













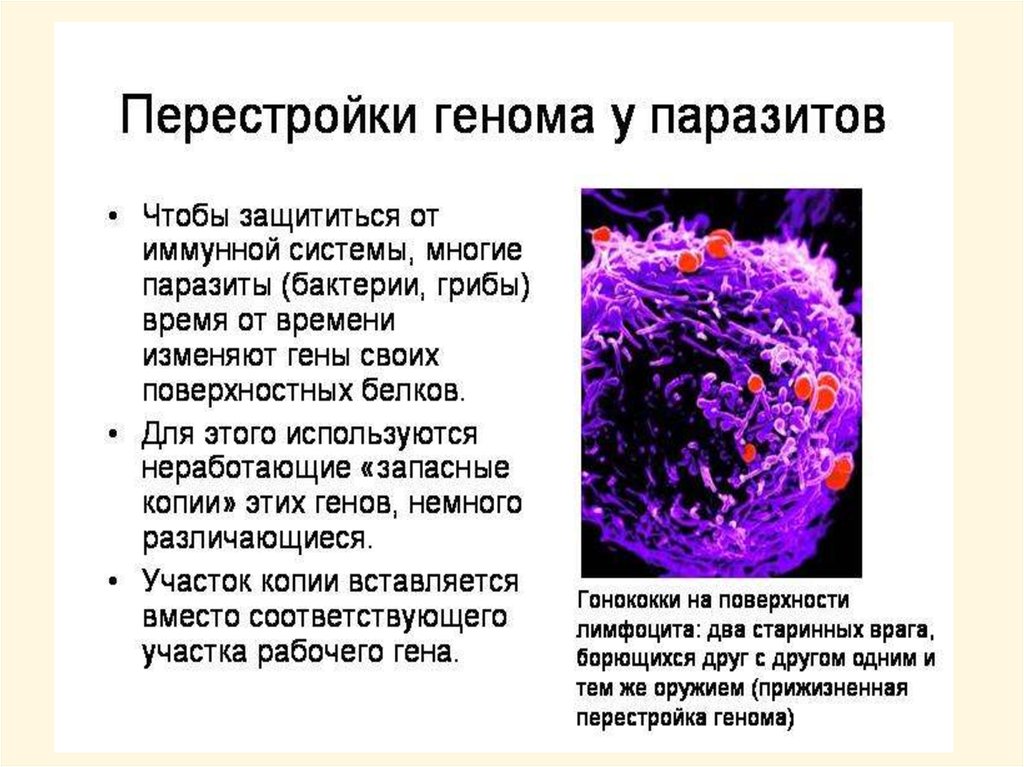







 biology
biologySimilar presentations:










Изменчивость. Формы изменчивости
1.
ИзменчивостьИзменчивость – способность организмов менять свои признаки и свойства
Формы изменчивости
Дискретная
Признаки – качественные
За формирование признака
отвечает 1 или 2
взаимодействующих гена; если
аллелей гена в популяции
несколько – полиморфизм
Примеры:
Группы крови, резус-фактор, масть у
свиней (эпистаз)
Непрерывная
Признаки – количественные
За формирование признака
отвечает несколько, иногда много
генов – полигенное наследование, в
т.ч. полимерия
Примеры:
Рост, жирность молока, интеллект
Иногда отдельно выделяют изменчивость, обусловленную плейотропным
(множественным) действием генов., например мутация белых глаз у дрозофилы
меняет цвет и размер внутренних органов, снижает жизнеспособность
2.
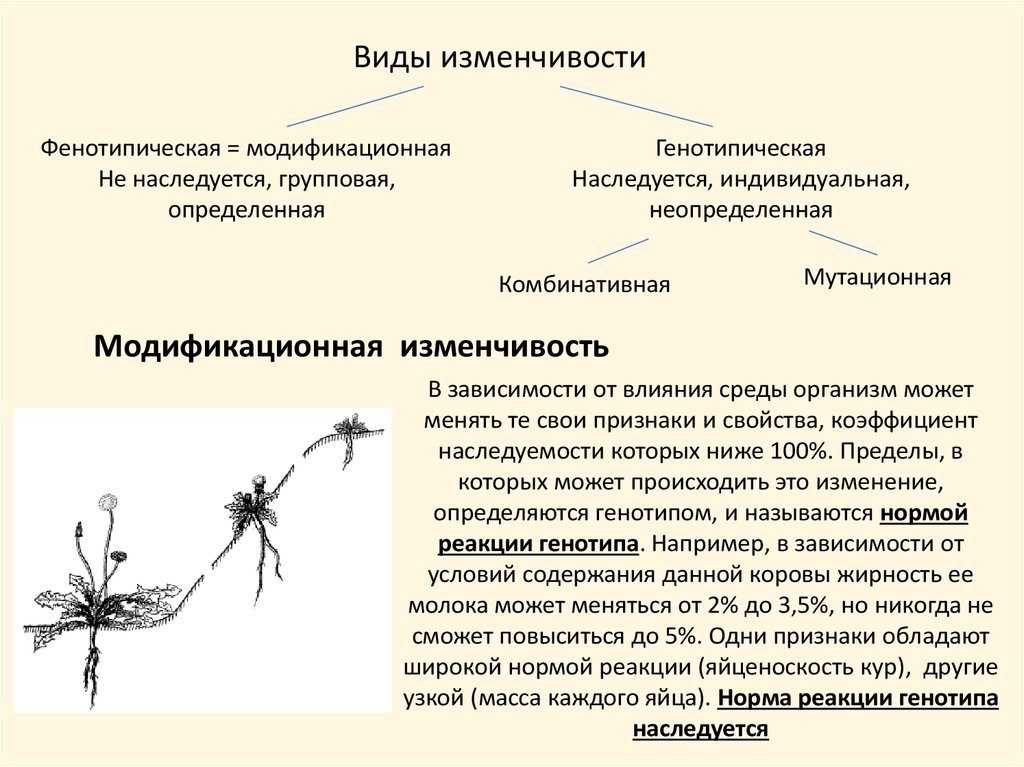
Виды изменчивостиФенотипическая = модификационная
Не наследуется, групповая,
определенная
Генотипическая
Наследуется, индивидуальная,
неопределенная
Комбинативная
Мутационная
Модификационная изменчивость
В зависимости от влияния среды организм может
менять те свои признаки и свойства, коэффициент
наследуемости которых ниже 100%. Пределы, в
которых может происходить это изменение,
определяются генотипом, и называются нормой
реакции генотипа. Например, в зависимости от
условий содержания данной коровы жирность ее
молока может меняться от 2% до 3,5%, но никогда не
сможет повыситься до 5%. Одни признаки обладают
широкой нормой реакции (яйценоскость кур), другие
узкой (масса каждого яйца). Норма реакции генотипа
наследуется
3.
Свойства модификационной изменчивости:• реализуется в пределах нормы реакции генотипа в ответ на влияние среды
• не наследуется
• носит групповой характер
• определенная, т.е. имеет предсказуемый адаптивный характер
Формы модификационной изменчивости:
• экологическая – формируется в
процессе развития организма в
результате взаимодействия
генотипа и среды
• сезонная – фенотип меняется под
влиянием гормонов, концентрация
которых определяется
фотопериодической реакцией
4.
Примеры, иллюстрирующие взаимодействие генотипа исреды при формировании признака.
Норма реакции генотипа кролика гималайской породы позволяет менять окраску.
Норма реакции стрелолиста позволяет менять форму листа
Генотип позволяет
синтезировать меланин, но
только на тех участках тела,
температура которых ниже
нормы.
1 – кролик содержится при
Форма
высокой температуре
листа у
2 – температура среды
стрелолиста
понижена
зависит от
3 – аналогично 2, но при
тех условий,
переводе кролика в
в которых
прохладное помещение у
развивается
него выбрили шерсть на
этот лист
бедре, поэтому новая шерсть
выросла черной
Модификационная изменчивость, хотя и не наследуется, но позволяет гибко
реагировать на изменения среды и сохранить в ряду поколений оптимальную норму
реакции генотипа
5.
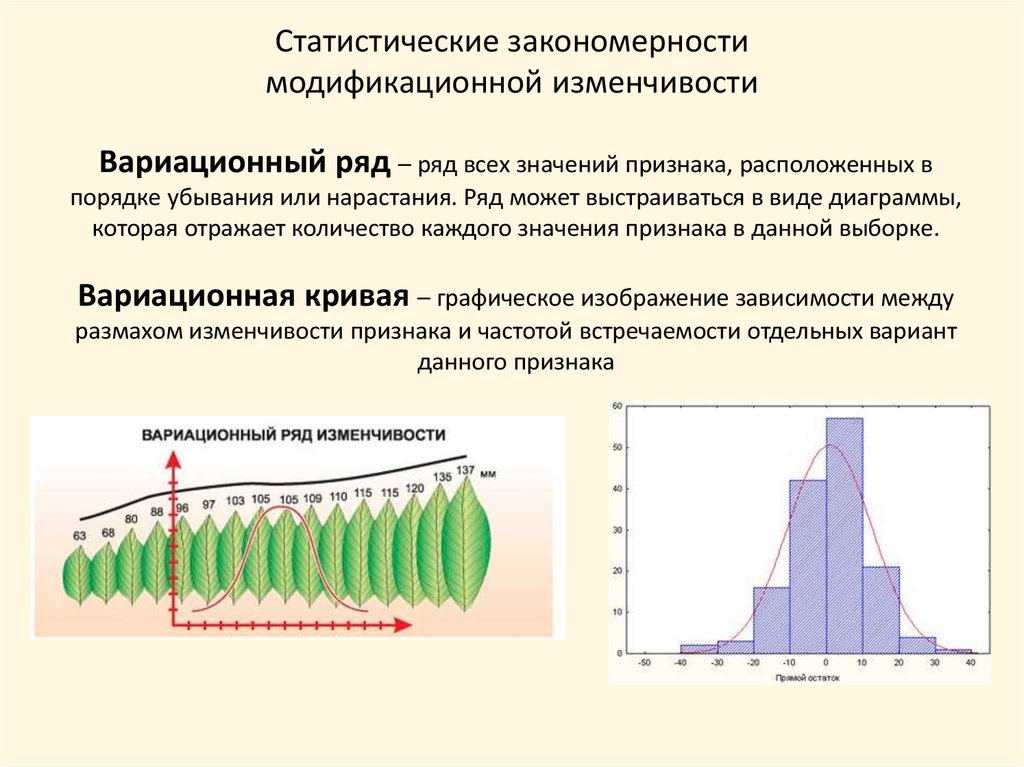
Статистические закономерностимодификационной изменчивости
Вариационный ряд – ряд всех значений признака, расположенных в
порядке убывания или нарастания. Ряд может выстраиваться в виде диаграммы,
которая отражает количество каждого значения признака в данной выборке.
Вариационная кривая – графическое изображение зависимости между
размахом изменчивости признака и частотой встречаемости отдельных вариант
данного признака
6.
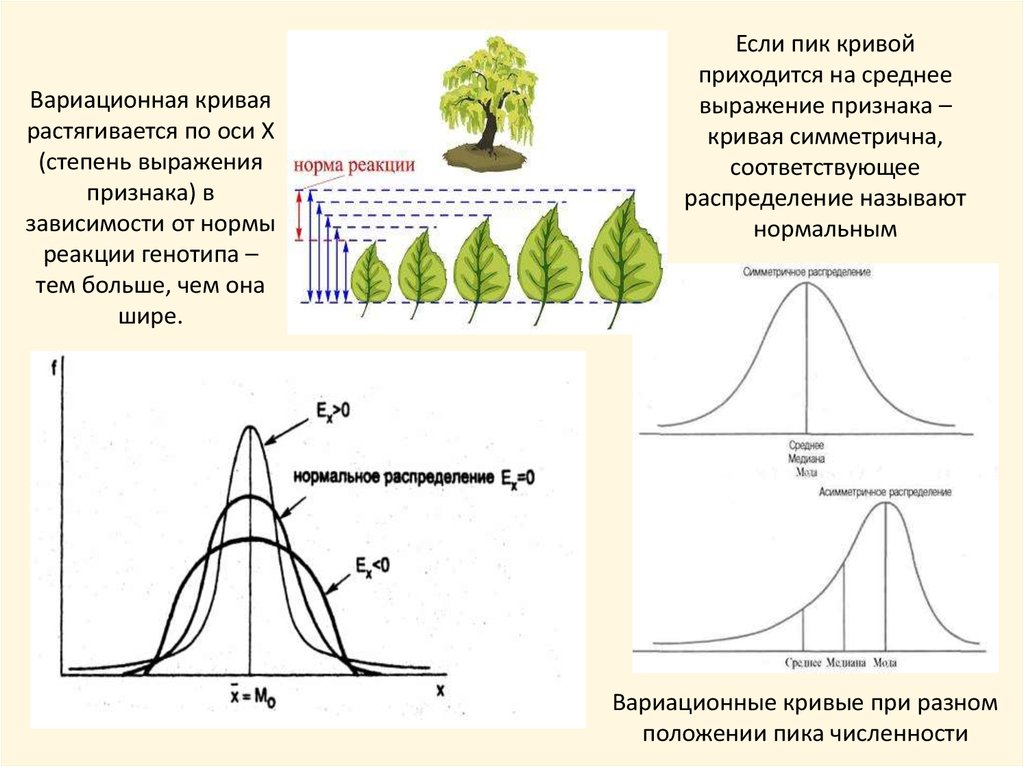
Вариационная криваярастягивается по оси Х
(степень выражения
признака) в
зависимости от нормы
реакции генотипа –
тем больше, чем она
шире.
Если пик кривой
приходится на среднее
выражение признака –
кривая симметрична,
соответствующее
распределение называют
нормальным
Вариационные кривые при разном
положении пика численности
7.
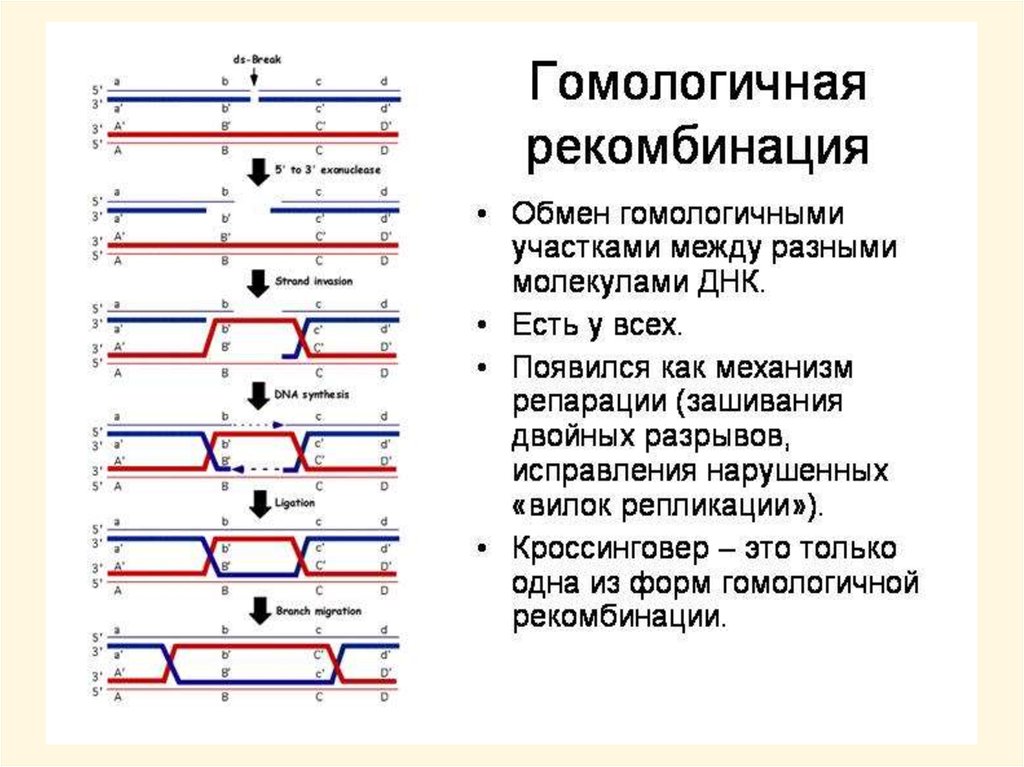
Генотипическая комбинативная изменчивостьКомбинативной изменчивостью называют индивидуальные ненаправленные
изменения генотипа, основанные на рекомбинации генетического материала. Для
ее реализации у прокариот необходим половой (парасексуальный) процесс, у
эукариот - мейоз и последующее половое размножение
Причины комбинативной изменчивости:
• рекомбинация генов в результате кроссинговера при мейозе – наиболее
устойчивый вариант рекомбинации, может долгое время удерживаться в ряду
поколений
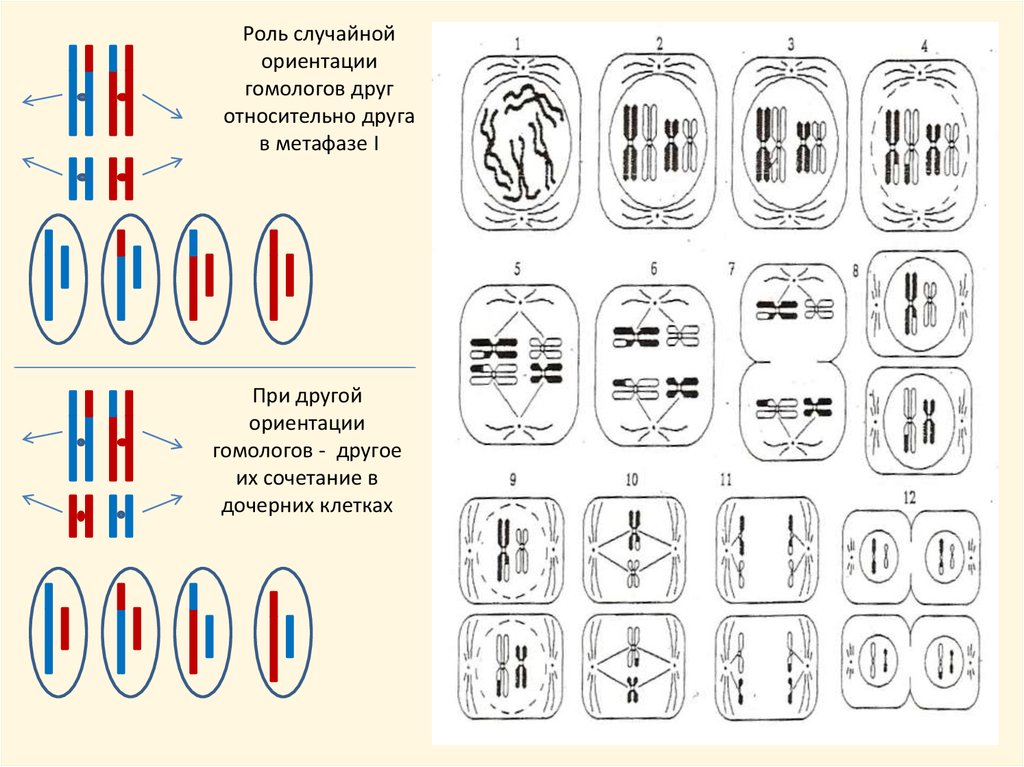
• новое сочетание хромосом в результате случайной взаимной ориентации
гомологов относительно полюсов и относительно друг друга в метафазе I
мейоза, и вследствие этого – их случайное сочетание при расхождении в
дочерние клетки в анафазе I мейоза. Каждое сочетание хромосом наследуется на
одно следующее поколение
• аналогично случайная ориентация кроссоверных хроматид в метафазе II
мейоза
• случайное сочетание гамет при оплодотворении – приводит к рекомбинации
генетического материала в следующем поколении. Не работает в случае
партеногенеза
Комбинативная изменчивость обеспечивает огромное разнообразие генотипов, но
не дает возможность получить новые аллели, и неустойчива в ряду поколений при
половом размножении
8.
Роль случайнойориентации
гомологов друг
относительно друга
в метафазе I
При другой
ориентации
гомологов - другое
их сочетание в
дочерних клетках
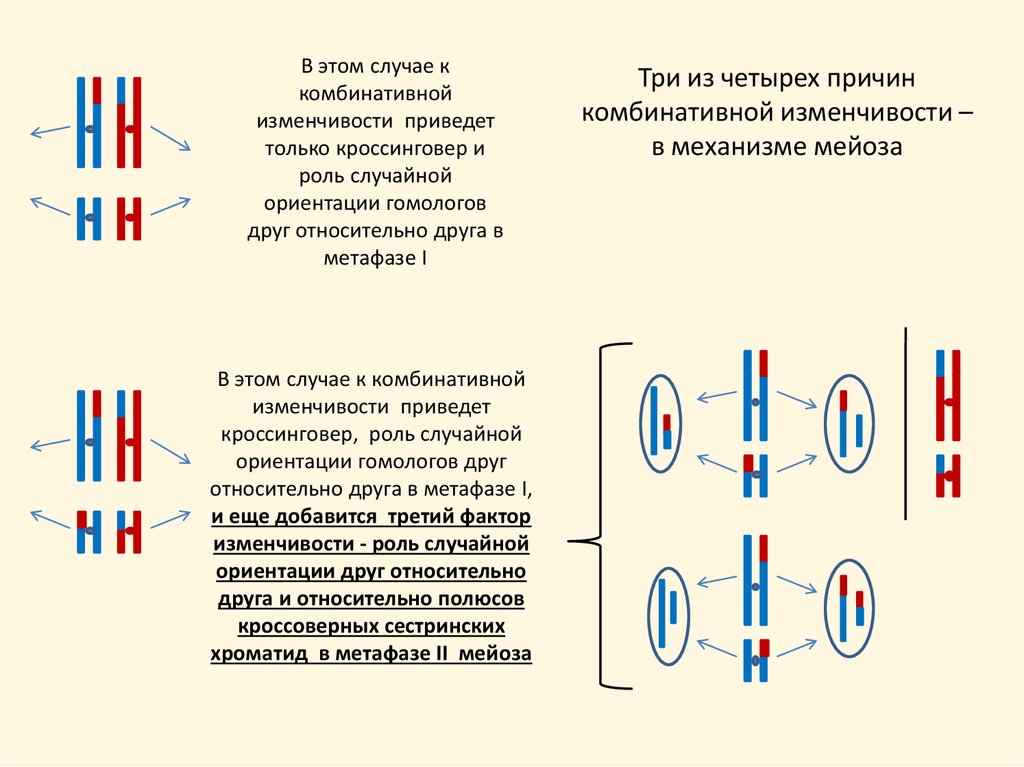
9.
В этом случае ккомбинативной
изменчивости приведет
только кроссинговер и
роль случайной
ориентации гомологов
друг относительно друга в
метафазе I
В этом случае к комбинативной
изменчивости приведет
кроссинговер, роль случайной
ориентации гомологов друг
относительно друга в метафазе I,
и еще добавится третий фактор
изменчивости - роль случайной
ориентации друг относительно
друга и относительно полюсов
кроссоверных сестринских
хроматид в метафазе II мейоза
Три из четырех причин
комбинативной изменчивости –
в механизме мейоза
10.
Генотипическая мутационная изменчивостьМутационной изменчивостью называют стойкие индивидуальные
ненаправленные, наследуемые в ряду поколений, изменения генотипа.
Мутация – отдельное событие мутационной изменчивости, т.е. стойкое …
Мутационную теорию начал разрабатывать в начале 20 века Г. Де Фриз
Мутации классифицируются разными способами, исходя из разных
1.
2.
3.
4.
5.
6.
7.
характеристик:
ядерные и цитоплазматические (в ДНК митохондрий, пластид)
генеративные (в гаметах или их предшественниках) и соматические (в
клетках тела; наследуются при вегетативном размножении)
прямые (меняют генотип) и обратные (возвращают фенотип в исходное
состояние; реверсия может быть истинной либо компенсаторной)
для данных условий среды полезные, нейтральные, вредные, в т.ч.
летальные
спонтанные (причина не понятна) и индуцированные (вызваны известным
мутагеном)
доминантные, рецессивные, кодоминантные
точечные = генные, хромосомные, геномные
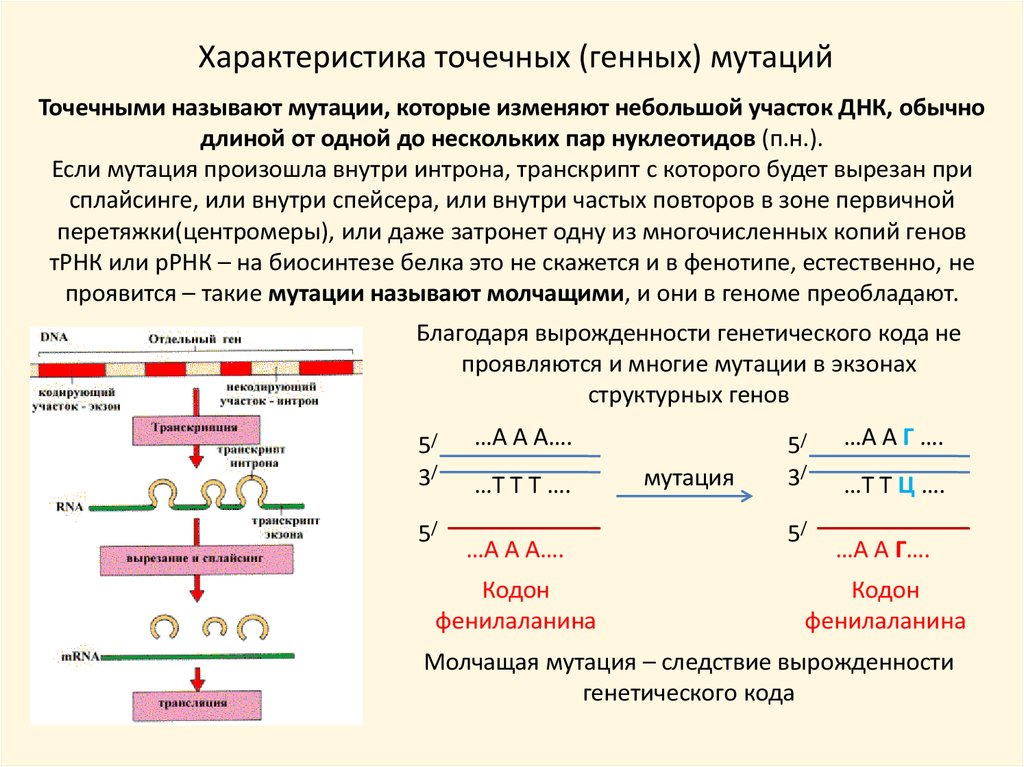
11.
Характеристика точечных (генных) мутацийТочечными называют мутации, которые изменяют небольшой участок ДНК, обычно
длиной от одной до нескольких пар нуклеотидов (п.н.).
Если мутация произошла внутри интрона, транскрипт с которого будет вырезан при
сплайсинге, или внутри спейсера, или внутри частых повторов в зоне первичной
перетяжки(центромеры), или даже затронет одну из многочисленных копий генов
тРНК или рРНК – на биосинтезе белка это не скажется и в фенотипе, естественно, не
проявится – такие мутации называют молчащими, и они в геноме преобладают.
Благодаря вырожденности генетического кода не
проявляются и многие мутации в экзонах
структурных генов
5/
3/
5/
…А А А….
…Т Т Т ….
…А А А….
Кодон
фенилаланина
мутация
5/
3/
5/
…А А Г ….
…Т Т Ц ….
…А А Г….
Кодон
фенилаланина
Молчащая мутация – следствие вырожденности
генетического кода
12.
Если мутация произойдет внутри экзона структурного гена таким образом, что вбелке заменится аминокислота, но не будут затронуты регуляторный и активный
центры белка – мутацию можно будет назвать «консервативным замещением».
Если в результате мутации свойства белка изменяются – ее называют «радикальной
заменой».
Пример точечной мутации, при
которой происходит замена одной
АК на другую
Наиболее известный пример
радикальной замены серповидноклеточная анемия, при
которой происходит нарушение в
строении бета-цепи гемоглобина: вместо
глутаминовой кислоты в 6-м положении
стоит валин.
13.
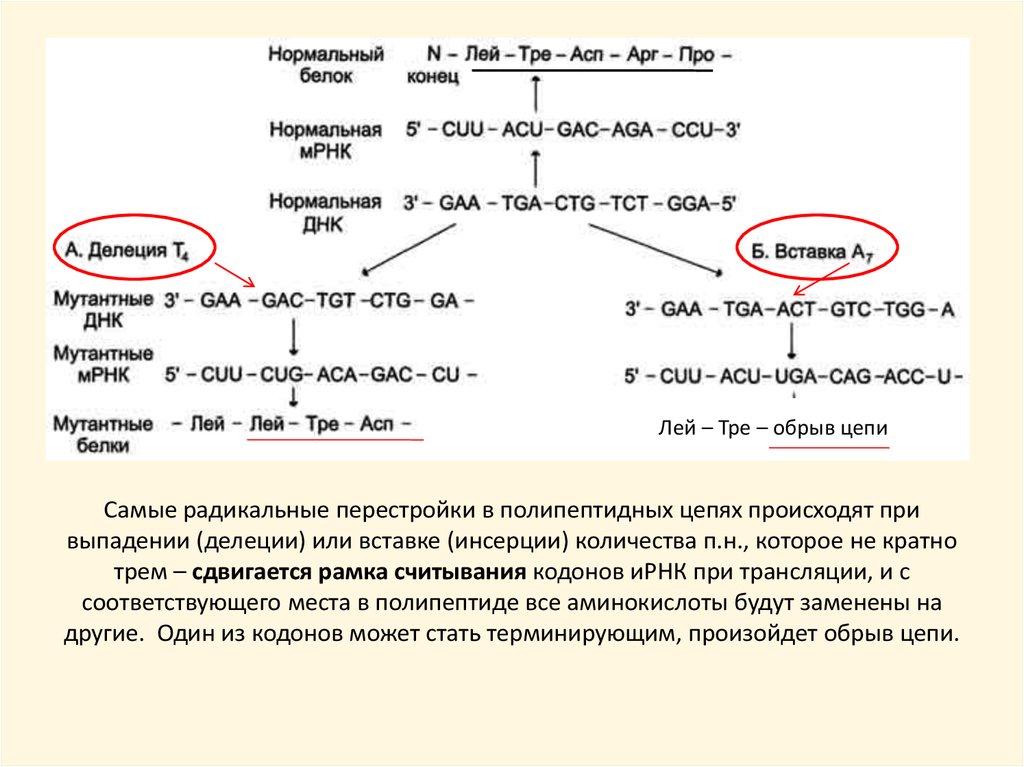
Лей – Тре – обрыв цепиСамые радикальные перестройки в полипептидных цепях происходят при
выпадении (делеции) или вставке (инсерции) количества п.н., которое не кратно
трем – сдвигается рамка считывания кодонов иРНК при трансляции, и с
соответствующего места в полипептиде все аминокислоты будут заменены на
другие. Один из кодонов может стать терминирующим, произойдет обрыв цепи.
14.
Генные мутации, происходящие в процессегаметогенеза, могут стать источником
получения новых аллелей – это увеличивает
гетерозиготность популяции, обогащает ее
генофонд
Некоторые гены очень устойчивы,
спонтанные мутации происходят в них с
частотой 3 раза на 10 миллиардов гамет.
Самые неустойчивые гены обнаружены у
кукурузы – 2 новые мутации на тысячу гамет.
Полиморфизм по признаку окраски
шерсти
У человека средняя частота мутаций каждого гена – от 30 до 100 на 1 миллион гамет,
общее число новых мутаций на зиготу – более 60 (среди них преобладают молчащие
мутации, новых радикальных замен только 1,6 на зиготу). Подавляющее большинство
мутаций не конструктивны, они могут нарушить транскрипцию гена, либо транскрипт
с него нормально не работает. В итоге у каждого человека на гаплоидный набор в
соматических клетках приходится 8 летальных доз мутаций (новые вместе с
полученными от предков), но поскольку такие мутации обычно рецессивны, а
соматические клетки диплоидны, организм выживает.
15.
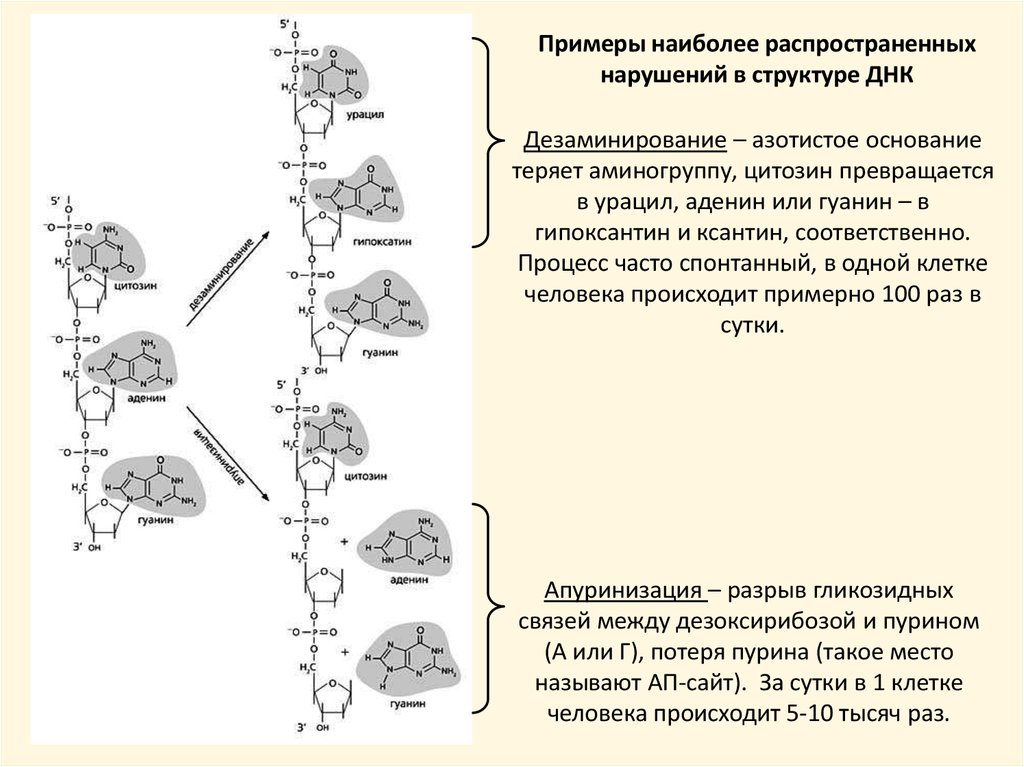
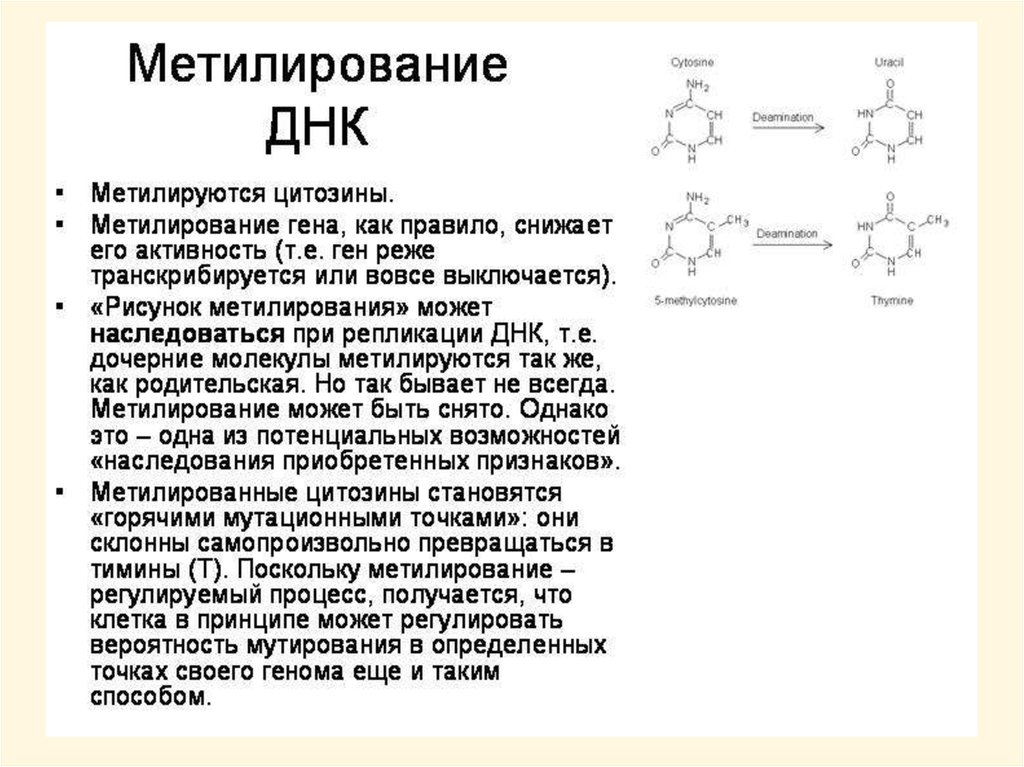
Примеры наиболее распространенныхнарушений в структуре ДНК
Дезаминирование – азотистое основание
теряет аминогруппу, цитозин превращается
в урацил, аденин или гуанин – в
гипоксантин и ксантин, соответственно.
Процесс часто спонтанный, в одной клетке
человека происходит примерно 100 раз в
сутки.
Апуринизация – разрыв гликозидных
связей между дезоксирибозой и пурином
(А или Г), потеря пурина (такое место
называют АП-сайт). За сутки в 1 клетке
человека происходит 5-10 тысяч раз.
16.
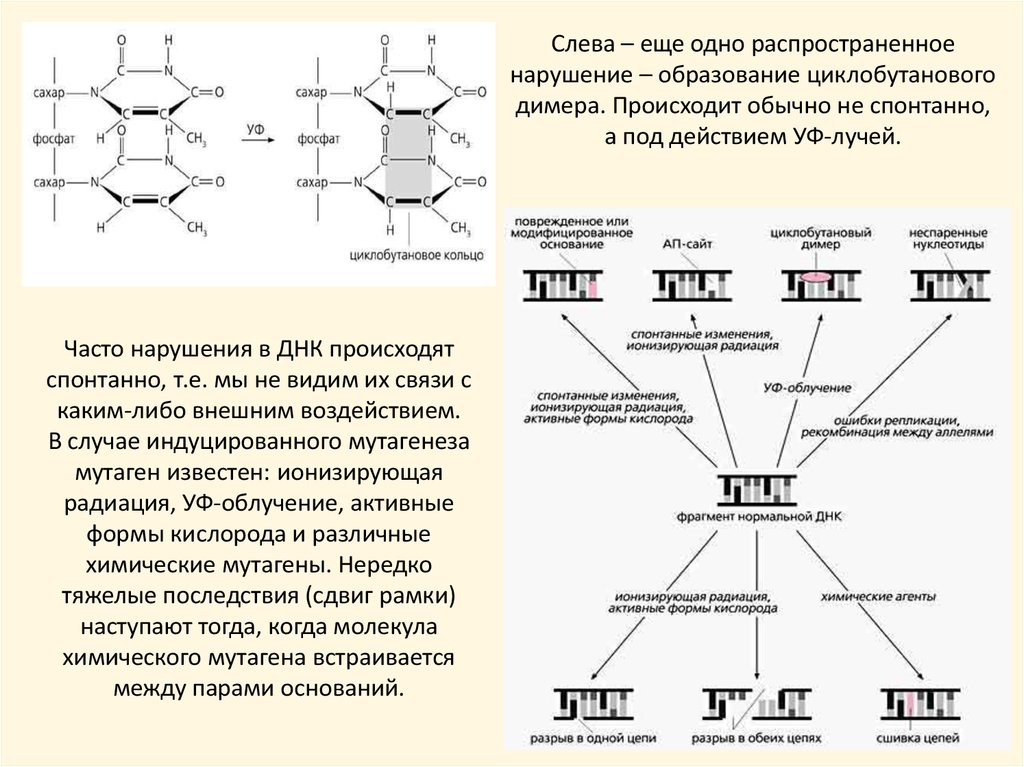
Слева – еще одно распространенноенарушение – образование циклобутанового
димера. Происходит обычно не спонтанно,
а под действием УФ-лучей.
Часто нарушения в ДНК происходят
спонтанно, т.е. мы не видим их связи с
каким-либо внешним воздействием.
В случае индуцированного мутагенеза
мутаген известен: ионизирующая
радиация, УФ-облучение, активные
формы кислорода и различные
химические мутагены. Нередко
тяжелые последствия (сдвиг рамки)
наступают тогда, когда молекула
химического мутагена встраивается
между парами оснований.
17.
Повреждения ДНК очень разнообразныи достаточно часто происходят. Если бы
клетки не имели системы репарации,
т.е. исправления повреждений,
генетический аппарат был бы очень
нестабильным, наследование признаков
было бы под большой угрозой. Реально
система репарации есть, ее обслуживает
несколько десятков генов, она позволяет
исправить почти все повреждения.
Генными мутациями мы называем
только те изменения в исходной
структуре ДНК, которые не удалось
исправить и они остались.
Предполагается, что на самых ранних этапах эволюции протобионтов к миру
однонитевых РНК добавилась ДНК именно потому, что повреждения двунитевой
молекулы существенно легче исправить, вследствие чего она намного стабильнее. Еще
большее увеличение точности копирования и безошибочного сохранения
генетической информации теоретически возможно, но не целесообразно – оно стоило
бы организмам слишком дорого, а главное – привело бы к такой стабильности генома,
которая затруднила бы адаптации к меняющейся среде.
Другая возможность попытаться нейтрализовать эффект мутаций в структурных
генах – синтез дополнительного количества шаперонов (см. дополнения)
18.
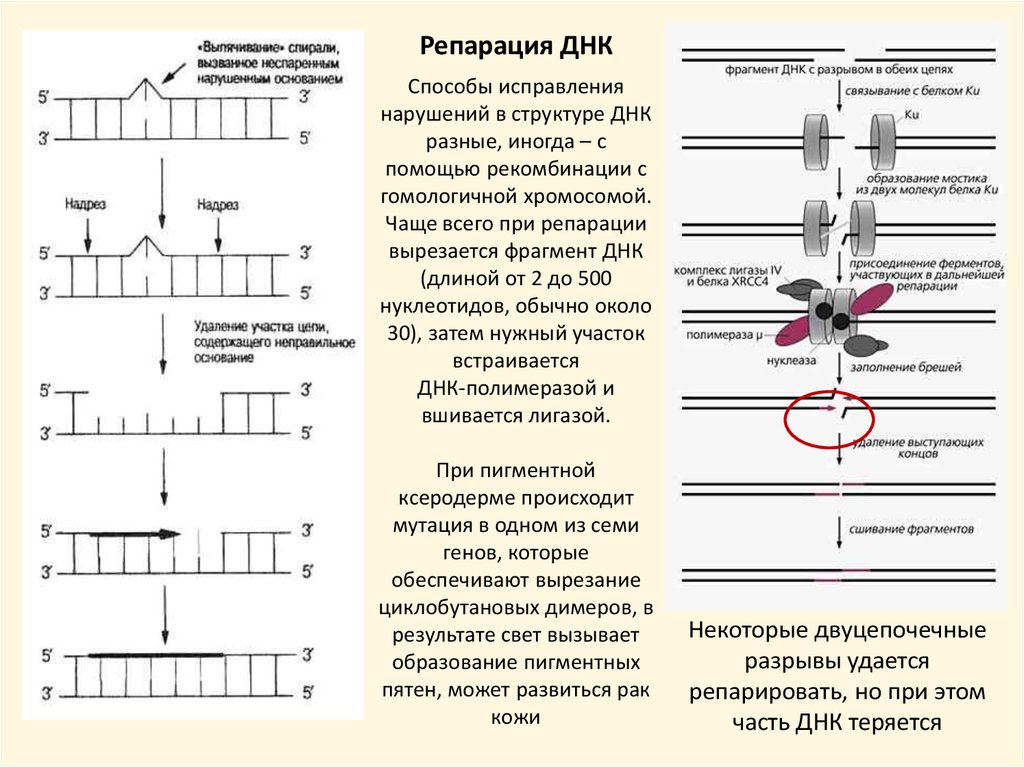
Репарация ДНКСпособы исправления
нарушений в структуре ДНК
разные, иногда – с
помощью рекомбинации с
гомологичной хромосомой.
Чаще всего при репарации
вырезается фрагмент ДНК
(длиной от 2 до 500
нуклеотидов, обычно около
30), затем нужный участок
встраивается
ДНК-полимеразой и
вшивается лигазой.
При пигментной
ксеродерме происходит
мутация в одном из семи
генов, которые
обеспечивают вырезание
циклобутановых димеров, в
результате свет вызывает
образование пигментных
пятен, может развиться рак
кожи
Некоторые двуцепочечные
разрывы удается
репарировать, но при этом
часть ДНК теряется
19.
20.
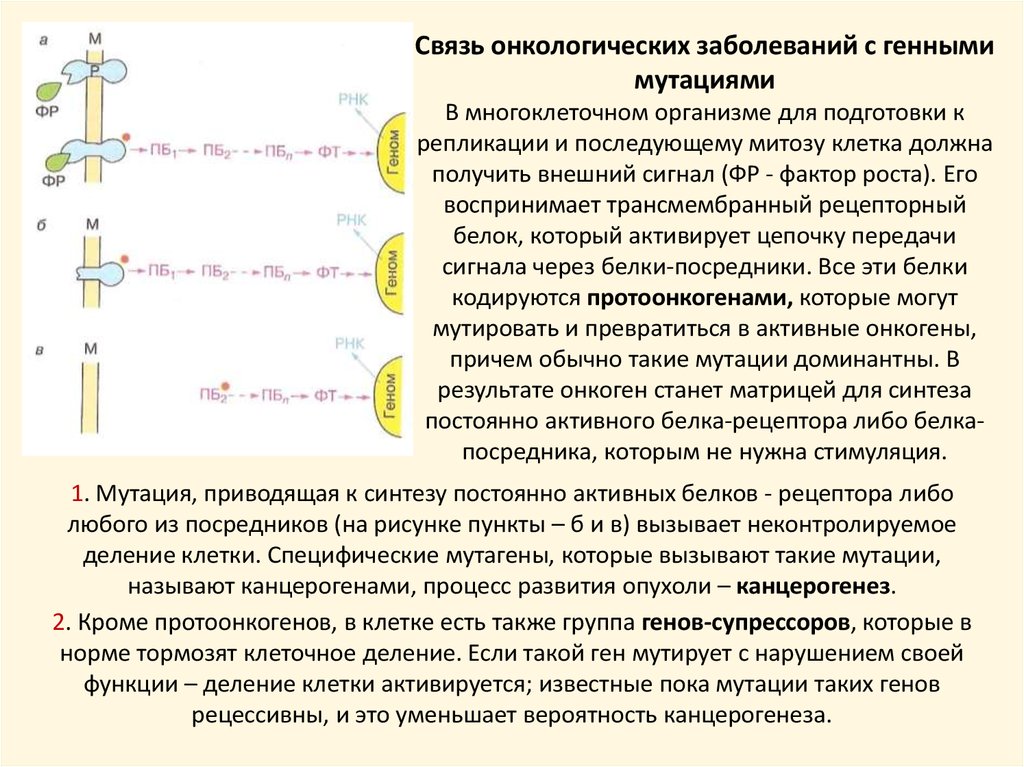
Связь онкологических заболеваний с геннымимутациями
В многоклеточном организме для подготовки к
репликации и последующему митозу клетка должна
получить внешний сигнал (ФР - фактор роста). Его
воспринимает трансмембранный рецепторный
белок, который активирует цепочку передачи
сигнала через белки-посредники. Все эти белки
кодируются протоонкогенами, которые могут
мутировать и превратиться в активные онкогены,
причем обычно такие мутации доминантны. В
результате онкоген станет матрицей для синтеза
постоянно активного белка-рецептора либо белкапосредника, которым не нужна стимуляция.
1. Мутация, приводящая к синтезу постоянно активных белков - рецептора либо
любого из посредников (на рисунке пункты – б и в) вызывает неконтролируемое
деление клетки. Специфические мутагены, которые вызывают такие мутации,
называют канцерогенами, процесс развития опухоли – канцерогенез.
2. Кроме протоонкогенов, в клетке есть также группа генов-супрессоров, которые в
норме тормозят клеточное деление. Если такой ген мутирует с нарушением своей
функции – деление клетки активируется; известные пока мутации таких генов
рецессивны, и это уменьшает вероятность канцерогенеза.
21.
3. Еще одна группа из нескольких десятков генов активна у эмбрионов,обеспечивая быстрое деление их клеток, а на более поздних стадиях развития
организма эти гены блокируются. Они могут быть активированы и тогда станут
онкогенами в любой клетке взрослого организма, и сделает это либо канцероген,
либо рядом встроится какая-то активно работающая ДНК
4. Канцерогены могут действовать и обходным путем – могут нарушать работу
генов, которые обеспечивают репарацию ДНК.
5. Некоторые виды онкологических заболеваний вызываются РНК-содержащими
ретровирусами, которые по строению и общей стратегии сходны с ВИЧ. Такие вирусы
синтезируют свою кДНК и встраивают ее в генетический аппарат клетки, активируя
прилегающие участки хозяйской ДНК и провоцируя, таким образом, канцерогенез.
Если такие интегрированные вирусы (их называют провирусы) будут встроены в ДНК
гамет или их предшественников – они перейдут к следующему поколению.
Предполагается, что многочисленные ретротранспозоны («прыгающие гены») в
наших хромосомах по своему происхождению связаны с интегрированными
ретровирусами.
С возрастом риск онкологических заболеваний увеличивается, так как различные
соматические мутации накапливаются, способность к репарации ДНК уменьшается.
22.
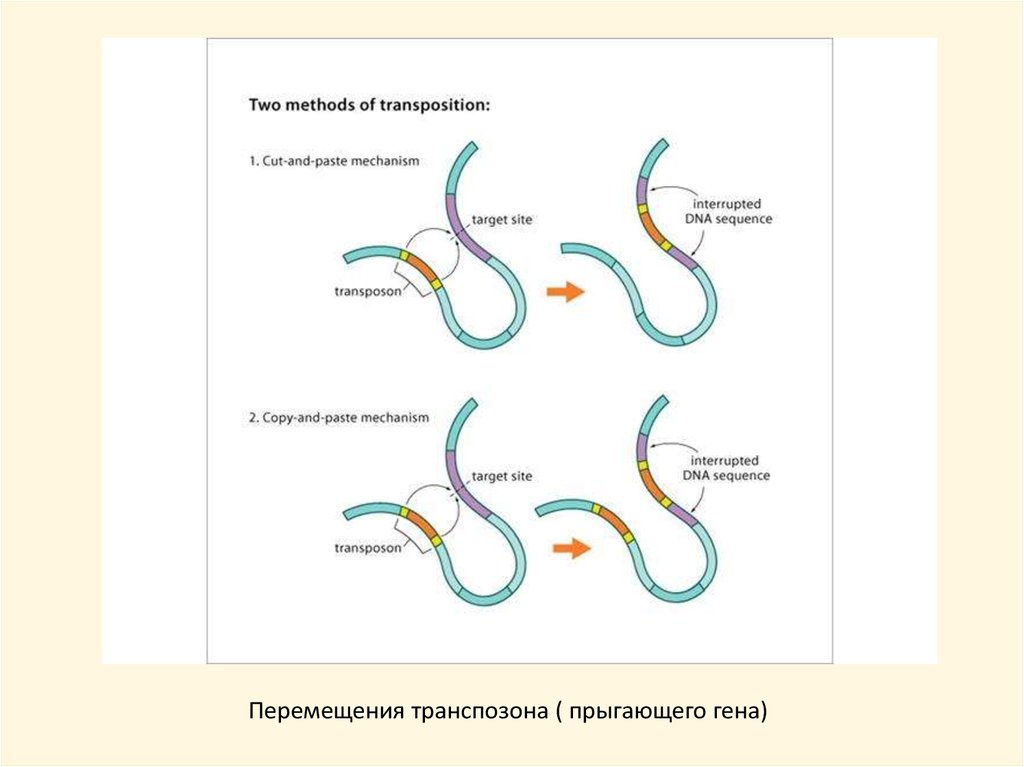
Перемещения транспозона ( прыгающего гена)23.
Характеристика хромосомных мутацийХромосомные перестройки – изменения в количестве и расположении
участков той или иной хромосомы. Если не летальны, то могут передаваться в
ряду поколений.
Последствия хромосомных перестроек могут быть весьма значительными:
• при делеции те гены, которые остались на неповрежденном гомологе,
проявляются в фенотипе
• при дупликации также изменяется соотношение тех аллелей, которые
находятся на дуплицированном локусе – могут проявиться в фенотипе даже
рецессивные аллели, если они удвоены
• при инверсии и транслокации проявляется «эффект положения» – те гены,
которые находились в составе гетерохроматина, после перестройки могут
попасть в активно работающий участок эухроматина, и тогда с них пойдет
транскрипция, и появятся те белки, которых в данной клетке быть не должно.
Может быть и прямо противоположная ситуация, тогда нужные гены работать
перестанут.
Иногда подобные перестройки активируют онкогены (см. выше пункт 3)
24.
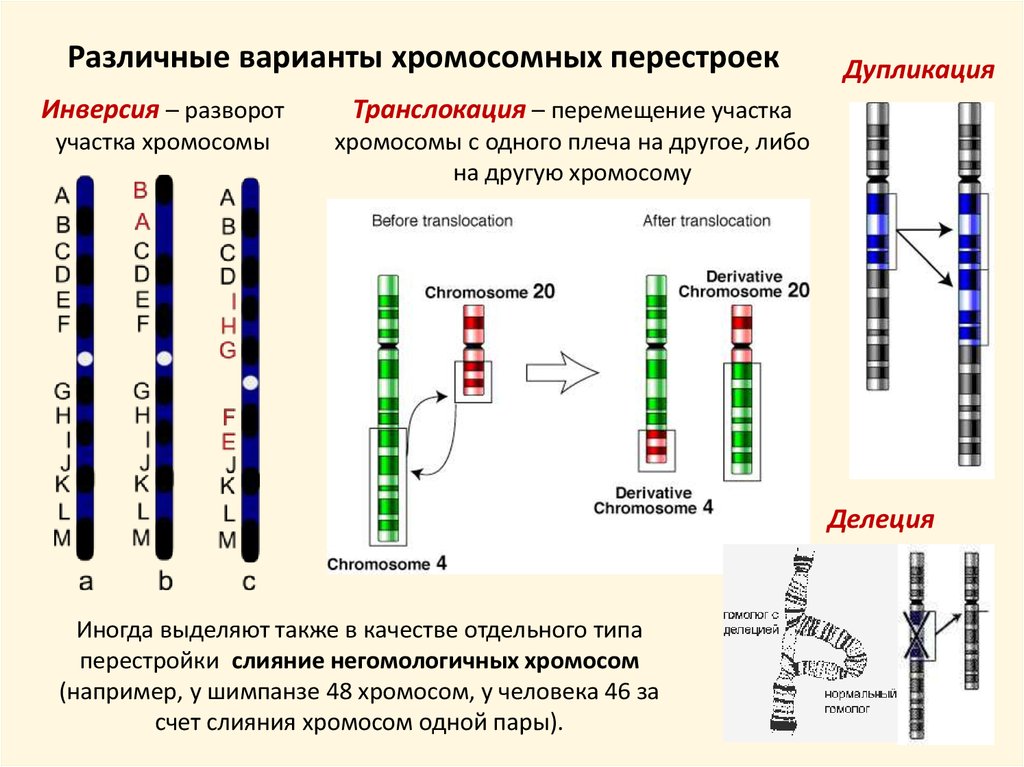
Различные варианты хромосомных перестроекИнверсия – разворот
Транслокация – перемещение участка
участка хромосомы
хромосомы с одного плеча на другое, либо
на другую хромосому
Дупликация
Делеция
Иногда выделяют также в качестве отдельного типа
перестройки слияние негомологичных хромосом
(например, у шимпанзе 48 хромосом, у человека 46 за
счет слияния хромосом одной пары).
25.
Характеристика геномных мутацийАнеуплоидия - изменение количества отдельных хромосом, обычно происходит в
результате неправильного расхождения хромосом при митозе в соматических клетках,
либо при мейозе в процессе гаметогенеза у животных. Моносомия по любой аутосоме
для клетки практически всегда летальна, трисомия приводит к серьезным нарушениям.
К геномным мутациям относят также увеличение числа наборов хромосом, в
результате чего организм становится полиплоидным (3n, 4n и т.д.). Такие мутации
широко распространены у растений – полиплоидны более трети из 300 тысяч видов
цветковых, а среди сельскохозяйственных культур - большинство. Обычно полиплоидия
приводит к увеличению вегетативной массы растений, увеличению скорости роста,
повышению устойчивости к неблагоприятным факторам (поэтому среди высокогорных
растений полиплоидны 80% видов). Однако у полиплоидных растений возникают
проблемы со спорообразованием, для которого требуется мейоз. Полиплоидность
мейоз затрудняет или даже исключает (если число наборов хромосом нечетное),
растения в этом случае размножаются вегетативно. Искусственный мутагенез для
получения полиплоидов широко используется в селекции. В Японии, в частности,
научились выращивать триплоидную форму арбузов, не имеющих семян - для этого
скрещивают особи с тетраплоидным и диплоидным набором. Полиплоидизация
помогает также преодолеть стерильность межвидовых гибридов
(работы Карпеченко Б.Д. – получен капустно-редечный гибрид)
Большинство животных и человек с такими мутациями жить не могут. Искусственно
были получены полиплоидные формы тутового шелкопряда для преодоления
стерильности межвидового гибрида (работы Астаурова Б.Л.)
26.
Для животных оптимальным является диплоидное состояниепо ряду причин:
• достаточная плоидность, чтобы в фенотипе не проявлялись вредные
рецессивные мутации – особи успешно выживают в процессе своего
онтогенеза; в генофонде популяции такие мутации хранятся в ряду
поколений, в новых условиях среды могут стать выгодными, особенно при
разнонаправленном отборе (серповидноклеточная анемия и др.)
• при диплоидности достаточно надежно сохраняются гены в поколениях с
помощью рекомбинационной репарации – она используется при особо
сложных повреждениях ДНК
• гетерозиготные комбинации разных аллелей создают внутривидовое
разнообразие, расширяют экологическую нишу вида
Важно отметить, что при гаплоидном состоянии скорость адаптивной
эволюции выше, чем при диплоидном, но для этого гаплоидность должна
сопровождаться высокой скоростью размножения и быстрой сменой
поколений (пример - бактерии)
27.
Закон гомологических рядов наследственной изменчивостиОстистые формы у разных
видов пшеницы (1-8) и
ячменя (9-12)
Н.И.Вавилов на основании изучения большого
количества различных видов растений в 1920 году
сформулировал Закон гомологических рядов в
наследственной изменчивости: «Виды и роды,
генетически близкие, характеризуются сходными
рядами наследственной изменчивости с такой
правильностью, что, зная ряд форм в пределах
одного вида, можно предвидеть нахождение
параллельных форм у других видов и родов».
С менее четкой формулировкой аналогичную
мысль высказывали и ранее – Ч.Дарвин и другие
ученые.
Закон гомологических рядов имеет
общебиологическое значение, справедлив для
всех живых организмов. На основании этого
закона можно предсказывать, какие мутантные
формы могут возникнуть у близкородственных
видов
28.
Дополнения- Устарело!
29.
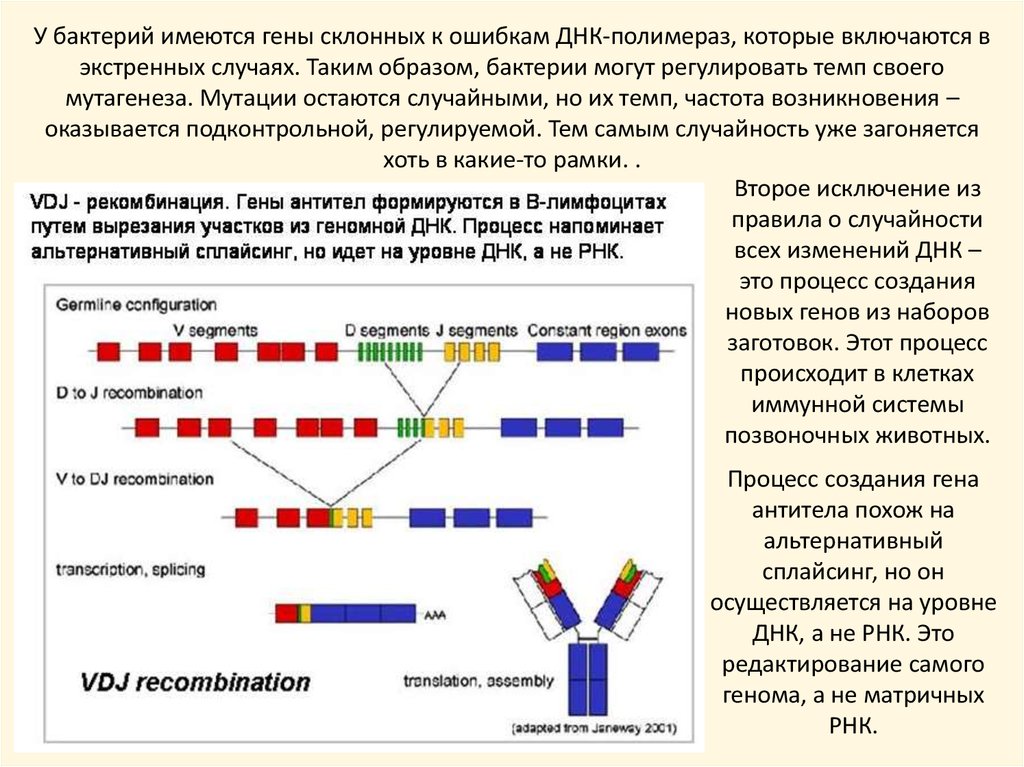
У бактерий имеются гены склонных к ошибкам ДНК-полимераз, которые включаются вэкстренных случаях. Таким образом, бактерии могут регулировать темп своего
мутагенеза. Мутации остаются случайными, но их темп, частота возникновения –
оказывается подконтрольной, регулируемой. Тем самым случайность уже загоняется
хоть в какие-то рамки. .
Второе исключение из
правила о случайности
всех изменений ДНК –
это процесс создания
новых генов из наборов
заготовок. Этот процесс
происходит в клетках
иммунной системы
позвоночных животных.
Процесс создания гена
антитела похож на
альтернативный
сплайсинг, но он
осуществляется на уровне
ДНК, а не РНК. Это
редактирование самого
генома, а не матричных
РНК.
30.
31.
32.
Возникающие вариации вирусов могут помогать друг другу за счет разделения функций. Например, в однойлинии может возникнуть мутация, повышающая скорость репликации вирусной РНК, в другой линии —
мутация, повышающая, например, эффективность самосборки белковых молекул в капсид. Поскольку обе
линии живут и размножаются в одной и той же хозяйской клетке, они могут пользоваться «достижениями»
друг друга: эффективная РНК-полимераза первой линии будет размножать и геномы второй, а более
эффективные капсидные белки второй линии будут «упаковывать» и РНК первой. Не исключено, что
совместить обе эти мутации в одном и том же геноме просто невозможно - вирусный геном очень мал
33.
34.
Вискаша – 102 хромосомы, уближайших родственных видов
– 56; результат полногеномной
дупликации до 112 хромосом,
затем потеря части хромосом
Тympanoctomys barrerae
•Мутации неизбежны.
•Вредные мутации, как правило,
возникают намного чаще, чем
полезные.
•Поэтому любая популяция
находится под угрозой
генетического вырождения.
•Главная защита от этой угрозы –
естественный отбор.
У животных и растений один из главных шаперонов называется Hsp-90 (от heat shock
protein). Установлено, что мутации, нарушающие работу Hsp90, приводят к
многочисленным аномалиям и уродствам. В основном эти аномалии представляют
собой проявления «скрытой изменчивости», то есть мутаций, которые уже
существовали в данной популяции, но не проявлялись, пока Hsp90 работал нормально.
Благодаря нормальной работе Hsp90 эти мутации не подпадали под действие отбора и
могли спокойно накапливаться, т.к. шаперон правильно укладывал даже белок с
нарушениями в первичной структуре.
35.
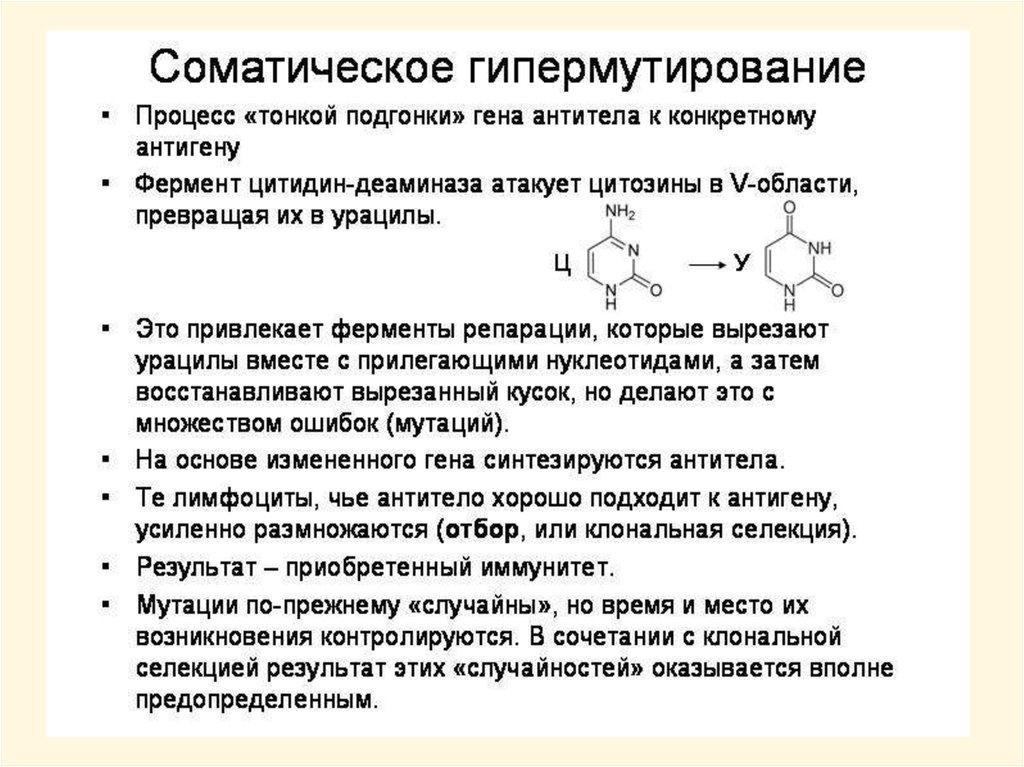
Когда вредных мутаций много, они не так вредныhttp://elementy.ru/news/165026
Эксперименты на червях C. elegans показали, что эффекты многих вредных мутаций
отрицательно коррелируют с силой реакции организма на перегрев. У особей,
энергично реагирующих на тепловой стресс, многие вредные мутации не проявляются
в фенотипе. Это объясняется тем, что белки-шапероны, вырабатывающиеся при
перегреве, помогают другим белкам восстановить «правильную» трехмерную
конфигурацию независимо от того, чем она была нарушена — перегревом или
мутацией. Кроме того, особи с сильной реакцией на стресс живут в среднем дольше.
Казалось бы, отбор должен был закрепить устойчиво высокий уровень реакции на
стресс, однако этот уровень сильно варьирует даже у генетически идентичных червей.
Как выяснилось, это связано с тем, что особи с сильной реакцией на стресс «платят» за
свои преимущества пониженной плодовитостью. Поэтому в переменчивых и
непредсказуемых условиях, характерных для природных местообитаний C. elegans,
оптимальной стратегией оказалось сохранение стохастической изменчивости по
данному признаку.
Снижение вредоносности мутаций по мере роста их числа может быть связано с
деятельностью белков-шаперонов, обеспечивающих правильное сворачивание
(укладку) других белковых молекул. Повышение числа мутантных (и потому
неправильно «свернутых») белков в клетке может привести к росту производства
шаперонов, которые иногда способны помочь даже «испорченному» белку свернуться
правильно. Для проверки этого предположения были измерены концентрации
шаперонов у бактерий с разным числом накопившихся мутаций. Оказалось, что в
линиях с большим числом мутаций уровень шаперонов действительно резко повышен.
36.
Всего известно 5000 наследственных заболеваний, из них 2000 - тяжелыхинвалидизирующих расстройств. Значительные успехи достигнуты в изучении
молекулярных причин наследственных заболеваний. Наследственные заболевания
связаны с изменением числа хромосом (табл. 1) или с мутациями. Большинство таких
мутаций на протяжении десятков тысяч лет передаются из поколения в поколение,
сохраняясь в популяции (табл. 2). Так, в случае заболевания фенилкетонурии частота
появления больных (гомозигот по рецессивному аллелю) составляет 1:15000
рождений, а частота гетерозиготного носительства болезнетворного аллеля - 1:62.
Именно поэтому генетический груз в популяции не уменьшается при таких
евгенических мерах, как уничтожение больных (как это происходило в античной
Спарте) или их стерилизации (как в США, где в первой половине ХХ века более 60 тыс.
человек было принудительно подвергнуто этой процедуре).
Для выявления болезнетворных мутаций исследуют семьи, в которых то или иное
заболевание встречается в нескольких поколениях. Для почти 500 заболеваний
найдены связанные с ними генами и выявлены мутации, приводящие к болезни.
Но иногда болезнетворные мутации возникают заново в клетках зародышевого пути.
Например, один из видов карликовости - ахондроплазия - встречается с частотой
1:100 000. В 80% случаев ахондроплазия вызывается вновь возникшей доминантной
мутацией (замене гуанина на цитозин), приводящей к замене аминокислоты глицина
на аргинин в рецепторе гормона роста. При этом ни у кого из родственников
больного нет этого заболевания. Эмбрионы, гомозиготные по данной мутации,
погибают.
37.
Другой механизм появления у детей заболевания, которого не было у их родителей накопление из поколения в поколение повторяющихся фрагментов ДНК внутринекоторых генов. К таким заболеваниям относится синдром ломкой Х-хромосомы. Это
заболевание проявляется уже в детстве и сопровождается нарушениями роста,
задержкой умственного развития, уходом ребенка от социальных контактов (аутизмом).
В одном из генов Х-хромосомы у больных обнаружено увеличенное число повторов
тринуклеотида ЦГГ. В норме число повторов не превышает 50, тогда как у больных оно
больше 90. Число повторов увеличивается при репликации хромосом.
Большинство вредных мутаций проявляется на ранних этапах развития - в детстве или
даже во внутриутробном периоде. Однако некоторые "поломки" генов могут не
проявляться до глубокой старости. Ученые нашли гены, мутации в которых связаны с
различными формами старческого слабоумия - болезнями Альцгеймера, Паркинсона,
Гентингтона. Наиболее часто встречающаяся болезнь Альцгеймера начинается в 60-80
лет с утраты памяти на недавние события и способности выполнять привычные
действия (одеваться, причесываться). Постепенно больной перестает узнавать своих
близких, забывает даже свое имя и через несколько лет умирает, находясь к этому
времени в совершенно беспомощном состоянии. Изучение найденных генов помогает
понять причины изменений работы мозга на молекулярном уровне и найти пути
лечения этой болезни. Одним из средств профилактики нарушения работы мозга в
старости является высокая интеллектуальная активность на протяжении всей жизни
человека.
38.
Данные о распространенности в России хромосомных болезней и врожденныхпороков развития (среди них имеются как наследственные, так и связанные с
неблагоприятными воздействиями на плод во время беременности) приведены в
табл. 3.
Не только наследственные, но и предрасположенность к инфекционным
заболеваниям имеет генетическую основу. Разные люди в разной степени
восприимчивы к различным инфекциям.
"Чума" ХХ века - СПИД - пока неизлечимое заболевание. Но некоторые люди (в Европе
около 1-2 %) невосприимчивы к вызывающему СПИД вирусу иммунодефицита
человека (ВИЧ) из-за мутации в гене хемокинового рецептора. Хемокиновый рецептор
расположен на поверхности клеток и служит "посадочной площадкой" для вируса
СПИДа. В отсутствии этого белка вирус, попав в организм, не способен проникнуть
внутрь клетки и не приводит к заболеванию. Описаны и другие мутации, приводящие
к повышенной устойчивости к ВИЧ.
Другой пример - эпидемия "коровьего бешенства" (губчатая болезнь мозга),
распространившаяся в Англии в связи с изменением технологии приготовления
костной муки, используемой в качестве кормовой добавки для скота (была снижена
температура обработки) и поразившая 160 000 животных. Пик эпидемии пришелся на
1992 г. Заболевание вызывается белком-прионом - удивительным инфекционным
агентом, не содержащим ДНК и вызывающим изменение конформации клеточных
белков и нарушение функций клеток нервной системы. Хотя зараженную говядину
употребляли многие, только два десятка человек заболели. Все заболевшие люди
имели одну и ту же мутацию, известную ранее и считавшуюся нейтральной.
39.
В последние годы особое внимание уделяют распространенности в различныхпопуляциях аллелей генов предрасположенности к тем или иным заболеваниям и так
называемых генов "внешней среды". В частности, это гены, контролирующие
детоксикацию чужеродных веществ. Все вещества, посутупающие в организм,
метаболизируются в два этапа. На первом этапе образуются промежуточные
генотоксические вещества. На втором этапе эти промежуточные метаболиты
превращаются в растворимые безвредные соединения, которые выводятся из
организма.
То, как организм реагирует на вредные воздействия среды, например, на табачный
дым, также в значительной мере определяется активностью системы детоксикации.
Например, у 3% женщин встречается сочетание генов, ответственных за
обезвреживание канцерогенов табачного дыма ферментами печени, которое в 10
раз повышает риск развития рака молочной железы. Таким женщинам курить строго
противопоказано. Влияют гены и на эффективность лечения различными
препаратами. Так, лечение эндометриоза - заболевания, встречающегося почти у 10%
женщин белой расы - широко используемым препаратом циклофероном у части
больных безрезультатно по причинам генетического характера.
С частотой 15% встречается аллель гена аполипопротеина Е (белок, отвественный за
связывание липидов), приводящий к ухудшению регенерации нервных тканей после
травм или сотрясений мозга, особенно если индивид гомозиготен по данному аллелю.
Значит, таким людям не стоит выбирать профессию боксера или автогонщика.