





















 biology
biologySimilar presentations:










Матричный принцип, как основа современной эволюционной теории
1. Матричный принцип, как основа современной эволюционной теории
2. Матричный принцип, как основа современной эволюционной теории
3.
Н.В.ТИМОФЕЕВ-РЕСОВСКИЙ«…то, что конвариантная
редупликация (идентичное
воспроизведение), протекающая
по определенным физическим
законам, все-таки наличествует —
во всяком случае на нашей
планете, а по-видимому, и в
космическом масштабе — лишь в
связи с жизнью, ведет к тому, что
логически вытекающий из
принципа конвариантной
редупликации принцип
естественного отбора является
действительно единственным
всеобщим биологическим
принципом».
4.
Два биологических макропринципа:• принцип отбора
(специфически биологический);
•принцип конвариантной редупликации
(подлежит физическому объяснению).
«… принцип естественного отбора …
возможен только как следствие из
принципов конвариантной редупликации и
изменчивости редуплицируемых структур»
5. Элементарный эволюционный материал в СТЭ
ИзменчивостьНаследственная
Мутационная
Ненаследственная
Комбинативная Репликационная Онтогенетическая
(Эпигенетическая)
6.
• 1909 г В.Иогансен: наследственныефакторы - гены.
• Т. Морган: ген, являющийся частью
хромосомы, мутирует как целостная
структура и контролирует определённый
признак организма.
• Н.В.Тимофеев-Ресовский, М.Дельбрюк,
К.Циммер, А. Серебровский и др.:
с помощью воздействия ионизирующего
излучения на мутационный процесс изучены
размер, протяжённость, структура
и делимость самого гена.
7.
• Дж. Бидл и Э. Тейтем: предположение - одинген кодирует синтез одного фермента.
• Дж. Уотсон и Ф. Крик: идентификация гена
как участка молекулы ДНК, завершившаяся
расшифровкой свойств самого генетического
кода.
• Ф. Жакоб и Ж. Моно: явление генетического
контроля синтеза ферментов у бактерий механизм регуляции активности генов.
• С. Бензер и Ч. Яновский: наименьшей
изменяемой единицей в пределах гена
является пара нуклеотидов.
8.
• Поток информации, записанной в ДНК,идёт от ДНК к информационной РНК,
на основе которой затем синтезируется
белковая молекула.
• Этот поток является дорогой
с односторонним движением:
информация, попавшая в белок, оттуда
не возвращается!
Т.е., не может быть синтезирована
кодирующая последовательность ДНК
на основе белковой последовательности
аминокислот.
9.
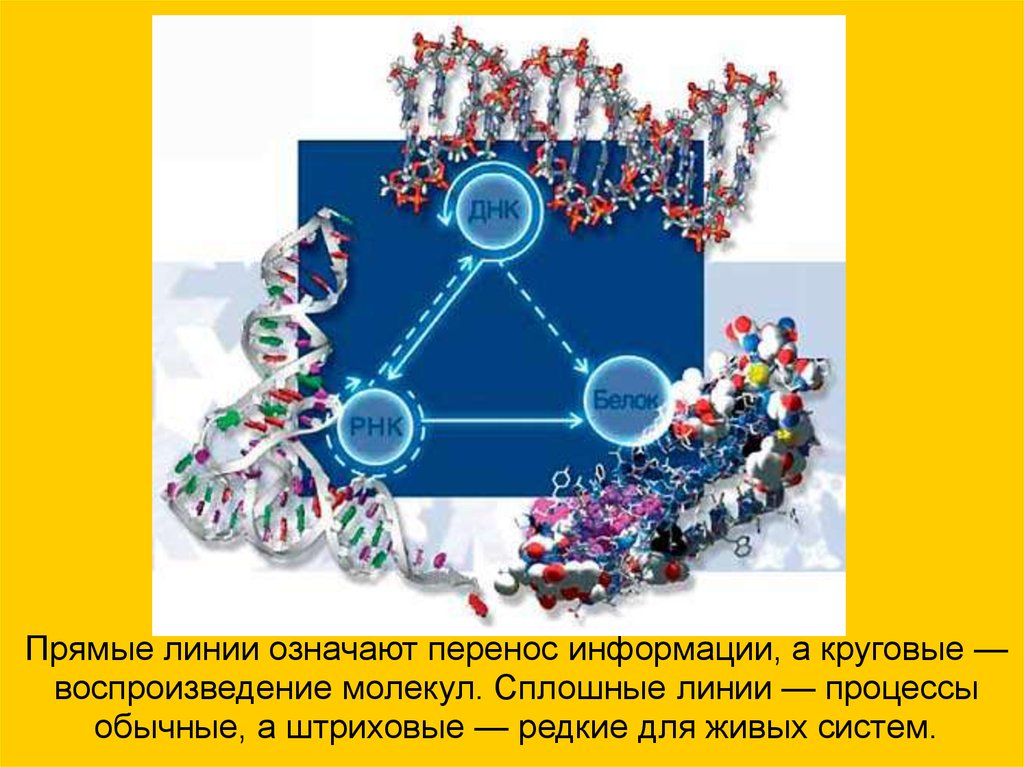
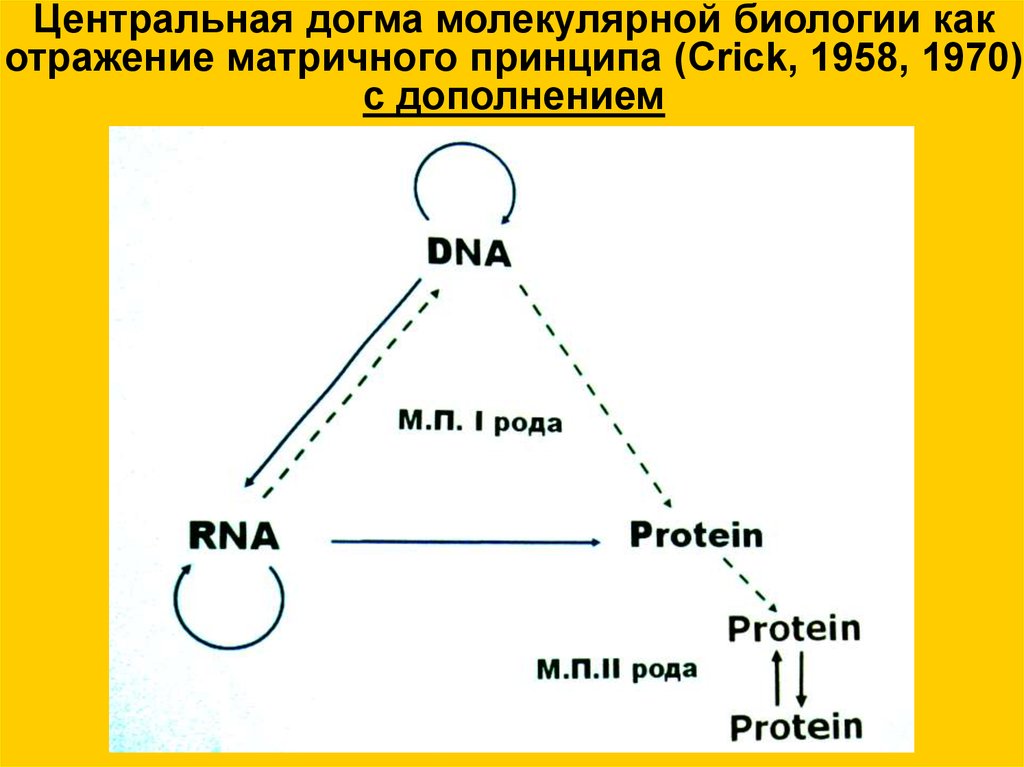
Прямые линии означают перенос информации, а круговые —воспроизведение молекул. Сплошные линии — процессы
обычные, а штриховые — редкие для живых систем.
10.
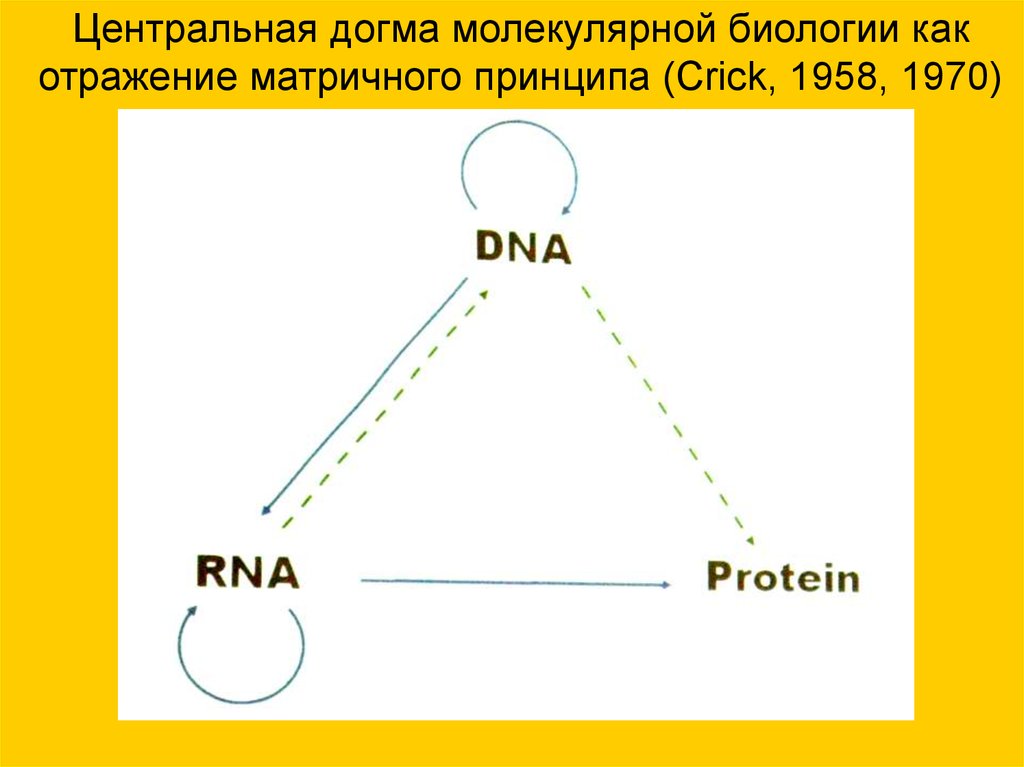
Центральная догма молекулярной биологии какотражение матричного принципа (Crick, 1958, 1970)
11.
«Центральная догма» молекулярнойбиологии:
• Генетическая информация, записанная
в виде последовательности оснований
ДНК, определяет последовательность
оснований дочернего полимера — РНК,
а та — последовательность
аминокислот в белковой цепи.
• Перенос информации возможен только
от ДНК к белкам, но не в обратном
направлении. Фактически „центральная
догма“ — современный (молекулярный)
аналог принципа А.Вейсмана о
ненаследовании приобретённых
признаков (Crick H.F. С, 1958)
12.
Формирование молекулярной парадигмыв 60-е годы прошлого века породило
иллюзию, наиболее ярко отразившуюся
во фразе-каламбуре Жака Моно:
«What is true for E.coli, is true for
E.lephant»
(«Что справедливо для кишечной
палочки, справедливо и для слона»)
Принцип биологической универсальности
дополняется
принципом биологической специфичности
13.
Прокариоты - оперонная организация геномаОперон — группа, имеющая общий регуляторный элемент.
Гены, контролирующие биосинтез аминокислоты гистидина у
бактерии Salmonella typhimurium (А), собраны в оперон,
представляющий собой единицу регуляции. Все эти гены
„переписываются“ на одну молекулу мРНК, которая служит
матрицей для синтеза отдельных белков-ферментов, которые
и осуществляют все 10 этапов биосинтеза гистидина.
14.
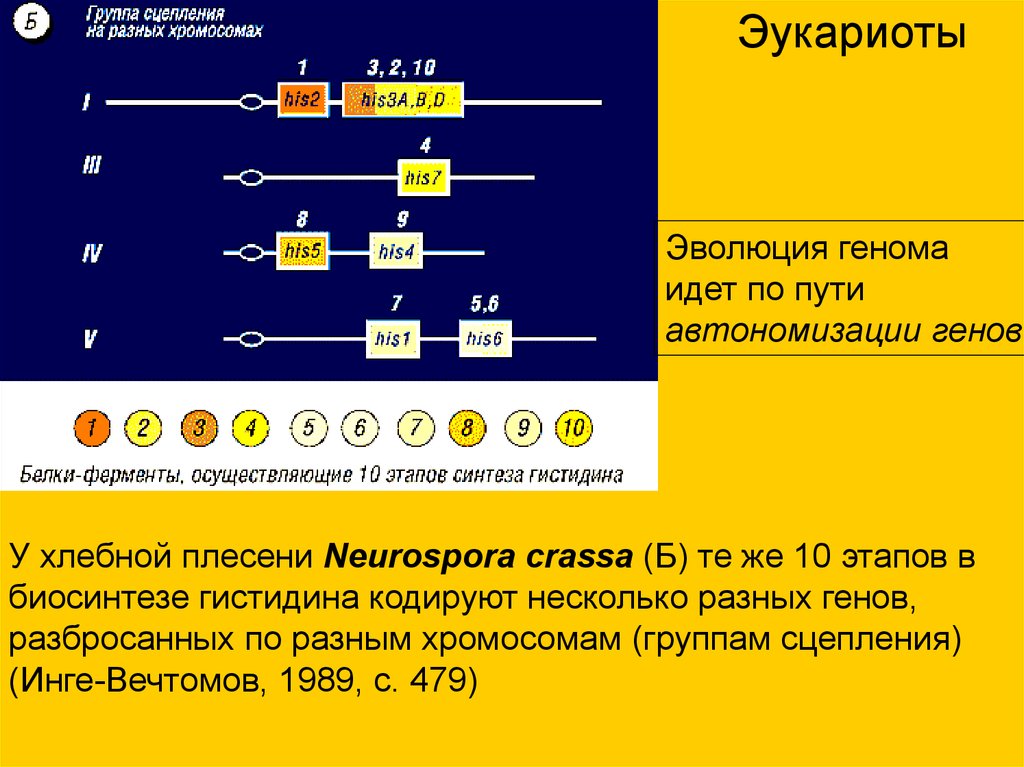
ЭукариотыЭволюция генома
идет по пути
автономизации генов
У хлебной плесени Neurospora crassa (Б) те же 10 этапов в
биосинтезе гистидина кодируют несколько разных генов,
разбросанных по разным хромосомам (группам сцепления)
(Инге-Вечтомов, 1989, с. 479)
15.
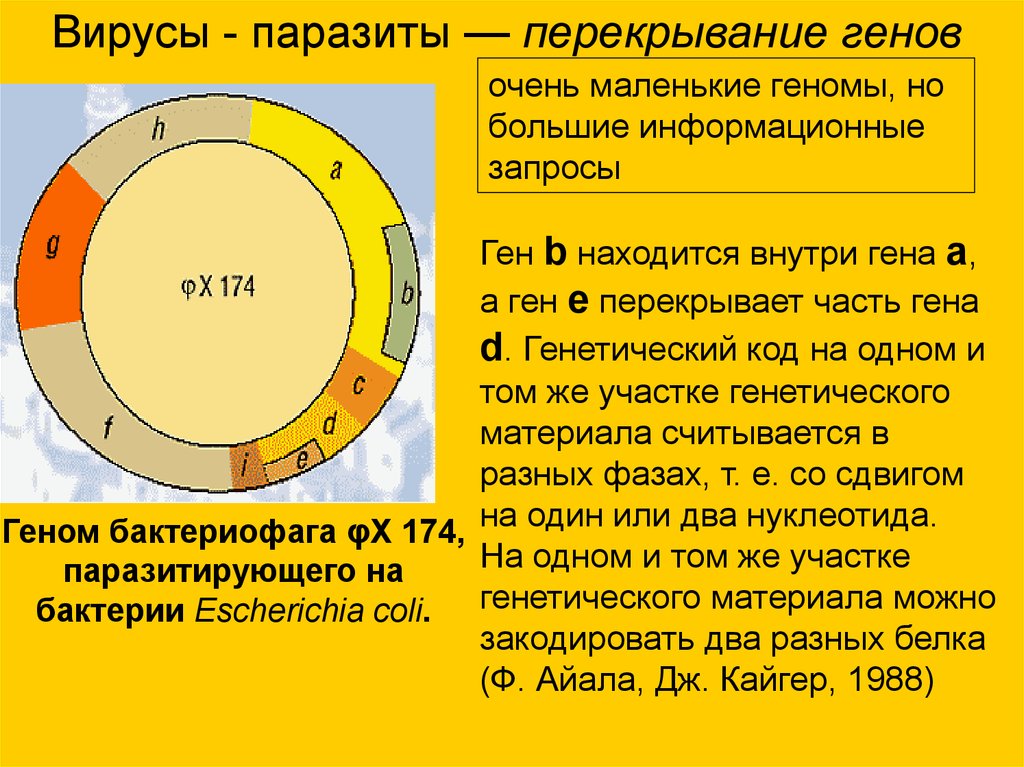
Вирусы - паразиты — перекрывание геновочень маленькие геномы, но
большие информационные
запросы
Ген b находится внутри гена a,
а ген e перекрывает часть гена
d. Генетический код на одном и
том же участке генетического
материала считывается в
разных фазах, т. е. со сдвигом
Геном бактериофага φX 174, на один или два нуклеотида.
На одном и том же участке
паразитирующего на
бактерии Escherichia coli. генетического материала можно
закодировать два разных белка
(Ф. Айала, Дж. Кайгер, 1988)
16.
Тенденции в эволюции генома• Автономизация.
• Олигомеризация.
• Возникновение мозаичной структуры
гена у эукариот (интроны и экзоны).
• Эволюция происходит за счёт
тиражирования (дуплицирования)
существующих генетических структур
и их дальнейшей дивергенции.
Блочный принцип: всё существующее генетическое
разнообразие произошло за счёт тиражирования и
комбинаторики относительно небольшого числа
исходных генетических структур.
17.
Экзоны - участки гена, содержащие кодирующиепоследовательности.
Интроны - участки гена не несущие генетической
информации.
Интроны (Mattick, Gagen, 2001; Fedorova, Fedorov, 2003)
содержат большое разнообразие регуляторных элементов,
влияющих на экспрессию генов — сайты связывания
транскрипционных факторов, энхенсеры, альтернативные
промоутеры, сайты метилирования и т.д. (Kolchanov et al.,
2002).
Т.е., «рыхлый» геном эукариот обеспечивает лучшие условия
для взаимодействия различных кодов, чем «плотный» геном
прокариот.
18.
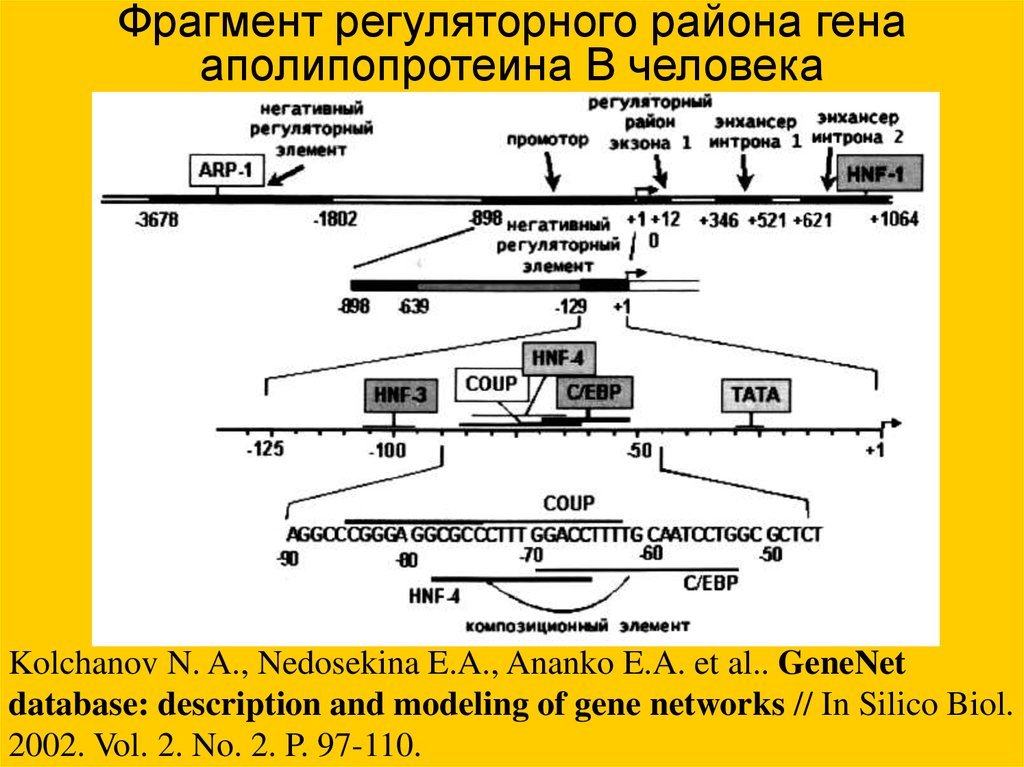
Фрагмент регуляторного района генааполипопротеина B человека
Kolchanov N. A., Nedosekina E.A., Ananko E.A. et al.. GeneNet
database: description and modeling of gene networks // In Silico Biol.
2002. Vol. 2. No. 2. P. 97-110.
19.
Комбинаторика альтернативного сплайсинга гена DSCAMDrosophila melanogaster
Black D.L. Protein diversity from alternative splicing: a challenge for
bioinformatics and post-genome biology // Cell. 2000. V. 103, № 3. P.
367-370.
20.
• Регуляторные районы генов эукариот, по сравнению спрокариотами, очень велики (на порядки больше размера
его кодирующей части).
• При количестве сайтов связывания регуляторных белков
равном-N количество состояний регуляторного района
равно 2N. При N=20 это около миллиона состояний!
• С огромной емкостью регуляторного кода согласуется
незначительная (6—15% от протеома) доля белковрегуляторов транскрипции у разных эукариот (Hermoso et
al., 2004, The Gene..., 2004)
• Блочно-комбинаторный принцип кодирования
генетической информации у эукариот — ароморфоз,
позволивший фактически безгранично наращивать
сложность генетических программ без существенного
роста размеров геномов.
21.
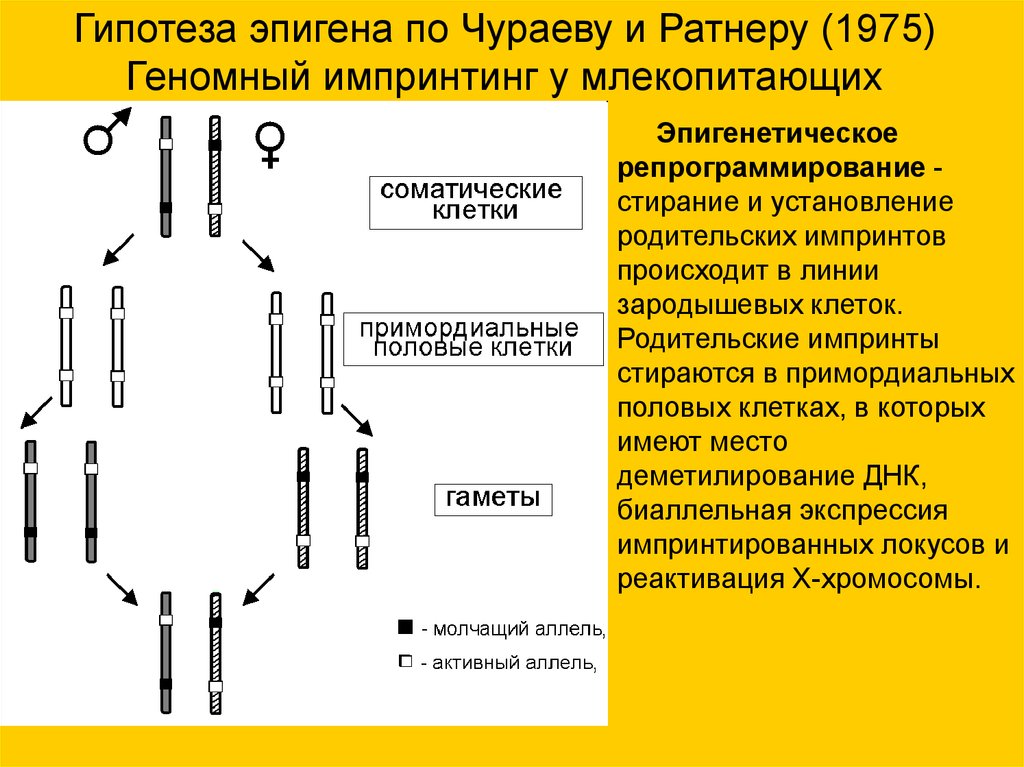
Гипотеза эпигена по Чураеву и Ратнеру (1975)Геномный импринтинг у млекопитающих
Эпигенетическое
репрограммирование стирание и установление
родительских импринтов
происходит в линии
зародышевых клеток.
Родительские импринты
стираются в примордиальных
половых клетках, в которых
имеют место
деметилирование ДНК,
биаллельная экспрессия
импринтированных локусов и
реактивация X-хромосомы.
22.
Спецификация Нох-генами тканей сегментовтела D.melanogaster эпигенетически наследуема и
зависит от белков семейств Polycomb.
У плацентарных млекопитающх белки
семейства Polycomb участвуют также в поддержании
геномного импринтинга, обеспечивающего родительспецифичную инактивацию одной из копий генов
диплоидного организма, предотвращая
партеногенез.
Эпигентическая разметка, в отличие от
генетической регуляции, может стираться под
влиянием среды (например, холодовой импринтинг)
(Kohler, Grossniklaus, 2002; Bastow et al., 2004).
Bastow R., Mylne J.S., Lister C. et al. 2004. Vernalization requires epigenetic
silencing of FLC by histone methylation // Nature. Vol. 427. No. 6970. P. 164-167.
Kohler C., Grossniklaus U. 2002. Epigenetics: the flowers that come in from the
cold // Curr. Biol. Vol. 12. No. 4. P. R129-R131.
23.
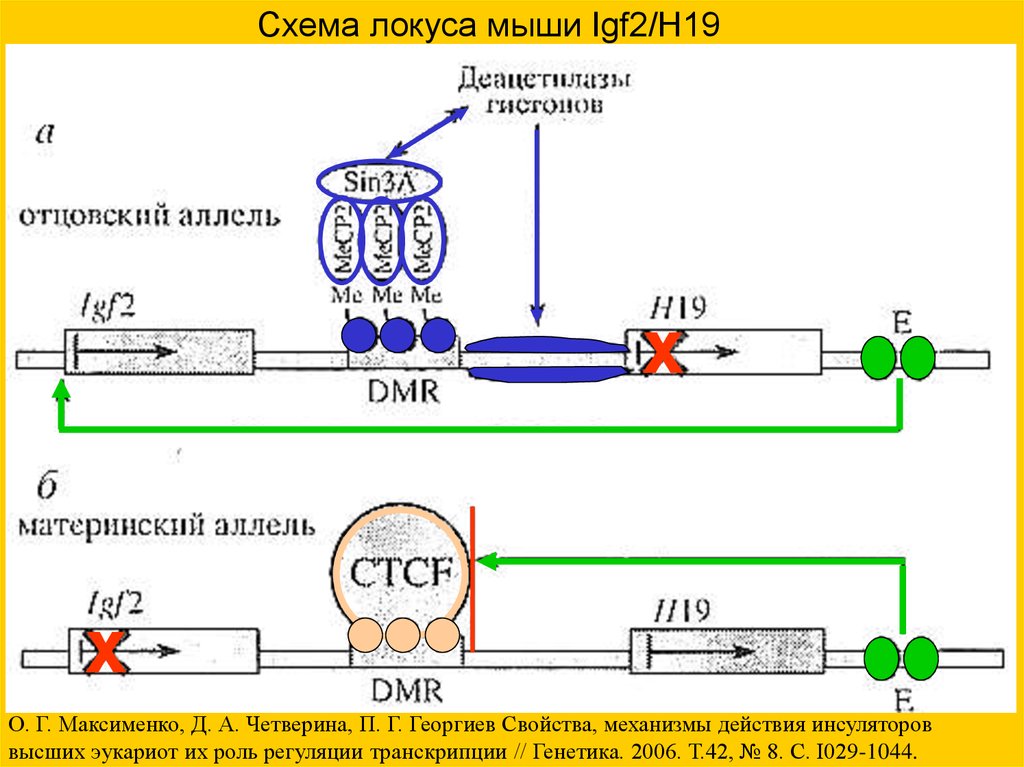
Схема локуса мыши Igf2/H19х
х
О. Г. Максименко, Д. А. Четверина, П. Г. Георгиев Свойства, механизмы действия инсуляторов
высших эукариот их роль регуляции транскрипции // Генетика. 2006. Т.42, № 8. С. I029-1044.
24.
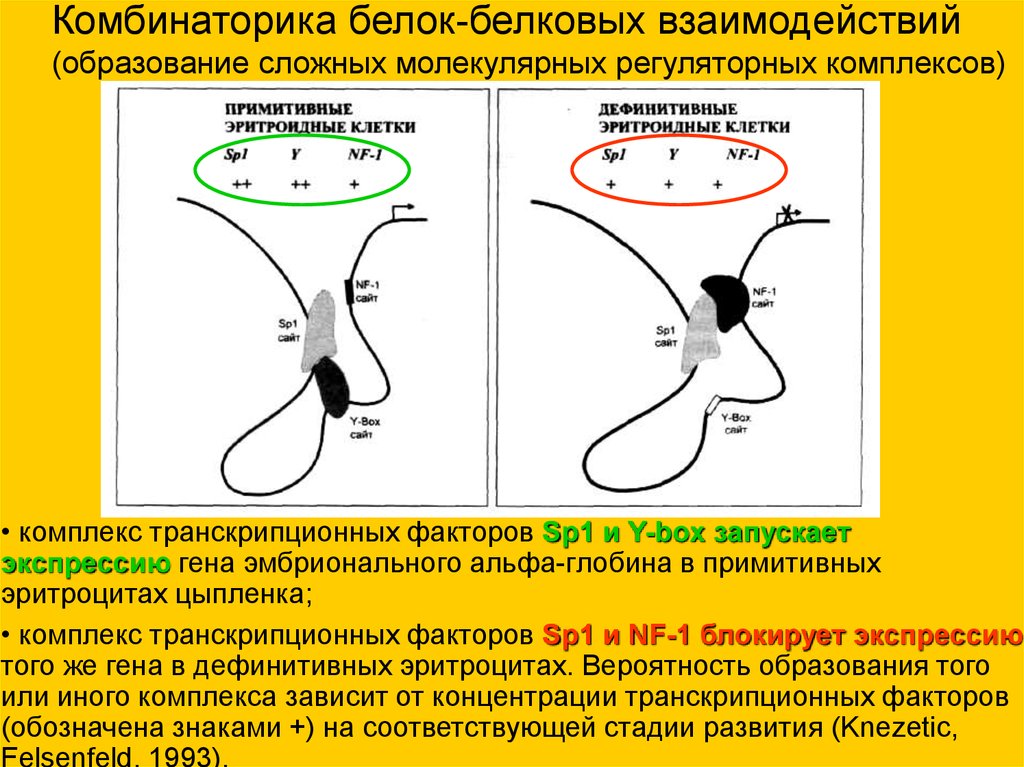
Комбинаторика белок-белковых взаимодействий(образование сложных молекулярных регуляторных комплексов)
• комплекс транскрипционных факторов Sp1 и Y-box запускает
экспрессию гена эмбрионального альфа-глобина в примитивных
эритроцитах цыпленка;
• комплекс транскрипционных факторов Sp1 и NF-1 блокирует экспрессию
того же гена в дефинитивных эритроцитах. Вероятность образования того
или иного комплекса зависит от концентрации транскрипционных факторов
(обозначена знаками +) на соответствующей стадии развития (Knezetic,
Felsenfeld, 1993).
25.
Различие в эпигенетической регуляции само посебе не ведет к эволюции паралогов. Их дивергенция
— всецело следствие накопленных в них мутаций.
Различная эпигенетическая регуляция лишь
нарушает нейтрализацию таких мутаций, время от
времени вводя их в сферу отбора.
Такая концепция согласуется с ранее
высказанной идеей об эволюционной роли
фенотипической супрессии, вызванной прионизацией
(Инге-Вечтомов, 1998, 2000).
Инге-Вечтомов С.Г. Прионы дрожжей и центральная догма
молекулярной биологии // Вестн. РАН. 2000. Т. 70. № 4. С. 299-306.
Инге-Вечтомов С.Г. Общая биология и частная генетика прионов //
Вестник ВОГиС. 1998. Т. 2. № 4.
26.
Прионы — «нестандартные» биологическиерепликаторы, представляющие собой особым
образом свернутые молекулы прионного
белка PrP. Этот белок в норме присутствует
на мембранах нейронов и выполняет какие-то
полезные функции, связанные с передачей
сигналов.
Chiesa R., Harris D.A. Fishing for Prion Protein
Function // PLoS Biol. 2009. 7(3): e1000075).
27. Инфекционные нейрогенеративные заболевания человека и животных:
Куру.
Болезнь Кройцфельда-Якоба.
Синдром Герштона-Штресслера-Шейнкера.
Семейная смертельная бессонница.
Скрэпи овец и коз.
Губчатая болезнь мозга крупного рогатого
скота («коровье бешенство»).
28.
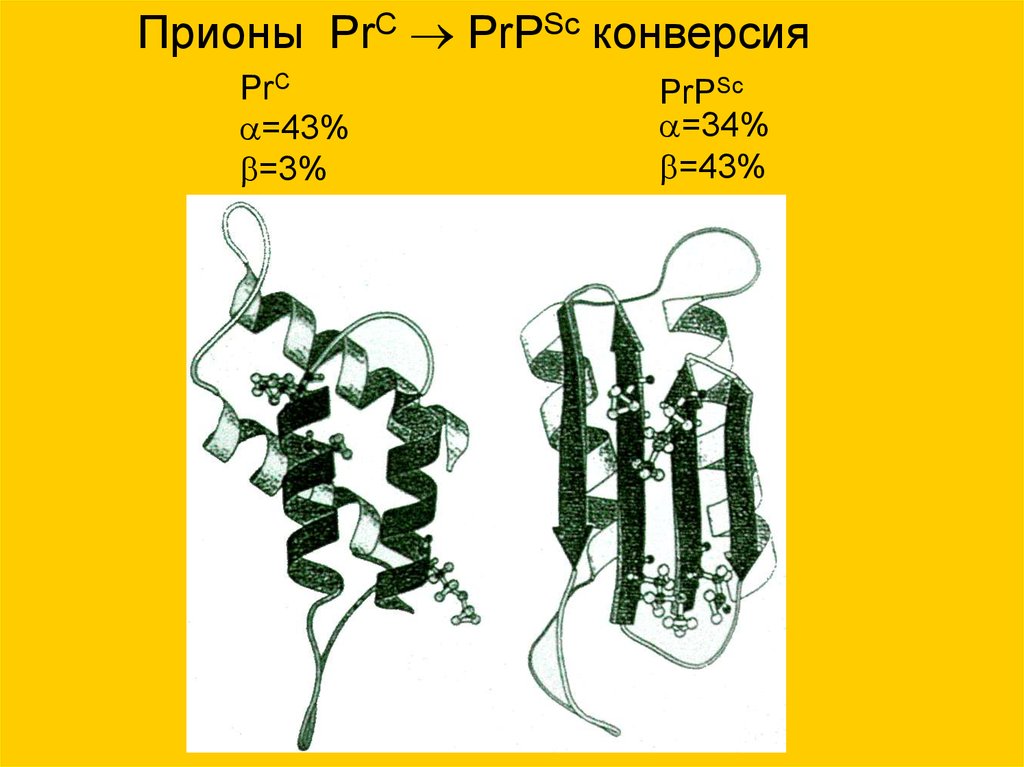
Прионы PrC PrPSc конверсияPrC
=43%
=3%
PrPSc
=34%
=43%
29.
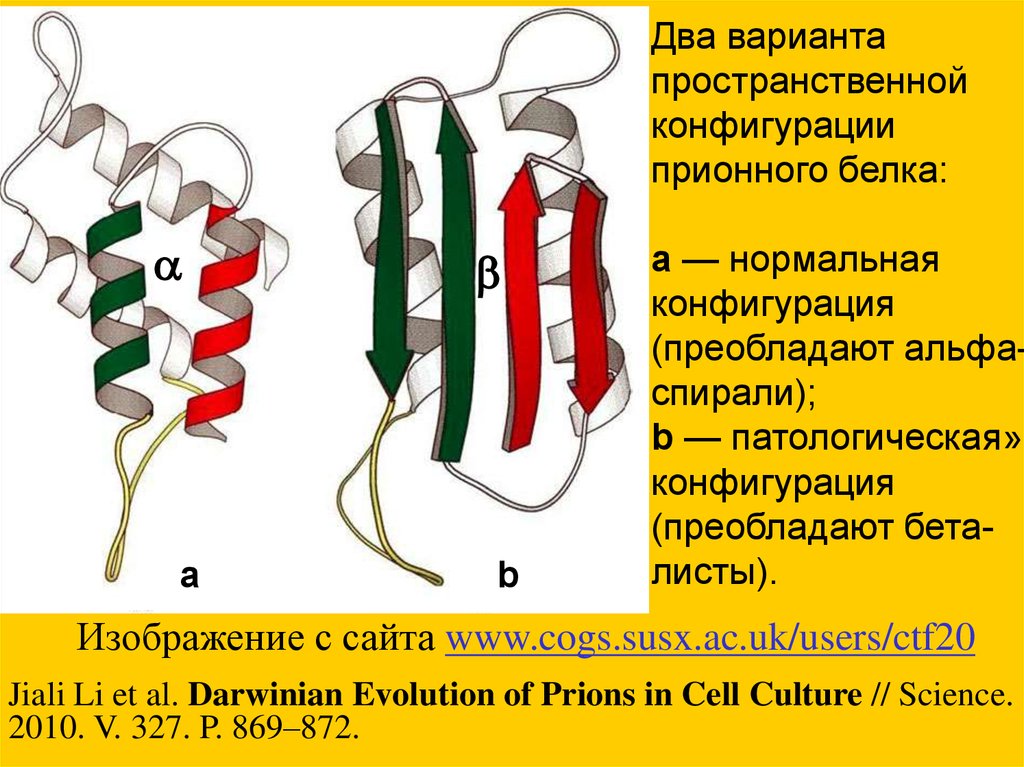
Два вариантапространственной
конфигурации
прионного белка:
a
b
a — нормальная
конфигурация
(преобладают альфаспирали);
b — патологическая»
конфигурация
(преобладают беталисты).
Изображение с сайта www.cogs.susx.ac.uk/users/ctf20
Jiali Li et al. Darwinian Evolution of Prions in Cell Culture // Science.
2010. V. 327. P. 869–872.
30.
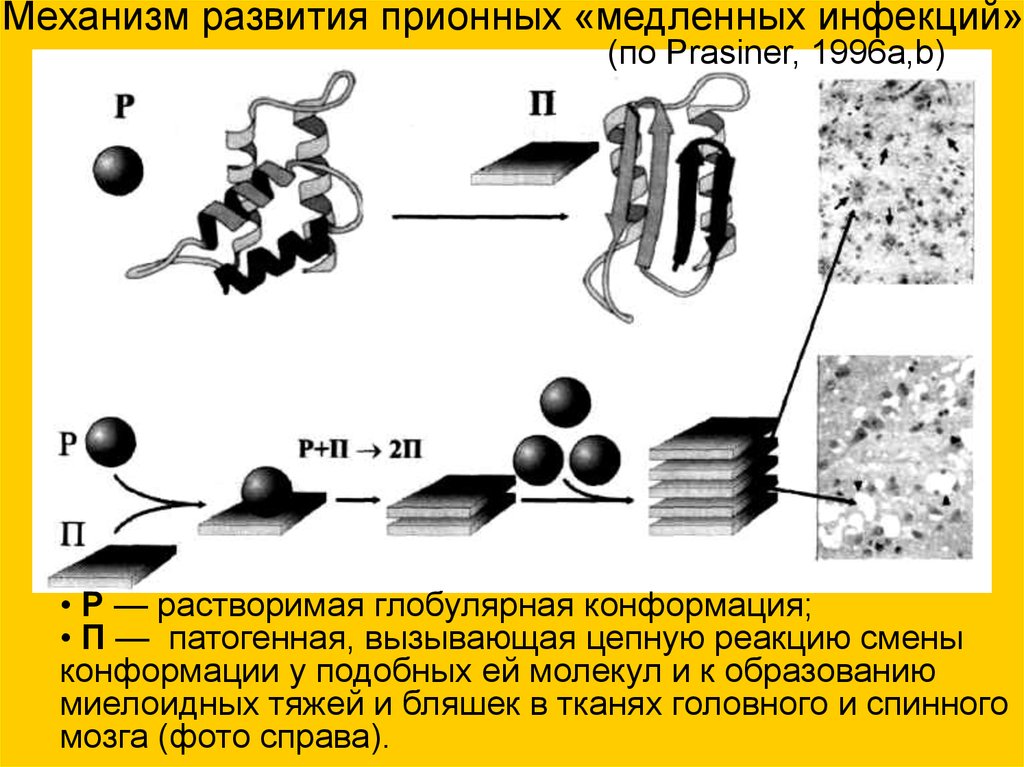
Механизм развития прионных «медленных инфекций»(по Prasiner, 1996a,b)
• Р — растворимая глобулярная конформация;
• П — патогенная, вызывающая цепную реакцию смены
конформации у подобных ей молекул и к образованию
миелоидных тяжей и бляшек в тканях головного и спинного
мозга (фото справа).
31. Прионы грибов
[Прион] (фенотип, продукт)[PSI] (нонсенс-супрессия)
[URE3] (усвоение уреидо сукцината)
[PIN+] (инициация [PSI]
[Неt-s](фактор несовместимости)
[ISP+] (антисупрессор к sup35)
[SWI+] (регуляция хроматина)
[ОСТ+] (транскрипционный
фактор)
[МСА] (метакаспаза)
[МОТЗ] (транскрипционный
фактор)
[GAR+] (устойчивость к
глюкозной репрессии)
Структурный
Вид
ген
SUP 35 Saccharomyces
cerevisiae
URE2
S. cerevisiae
RNQ1
HET-s
S. cerevisiae
Podospora
anserina
SFP1
S. cerevisiae
SWI1/SNF5 S. cerevisiae
CYC8/SSN6 S. cerevisiae
МСА1
МОТЗ
S. cerevisiae
S. cerevisiae
РМА1, STD1 S. cerevisiae
Источник
Cox, 1965; Wickner,
1994
Wickner, 1994
Derkatch et al, 2001
Coustou et al, 1997
Рогоза и др., 2009
Du et al., 2008
Patel et al., 2009
Nemecek et al., 2009
Alberty et al., 2009
Brown, Lindquist,
2009
32.
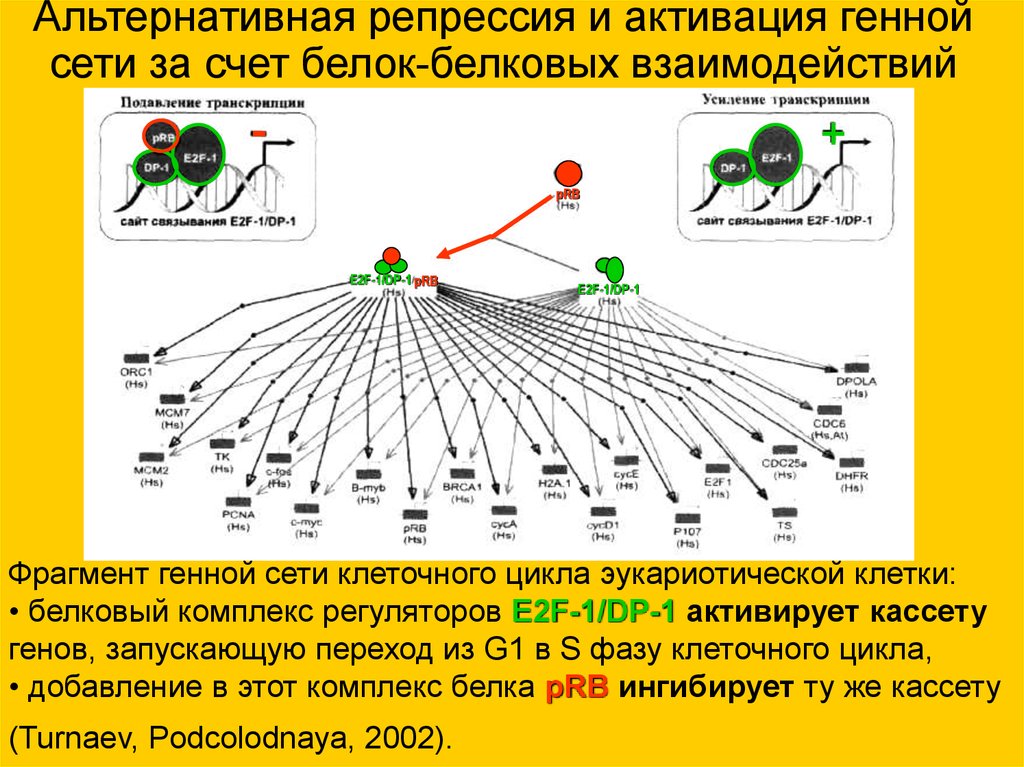
Альтернативная репрессия и активация геннойсети за счет белок-белковых взаимодействий
-
+
pRB
E2F-1/DP-1 pRB
E2F-1/DP-1
Фрагмент генной сети клеточного цикла эукариотической клетки:
• белковый комплекс регуляторов E2F-1/DP-1 активирует кассету
генов, запускающую переход из G1 в S фазу клеточного цикла,
• добавление в этот комплекс белка pRB ингибирует ту же кассету
(Turnaev, Podcolodnaya, 2002).
33.
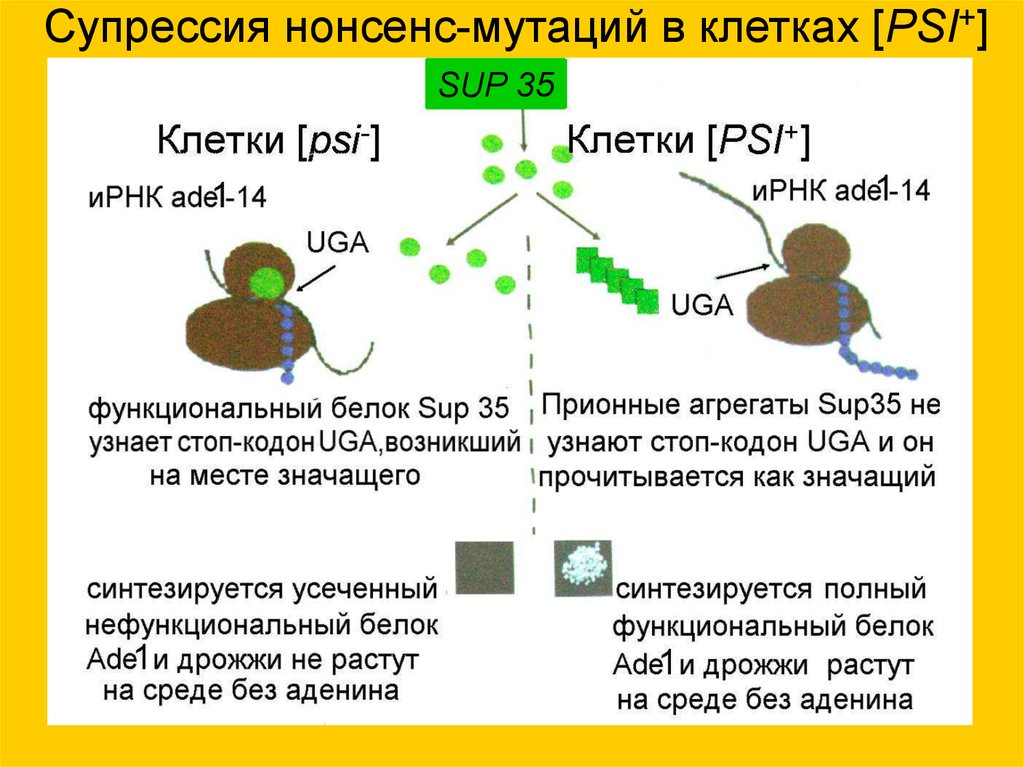
Супрессия нонсенс-мутаций в клетках [PSI+]SUP 35
34.
• Прионы мутируют, передают мутации понаследству, и эти мутации влияют на
эффективность размножения прионов в разных
условиях.
• Т.е., у прионов есть всё необходимое, чтобы
эволюционировать под действием естественного
отбора.
• Не ясно, может ли естественный отбор у прионов
быть «накопительным», то есть создавать новые
свойства путем последовательного закрепления
множества небольших наследственных изменений.
Jiali Li et al. Darwinian Evolution of Prions in Cell
Culture // Science. 2010. V. 327. P. 869–872.
35.
Центральная догма молекулярной биологии какотражение матричного принципа (Crick, 1958, 1970)
с дополнением
36.
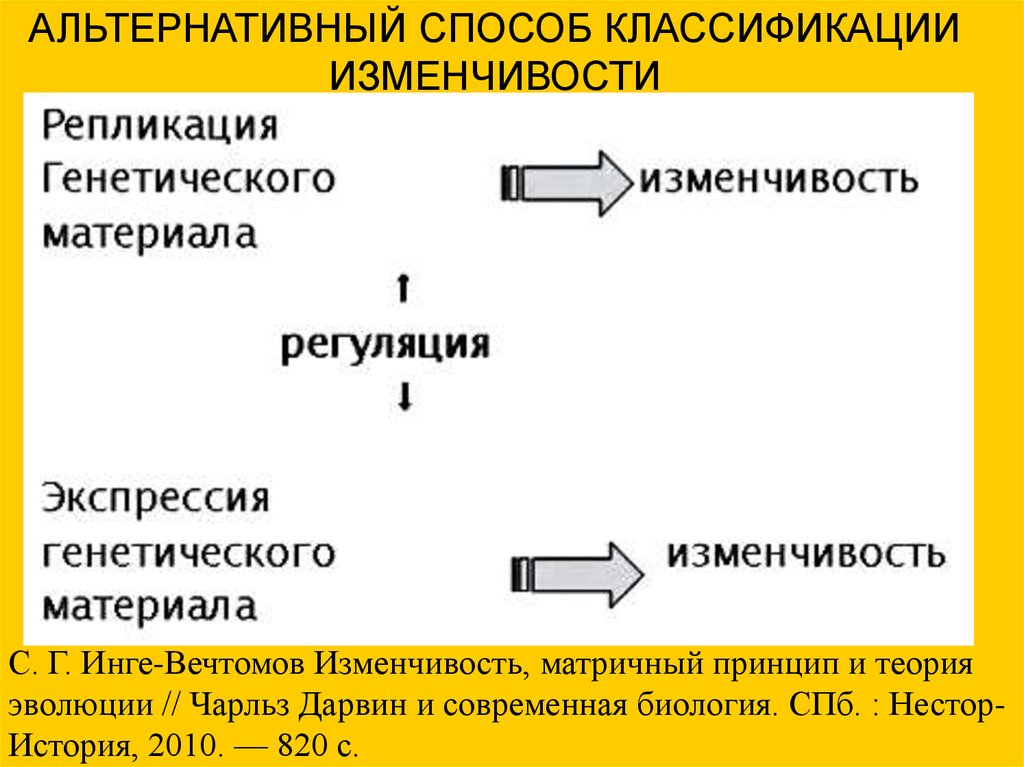
АЛЬТЕРНАТИВНЫЙ СПОСОБ КЛАССИФИКАЦИИИЗМЕНЧИВОСТИ
С. Г. Инге-Вечтомов Изменчивость, матричный принцип и теория
эволюции // Чарльз Дарвин и современная биология. СПб. : НесторИстория, 2010. — 820 с.
37.
• Два важнейших биологических макропринципа:1) конвариантная редупликация и отбор,
2) блочный принцип и негэнтропия,
3) ограниченность пространства и неограниченность времени.
Интроны:
1) участки гена без кодирующих последовательностей,
2) участки гена с кодирующими последовательностями,
3) транскрипционные факторы.
• Патогенная форма прионов:
1) мозаичная структура,
2) -спирали,
3) -листы.