• Пострепликативная репарация
54. Прямое восстановление структуры ДНК:
• Прямая метилтрансферазная реактивация :• Корректирует метилирование оснований с
помощью белка метилтрансферазы
• Фотореактивация
• необходим фермент фотолиаза и видимый свет
• корректирует тиминовые димеры без вырезания
ДНК
• *Система обнаружена у бактерий и растений, у
млекопитающих не обнаружена.
55. Вырезание поврежденного основания
56. Эксцизионная репарация у животных
57. Репарация неспаренных оснований
58. репарация неспаренных нуклеотидов
59. Пострепликативная репарация
60. SOS - индуцированная репарация
• Повреждения ДНК достигают критическогоуровня
• синтезируется белок RecA
• Он связывается с белком LexA (ингибитор
генов SOS-репарации ) и разрушает его
• Начинается синтез белков, которые
осуществляют SOS-репарацию
61. Нарушения системы репарации ДНК - причина наследственных болезней
• Пигментная ксеродерма (XP) - мутации генов XPB и XPDприводят к нарушениям системы эксцизионной репарации ДНК
после УФ-облучения (развитие рака кожи)
• Синдром Вернера - мутации гена WRN (8p12) приводят к
неправильной репарации (быстрое старение, риск развития
диабета и рака)
• Синдром Линча - мутации генов hMSH2 hMLH1 приводят к
нарушениям системы репарации неспаренных нуклеотидов
(развитие рака прямой кишки)
• *Нарушение системы репарации приводит к нестабильности
генома и развитию онкологических заболеваний.
62. Болезни экспансии тринуклеотидов
• Хорея Хантингтона - аутосомно-доминантноезаболевание с поздней манифестацией (после
40 лет).
• У здоровых людей ген хантигтин содержит от 8
до 31 повтора триплета ЦАГ, у больных - от 36
до 80
• добавление триплетов ЦАГ приводит к синтезу
аномального белка хантигтина, который
вызывает апоптоз клеток мозга, нарушениям
ЦНС и смерти
63. 4. Биосинтез молекул РНК. Транскрипция в клетках прокариот и эукариот
Этапы экспрессии генов
Принципы синтеза РНК на ДНК-матрице
Этапы транскрипции
РНК-полимераза прокариот
Стадии инициации транскрипции
Элонгация
Терминация транскрипции прокариот
РНК-полимеразы эукариот
Инициация транскрипции
образование базового
транскрипционного комплекса
• Транскрипционный цикл
64. Этапы экспрессии генов
65. Принципы синтеза РНК на ДНК-матрице: комплементарность и антипараллельность
66. РНК-полимеразы катализируют реакцию синтеза молекул РНК на ДНК матрице
67. На 5’-конце РНК содержится трифосфат (АТФ или ГТФ)
68. Этапы транскрипции
• Связывание ДНК-матрицы – узнаваниепромотора, образование открытого
двойного комплекса
• Инициация – соединение 2-х первых
нуклеотидов, образование открытого
тройного комплекса, начало синтеза РНК
• Элонгация – продолжение синтеза РНК
• Терминация – завершение синтеза РНК
69. РНК-полимераза прокариот
• Кор фермент α2ββ’обладает
полимеризующей
активностью
• σ – фактор –
обеспечивает
узнавание промотора и
инициацию
• Полный фермент –
холофермент (Mg2+ )
α2ββ’σ
70. Структура промоторов генов прокариот
• Промотор – специфическая последовательность ДНК,необходимая для образования комплекса РНКполимеразы с ДНК-матрицей.
• -10 последовательность - ТАТААТ (блок Прибнова)
• -35 последовательность - ТТГАЦА
• Стартовая точка (+1) – нуклеотид, с которого начинается
синтез РНК
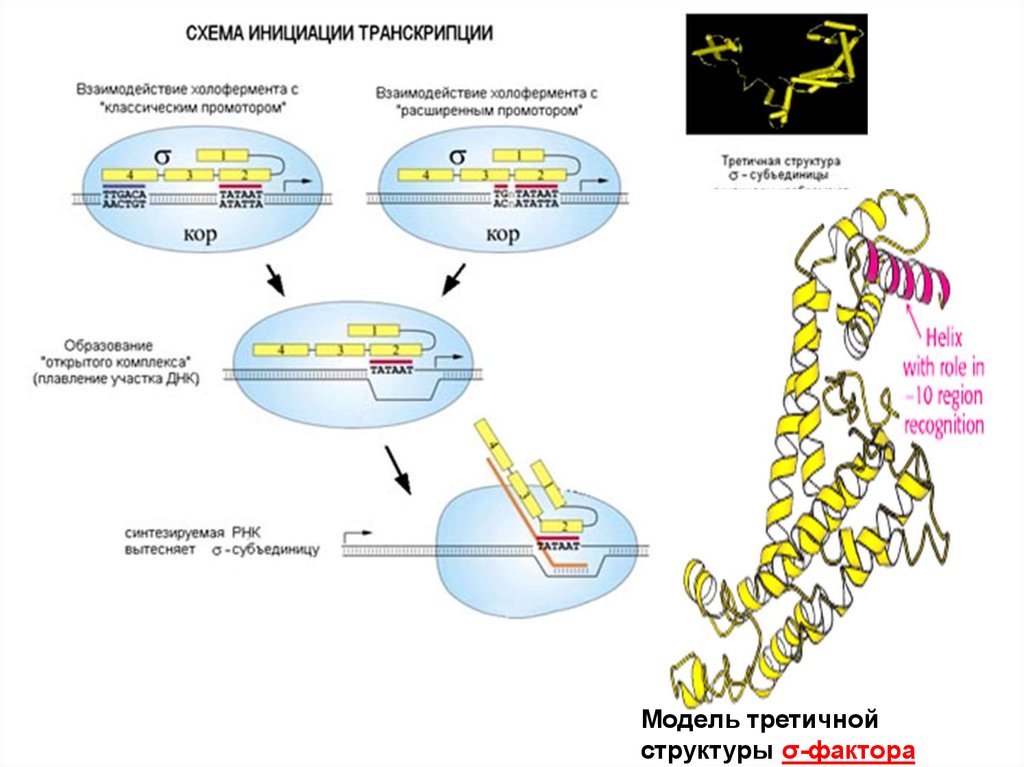
71. Стадии инициации транскрипции:
• образование «закрытого» комплекса,• образования «открытого двойного»
комплекса (с расплетанием участка ДНК)
• образования «открытого тройного»
комплекса (синтез коротких РНК без
диссоциации σ-фактора)
• После синтеза фрагмента РНК (9-12
нуклеотидов) σ-фактор покидает промотор
и начинается стадия элонгации
72.
Модель третичнойструктуры σ-фактора
73.
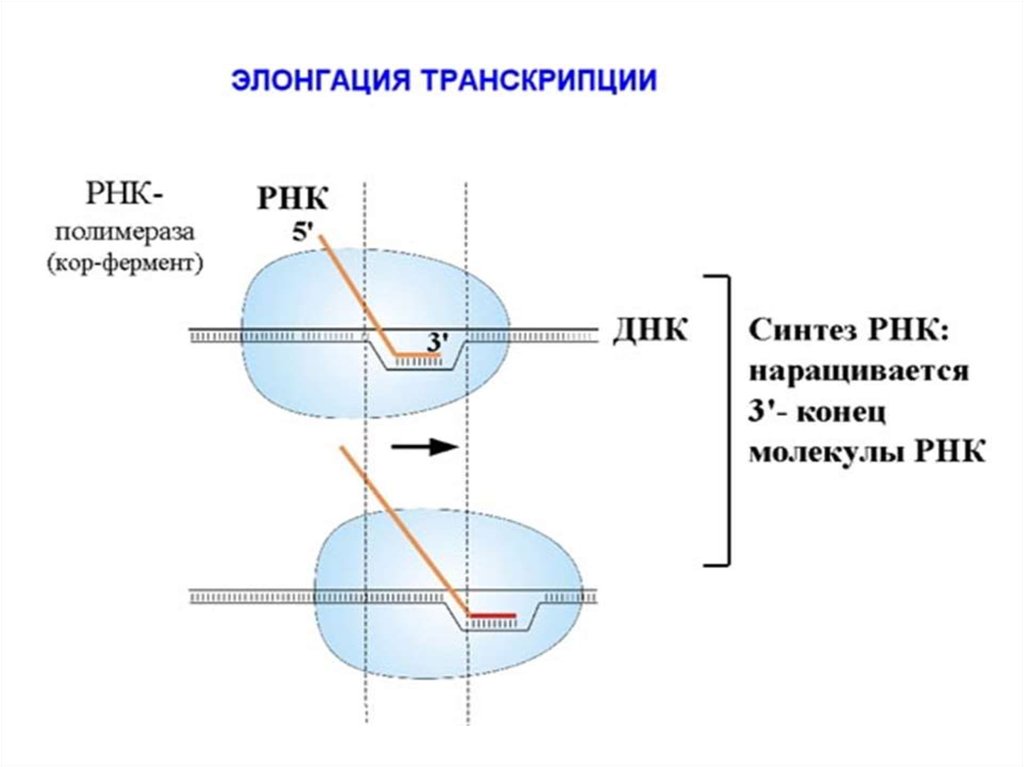
74. Элонгация - синтез молекулы РНК на ДНК-матрице
75. Терминация транскрипции прокариот
Терминация транскрипциипрокариот
• Терминация транскрипции прокариот
может происходить двумя способами: с
участием ρ (ро)-фактора и без него
• терминатор – это специфическая
последовательность ДНК, на которой
происходит терминация транскрипции
76. ρ-независимая терминация транскрипции
• На схеме показан 3’-конец молекулы РНК• После прохождения терминатора молекула РНК образует
шпильку, которая приводит к остановке транскрипции
• наименее стабильный уридил-адениловый дуплекс (выделен
розовым цветом) разрушается
77. ρ-зависимая терминация транскрипции
• Rho (ρ)-фактор – специальный белок терминации транскрипции• Во время синтеза движется по РНК к 3’-концу, используя
энергию
• Когда r-фактор “догоняет” РНК-полимеразу происходит
терминация транскрипции
78. Рифампицин ингибирует инициацию транскрипции прокариот
Рифампицин блокирует боковой проход РНК-полимеразы для трифосфатов.79. Отличия транскрипции в клетках прокариот и эукариот
80. РНК-полимеразы эукариот
• РНК-полимераза I – синтез рРНК (28S, 18S и5,8S рРНК)
• РНК-полимераза II – синтез мРНК и мяРНК
• РНК-полимераза III – синтез тРНК, 5S рРНК,
некоторых мяРНК
• РНК-полимераза митохондрий – состоит из
одной субъединицы, ген которой находится
в ядерной ДНК
81. Amanita phalloides синтезирует α – аманитин, к которому чувствительна РНК-полимераза II эукариот
Amanita phalloides82. Для инициации синтеза РНК у эукариот необходимы
• Специфические белки – факторытранскрипции (транс-факторы)
• Регуляторные последовательности ДНК
(цис-элементы) – промоторы, энхансеры и
сайленсеры
83. Универсальные последовательности промоторов класса II
• ТАТА-бокс (-25 п.н.) (блок Хогнесса)• ЦААТ-боксы (- 50/-150 п.н.)
• ГЦ-мотивы (-300 п.н.)
• Классический промотор класса II содержит
ТАТА-бокс последовательность ТАТААА (блок
Хогнесса)-, – точно определяют точку начала
транскрипции.
84. Инициация транскрипции
• в зоне ТАТА-бокса должны собратьсяфакторы транскрипции TFII (Transcriptional
Factors of RNA-polymerase II):
• TFIIA, TFIIB, TFIID, TFIIE, TFIIF и TFIIH
• первым с ДНК связывается белок TFIID
• TFIID – это комплекс белков, в состав
которого входит белок TBP и TAF белки
85. образование базового транскрипционного комплекса
• факторы транскрипции связываются спромотором в зоне ТАТА-бокса в
определенной последовательности:
• TFIIА, TFIIВ, TFIIF
• Затем присоединяется РНК-пол. II
и TFIIE
• TFIIH присоединяется последним
и фосфорилирует РНК-полимеразу
Для инициации транскрипции генов эукариот
необходимы многочисленные белки – факторы
транскрипции. Поверхность белка ТВР обеспечивает
связывание промотора с другими компонентами.
Другие факторы транскрипции собираются в
определенной последовательности: TFIIА, TFIIВ, TFIIF.
Затем присоединяются РНК-полимераза II и TFIIE,
образуя основной транскрипционный комплекс.
86. Ключевым моментом инициации является узнавание ТАТА-бокса белком TBP
• Белок TBP (TATA Binding Protein) – входит всостав комплекса TFIID
• TBP связывается с ТАТА боксом в 100000 раз
прочнее, чем с другими
последовательностями ДНК
• Это связывание индуцирует
пространственные изменения в молекуле
ДНК.
87. Белок TBP (TATA Binding Protein) обеспечивает изменение структуры ДНК
• Двойная спираль частично раскручивает,расширяя малую бороздку и обеспечивая
гидрофобные взаимодействия между ДНК и
белком TBP
ДНК связывается с вогнутой
поверхностью белка.
88. Транскрипционный цикл
• для активации фермента и успешной транскрипции необходимофосфорилирование фермента. Большинство транскрипционных
факторов высвобождается до того как, РНК-полимераза покидает
промотор.
• Терминация. Дефосфорилирование РНК-полимеразы связано с
терминацией транскрипции
89. 5. Процессинг РНК в клетках прокариот и эукариот
Этапы реализации генетической информации
Процессинг пре- РНК эукариот происходит в ядре и в митохондриях
Процессинг рРНК и тРНК
Экзон
Этапы экспрессии генов эукариот
Полиаденилирование
Механизмы сплайсинга интронов:
Механизмы альтернативного сплайсинга:
Альтернативный выбор сигнала полиаденилирования
90. Этапы реализации генетической информации
• Транскрипция – синтез молекул РНК,образование первичного транскрипта (преРНК)
• Процессинг –модификация первичного
транскрипта (пре-РНК) и образование
зрелых молекул РНК
91. Процессинг пре- РНК эукариот происходит в ядре и в митохондриях
92. Процессинг РНК в клетках прокариот
• Ген – это участок молекулы ДНК, кодирующий синтезфункциональной молекулы РНК
• Цистрон – участок молекулы мРНК, который
кодирует синтез одной полипептидной цепи
• Молекулы мРНК прокариот полицистронны, т.е. они
служат матрицей для одновременного синтеза
нескольких полипептидов
мРНК прокариот не процессируются
мРНК прокариот обычно являются Полицистронными , т.е. включают в
себя последовательности нуклеотидов нескольких генов одного оперона
мРНК бактерий при выполнении функций матричных РНК в трансляции не
требуют разбиения на последовательности отдельных генов и могут
транслироваться непосредственно рибосомами с образованием
функционально активных белков
93. Процессинг рРНК и тРНК
• Зрелые молекулы рРНК и тРНК образуютсяу прокариот и эукариот в результате эндо- и
экзонуклеазных воздействий на их
предшественники.
• У эукариот в некоторых случаях
вырезаются интроны из пре-рРНК и претРНК
94.
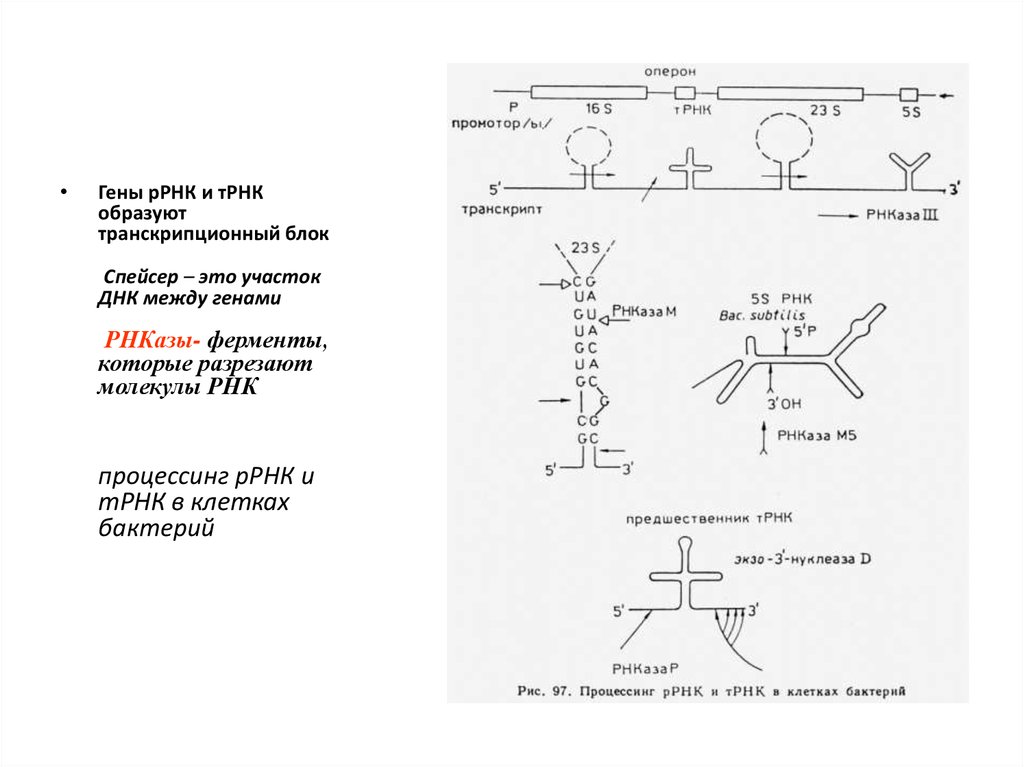
Гены рРНК и тРНК
образуют
транскрипционный блок
Спейсер – это участок
ДНК между генами
РНКазы- ферменты,
которые разрезают
молекулы РНК
процессинг рРНК и
тРНК в клетках
бактерий
95. Процессинг тРНК и рРНК у прокариот
• Молекулы 5S, 16S и 23S рРНК синтезируются в составеобщего первичного транскрипта. В спейсерных участках
между генами 16S и 23S рРНК расположены гены тРНК
• РНКаза III участвует в процессинге пре- рРНК у
бактерий,
• В процессинге пре- тРНК у бактерий участвует РНКаза P
и РНКаза D
• РНКаза Р является рибозимом, т.к. содержит
собственную РНК, которая обладает эндонуклеазной
активностью.
• Некоторые РНК могут катализировать химические
реакции
96. Этапы процессинга тРНК прокариот:
• Модификация 5’ –конца - РНКаза Р
• Модификацию 3’ –
конца осуществляет
РНКаза D
• Модификация
некоторых азотистых
оснований и
формирование зрелой
структуры тРНК
97. Этапы процессинга тРНК дрожжей (эукариоты)
• Удалениеинтрона
• Вырезание 5’
конца (лидер)
• Удаление УУ с
3’конца
• Присоединен
ие ЦЦА к
3’концу -тРНК
• Модификаци
я азотистых
оснований
98. Процессинг рРНК эукариот
• Транскрипционный блок содержит гены 18S; 5,8S; 28S - рРНК, разделенные спейсерами
• три зрелые молекулы рРНК образуются при
расщеплении спейсеров эдонуклеазой
• Некоторые эукариоты содержат интрон в
28S пре- рРНК, который вырезается
(аутосплайсинг) и образуется 26S рРНК
99. Экзон-интронная структура генов эукариот
Структура α- и β-глобиновых генов
Экзоны (тёмно-красный цвет ), разделены интронами (голубой цвет).
Цифры над генами указывают аминокислотные остатки кодируемого
полипептида.
5’- 3’- не транслируемые области содержатся в первом и последнем экзонах
(розовый цвет). Они присутствуют в зрелой мРНК, но не транслируются.
Экзон-интронная структура характерна для генов вирусов и эукариот.
100. Этапы процессинга пре - мРНК эукариот
• Кэпирование модификация 5’конца• Полиаденилировани
е - модификация 3’конца
• Сплайсинг - удаление
интронов и
соединение экзонов
101. Этапы экспрессии генов эукариот
102.
103. Модификация 5’-конца – кэпирование
Кэп – это 7-метил-гуанозин соединенный в 5’-5’-ориентации с
первым нуклеотидом мРНК
• Кэп присоединяется с помощью фермента гуанозил-7метилтрансферазы к первому 5’-трифосфату мРНК сразу после
транскрипции с помощью особой 5’ - 5’- связи
104. Модификация 3’-конца пре-мРНК - полиаденилирование
Модификация 3’-конца пре-мРНК полиаденилирование105. Модификация 3’-конца – полиаденилирование
• Последовательность ААУААА служитсигналом полиаденилирования
• Специальная эндонуклеаза узнает эту
последовательность и отрезает 10-30
оснований от 3’-конца молекулы премРНК
• Фермент поли(А)-полимераза добавляет
100 – 200 адениловых нуклеотидов к 3’концу мРНК, образуя поли(A) «хвост»
106. Механизмы сплайсинга интронов:
• Тип I - интроны подвергаются аутосплайсингув присутствии только ионов Mg +2 и гуанозина
(пре - рРНК Tetrahymena рhysarum)
• Тип II – интроны подвергаются
аутосплайсингу и имеют концевые
последовательности 5’-GU_ AG-3’ (некоторые
РНК митохондрий у дрожжей)
• Тип III - интроны мРНК, имеющие концевые
последовательности 5’GU_ AG3’,
подвергаются сплайсингу в ядре с участием
мяРНК
107. Сплайсинг интронов типа I
108. Последовательности интронов, необходимые для сплайсинга 5'- GU и 3'- AG
Последовательности интронов, необходимые длясплайсинга 5'- GU и 3'- AG
• Показаны два экзона (красным и зеленым цветом) и один
интрон
• На границе экзон-интрон находятся последовательности GU –
AG
• Для вырезания интронов также необходим сайт ветвления – А
109. Сплайсинг ядерной мРНК происходит в сплайсосоме
• Сплайсосома - специальная ядернаяструктура, в которой происходит сплайсинг
• В состав сплайсосомы входят мяРНК (U1, U2,
U4, U5 и U6) 145 молекул белков
110. Взаимодействие компонентов сплайсосомы с экзонами и интронами РНК
111. Механизмы альтернативного сплайсинга:
• Альтернативный выбор промотора• Альтернативный выбор сигнала
полиаденилирования
• Альтернативный выбор разных наборов
экзонов
• Транс-сплайсинг
112.
113. Альтернативный выбор сигнала полиаденилирования
mРНК кальцитонинового гена умлекопитающих (крыса)
Во всех клетках есть
кальцитониновый ген, но в клетках
щитовидной железы он
экспрессируется в виде гормона
кальцитонина, а в клетках гипофиза
- нейропептида CGRP (пептида,
имеющего отношение к гену
кальцитонина). Ген один, а белки
получаются разные в результате
сплайсинга mРНК и процессинга
полипептидов. В клетках других
тканей этот ген не экспрессируется.
114. Структура мРНК прокариот
• Лидер - это 5’ не транслируемый участок - 5’ UTR (UnTranslatedRegion)
• Трейлер – это 3’ не транслируемый участок (3’UTR)
• Рамка считывания –участок мРНК, кодирующий синтез
полипептида – от старт кодона до стоп-кодона
115. Строение мРНК эукариот
• 5’-кэп- 7метил-гуанозин• Лидер – 5’ нетранслируемый участок - 5’ UTR
(UnTranslated Region)
• Кодирующая последовательность
• Трейлер – 3’ нетранслируемый участок (3’UTR)
• 3’-поли(А)-фрагмент