




























")













 и транспортируемые по флоэме (В)")









 biology
biologySimilar presentations:




")





Водный обмен
1.
2. Водный обмен растений
1.Водная среда объединяет все части организма. В теле растения
водная фаза представляет собой непрерывную среду.
2. Вода – важнейший растворитель и важнейшая среда для протекания
биохимических реакций
3. Вода участвует в упорядочивании структур в клетках
4. Вода – метаболит и непосредственный компонент биохимических
реакций
5. Вода – главный компонент в транспортной системе высших
растений
6. Вода – терморегулирующий фактор
7. Вода амортизатор при механических воздействиях
8. Вода обеспечивает упругость растительных тканей и рост
растяжением
3.
Водичка в растительной клеткеСвободная
85 – 90%
3/4
1/4
1/20
Связанная
10 - 15%
подвижность
Констутивная
(химически связанная)
Гидратационная:
-коллоидносвязанная
(иммобилизованная)
-осмотически связанная
ЯМР
вакуоль
КС
цитозоль
КС 50%
Подвижная и малоподвижнная
Хл 50%
Мх 50%
95% коллоидносвязанная
Вакуоль 98%
Соли, сахара,
Орг. кислоты
Аминокислоты
Белки
Таннины
4.
РастенияПойкилогидрические
Гомойогидрические
Гигрофиты
Мезофиты
Ксерофиты
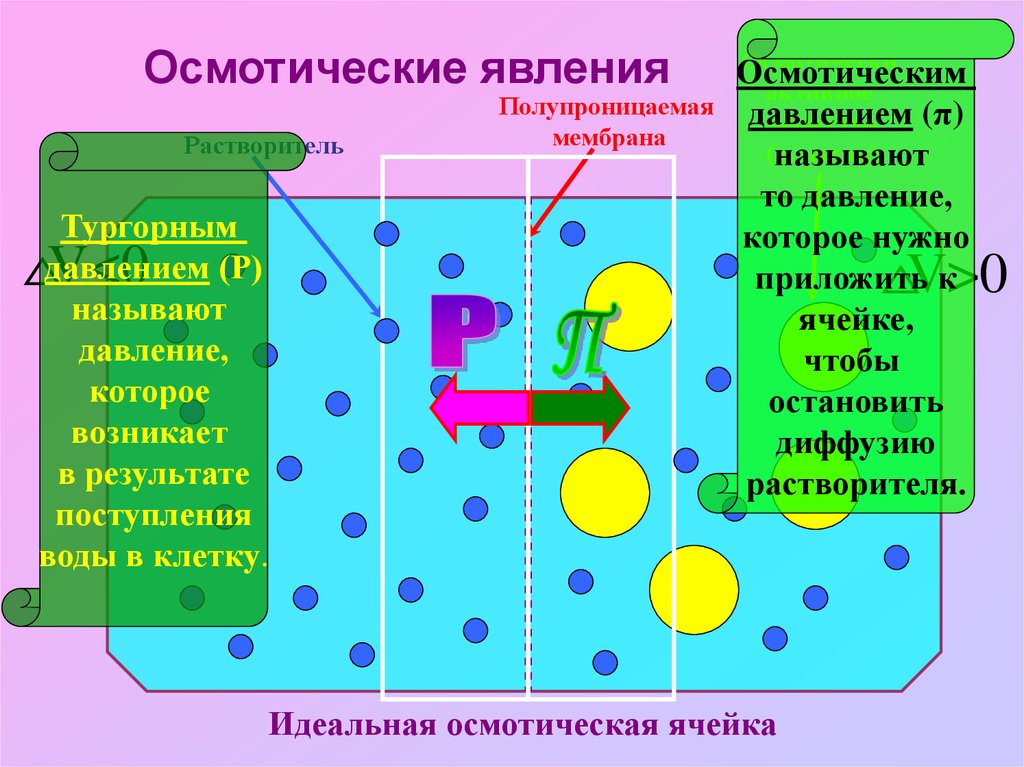
5.
Осмотические явленияРастворитель
Тургорным
давлением (Р)
называют
давление,
которое
возникает
в результате
поступления
воды в клетку.
V<0
Полупроницаемая
мембрана
Осмотически
Осмотическим
активное
давлением
(π)
вещество
(осмотик)
называют
то давление,
которое нужно
приложить к
ячейке,
чтобы
остановить
диффузию
растворителя.
Идеальная осмотическая ячейка
V>0
6.
Осмотические явленияРастворитель
Биополимер
(не образует
Полупроницаемая истинного раствора )
мембрана
Матричная
компонента
осмотического
давления
возникает
в результате
взаимодействия
воды с
гидрофильными
группами
полимеров.
Активность
воды
уменьшается. Идеальная осмотическая ячейка:
взаимодействие растворителя с веществом
7. Осмотическое давление и проблема «лишней» воды
В клетке ψ < 0,в наружной среде ψ ~ 0
Водичка идет по
градиенту…
Осмотическое
давление π избыточное давление,
которое надо приложить
к раствору вещества,
чтобы привести его в
равновесие с чистым
растворителем,
отделенным от него
полупроницаемой
мембраной.
π = RT * ∑ Ci
8. Растительная клетка как осмотическая система
Осмос приводит к возникновению тургорного давления в клетках.Тургорным давлением называется гидростатическое давление, возникающее в
клетках при поглощении ими воды в результате осмоса. Это давление придает
форму и упругость растительным тканям.
Часто используемый в физиологии растений термин “тургор” является качественной
характеристикой клеток, указывающий на наличие в них тургорного давления.
Тонопласт
Клеточная
стенка
Осмотическое
давление
Тургорное
давление
Противодавление
клеточной стенки
Плазмалемма
Противодавление клеточной стенки N равно
гидростатическому или тургорному давлению
N=-P
9.
Термодинамические показатели водыa w - активность воды.
Эффективная (реальная) концентрация, соответственно которой вода
участвует в различных процессах,
для чистой воды = 1;
в растворе или клетке < 1
m w - химический потенциал воды.
Выражает максимальное количество внутренней энергии молекул воды,
которая может быть превращена в работу
Y - водный потенциал
Выражает способность воды в данной системе совершить работу в
сравнении с той работой, которую совершила бы чистая вода при тех же
условиях
10. Итак, основные показатели водного баланса клетки, количественно.
Активность воды:аН О = pН О / poН О
2
2
аН О ≤ 1
2
2
pН2О и
– давление пара воды над системой и
над чистой водой соответственно
poН2О
Химический потенциал воды:
µН О = RT ln аН О
2
2
Водный потенциал: ΨН2О = (µН2О - µоН2О) / V [джоуль/см3]
где V – парциальный мольный объем воды
µН О ≤ 0
2
ΨН О ≤ 0
2
Водный потенциал клетки:
Ψкл = Ψs + Ψp + Ψm + Ψg
Где: Ψs – осмотический потенциал, влияние на Ψ растворенных веществ
Ψp – потенциал давления, влияние на Ψ механического давления
Ψm – матричный потенциал, влияние на Ψ связанной
макромолекулами воды
Ψg – гравитационный потенциал, влияние на Ψ силы тяжести
Ψs < 0; Ψm < 0; Ψp > 0 (как правило); Ψg – для высоких деревьев
Из старых классических учебников: «Сосущая сила клетки» S =
Ψкл
11. Тургор обеспечивает рост клетки «растяжением»
При Yw внешней среды > Yw внутренней среды клеткивода поступает в клетку.
Происходит неупругое растяжение клеточной стенки.
12. Плазмолиз
Yw внешней среды ≤ Yw внутренней среды клеткивода выходит из клетки
Клетка теряет тургор, а затем происходит плазмолиз
13. Водный баланс клетки
Ψкл = [ Ψs + Ψm] + ΨpΨ
Ψкл = Ψнар. р-ра
Ψр
Ψнар. р-ра
0
Ψкл
Ψs + Ψm
14.
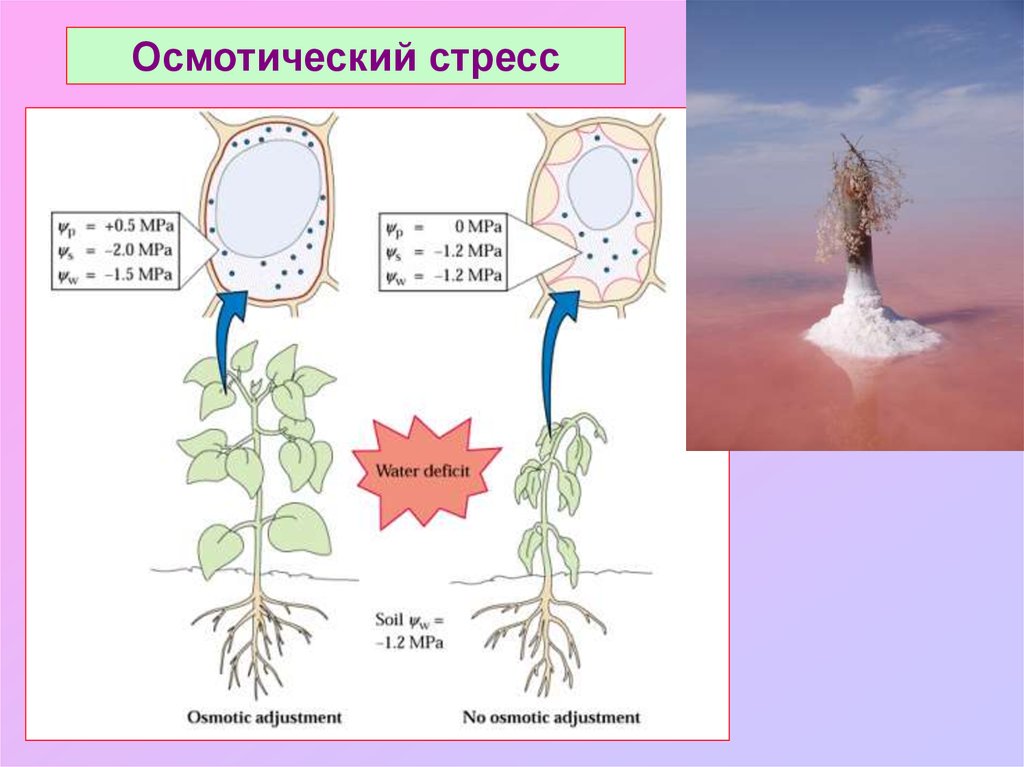
Осмотический стресс15.
4. Понижение1. Понижение
матричного
потенциалас осмотическим
Как бороться
стрессом?
осмотического
потенциала
за счет увеличения числа
за счет синтеза осмолитов
свободных –COOH групп
(глицин-бетаин,
(деметилирование,
Пролин, пролин-бетаин,
изменение доли пектина,
холинсульфат,
окислительные реакции)
маннитол, и др.)
2. Понижение
осмотического потенциала
за счет накопления ионов
(K+, Na+, Cl-, малат и др.
органические кислоты)
3. Понижение
матричного потенциала
за счет синтеза
гидрофильных белков
(осмотины, Lea-белки)
16.
Растения-галофитыSalicornia
Suaeda
17.
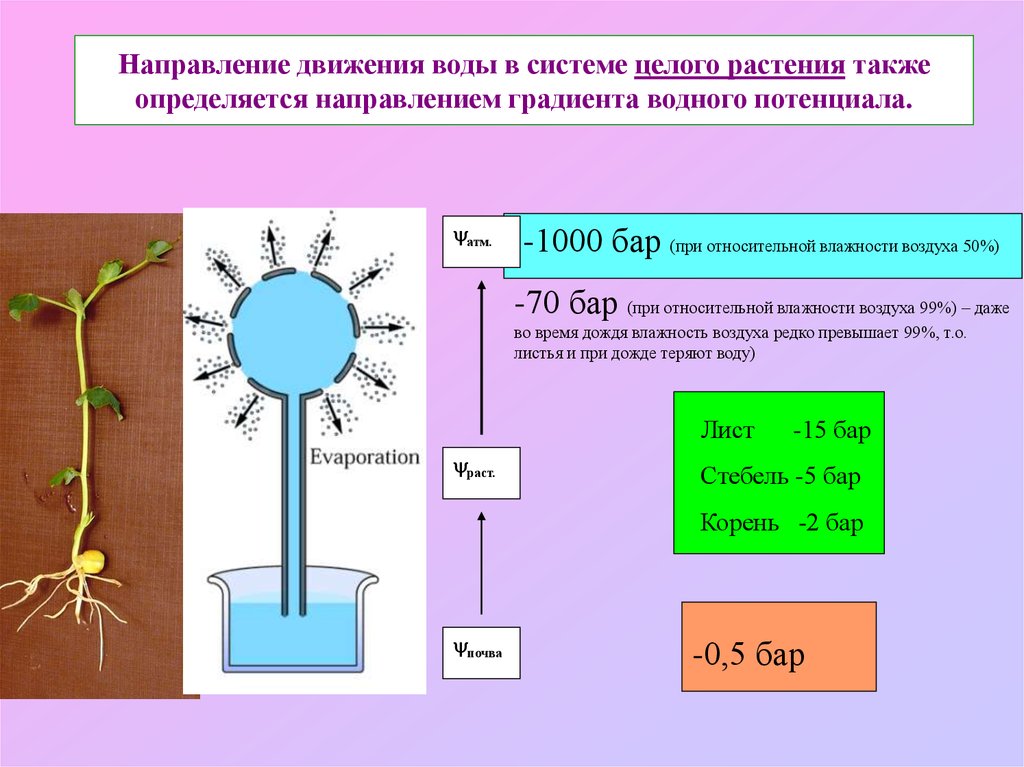
Направление движения воды в системе целого растения такжеопределяется направлением градиента водного потенциала.
атм.
-1000 бар (при относительной влажности воздуха 50%)
-70 бар (при относительной влажности воздуха 99%) – даже
во время дождя влажность воздуха редко превышает 99%, т.о.
листья и при дожде теряют воду)
Лист
раст.
-15 бар
Стебель -5 бар
Корень -2 бар
почва
-0,5 бар
18. Колебания водного потенциала в растительных тканях
Днем водный потенциал клеток обычно снижается,но для разных тканей и органов в разной степени..
19. Динамические характеристики потока воды в клетку Почти закон Ома – I = U/R…
Jw = Lр Y = Lр (Yсреды - Yклетки)R = 1/L p
Jw - объемный поток воды,· выражаемый в см3·см-2·с-1 или см·с-1;
Lр – коэффициент гидравлической проводимости, см·c-1·бар-1;
R – сопротивление потоку воды
Y - градиент водного потенциала
20. Динамические характеристики потока воды в клетку
Jw = Lр Y = Lр (Yср - Yкл)Lр зависит от двух
процессов:
1. Диффузии через
липидный бислой;
2. Проникновения через
специальные белки аквапорины
Повышается при изменении
состава мембран:
Повышение доли
ненасыщенных
жирных кислот (десатурация) ,
стероидов, изопреноидов,
появление окисленных
производных и др.
Изменение числа аквапоринов и
их активности (зависит от
фосфорилирования)
21. Аквапорины
Молекулы воды преодолеваютмембраны двумя путями: через
липидный бислой и через поры,
образуемые специфическими
белками – аквапоринами.
Регулирование:
• Изменение числа каналов для
осмотически активных веществ.
• Изменение активности каналов
(зависит от фосфорилирования,
мембранного потенциала, сигналов и др.)
• Изменение типа каналов.
• Изменение расположения каналов на
мембране.
Аквапорины ПМ
входят в семейство
интегральных белков
PIP (plasma
membrane intrinsic
protein),
Аквапорины
тонопласта
входят в семейство
интегральных белков
TIP (tonoplast intrinsic
protein).
22. Структура водных каналов - аквапоринов
NPA-бокс: Asp-Pro-Ala23. Регулирование аквапоринами водного баланса клетки и растения
много воды вклетке
мало воды в
клетке
Арабидопсис с введенным
геном аквапорина PIP1a и
PIP1b в антисенс
конфигурации.
У трансгена в 5 раз больше
корневая система.
24.
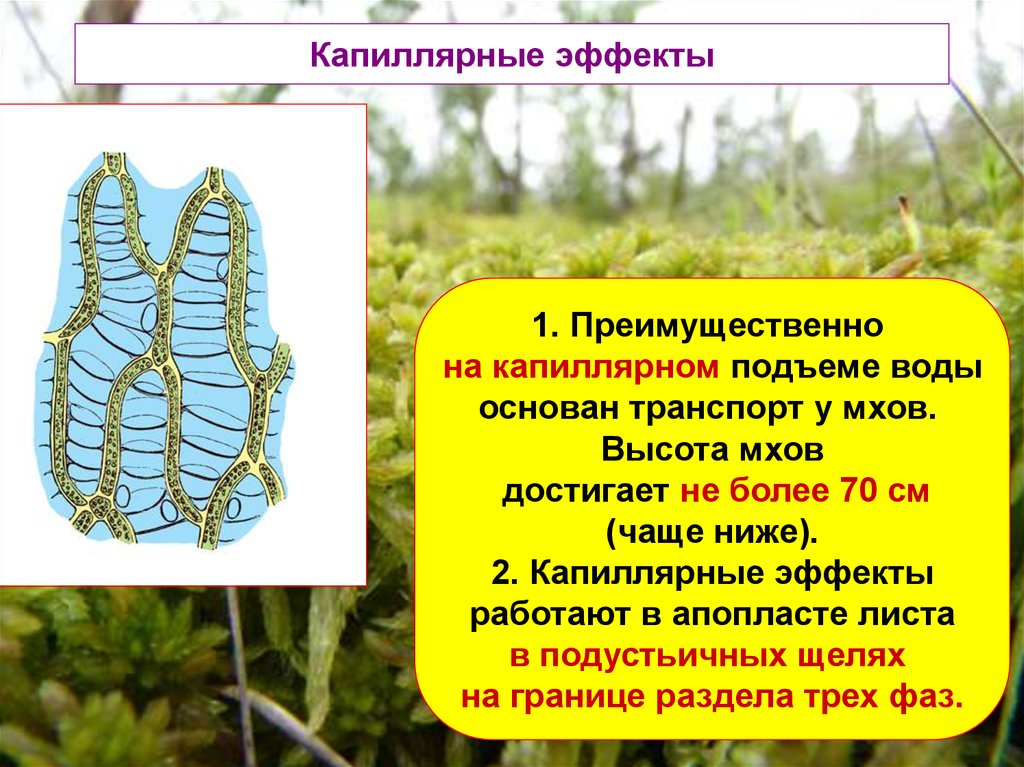
Капиллярные эффекты1. Преимущественно
на капиллярном подъеме воды
основан транспорт у мхов.
Высота мхов
достигает не более 70 см
(чаще ниже).
2. Капиллярные эффекты
работают в апопласте листа
в подустьичных щелях
на границе раздела трех фаз.
25. Поглощение воды корнем
Поздняяметаксилема
Поглощение воды корнем
Ароморфоз:
Появление корней
Ризодерма
с корневыми
«коллективное» выполнение
волосками
осмотических функций.
Нижний концевой двигатель
Кора
Древовидные папоротники
достигали 40 метров в высоту
Эндодерма
с поясками
Каспари
Протоксилема
Ранняя
метаксилема
Перицикл
Флоэма
26. Радиальное строение корня
Ризодерма: первичная загрузкасимпласта, локально –
в трихобластах.
Кора: «метаболический котел»:
1. Реакции ассимиляции
2. Буферная емкость (вакуоли)
Эндодерма: двойной барьер:
а) диффузный – контроль потока
ионов в центральный цилиндр
по апопласту
б) осмотический - барьер
для центробежного потока
ионов и воды из центрального
цилиндра.
Эндодерма отделяет
низкосолевое пространство
(ризодерма и кора) от
высокосолевого (ц. цилиндр ).
Перицикл: кольцевой коллектор,
Собирает ионы и направляет их
к ксилеме.
Ксилема: финиш радиального и
старт дальнего тр-та ионов.
27. Транспортные системы клеток корня
Загрузка ризодермыидет за счет работы
высокоаффинных
переносчиков.
Загрузка ксилемы
обычно идет пассивно
(через каналы),
но за счет работы
Н+-АТФ-аз
28.
Ароморфоз:Образования трахей (сосудов ксилемы):
повышение гидравлической проводимости,
увеличение скорости транспорта.
Увеличение роли
«верхнего концевого двигателя»
Максимальная известная высота деревьев – 116 м..
29.
Верхний эпидермисКУТИКУЛА
(экстрацеллюлярный
слой восковых
веществ)
Столбчатый
мезофилл
Губчатый
мезофилл
УСТЬИЦЕ
Нижний эпидермис
30.
Преобладающие компоненты:сложные эфиры неразветвленных
алкан-1-олов (четные С24-С28)
и алкан-1-оевых кислот (четные С20 – С24)
КУТИКУЛА
(экстрацеллюлярный
слой восковых
веществ)
Примеси и минорные компоненты:
свободные неразветвленные
алкановые спирты, альдегиды, кислоты (четные С20-С28),
иногда – соответствующие кетоны.
Часто алканы (нечетные С25 – С31)
31.
Hosta ‘Love Pat’Микроструктура воска
придает
«пластмассовый»
блеск и устойчивость
к болезням
Hosta ‘Queen Josephine’
32. УСТЬИЦА – ОСОБЫЕ КЛЕТКИ ЭПИДЕРМИСА
1. СОДЕРЖАТ ХЛОРОПЛАСТЫ2. НЕТ ПЛАЗМОДЕСМ
33.
замыкающие клетки устьицаклеточная стенка
более толстая
(ДВУДОЛЬНЫЕ)
Диаметр устьичных щелей 3-12 мкм
расположены (у 75% видов) на нижней
стороне листа
Состояние устьиц зависит от:
• Водного потенциала клеток листа;
• Концентрации СО2 в подъустьичной щели;
• Циркадного (суточного) ритма;
• Фоторецепторов (фототропины и фитохромы)
• Абсцизовой кислоты (АБК) – гормона растений,
вызывающего быстрое закрытие устьиц;
34. ОБРАЗОВАНИЕ МАЛАТА В КЛЕТКАХ УСТЬИЦ (С-3!)
ДЕНЬ:ГЛИКОЛИЗ
КРАХМАЛ
ГЛЮКОЗА
ФЕП
ФЕП-К
ФЕП + НСО3-
МДГ
ОАА
МАЛАТ
Фн
НАДФН
НОЧЬ:
МАЛИК- ЭНЗИМ
МАЛАТ
НАД+
СО2
НАДН
НАДФ+
ГЛЮКОНЕОГЕНЕЗ
ПИРУВАТ
ГЛЮКОЗА
КРАХМАЛ
35.
Открывание устьиц ( 2-3 часа)К+in-канал
+ + + Н+
Н+ Cl
К+
АТФ
Н2 О
Cl-in
К+in
АДФ
К+
Cl-
Н2О
малат
АТФ
АДФ
Синий свет, ИУК, низкие конц.
СО2 –факторы открывания
Н+
36.
Закрывание устьиц (30 мин)Cl-out
К+out
Н2О
Са2+
К+out
Высокая конц. СО2, АБК- (образуется
при водном дефиците), снижение
водного потенциала клеток листафакторы закрывания
Cl-out
малат
мтх
Ночь:
малат
СО2
НАДН
ПК
глюкоза
крахмал
37.
ГуттацияКаждый лист таро (Colocasia esculenta)
в течение суток выделяет до 100 мл воды,
причем интенсивность гуттации
достигает
38.
ГуттацияРеабсорбция минеральных веществ
39.
Лист: ВЕРХНИЙ КОНЦЕВОЙДВИГАТЕЛЬ водного потока.
1. Транспирация (контакт с
атмосферой)
2. Фиксация СО2 (образование
органических осмотиков)
3. Гуттация
4. Рециркуляция воды (активный
отток по флоэме)
5. Рост молодого листа
Корень: НИЖНИЙ КОНЦЕВОЙ
ДВИГАТЕЛЬ водного потока.
1. Создание корневого давления
(загрузка ксилемы)
2. Рециркуляция воды (активный
обмен ксилемы с флоэмой)
3. Рост молодого корня
40. Радиальное строение корня и механизм возникновения корневого давления
Ризодерма: первичная загрузкасимпласта, локально –
в трихобластах.
Кора: «метаболический котел»:
1. Реакции ассимиляции
2. Буферная емкость (вакуоли)
Эндодерма: двойной барьер:
а) диффузный – контроль потока
ионов в центральный цилиндр
по апопласту
б) осмотический - барьер
для центробежного потока
ионов и воды из центрального
цилиндра.
Эндодерма отделяет
низкосолевое пространство
(ризодерма и кора) от
высокосолевого (ц. цилиндр ).
Перицикл: кольцевой коллектор,
Собирает ионы и направляет их
к ксилеме.
Ксилема: финиш радиального и
старт дальнего тр-та ионов.
41.
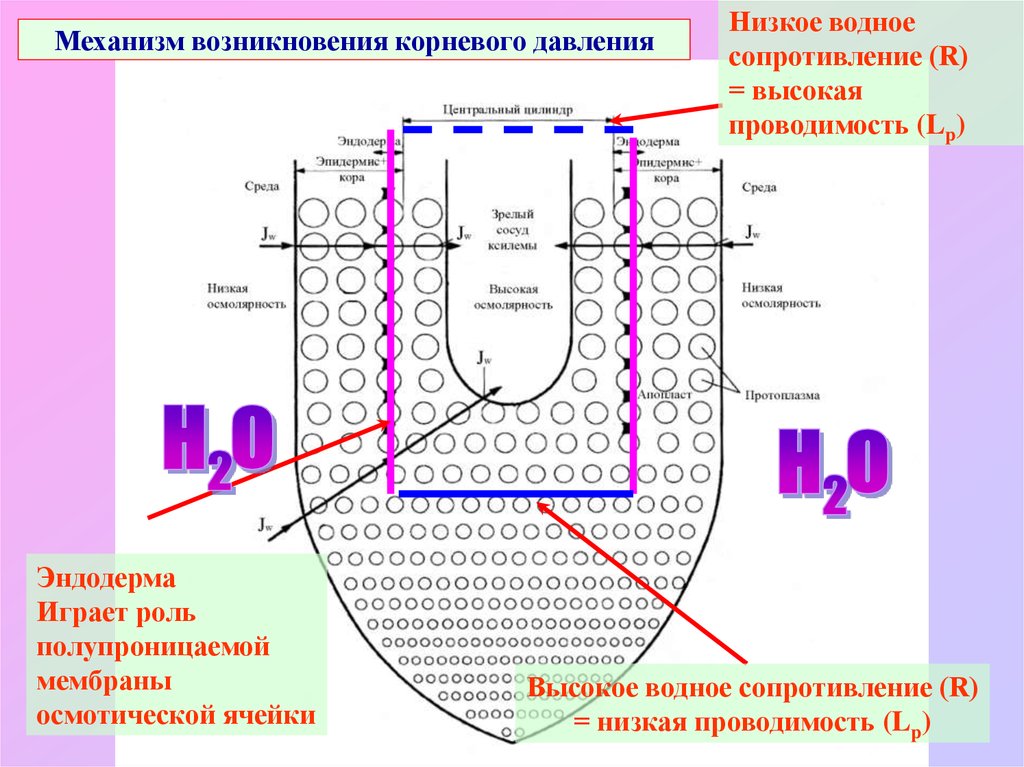
Механизм возникновения корневого давленияЭндодерма
Играет роль
полупроницаемой
мембраны
осмотической ячейки
Низкое водное
сопротивление (R)
= высокая
проводимость (Lp)
Высокое водное сопротивление (R)
= низкая проводимость (Lp)
42. Радиальный транспорт воды в корне
43. Варианты транспортных путей в корне
СимпластныйТрансцеллюлярный (от клетки к клетке)
Апопластный
В зависимости от внешних условий и вида
растений могут преобладать разные пути
транспорта. При интенсивной транспирации
вода обычно передвигается по апопласту
Апопластный транспорт воды быстрее
(~ в 50 раз) симпластного.
Для гороха, ячменя, фасоли вода
передвигается от клетки к клетке при любых
условиях, для кукурузы – по апопласту.
По апопласту обычно передвигаются Ca2+ и
бор. По симпласту – фосфаты.
44.
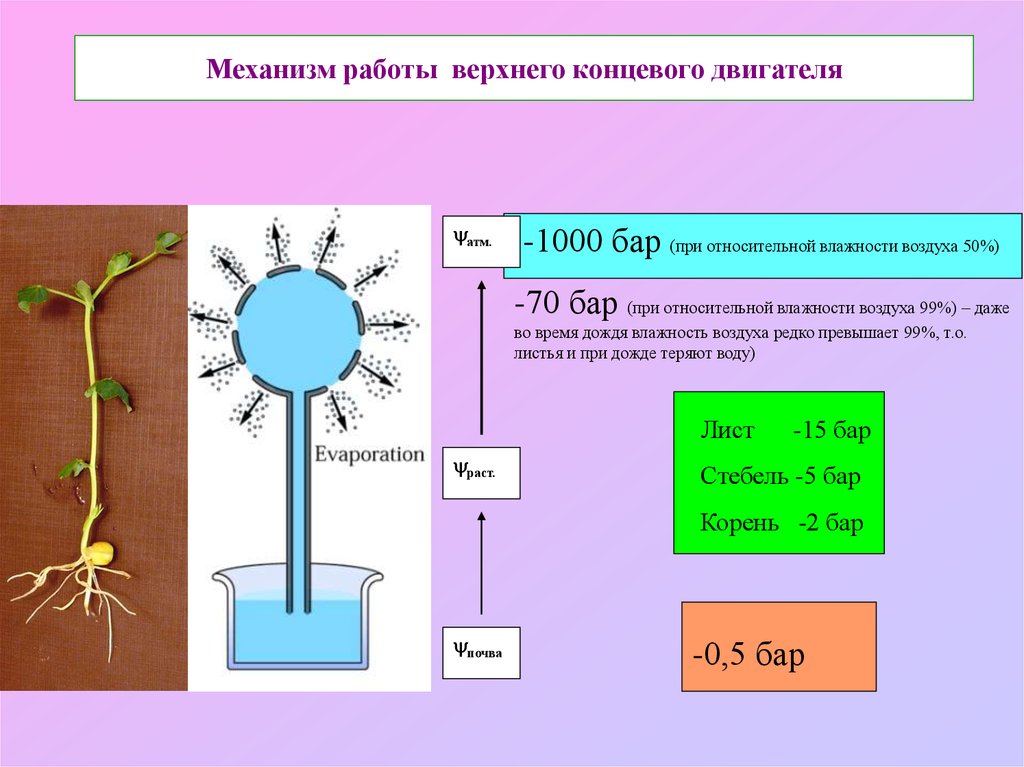
Механизм работы верхнего концевого двигателяатм.
-1000 бар (при относительной влажности воздуха 50%)
-70 бар (при относительной влажности воздуха 99%) – даже
во время дождя влажность воздуха редко превышает 99%, т.о.
листья и при дожде теряют воду)
Лист
раст.
-15 бар
Стебель -5 бар
Корень -2 бар
почва
-0,5 бар
45. Схема флоэмного и ксилемного транспорта
46. Взаимосвязь ксилемного и флоэмного транспорта
47. Строение флоэмы
48.
Афиднаяметодика
(от Aphid –
тля)
49. Состав флоэмного и ксилемного экссудата
Компонент илисвойство
Флоэма
Ксилема
100 –
300 г/л
0
5 – 40 г/л
0,1 – 2 г/л
1 – 5 г/л
0,2 – 4 г/л
Всего
растворено
250 –
1200 mM
π = 0,6 –
3,0 MПa
0,02 –
0,2 mM
π = 0.02 –
0,2 MПa
рН
7.3 – 8.0
5.0 – 6.5
Сахара
Аминокислоты
(glu, asp, gln, аsn)
Неорганические
соли
50. Вещества, не транспортируемые (А) и транспортируемые по флоэме (В)
Не транспортируются:Углеводы с альдегидной
или кетонной группой
(глюкоза, манноза,
фруктоза…)
Транспортируются:
Сахароза – чаще всего;
У некоторых видов:
Галактопроизводные
(раффиноза, стахиоза) тыквенные
Сахароспирты (маннит,
сорбит) – розоцветные
Уреиды – у видов с
симбиотичесой фиксацией
азота.
Кроме того, короткие РНК,
сигнальные молекулы
(системин и др).
51. Клетки-спутницы бывают трех типов. Зачем?
A – обычные и промежуточныеклетки-спутницы у Mimulus cardinalis
В – трансферные клетки у Pisum
sativum
С - промежуточные клетки у Alonsoa
warscewicziij с множеством
плазмодесм
52. Симпластная и апопластная загрузка флоэмы
53.
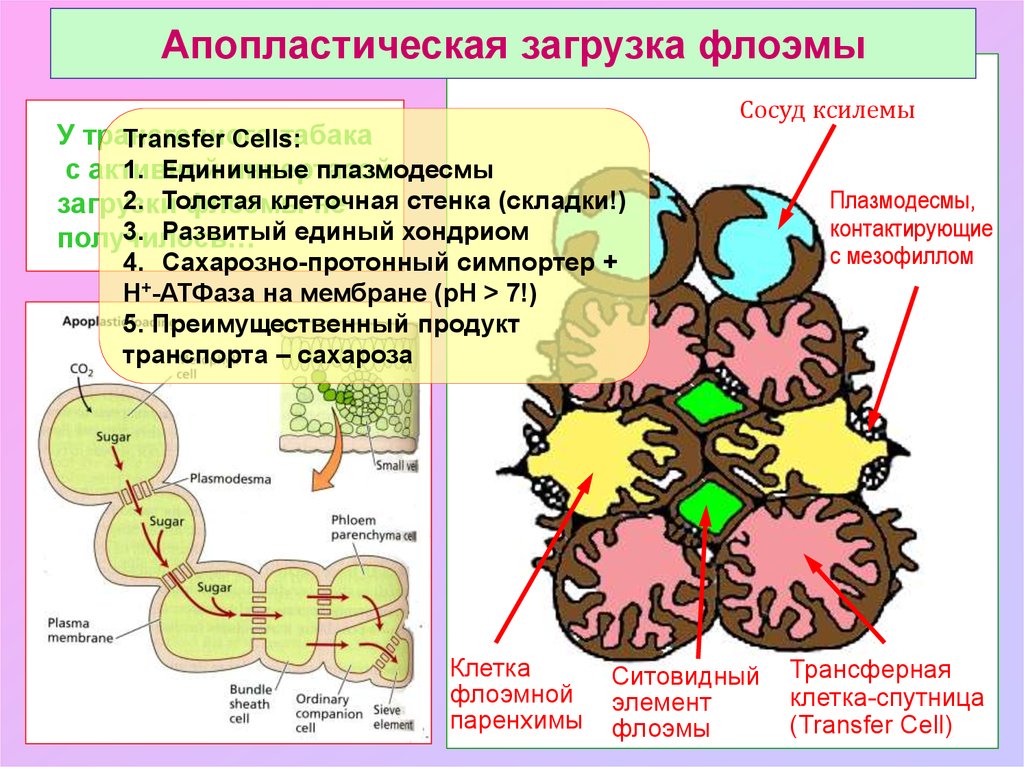
Апопластическая загрузка флоэмыУ трансгенного
табака
Transfer Cells:
1. Единичные
плазмодесмы
с активной
инвертазой
2. Толстая
клеточная
стенка (складки!)
загрузки
флоэмы
не
3. Развитый единый хондриом
получилось…
Сосуд ксилемы
4. Сахарозно-протонный симпортер +
Н+-АТФаза на мембране (рН > 7!)
5. Преимущественный продукт
транспорта – сахароза
Клетка
флоэмной
паренхимы
Ситовидный
элемент
флоэмы
Плазмодесмы,
контактирующие
с мезофиллом
Трансферная
клетка-спутница
(Transfer Cell)
54. Симпластическая загрузка флоэмы
Intermediary Cells:1. Обильные плазмодесменные поля
2. Толстая клеточная стенка (ровная!)
3. Развитый единый хондриом
4. Специфические ферменты синтеза
5. Разнообразные продукты
транспорта фотоассимилятов (смесь!)
Ситовидный элемент
флоэмы
Сосуд ксилемы
Плазмодесмы,
контактирующие
с мезофиллом
Промежуточная
клетка-спутница
(Intermediary Cell)
55.
Alonsoa meridionalis(Scrophulariaceae)
Вещества, содержащиеся во флоэмном экссудате
Alonsoa meridionalis, mM (C.Knop, 1998)
Сахароза
174 85
Галактинол
traces
Мезо-инозитол
traces
Раффиноза
Стахиоза
249 51
397 68
Морфология проводящей системы листа Alonsoa
Промежуточные клетки
со множеством
плазмодесм –
симпластная
загрузка
56. Роль промежуточных клеток в симпластной загрузке флоэмы: «полимеризационная модель»
Mesophyll CellSieve Element
IC
raffinose
raffinose
stachyose
stachyose
sucrose
galactinol
SEL:
SEL:
<0.7 kDa
1-3 kDa
1. Синтез сахарозы и галактинола в мезофилле
2. Передвижение сахарозы и галактинола в промежуточные клетки
через плазмодесмы
3. Синтез стахиозы и раффинозы в промежуточных клетках
4. Невозможность обратного перемещения стахиозы и раффинозы в
мезофилл (из-за размера молекул и особенностей плазмодесм)
57. Промежуточные клетки всегда присутствуют в листьях, содержащих стахиозу, и наоборот
СемействаКлетки-спутницы
Наличие
стахиозы
Bignoniaceae: Catalpa bignonioides
IC (промежуточные клетки)
Стахиоза и др.
Buddlejaceae: Buddleja davidii
IC (промежуточные клетки)
Стахиоза и др.
Celastraceae (4)
IC (промежуточные клетки)
Стахиоза и др.
Симпластическая загрузка флоэмы
Cucurbitaceae (5)
IC (промежуточные клетки)
Стахиоза и др.
Lamiaceae (2)
IC (промежуточные клетки)
Стахиоза и др.
Oleaceae (2)
IC (промежуточные клетки)
Стахиоза и др.
Onagraceae (3)
IC (промежуточные клетки)
Стахиоза и др.
Verbenaceae: Verbena hybrida
IC (промежуточные клетки)
Стахиоза и др.
OC(обычные клетки)
нет
Araliaceae: Hedera helix
OC(обычные клетки)
нет
Brassicaceae: Arabidopsis thaliana (Columbia)
OC(обычные клетки)
нет
Chenopodiaceae (4)
OC(обычные клетки)
нет
Rosaceae (4)
OC(обычные клетки)
следы стахиозы
Violaceae (2)
OC(обычные клетки)
следы стахиозы
OC,TC (обычные и трансферные
клетки)
нет
Asteraceae (2)
TC (трансферные клетки)
нет
Boraginaceae(3)
TC (трансферные клетки)
нет
Fabaceae (10)
TC (трансферные клетки)
нет
Apiaceae: Petroselinum crispum
Апопластическая загрузка флоэмы
Solanaceae (2)
Апопластическая загрузка флоэмы
58.
Взаимосвязь ксилемного и флоэмного транспортаГетеробарический лист:
1. Разгрузка ксилемы
по апопласту
2. Загрузка флоэмы
по симпласту
Изобарический лист:
1. Разгрузка ксилемы
по симпласту
2. Загрузка флоэмы
по апопласту
59.
Способы загрузки и разгрузки ксилемы и флоэмы зависит от многихфакторов – жизненных форм, условий существования, типа органов