








 biology
biologySimilar presentations:




")



")

Метаболические пути
1. МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ОБРАЗО
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО
ОБРАЗОВАНИЯ
РОССИЙСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ – МСХА ИМЕНИ К.А. ТИМИРЯЗЕВА
(ФГБОУ ВО РГАУ - МСХА ИМЕНИ К.А. ТИМИРЯЗЕВА)
Факультет зоотехнии и биологии
Метаболические пути
Выполнил студент 405 группы
Гарбузов С.Г.
Москва, 2017
2. ОСНОВНЫЕ МЕТАБОЛИЧЕСКИЕ ПУТИ
Характер метаболизма в тканях во многом определяетсяпитанием. У человека и ряда других млекопитающих
метаболическим превращениям подвергаются продукты,
абсорбируемые после переваривания содержащихся в пище
углеводов, липидов и белков. Это главным образом глюкоза,
триацилглицерол и, аминокислоты. У жвачных животных (и в
меньшей степени у других травоядных) целлюлоза
переваривается симбиотическими микроорганизмами с
образованием низших гомологов органических кислот
(уксусной, пропионовой, масляной); тканевый метаболизм у
этих животных адаптирован к утилизации в качестве основного
субстрата низших жирных кислот. (Марри Р., Греннер Д.,
Мейес П., Родуэлл В.)
3.
Тканевое дыханиеЭто процесс потребление клетками тканей организма кислорода, который
участвует в биологическом окислении. Такой вид окисления называют аэробным
окислением. Если конечным акцептором в цепи переноса водорода выступает не
кислород, а другие вещества (например пировиноградная кислота), то такой тип
окисления называют анаэробным.
Т.о. биологическое окисление - это дегидрирование субстрата с помощью
промежуточных переносчиков водорода и его конечного акцептора.
Дыхательная цепь (ферменты тканевого дыхания) - это переносчики протонов и
электронов от окисляемого субстрата на кислород. Окислитель - это соединение,
способное принимать электроны. Такая способность количественно
характеризуется окислительно-восстановительным потенциалом по отношению к
стандартному водородному электроду, рН которого равен 7,0. Чем меньше
потенциал соединения, тем сильнее его восстанавливающие свойства и наоборот.
Т. о. любое соединение может отдавать электроны только соединению с более
высоким окислительно-восстановительным потенциалом. В дыхательной цепи
каждое последующее звено имеет более высокий потенциал, чем предыдущее.
(Кнорре Д. Г., Мызина С. Д.)
4.
Дыхательная цепь состоит из:1.
НАД - зависимой дегидрогеназы;
2.
ФАД- зависимой дегидрогеназы;
3.
Убихинона (КоQ);
4.
Цитохрмов b, c, a+a3 .
• НАД-зависимые дегидрогеназы. В качестве кофермента содержат НАД и НАДФ.
Пиридиновое кольцо никотинамида способно присоединять электроны и протоны водорода.
• ФАД и ФМН-зависимые дегидрогеназы содержат в качестве кофермента фосфорный
эфир витамина В2 (ФАД).
• Убихинон (КоQ) отнимает водород у флавопротеидов и превращается при этом в
гидрохинон.
• Цитохромы - белки хромопротеиды, способные присоединять электроны, благодаря
наличию в своем составе в качестве простетических групп железопорфиринов. Они
принимают электрон от вещества, являющегося немного боле сильным восстановителем, и
передают его более сильному окислителю. Атом железа связан с атомом азота
имидазольного кольца аминоксилоты гистидина с одной стороны от плоскости
порфиринового цикла, а с другой стороны с атомом серы метионина. Поэтому
потенциальная способность атома железа в цитохромах к связыванию кислорода подавлена.
В цитохроме с порфириновая плоскость ковалентно связана с белком через два остатка
цистеина, а в цитохромах b и а, она ковалентно не связано с белком.
В цитохроме а+а3 (цитохромоксидазе) вместо протопорфирина содержатся порфирин А,
который отличатся рядом структурных особенностей. Пятое координационное положение
железа занято аминогруппой, принадлежащей остатку аминосахара, входящего в состав самого
белка. (Кнорре Д. Г., Мызина С. Д. )
5.
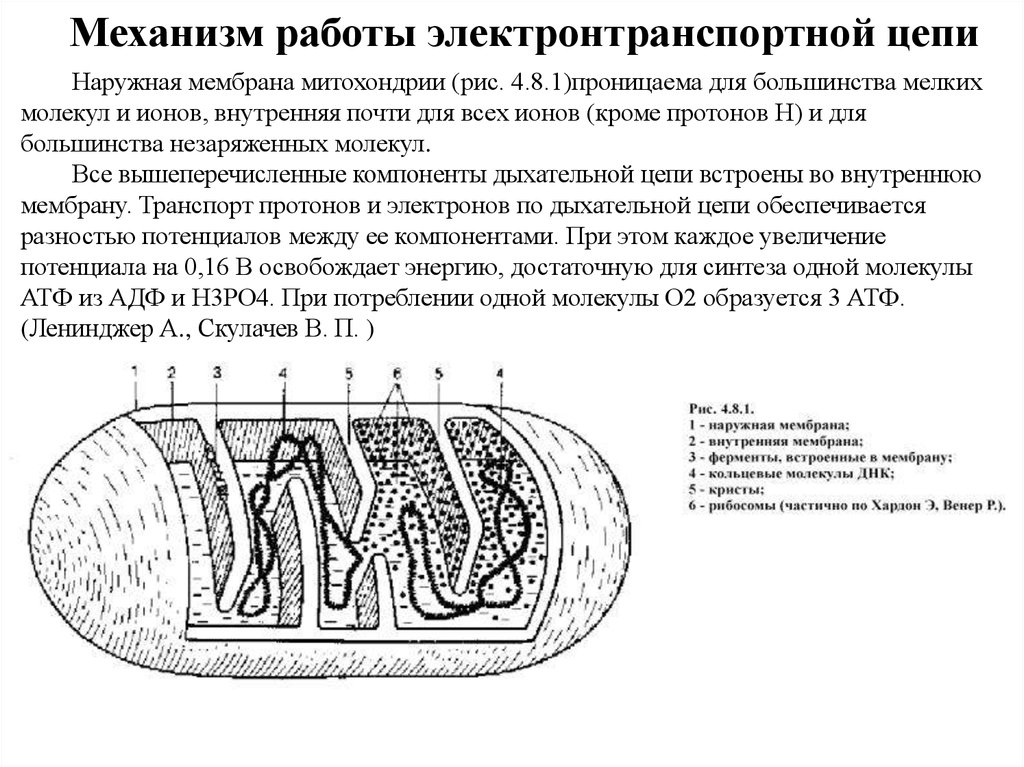
Механизм работы электронтранспортной цепиНаружная мембрана митохондрии (рис. 4.8.1)проницаема для большинства мелких
молекул и ионов, внутренняя почти для всех ионов (кроме протонов Н) и для
большинства незаряженных молекул.
Все вышеперечисленные компоненты дыхательной цепи встроены во внутреннюю
мембрану. Транспорт протонов и электронов по дыхательной цепи обеспечивается
разностью потенциалов между ее компонентами. При этом каждое увеличение
потенциала на 0,16 В освобождает энергию, достаточную для синтеза одной молекулы
АТФ из АДФ и Н3РО4. При потреблении одной молекулы О2 образуется 3 АТФ.
(Ленинджер А., Скулачев В. П. )
6.
Механизм работы электронтранспортной цепиПроцессы окисления и образования АТФ из АДФ и фосфорной кислоты т.е.
фосфорилирования протекают в митохондриях. Внутренняя мембрана образует
множество складок - крист. Пространство органиченное внутренней мембраной матриксом. Пространство между внутренней и наружной мембранами называется
межмембранным.
Такая молекула содержит в себе три макроэргических связи. Макроэргической
или богатой энергией называют химическую связь, при разрыве которой
высвобождается более 4 ккал/моль. При гидролитическом расщеплении АТФ до АДФ
и фосфорной кислоты высвобождается 7,3 ккал/моль. Ровно столько же тратится для
образования АТФ из АДФ и остатка фосфорной кислоты и это один из основных
путей запасания энергии в организме.
В процессе транспорта электронов по дыхательной цепи высвобождается
энергия, которая тратится на присоединение остатка фосфорной кислоты к АДФ с
образованием одной молекулы АТФ и одной молекулы воды. В процессе переноса
одной пары электронов по дыхательной цепи высвобождается и запасается в виде
трех молекул АТФ 21,3 ккал/моль. Это составляет около 40 % высвободившейся при
электронном транспорте энергии.
Такой способ запасания энергии в клетке называется окислительным
фосфорилированием или сопряженным фосфорилированием.
Молекулярные механизмы этого процесса наиболее полно объясняет
хемоосмотическая теория Митчелла, выдвинутая в 1961 году. (Ленинджер А.
Скулачев В. П. )
7.
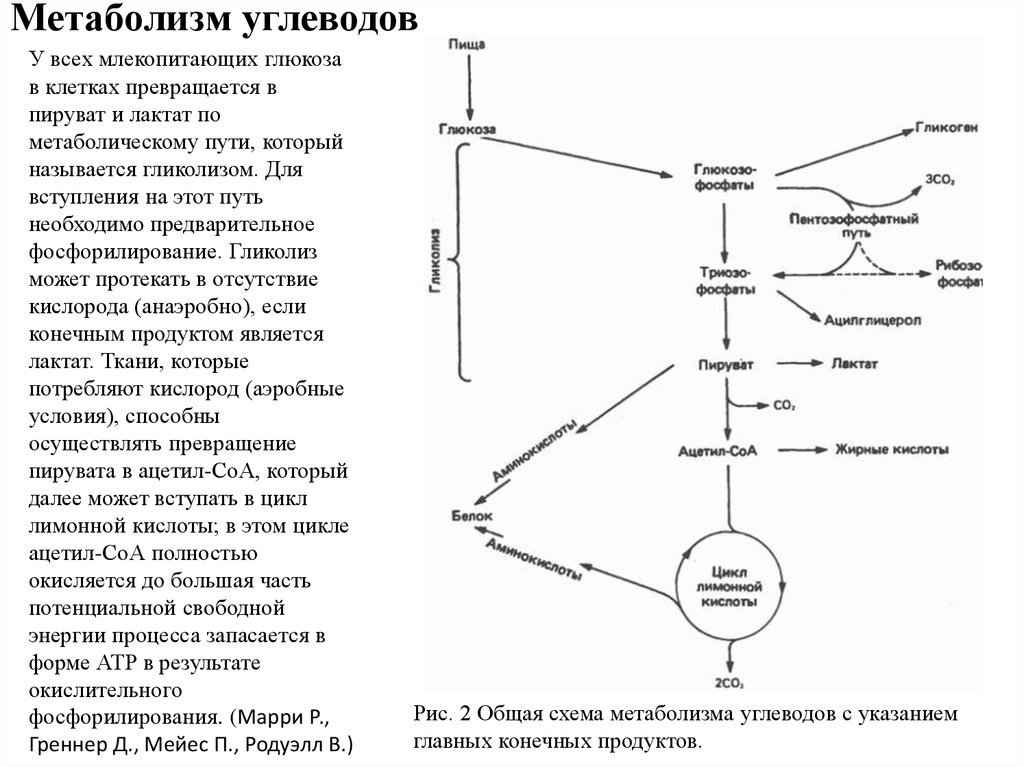
Метаболизм углеводовУ всех млекопитающих глюкоза
в клетках превращается в
пируват и лактат по
метаболическому пути, который
называется гликолизом. Для
вступления на этот путь
необходимо предварительное
фосфорилирование. Гликолиз
может протекать в отсутствие
кислорода (анаэробно), если
конечным продуктом является
лактат. Ткани, которые
потребляют кислород (аэробные
условия), способны
осуществлять превращение
пирувата в ацетил-СоА, который
далее может вступать в цикл
лимонной кислоты; в этом цикле
ацетил-СоА полностью
окисляется до большая часть
потенциальной свободной
энергии процесса запасается в
форме АТР в результате
окислительного
фосфорилирования. (Марри Р.,
Греннер Д., Мейес П., Родуэлл В.)
Рис. 2 Общая схема метаболизма углеводов с указанием
главных конечных продуктов.
8.
Метаболизм углеводовТаким образом, глюкоза служит главным видом топлива для многих тканей,
однако она (а также ее метаболиты) участвует и в других процессах.
1. Глюкоза превращается в полимер гликоген, который
запасается в ряде тканей, в особенности в скелетных мышцах и в печени.
2. Субстрат пентозофосфатного пути является одним из промежуточных
продуктов гликолиза. Этот путь служит источником восстановительных
эквивалентов используемых в процессах биосинтеза, например в биосинтезе
жирных кислот; кроме того, он является источником рибозы, необходимой для
синтеза нуклеотидов и нуклеиновых кислот.
3. Трнозофосфат, образующийся на одной из стадий гликолиза, является
источником глицерола, используемого в синтезе ацилглицеролов (жиров).
4. Пируват и ряд промежуточных соединений цикла лимонной кислоты—это
источники углеродных скелетов, используемых в синтезе аминокислот, а
ацетил-СоА служит основным строительным блоком в синтезе
длинноцепочечных жирных кислот и холестерола—предшественника всех
синтезируемых в организме стероидов (Марри Р., Греннер Д., Мейес П.,
Родуэлл В.).
9. Метаболизм липидов
Источником длинноцепочечных жирныхкислот служат синтез de novo из ацетил-СоА (в
свою очередь образующегося из углеводов) и
пищевые липиды. В тканях жирные кислоты
могут либо окисляться до ацетил-СоА (Рокисленне), либо эстерифицироваться в
ацилглицеролы (триацилглицерол является
главным энергетическим резервом организма).
образующийся при -окислении, участвует в
ряде важных процессов.
1. Ацетил-СоА может полностью окисляться до
в цикле лимонной кислоты. Жирные кислоты
являются источником значительных количеств
энергии (тканевым топливом) при утилизации
в процессе Р-окисления, а затем в ходе реакций
цикла лимонной кислоты.
2. Ацетил-СоА служит источником атомов
углерода для холестерола.
3. В печени из него образуется ацетоацетат —
исходное кетоновое тело. Кетоновые тела
являются альтернативным водорастворимым
тканевым топливом, которое при определенных Рис. 3. Общая схема метаболизма липидов с
условиях может стать важным источником
указанием главных конечных продуктов. Кетоновые
энергии (например, при голодании) (Марри Р.,
тела включают ацетоацетат, 3-гидроксибутират и
Греннер Д., Мейес П., Родуэлл В.).
ацетон.
10.
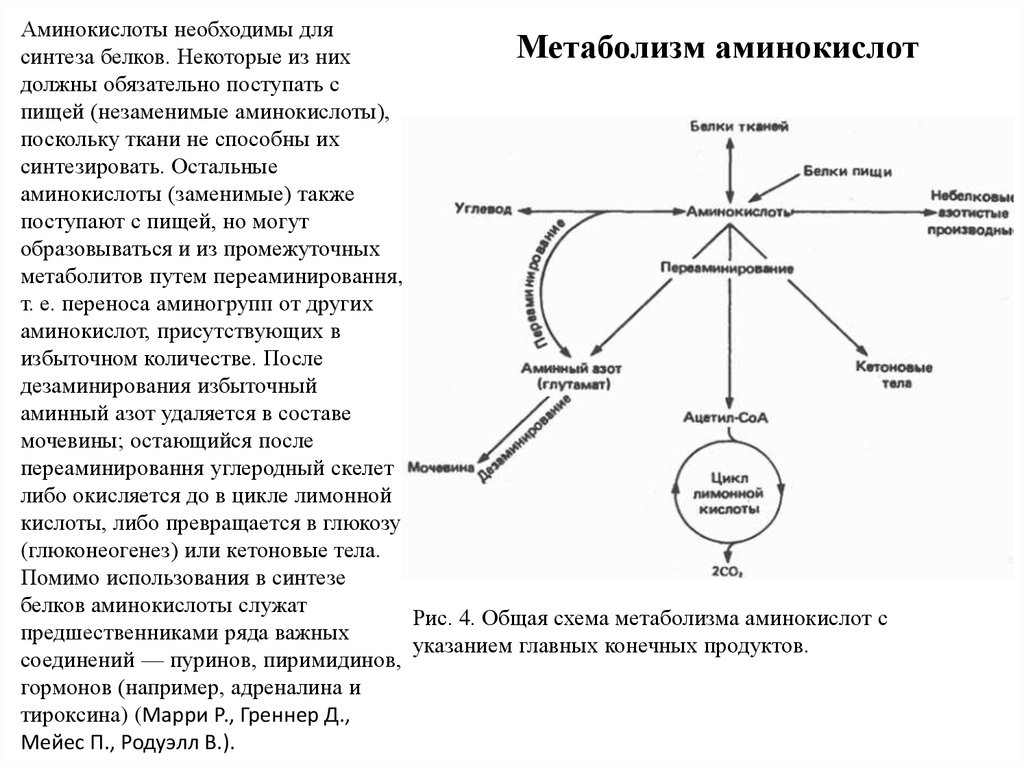
Аминокислоты необходимы дляМетаболизм аминокислот
синтеза белков. Некоторые из них
должны обязательно поступать с
пищей (незаменимые аминокислоты),
поскольку ткани не способны их
синтезировать. Остальные
аминокислоты (заменимые) также
поступают с пищей, но могут
образовываться и из промежуточных
метаболитов путем переаминировання,
т. е. переноса аминогрупп от других
аминокислот, присутствующих в
избыточном количестве. После
дезаминирования избыточный
аминный азот удаляется в составе
мочевины; остающийся после
переаминировання углеродный скелет
либо окисляется до в цикле лимонной
кислоты, либо превращается в глюкозу
(глюконеогенез) или кетоновые тела.
Помимо использования в синтезе
белков аминокислоты служат
Рис. 4. Общая схема метаболизма аминокислот с
предшественниками ряда важных
указанием главных конечных продуктов.
соединений — пуринов, пиримидинов,
гормонов (например, адреналина и
тироксина) (Марри Р., Греннер Д.,
Мейес П., Родуэлл В.).
11. Список литературы
1. Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимиячеловека: В 2-х томах. Т. 1. Пер. с англ.: — М.: Мир,
1993. - 384 с
2. Кнорре Д. Г., Мызина С. Д. Биологическая химия. – М.:
Высш. шк. 1998, 479 с.;
3. Ленинджер А. Биохимия. Молекулярные основы
структуры и функций клетки // М.: Мир, 1974, 956 с.;
4. Скулачев В. П. Энергетика биологических мембран // М.:
Наука, 1989, 565 с.;
5. Карякин А. В., Арчаков А. И. Межмембранный перенос
электронов // Успехи современной биологии, 1981, Т. 91,
вып. 1, с. 74-89;