














































 biology
biologySimilar presentations:










Гормональная регуляция как механизм межклеточной и межорганной координации обмена веществ
1.
Лекция 15:Гормональная регуляция как механизм межклеточной и
межорганной координации обмена веществ. Механизм
передачи гормонального сигнала в клетку. Инсулин, роль
гормона.
Дисциплина: Биохимия
для специальности 31.05.01 Лечебное дело
ИФОИТМ ТГМУ
доц. О. А. Артюкова
2.
Основные вопросы, рассматриваемые на лекции:■
■
■
■
■
■
■
■
Роль гормонов в системе регуляции метаболизма, клетки-мишени и клеточные
рецепторы гормонов. Строение G-белков.
Передача гормонального сигнала в клетку. Образование вторичных посредников.
Метаболические изменения в ответ на сигнал.
Классификация гормонов по химическому строению, механизму действия и
биологическим функциям.
Гормоны гипоталамуса: либерины и статины. Гормоны гипофиза.
Регуляция энергетического метаболизма, роль инсулина и контринсулярных
гормонов в обеспечении гомеостаза в постабсорбтивный период и при голодании.
Изменение гормонального статуса и метаболизма при сахарном диабете.
Патогенез основных симптомов сахарного диабета, диабетичеcкой комы.
Патогенез поздних осложнений сахарного диабета.
3.
Для нормального функционирования многоклеточногоорганизма необходима взаимосвязь между отдельными клетками, тканями, органами.
Эту связь осуществляют:
Все уровни регуляции в организме интегрированы и действуют как единое целое!
4.
Системы регуляции обмена веществ и метаболизма клетки образуют3 иерархических уровня:
ЦНС
Эндокринная система –
гормоны
Внутриклеточные механизмы регуляции:
1. Изменение активности ферментов
2. Изменение количества ферментов
3. Изменение скорости мембранного транспорта
5.
Гормоны - сигнальные молекулы межклеточного общения!Гормоны (греч. hormao – привожу в движение) – вещества, вырабатываемые
специализированными клетками, регулирующие обмен веществ в отдельных
органах и во всем организме в целом. Синтез и секреция гормонов
стимулируется внешними и внутренними сигналами, поступающими в ЦНС.
6.
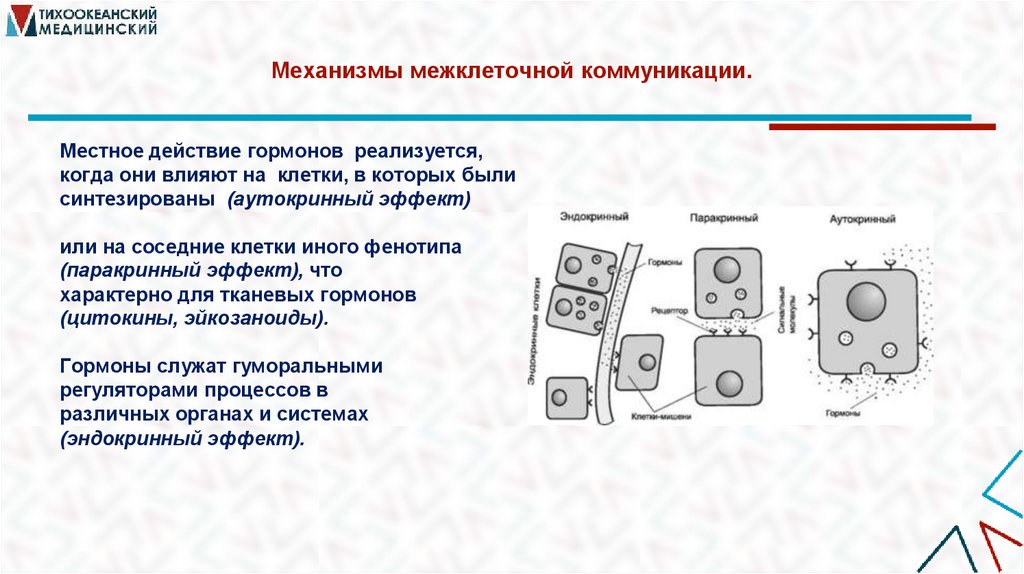
Механизмы межклеточной коммуникации.Местное действие гормонов реализуется,
когда они влияют на клетки, в которых были
синтезированы (аутокринный эффект)
или на соседние клетки иного фенотипа
(паракринный эффект), что
характерно для тканевых гормонов
(цитокины, эйкозаноиды).
Гормоны служат гуморальными
регуляторами процессов в
различных органах и системах
(эндокринный эффект).
7.
Интегральные свойства гормонов8.
Гормоны - внеклеточные химические сигналы (первичные мессенджеры).Особенности действия гормонов:
● Действуют в малых количествах (10-6-10-12 ммоль/л).
● Обладают беспороговым принципом действия
(даже 1 молекула гормона способна оказать эффект!)
● Обладают дистантным характером действия.
● Обладают высокой специфичностью действия.
● ереносят только информацию!
(не используются в энергетических и строительных целях).
9.
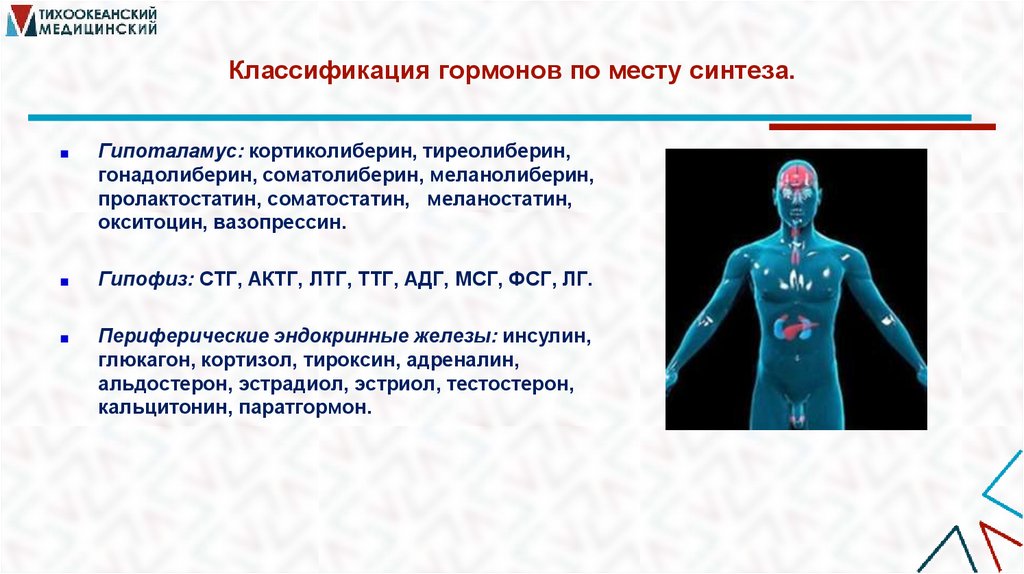
Классификация гормонов по месту синтеза.■
Гипоталамус: кортиколиберин, тиреолиберин,
гонадолиберин, соматолиберин, меланолиберин,
пролактостатин, соматостатин, меланостатин,
окситоцин, вазопрессин.
■
Гипофиз: СТГ, АКТГ, ЛТГ, ТТГ, АДГ, МСГ, ФСГ, ЛГ.
■
Периферические эндокринные железы: инсулин,
глюкагон, кортизол, тироксин, адреналин,
альдостерон, эстрадиол, эстриол, тестостерон,
кальцитонин, паратгормон.
10.
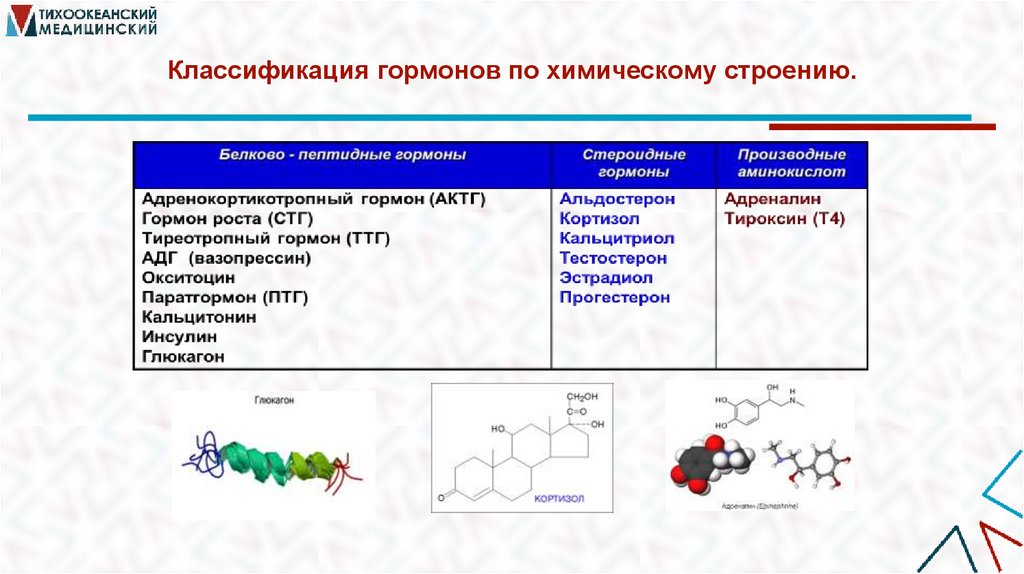
Классификация гормонов по химическому строению.11.
Метаболизм гормонов зависят от природы гормона.Обмен белковых (пептидных) гормонов.
■
■
■
■
■
Инсулин
Синтез: в неактивной форме (прогормон).
Активация: частичный протеолиз.
Хранение: накапливаются и хранятся в секреторных гранулах (5 дней)
Время действия: мин–час.
Циркуляция: хорошо растворимы в крови.
Инактивация: гидролиз до аминокислот в печени.
12.
Метаболизм гормонов зависят от природы гормона.Обмен стероидных гормонов.
■
■
■
■
Синтез: производные холестерина.
Время действия: часы.
Циркуляция: транспортируются
специфическими белками крови.
Инактивация: окисление в печени.
13.
Метаболизм гормонов зависят от природы гормона.Обмен гормонов - производных аминокислот.
■
■
■
■
■
■
■
■
■
Адреналин
Синтез: в мозговом слое надпочечников из ТИР.
Время действия: миллисек.
Хранение: происходит в секреторных гранулах.
Транспорт: растворим в крови.
Инактивация: при участии специфических ферментов.
Тиреоидные гормоны
Синтез: в клетках щитовидной железы из ТИР.
Время действия: несколько суток.
Хранение: накапливаются в коллоиде железы.
Транспорт: специфическими белками крови.
Инактивация происходит длительно в результате
дейодирования гормонов.
14.
Эйкозаноиды (простагландины, тромбоксаны, лейкотриены) –производные арахидоновой кислоты
Арахидоновая кислота (С20:4) входит в
состав мембранных фосфолипидов.
15.
Эйкозаноиды - большая группа паракринных гормонов.Эйкозаноиды действуют на клетки-мишени через мембранные рецепторы
и механизм образования внутриклеточного вестника сигнала:
ц-АМФ, ц-ГМФ, инозитолтрифосфата, Са2+.
В разных тканях и ситуациях образуются неодинаковые эйкозаноиды.
Эйкозаноиды выполняют разнообразные физиологические функции:
■
сокращение /расслабление гладких мышц,
■
агрегация тромбоцитов,
■
расширение /сокращение сосудов,
■
развитие воспалительного процесса.
■
Ингибитор циклооксигеназы (ЦОГ) – аспирин
тормозит синтез эйкозаноидов
(противовоспалительное средство).
16.
Классификация гормонов по биологическому действию.17.
Клетка-мишень для гормона определяется по наличиюрецептора для сигнальной молекулы (гормона).
Рецепторы – белки, комплементарные
сигнальной молекуле, позволяют клетке
получать инструкции от гормонов.
Число рецепторов гормонов варьирует
от 500 до 100 000 на клетку.
Рецепторы передают полученную информацию
через вторичные мессенджеры в клетке системе
белков и ферментов, которые образуют каскад
реакций, обеспечивающий усиление
сигнала в несколько сот раз.
В зависимости от природы гормона
и локализации рецептора различают
следующие механизмы передачи
гормонального сигнала:
1. мембранно–внутриклеточный,
2. цитозольный.
18.
Особенности мембранно-внутриклеточного механизмапередачи гормонального сигнала.
Механизм характерен для гормонов
белково-пептидной природы,
адреналина, эйкозаноидов.
Гормоны в клетку не проникают!
В клетке образуются вторичные
вестники (мессенджеры):
ц-АМФ, ц-ГМФ, Са 2+ и др.
Метаболические эффекты гормонов
реализуются через повышение
активности ферментов в клетке.
19.
Рецепторы, встроенные в мембрану клеток.Рецепторы
Сопряженные
с G – белками
Аденилатциклазна
я
система 3
Гуанилатциклаза
Инозитолфосфатная
система 4
Каталитические
2
20.
Трансмембранная передача гормонального сигнала,сопряженная с G–белками (G-proteins).
G-белки - это сигнальные белки, которые являются
универсальными посредниками при передаче
гормональных сигналов к эффекторным белкам,
вызывающим конечный клеточный ответ.
G-белки состоят из 3 субъединиц – αßγ.
G-белки - семейство белков, относящихся к ГТФ-азам.
G-белки используют замену ГДФ на ГТФ
как молекулярный «включатель» для
регулировки клеточных процессов.
G-белки были обнаружены и изучены
А. Гилманом и М. Родбеллом
(Нобелевская премия по физиологии и
медицине 1994 г.)
21.
Аденилатциклаза катализирует превращение АТФ в ц-АМФ.ц-АМФ является вторичным мессенджером.
ц-АМФ используется для внутриклеточного
распространения сигналов тех гормонов,
которые не могут проходить через клеточную
мембрану.
Сазерленд Эрл Уилбур (1915- 1974) -лауреат
Нобелевской премии по физиологии и медицине
(1971). Открыл в животных тканях ц–АМФ и
установил его роль в проведении гормонального
сигнала в клетку.
22.
G-белки стимулируют аденилатциклазу.Рецептор имеет центры связывания гормона
на наружной поверхности мембраны и G-белка
на внутренней поверхности мембраны. Взаимодействие
приводит к изменению конформации рецептора.
Снижается сродство α-субъединицы к ГДФ и
увеличивает сродство к ГТФ(ГДФ замещается ГТФ).
Отделившаяся субъединица α-ГТФ перемещается
в липидном слое мембраны к аденилатциклазе.
Взаимодействие α-ГТФ с регуляторным центром
аденилатциклазы изменяет конформацию фермента и
увеличивает скорость образования ц-АМФ из АТФ
(концентрация ц-АМФ в клетке возрастает).
23.
Последовательность процессов трансмембранной передачигормонального сигнала через аденилатциклазную систему.
α-ГТФ мигрирует к аденилатциклазе (АЦ).
АЦ увеличивает скорость образования
ц-АМФ из АТФ.
ц-АМФ – вторичный вестник гормонального
сигнала в клетке.
4 мол. ц-АМФ активируют протеинкиназу А
(происходит диссоциация R и С-протомеров).
Протеинкиназа А (ПК А) фосфорилирует
ферменты, их активность повышается.
На каждом этапе каскада происходит
значительное усиление сигнала.
24.
Последовательность процессов трансмембранной передачигормонального сигнала через инозитолфосфатную систему.
Взаимодействие α-ГТФ с центром связывания
фосфолипазы С изменяет конформацию и активность
фермента, возрастает скорость гидролиза фосфолипида
мембраны - фосфатидилинозитол-4,5-бисфосфата (ФИФ2).
Образуются 2 продукта (вторичные мессенджеры):
диацилглицерол (ДАГ), который остается в мембране
и участвует в активации фермента протеинкиназы С;
инозитол-1,4,5-трифосфат (ИТФ, ИФ3), который
уходит в цитозоль.
25.
Последовательность процессов трансмембранной передачигормонального сигнала через инозитолфосфатную систему.
Инозитол-1,4,5-трифосфат (ИТФ, ИФ3) вторичный вестник гормонального
сигнала в клетке.
ИФ3 связывается со специфическими
центрами Са2+-канала мембраны.
Ионы Са2+ поступают в цитозоль.
26.
Последовательность процессов трансмембранной передачигормонального сигнала через инозитолфосфатную систему.
Повышение концентрации Са2+ в цитозоле
увеличивает скорость взаимодействия Са2+
с неактивным ферментом протеинкиназой С (ПК С)
и белком кальмодулином.
Кальмодулин функционирует как внутриклеточный
рецептор, он имеет 4 центра связывания Са2+.
Взаимодействие комплекса [кальмодулин] - [4 Са2+]
с различными белками и ферментами приводит
к их активации путем фосфорилирования.
На каждом этапе каскада происходит усиление
гормонального сигнала в клетке.
27.
Последовательность процессов трансмембранной передачигормонального сигнала через гуанилатциклазу.
Присоединение гормона к рецептору приводит
к возрастанию уровня ц–ГМФ в клетке.
ц-ГМФ – вторичный вестник (мессенджер)
гормонального сигнала!
ц-ГМФ активирует протеинкиназу G (ПК G),
которая фосфорилирует специфические белки,
участвующие в реализации гормонального
сигнала. Одним из субстратов ПК G является
предсердный натриуретический фактор (ПНФ),
регулирующий гомеостаз жидкости
в организме.
28.
Трансмембранная передача гормонального сигнала.Каталитические рецепторы (рецептор инсулина)
Каталитические рецепторы являются ферментами. Их активаторами могут быть
белковые гормоны, цитокины. В активной форме рецепторы-ферменты
фосфорилируют специфические белки по ОН-группам тирозина (тирозиновые
протеинкиназы).
Рецептор инсулина построен из 2α- и 2 β-субъединиц.
■
α-субъединицы расположены на наружной поверхности мембраны,
они связывают инсулин.
■
Каталитический центр находится на внутриклеточных доменах β-субъединиц.
29.
Каталитические рецепторы (рецептор инсулина)Цитозольная часть рецептора имеет несколько остатков
тирозина, которые могут фосфорилироваться и
дефосфорилироваться (АТФ). Фосфорилирование
приводит к изменению заряда, конформации и
субстратной специфичности фермента
(тирозиновая протеинкиназа, Тир-ПК).
Тирозиновая-ПК фосфорилирует определенные
клеточные белки и ферменты, т.к.
• фосфопротеинфосфатазы,
• фосфодиэстеразы,
• ГЛЮТ- 4
• регуляторные белки ядра
(факторы транскрипции, повышающие/снижающие
экспрессию генов определенных ферментов).
30.
Передача сигнала с помощью внутриклеточных рецепторовСтероидные и тиреоидные гормоны могут
проходить через плазматическую мембрану клетки.
Гормон связывается с белком-рецептором в
цитозоле клетки.
Комплекс гормон-рецептор проходит в ядро и
взаимодействует с энхансерами /сайленсерами в ДНК,
что приводит к увеличению/уменьшению скорости
транскрипции м-РНК.
В клетке увеличивается/уменьшается
скорость синтеза белков, ферментов.
31.
Влияние гормонов на метаболические процессы в клетке-мишени.32.
Взаимосвязь ЦНС и эндокринной регуляции.Большинство гормональных систем взаимосвязаны между собой и
регулируются по механизму обратной связи.
ЦНС воспринимают сигналы из внешней и внутренней среды.
По нейронам нервных клеток сигналы поступают в гипоталамус,
где синтезируются гормоны пептидной природы рилизинг-факторы (либерины, статины), влияющие
на образование тропных гормонов гипофиза, которые
стимулируют периферические железы.
Изменение концентрации метаболитов в клетках-мишенях/крови
по механизму отрицательной обратной связи подавляет
синтез гормонов, действуя либо на периферические железы,
либо на синтез гипоталамо-гипофизарных гормонов.
33.
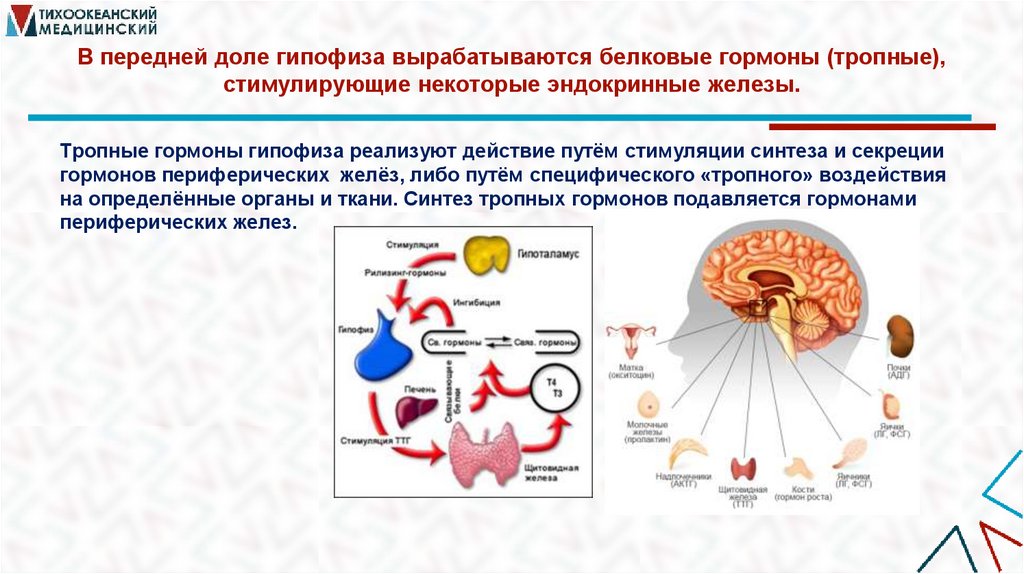
В передней доле гипофиза вырабатываются белковые гормоны (тропные),стимулирующие некоторые эндокринные железы.
Тропные гормоны гипофиза реализуют действие путём стимуляции синтеза и секреции
гормонов периферических желёз, либо путём специфического «тропного» воздействия
на определённые органы и ткани. Синтез тропных гормонов подавляется гормонами
периферических желез.
34.
Причины развития регуляторных (гормональных) нарушений.35.
Биохимические методы диагностики гормональных нарушений.36.
Гормональная регуляция метаболизма основных энергетических субстратов(белков, жиров, углеводов)
Функциональная активность тканей организма требует затрат энергии,
которая образуется в процессе катаболизма компонентов пищи.
37.
Регуляция метаболизма энергоносителей в абсортивный период.После приема пищи в крови повышается
концентрация глюкозы, аминокислот, ХМ.
Стимул к секреции инсулина.
Повышение индекса инсулин/глюкагон.
Инсулин способствует поступлению:
- глюкозы в инсулинозависимые ткани
(мышцы, жировая) через ГЛЮТ- 4;
- жирных кислот из ХМ в разные ткани.
38.
Анаболическое действие инсулина(стимуляция процессов в органах и тканях):
39.
Регуляция метаболизма энергоносителей в постабсортивный период.Переваривание и всасывание углеводов заканчивается
через 2 час. после приема пищи, белков и жиров –
через 4–6 час. Время между завершением пищеварения
и следующим приемом пищи - постабсортивный
период (от4 до 12 часов после приема пищи).
Уровень глюкозы в крови снижается.
Снижается индекс инсулин/глюкагон
Основные изменения метаболизма происходят в печени и жировой ткани,
они направлены на пополнение глюкозы крови за счет резервов углеводов
(гликогенолиз, глюконеогенез) и на использование других субстратов
(ТАГ, аминокислоты).
40.
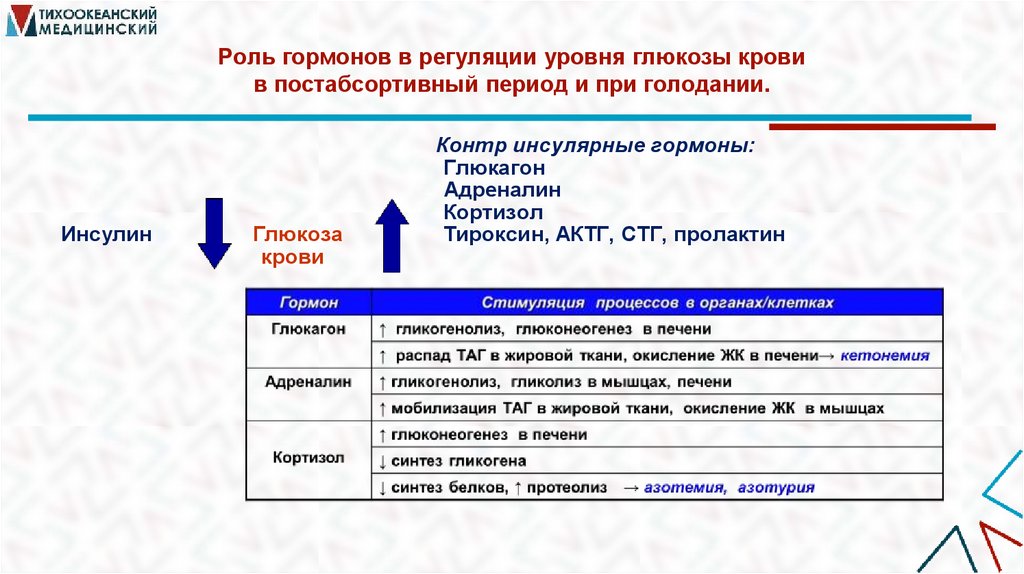
Роль гормонов в регуляции уровня глюкозы кровив постабсортивный период и при голодании.
Инсулин
Глюкоза
крови
Контр инсулярные гормоны:
Глюкагон
Адреналин
Кортизол
Тироксин, АКТГ, СТГ, пролактин
41.
Изменение метаболизма основных энергоносителей при голодании.В отсутствие пищи в крови снижается уровень глюкозы, аминокислот, ТАГ.
Инсулин/глюкагоновый индекс ↓, уровень контринсулярных гормонов ↑.
В этих условиях преобладают процессы катаболизма жиров, гликогена и
белков на фоне общего снижения скорости метаболизма.
Под влиянием контринсулярных гормонов происходит обмен субстратами между
печенью, жировой тканью, мышцами и мозгом. Этот обмен служит двум целям:
1) поддержанию концентрации глюкозы в крови для обеспечения
глюкозозависимых тканей (мозг, эритроциты);
2) мобилизации других источников энергии (ТАГ) для
обеспечения энергией всех других тканей (мышцы и др.).
42.
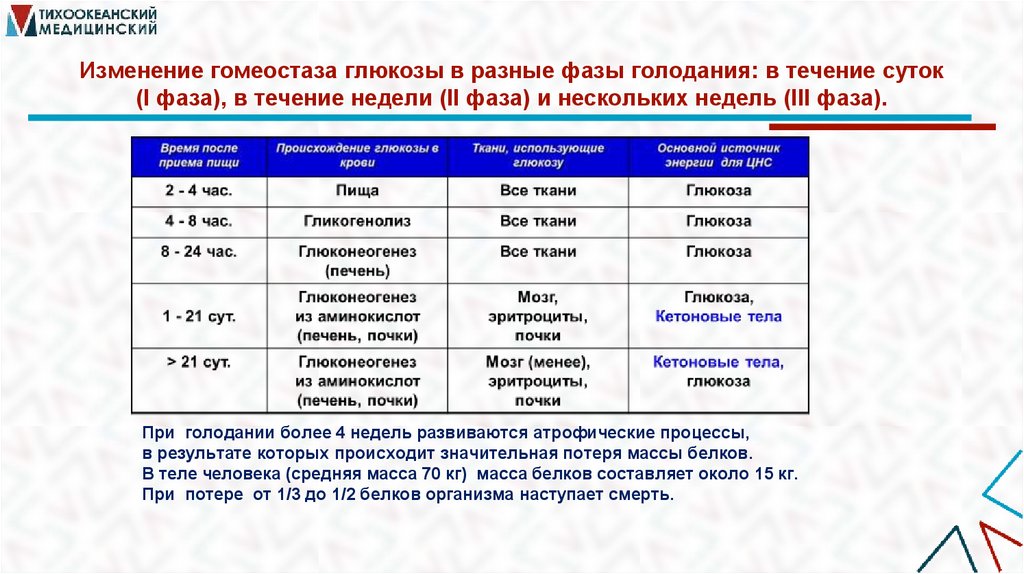
Изменение гомеостаза глюкозы в разные фазы голодания: в течение суток(I фаза), в течение недели (II фаза) и нескольких недель (III фаза).
При голодании более 4 недель развиваются атрофические процессы,
в результате которых происходит значительная потеря массы белков.
В теле человека (средняя масса 70 кг) масса белков составляет около 15 кг.
При потере от 1/3 до 1/2 белков организма наступает смерть.
43.
Изменение метаболизма основных энергоносителей при голодании.44.
Сахарный диабет (СД) –хроническое полиэтиологичное заболевание, характеризующееся глубокими
нарушениями углеводного, белкового, жирового и водно-солевого обменов.
ИЗСД возникает вследствие дефицита
инсулина, вызванного аутоиммунным
разрушением β-клеток поджелудочной
железы.
ИЗСД сопровождается тяжёлыми
метаболическими нарушениями,
развитием кетоацидоза и комы.
ИЗСД поражает в большинстве случаев
детей, но может проявиться в любом
возрасте.
ИНСД представляет собой
группу гетерогенных нарушений
углеводного обмена
(нарушение толерантности к глюкозе,
инсулинорезистентность).
Поражает людей старше 40 лет,
преимущественно жителей
развитых стран, горожан.
45.
Инсулинорезистентность –нарушение метаболического ответа на эндогенный или экзогенный инсулин.
В основе инсулинрезистентности могут лежать разные причины:
■
генетические дефекты рецепторов инсулина,
■
нарушении превращения проинсулина в инсулин,
■
повреждении инсулина и рецепторов антителами,
■
действия контринсулярных гормонов.
46.
Изменение метаболизма при сахарном диабете.При диабете как I типа, так и II типа
индекс инсулин/глюкагон снижается.
Это приводит к тому, что даже в абсортивный период метаболизм
энергоносителей осуществляется в режиме голодания!
47.
Изменение метаболизма при сахарном диабете.Причины развития гипергликемии (гиперглюкоземии)/глюкозурии:
■
■
■
■
■
■
уменьшение поступления глюкозы в инсулинзависимые ткани;
торможение синтеза гликогена в печени и мышцах;
снижение синтеза ТАГ из глюкозы в печени и жировой ткани;
ускорение глюконеогенеза из аминокислот в печени.
повышение концентрации глюкозы в крови (гиперглюкоземия).
Когда уровень глюкозы крови превысит почечный порог (9-10 ммоль/л),
начинается выделение глюкозы с мочой (глюкозурия).
48.
Изменение метаболизма при сахарном диабете.Причины развития кетонемии/кетонурии:
Глюкагон ускоряет липолиз в жировой ткани.
Увеличивается поступление жирных кислот в печень.
Окисление жирных кислот в печени и накопление ацетил~КоА.
Синтез кетоновых тел в печени и повышение их в крови.
Кетонемия (диабетический кетоацидоз!)
Кетонурия
49.
Изменение метаболизма при сахарном диабете.Причины развития азотемии/азотурии:
Снижение синтеза белка и активация протеолиза в ряде тканей
(мышечная, соединительная, костная, лимфоидная)
под влиянием глюкокортикостероидов (кортизол)
Поступление аминокислот в кровь и печень.
Дезаминирование аминокислот и активация глюконеогенеза.
Активация синтеза мочевины в печени (орнитиновый цикл ↑)
Азотемия
Азотурия
50.
Изменение метаболизма при сахарном диабете.Причины развития полифагии, полидипсии, полиурии:
Реакции глюконеогенеза протекают с участием оксалоацетата (ЩУК).
ЩУК активно вовлекается в глюконеогенез. В результате
цикл Кребса тормозится, снижается образование АТФ,
возникает энергодефицит, который компенсируется
повышенным потреблением пищи
Полифагия
Выведение из организма с мочой избытка глюкозы, кетоновых тел и
мочевины (осмотически активные соединения) сопровождается
потерей большого количества жидкости, что компенсируется
повышением потребления воды
Полиурия (3 - 4 л/сут), полидипсия (жажда)
51.
Поздние осложнения сахарного диабета.Неферментативное гликозилирование белков.
Причина гликозилирования- избыточная глюкоза крови
взаимодействует с NH2-группами белков;
белки изменяют свою конформацию и функции.
■
■
■
Примеры:
При гликозилировании кристаллинов прозрачность хрусталика глаза
уменьшается, возникает помутнение - катаракта.
При гликозилировании белков базальных мембран нарушается
их структура, происходит их утолщение и развитие ангиопатий
(повреждения капилляров, мелких и крупных сосудов).
При гликозилировании гемоглобина нарушается
транспорт О2 и развивается гипоксия.
52.
Острые осложнения сахарного диабета.Кома - угрожающее жизни состояние,
характеризующееся полной утратой сознания.