
















 biology
biologySimilar presentations:
")


")






Анаболизм углеводов (лекция № 5)
1.
Лекция №5.Тема: Анаболизм углеводов.
2.
Вопросы лекции №5.1.
Глюконеогенез.
2.
Регуляция глюконеогенеза.
3.
Энергетический баланс глюконеогенеза.
4.
Гликогеногенез.
5.
Мобилизация гликогена.
6.
Роль гормонов в регуляции резервирования и
мобилизации гликогена.
3.
4.
5.
6.
7.
Организационные принципы анаболизма:1
• пути синтеза и расщепления биомолекул не
идентичны ( всегда имеется хотя бы одна
ферментативная реакция по которой они
отличаются);
2
• анаболические и катаболические пути
регулируются
разными
регуляторными
ферментами;
3
• биосинтетические процессы ( идущие с
затратой энергии) связаны с расщеплением
АТФ (необратимость процесса)
8.
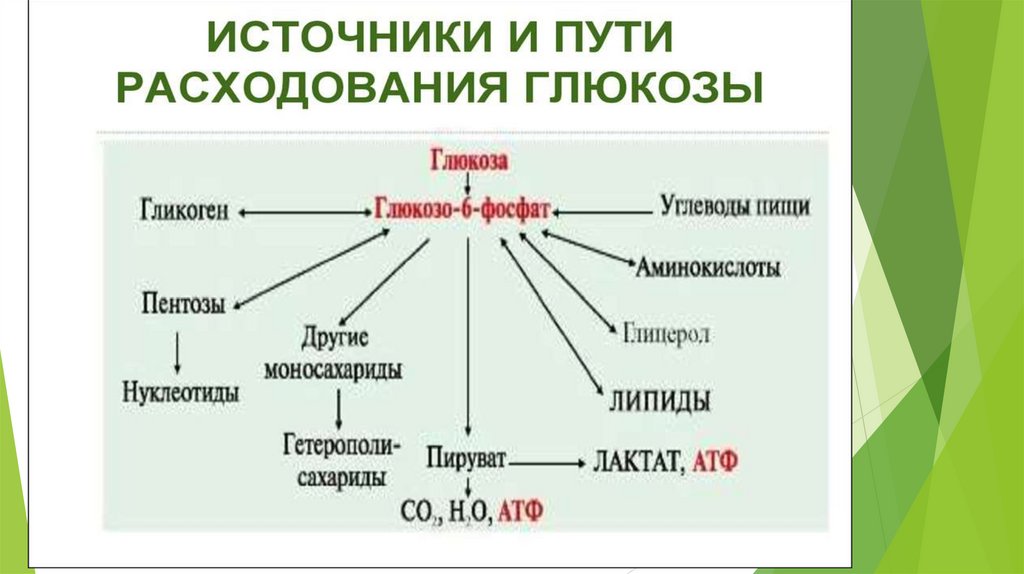
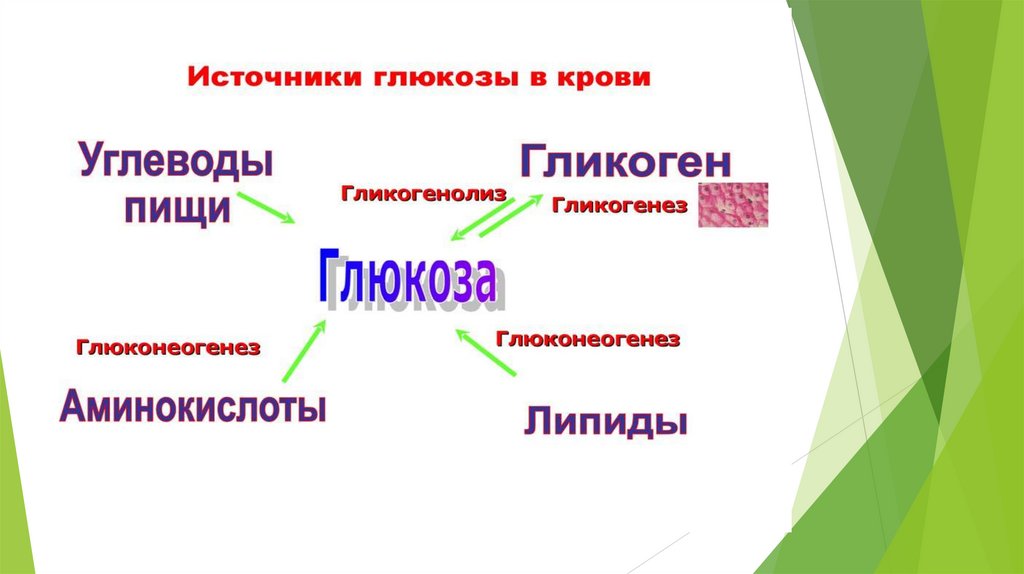
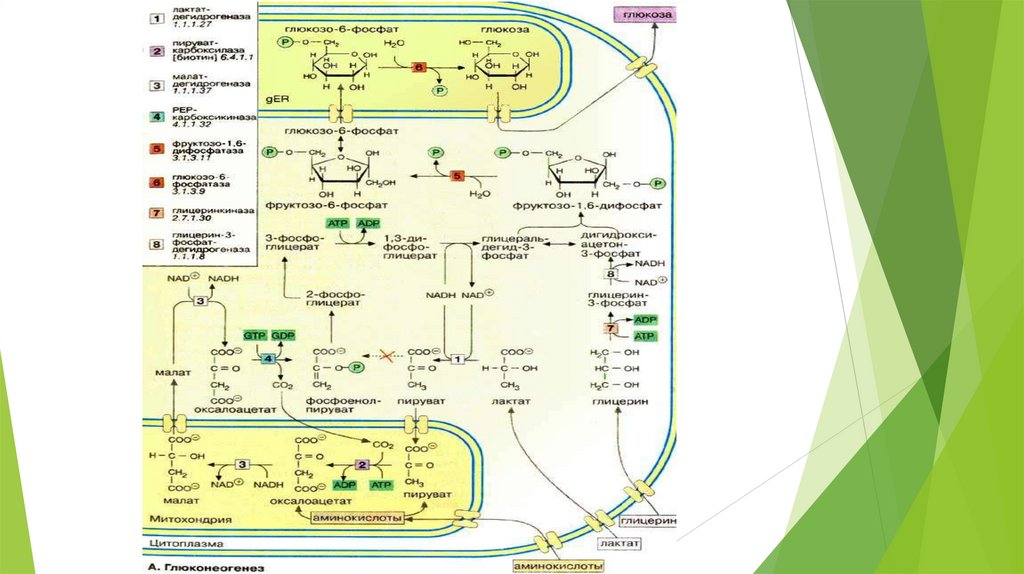
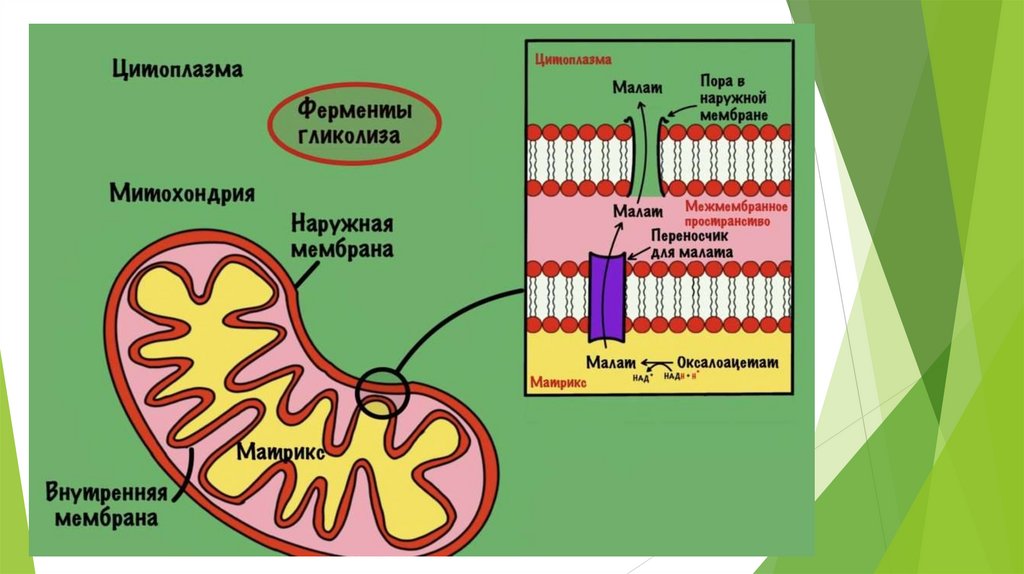
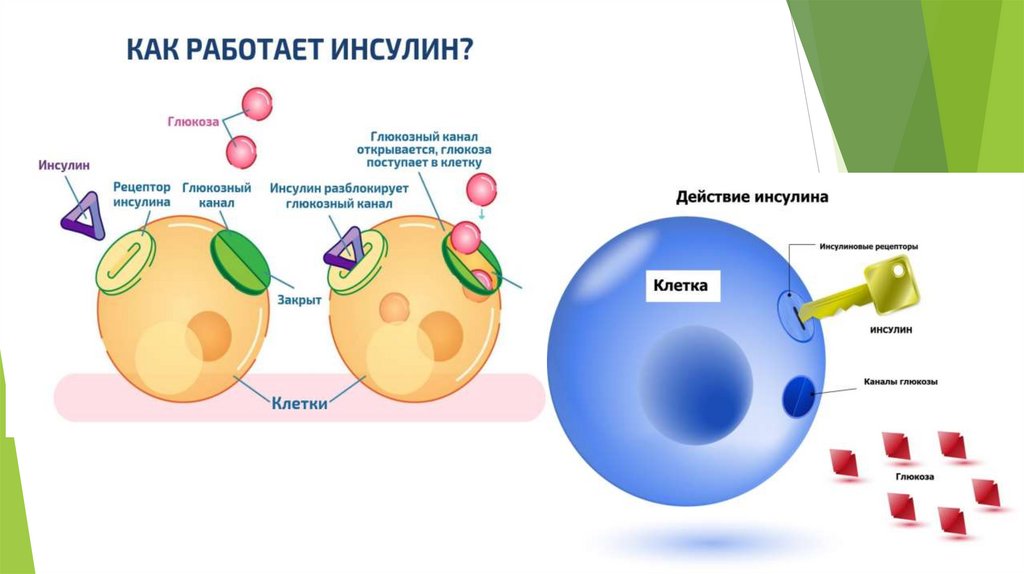
ГлюконеогенезD-глюкоза крови – главный источник топлива для нервной ткани, почек,
эритроцитов и т.д.
Не протекает в поперечно-полосатых мышцах, так как там нет глюкозо6-фосфатазы.
В сутки мозг человека потребляет 120 гр. глюкозы.
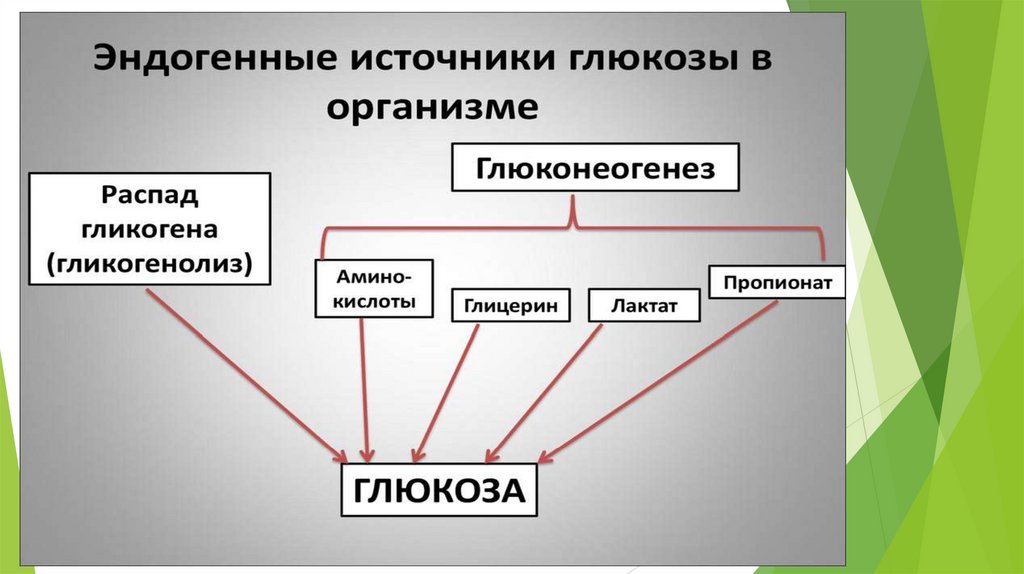
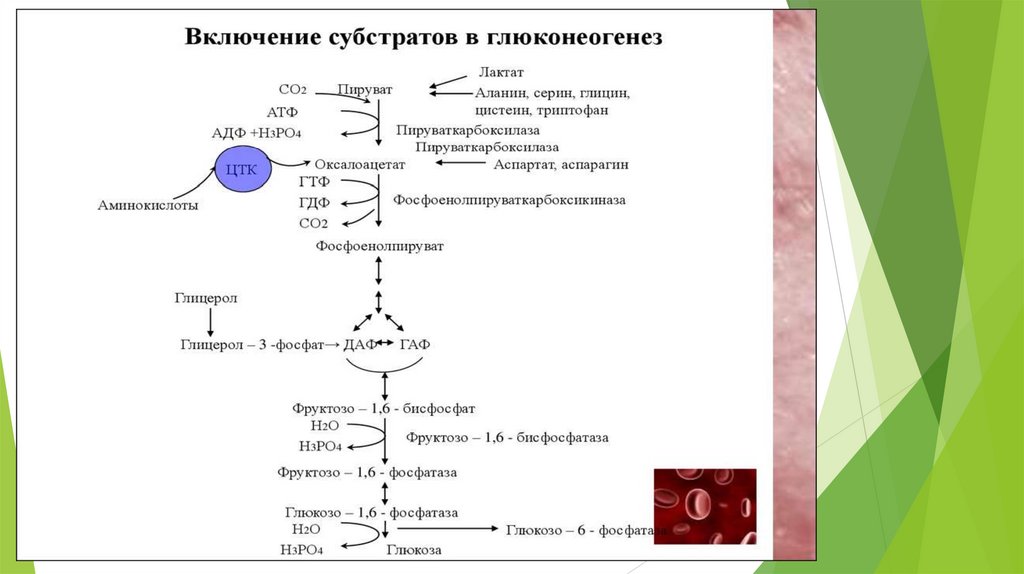
Глюконеогенез – синтез глюкозы из неуглеводных предшественников.
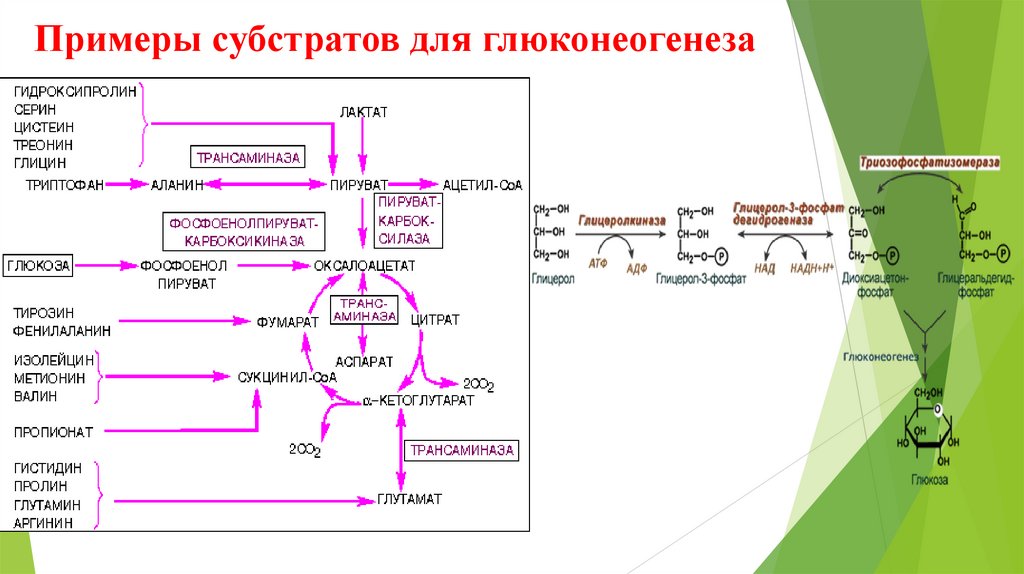
Предшественники глюкозы: пируват, лактат, глюкогенные аминокислоты,
глицерол, промежуточные продукты ЦТК.
Локализация: печень ( 80%), корковое вещество почек (всего 15%.),
слизистая кишечника. Эти ткани могут обеспечивать синтез 80-180 г
глюкозы в сутки.
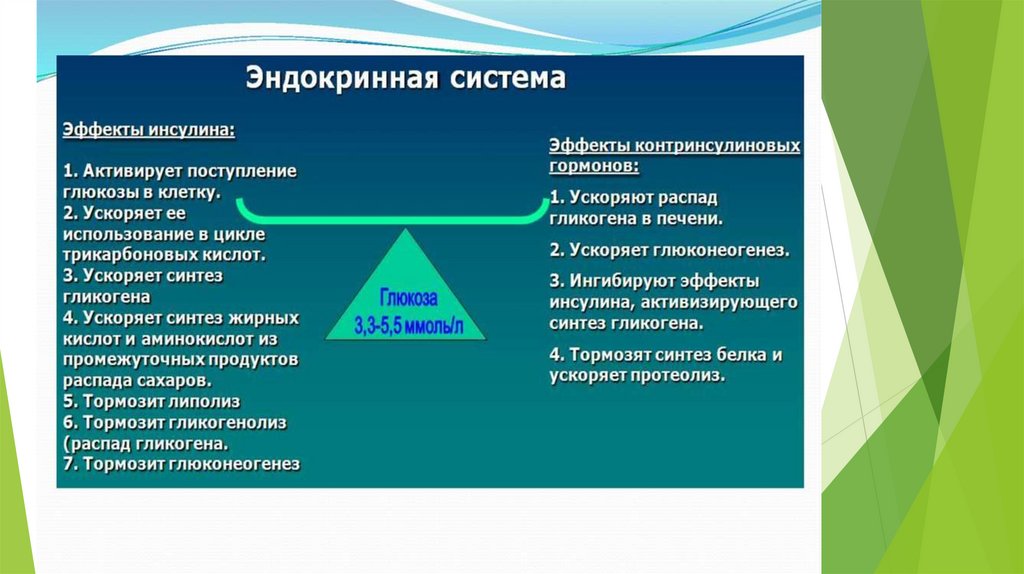
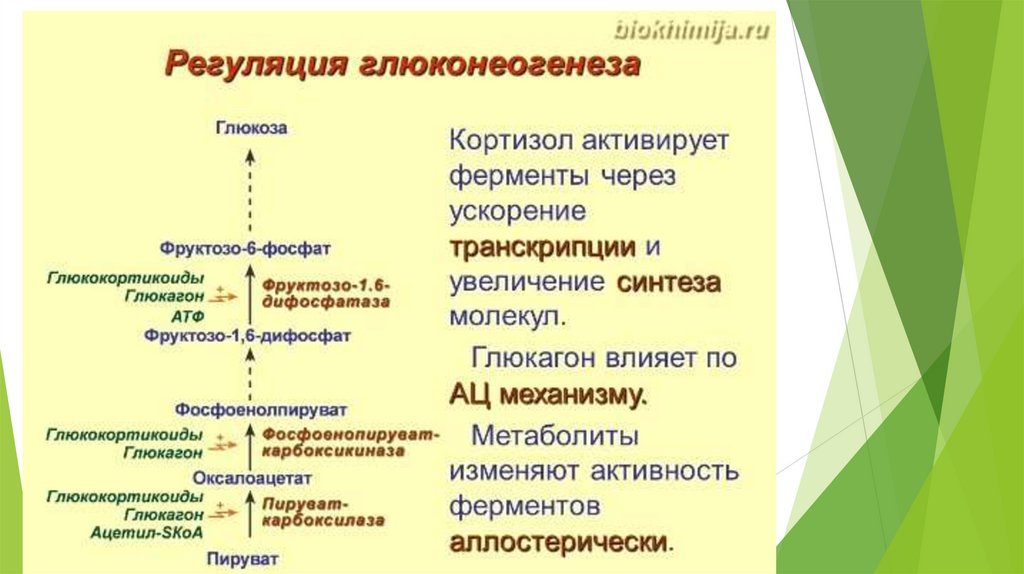
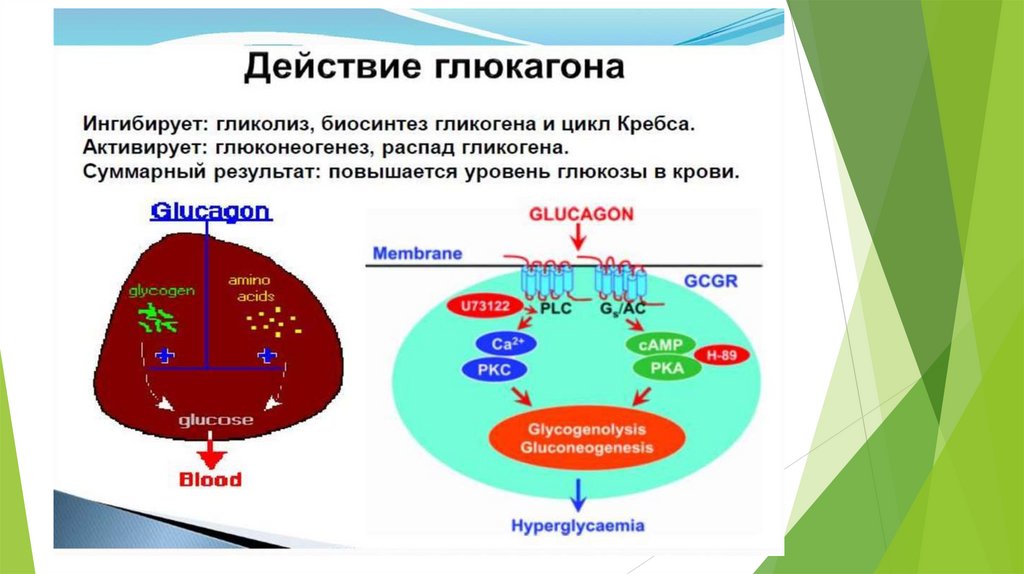
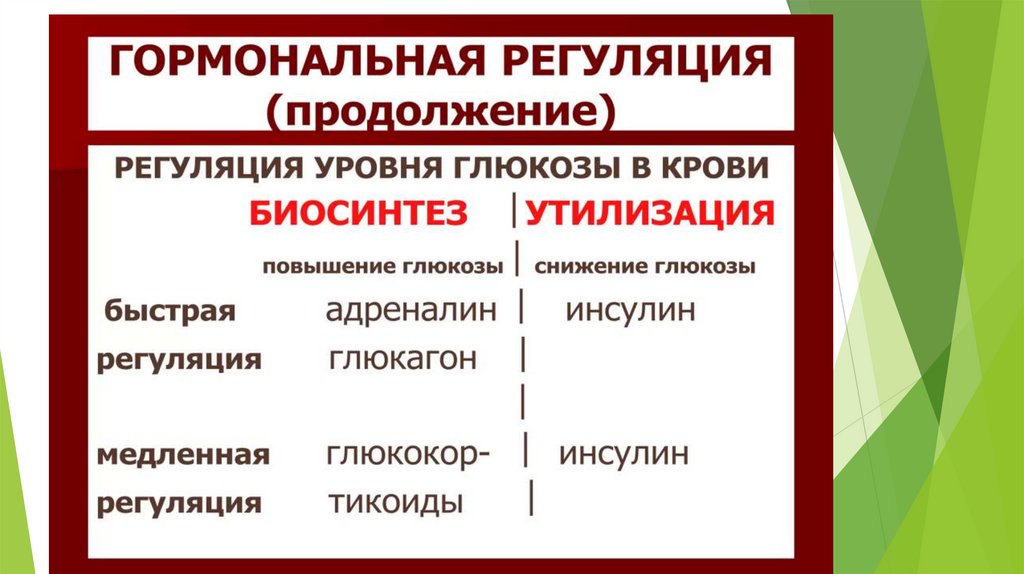
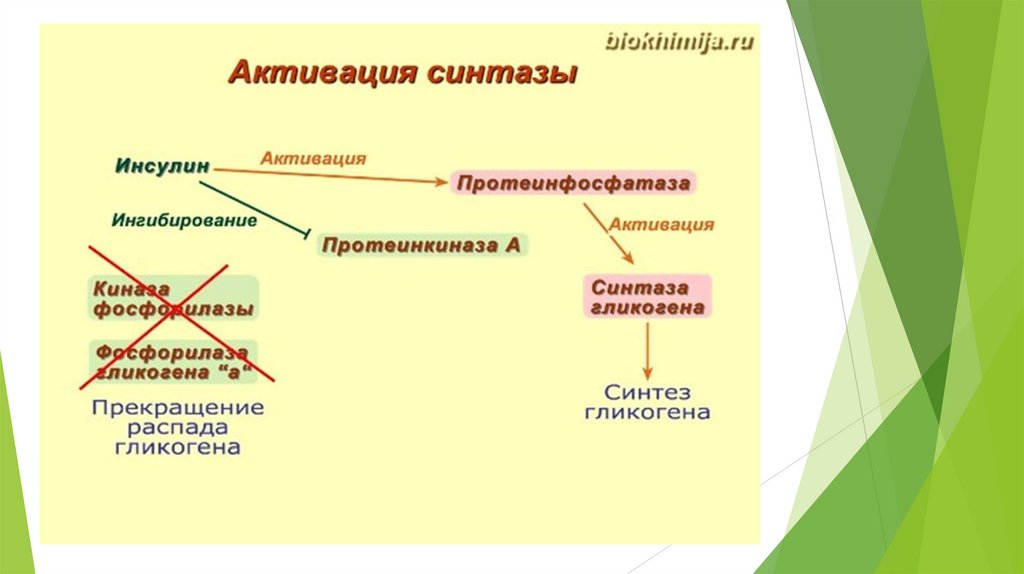
Глюконеогенез контролируется гормонами.
Кортизол, глюкагон и адреналин стимулируют этот процесс, а инсулинподавляет.
Центральный путь глюконеогенеза – превращение пирувата в глюкозу.
7 стадий гликолиза обратимы; 3 стадии необратимы.
9.
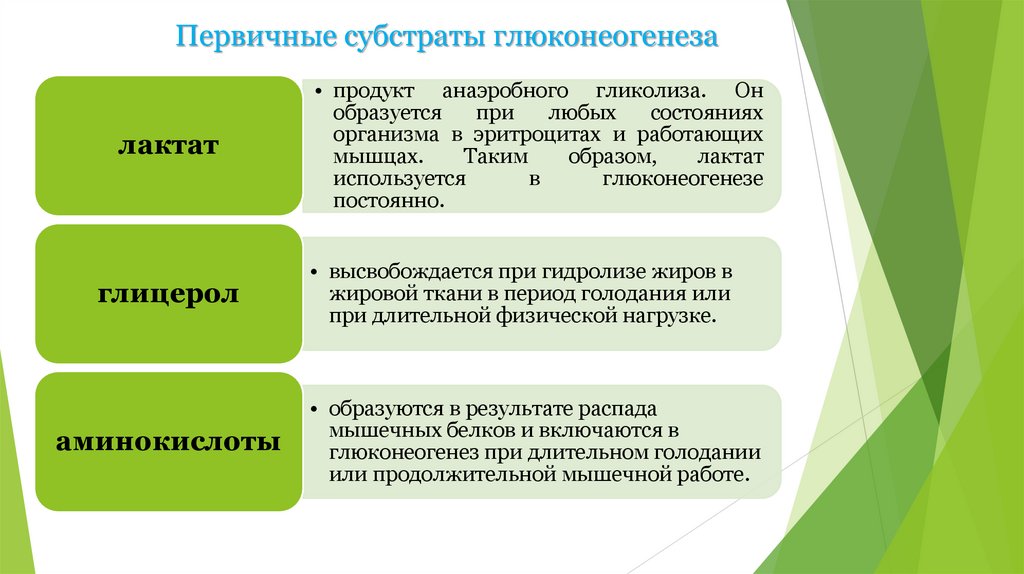
Первичные субстраты глюконеогенезалактат
• продукт анаэробного гликолиза. Он
образуется
при
любых
состояниях
организма в эритроцитах и работающих
мышцах.
Таким
образом,
лактат
используется
в
глюконеогенезе
постоянно.
глицерол
• высвобождается при гидролизе жиров в
жировой ткани в период голодания или
при длительной физической нагрузке.
аминокислоты
• образуются в результате распада
мышечных белков и включаются в
глюконеогенез при длительном голодании
или продолжительной мышечной работе.
10.
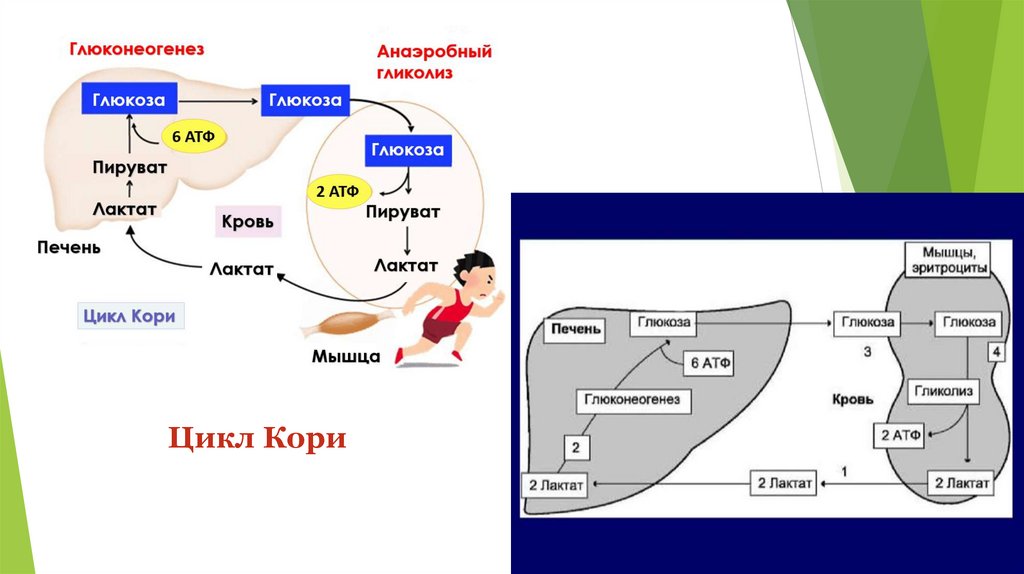
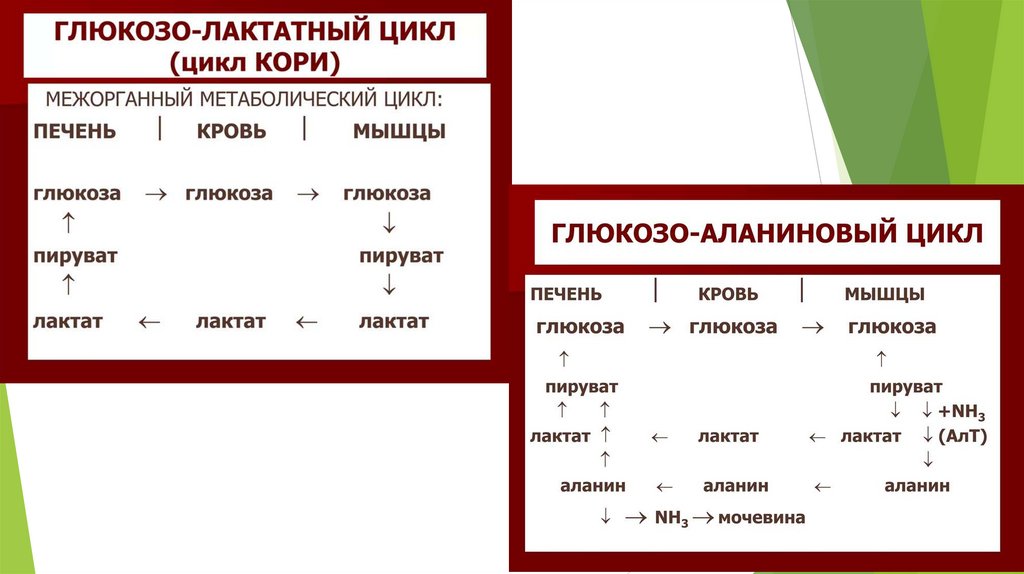
Примеры субстратов для глюконеогенеза11.
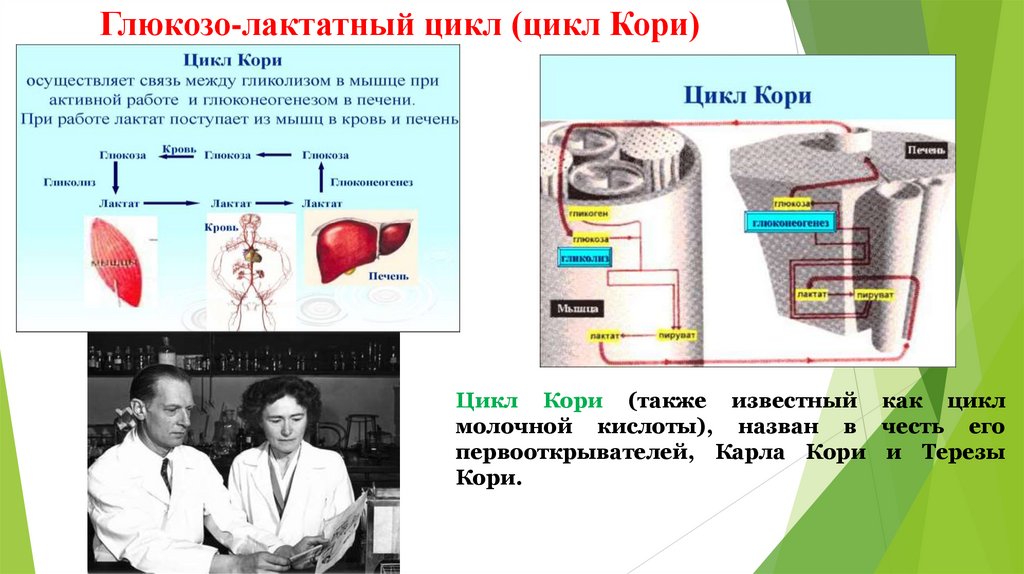
Глюкозо-лактатный цикл (цикл Кори)Цикл Кори (также известный как цикл
молочной кислоты), назван в честь его
первооткрывателей, Карла Кори и Терезы
Кори.
12.
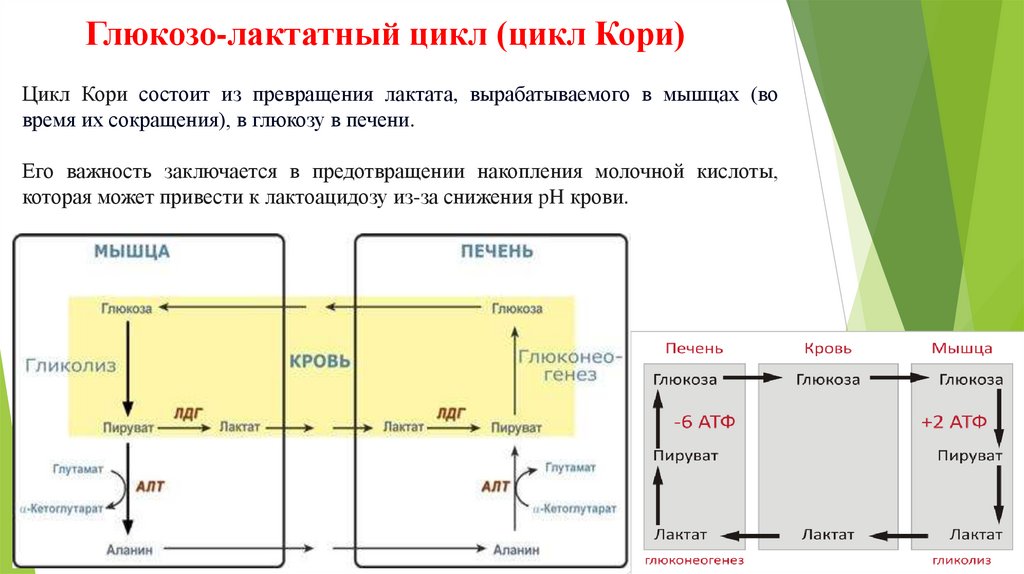
Глюкозо-лактатный цикл (цикл Кори)Цикл Кори состоит из превращения лактата, вырабатываемого в мышцах (во
время их сокращения), в глюкозу в печени.
Его важность заключается в предотвращении накопления молочной кислоты,
которая может привести к лактоацидозу из-за снижения pH крови.
13.
Глюкозо-лактатный цикл (цикл Кори)Глюкозо-лактатный цикл – это циклический процесс, объединяющий реакции
глюконеогенеза и реакции анаэробного гликолиза. Глюконеогенез происходит в
печени, субстратом для синтеза глюкозы является лактат, поступающий в
основном из эритроцитов или мышечной ткани.
В эритроцитах молочная кислота образуется непрерывно, так как для них
анаэробный гликолиз является единственным способом образования энергии.
В скелетных мышцах высокое накопление молочной кислоты (лактата)
является следствием гликолиза при очень интенсивной, субмаксимальной
мощности, работе, при этом внутриклеточный рН снижается до 6,3-6,5. Но даже
при работе низкой и средней интенсивности в скелетной мышце всегда
образуется некоторое количество лактата.
Убрать молочную кислоту можно только одним способом – превратить ее в пировиноградную кислоту.
Однако сама мышечная клетка ни при работе, ни во время отдыха не способна превратить лактат в
пируват из-за особенностей изофермента лактатдегидрогеназы5. Зато клеточная мембрана высоко
проницаема для лактата, он движется по градиенту концентрации наружу и переносится кровью в
печень. Поэтому во время и после нагрузки (при восстановлении) лактат легко удаляется из мышцы.
Это происходит довольно быстро, всего через 0,5-1,5 часа в мышце лактата уже нет. Малая часть
молочной кислоты выводится с мочой.
14.
Цикл Кори15.
16.
17.
18.
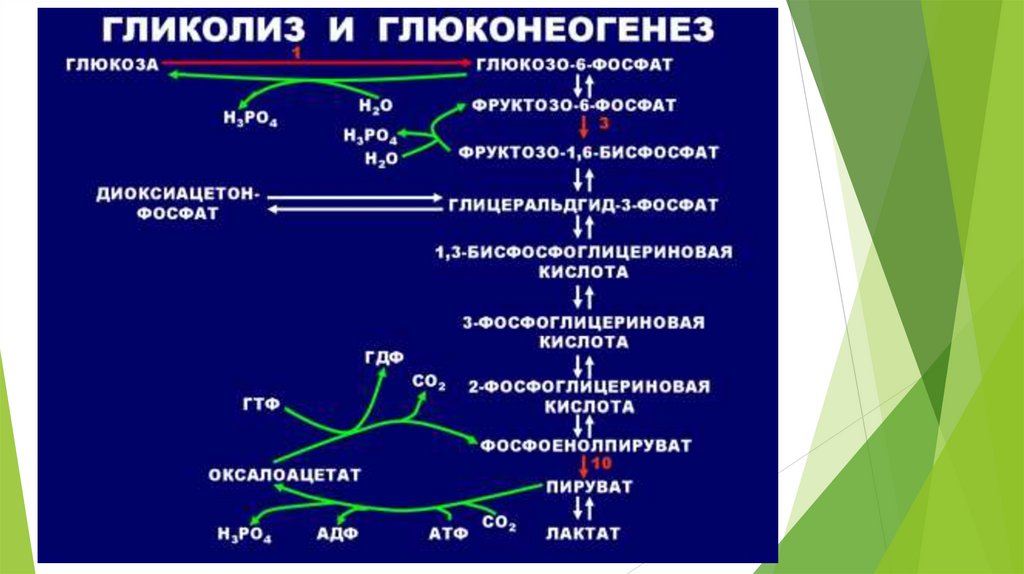
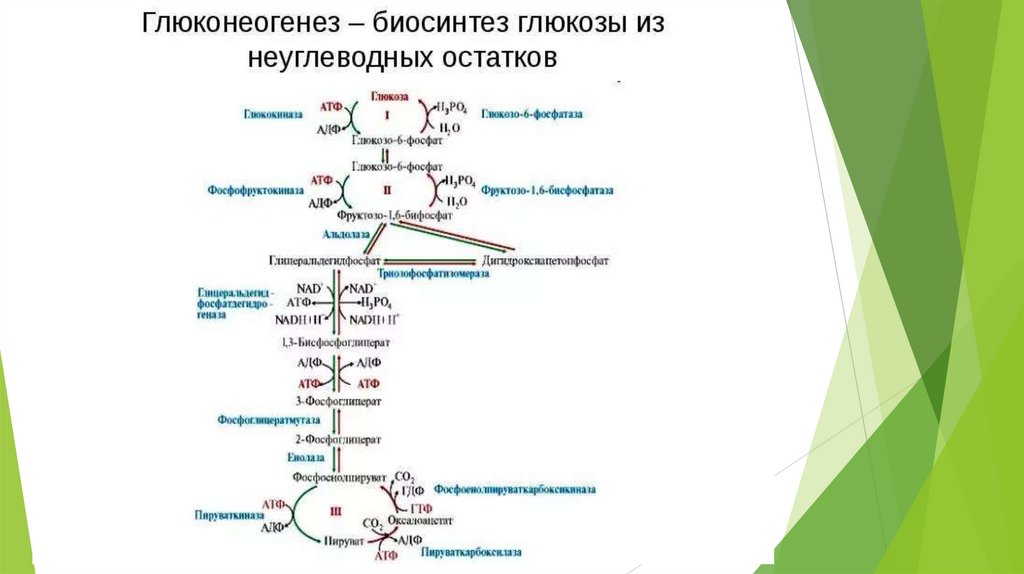
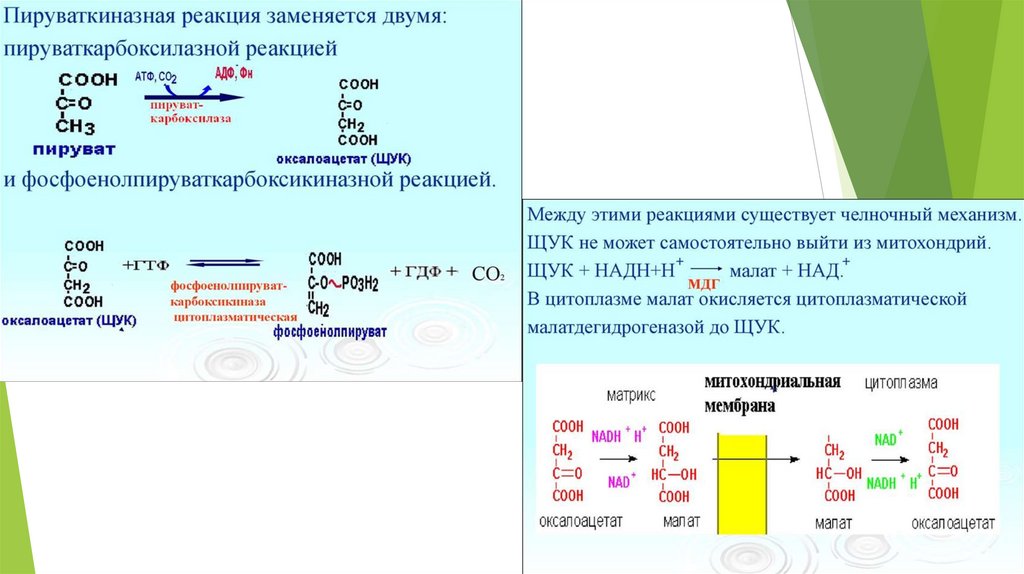
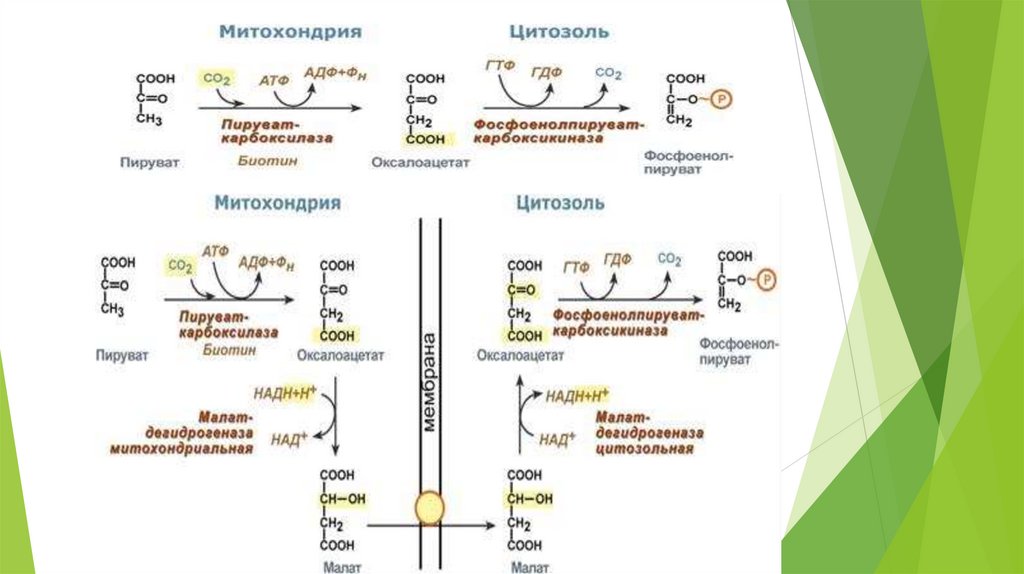
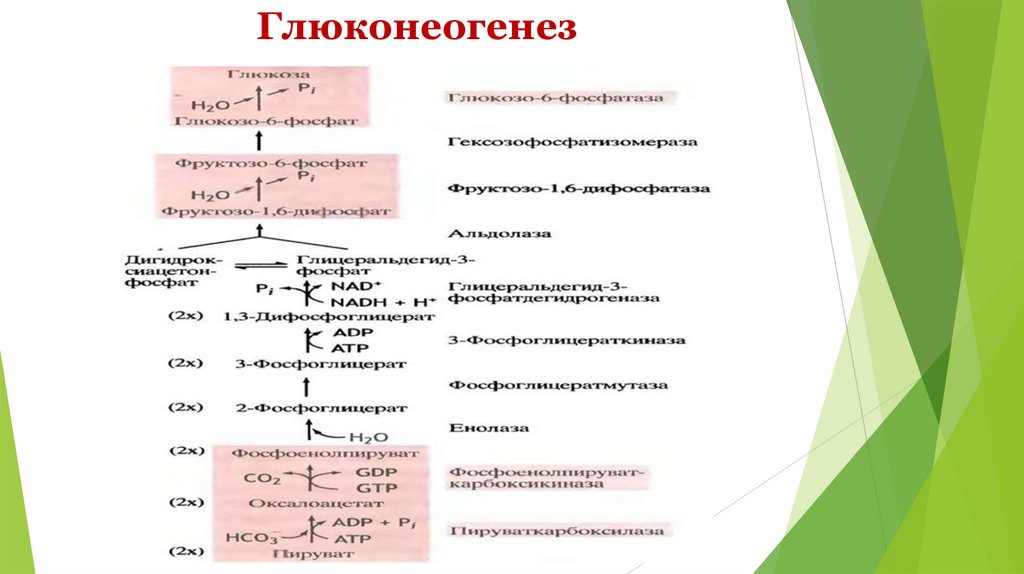
Необратимые стадии гликолиза( обходные пути глюконеогенеза)
1.
превращение пирувата в ФЕП – для осуществления
требуются цитозольные и митохондриальные ферменты клеток
печени; расход энергии = 14,8 ккал/моль);1АТФ+1ГТФ;
Ферменты: пируваткарбоксилаза, малатдегидрогеназа, NADзависимая малатдегидрогеназа, фосфоенолпируват-карбоксилаза.
превращение фруктозо-1,6-дифосфата во фруктозо- 6фосфат
Ферменты: фруктозодифосфатаза ( Mg-зависимая)
3. превращение глюкозо-6-фосфата в глюкозу
Ферменты: глюкозо-6-фосфатаза ( Mg-зависимая)
2.
Энергетические затраты глюконеогенеза: для образования 1
молекулы глюкозы расходуется: 6 фосфатных групп – 4 от АТФ и 2
от ГТФ; для восстановительных этапов требуется 2 молекулы
NADH.
19.
20.
21.
22.
23.
24.
Глюконеогенез25.
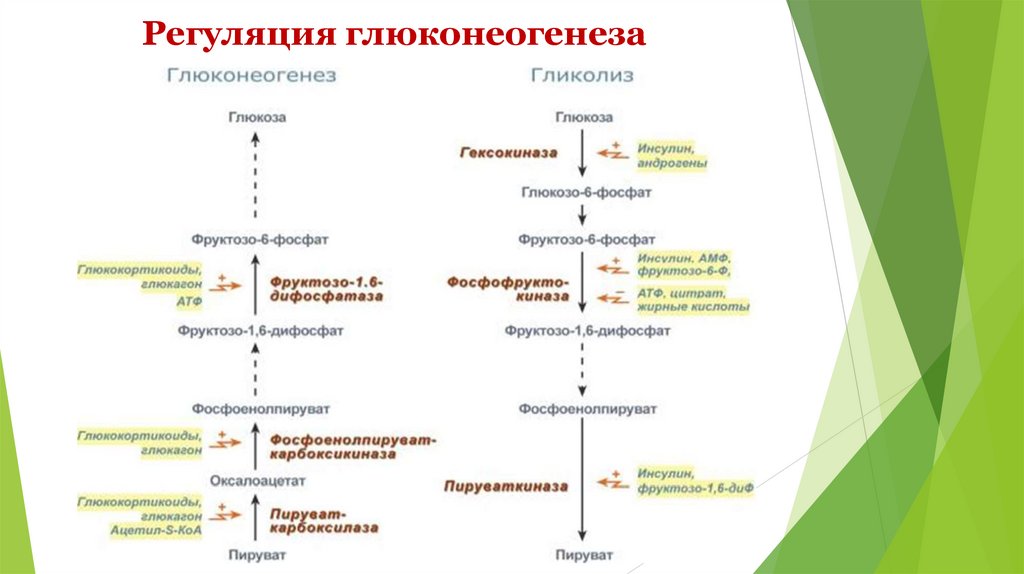
Регуляция глюконеогенеза26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
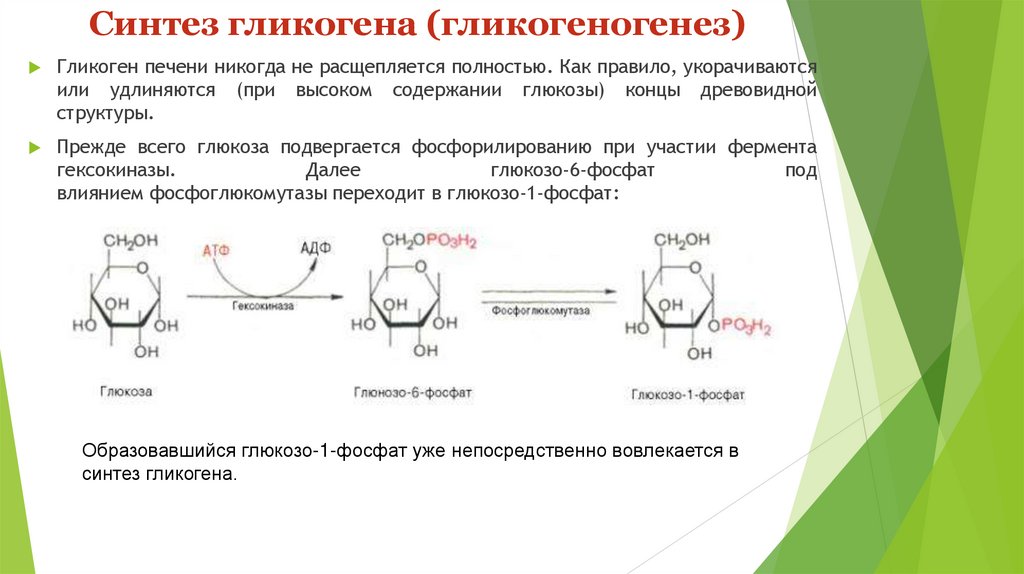
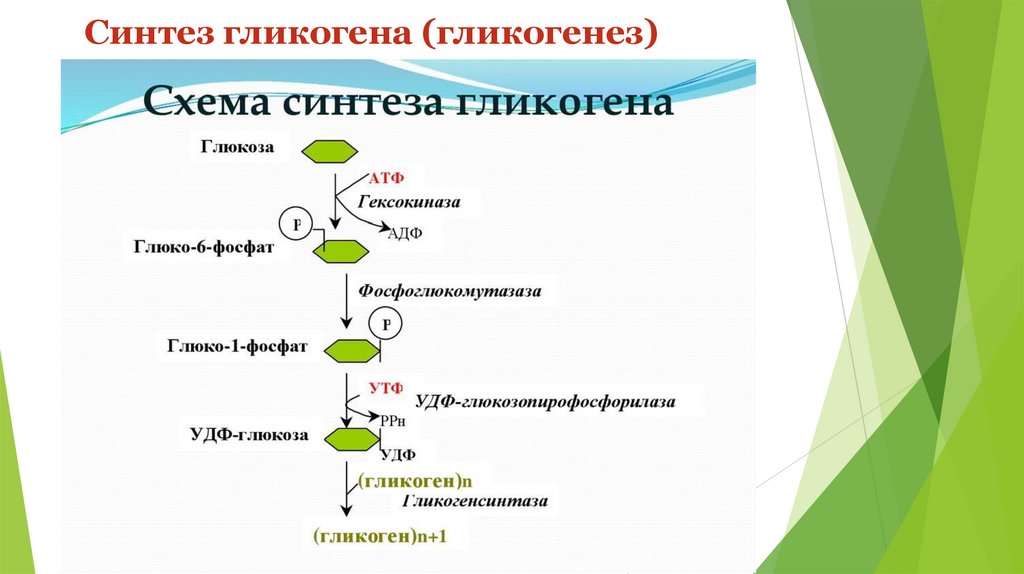
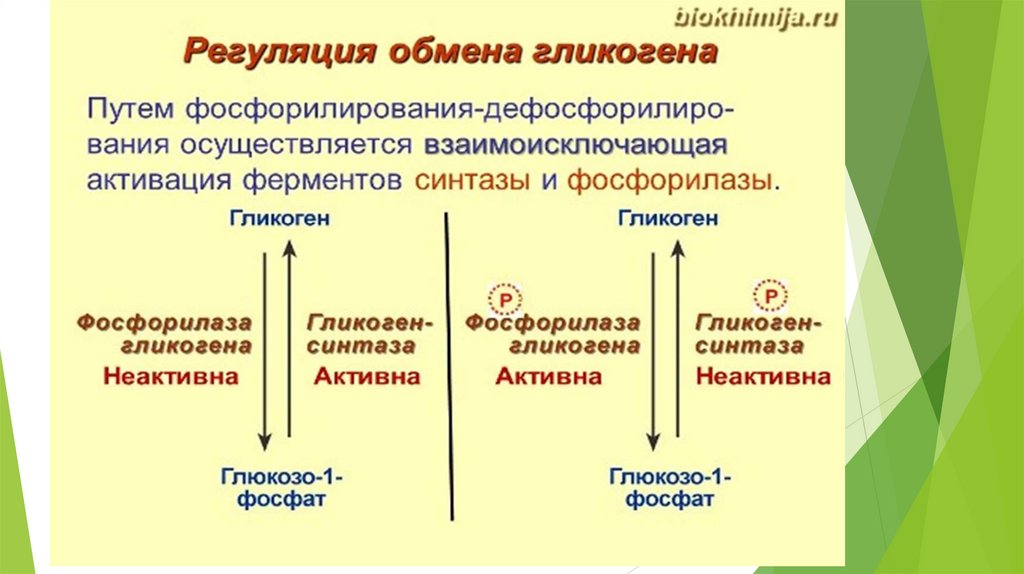
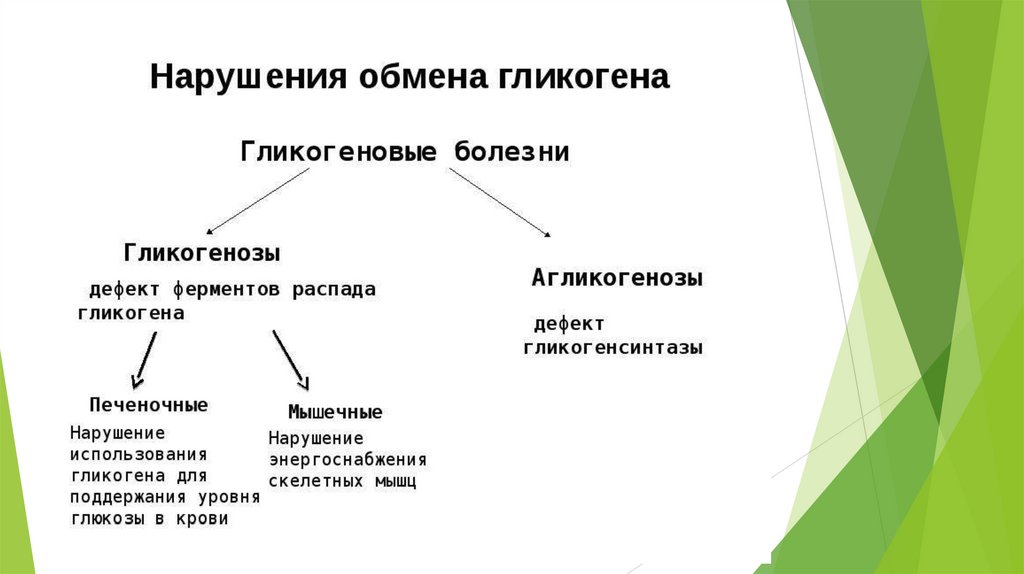
Синтез гликогена (гликогеногенез)Гликоген печени никогда не расщепляется полностью. Как правило, укорачиваются
или удлиняются (при высоком содержании глюкозы) концы древовидной
структуры.
Прежде всего глюкоза подвергается фосфорилированию при участии фермента
гексокиназы.
Далее
глюкозо-6-фосфат
под
влиянием фосфоглюкомутазы переходит в глюкозо-1-фосфат:
Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в
синтез гликогена.
41.
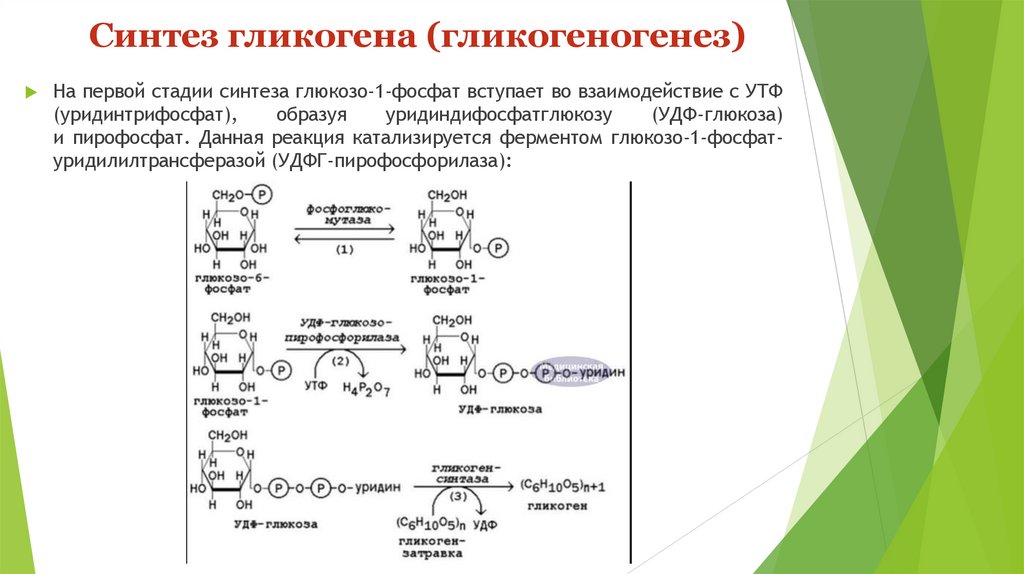
Синтез гликогена (гликогеногенез)На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ
(уридинтрифосфат),
образуя
уридиндифосфатглюкозу
(УДФ-глюкоза)
и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфатуридилилтрансферазой (УДФГ-пирофосфорилаза):
42.
Синтез гликогена (гликогенез)43.
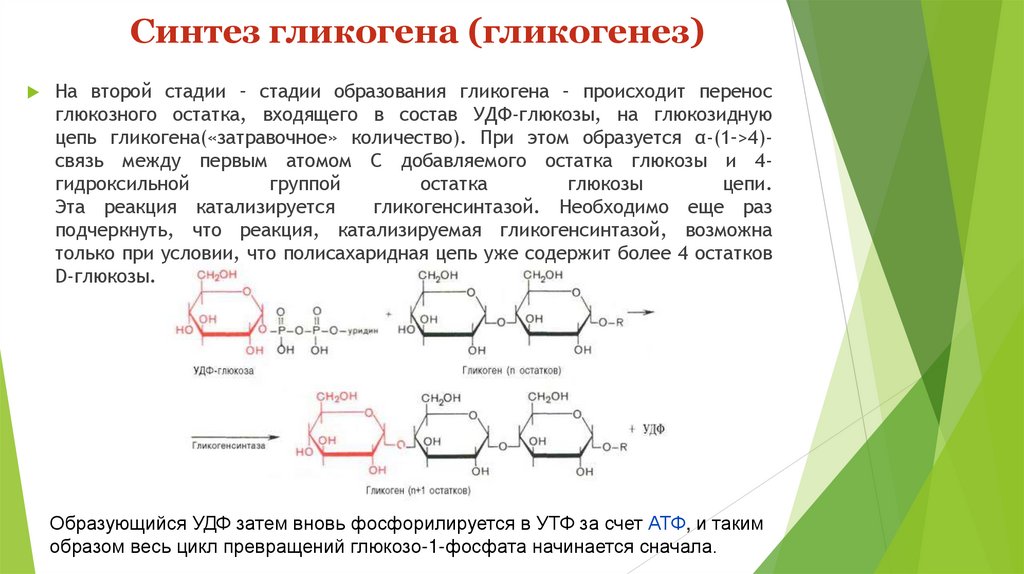
Синтез гликогена (гликогенез)На второй стадии – стадии образования гликогена – происходит перенос
глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную
цепь гликогена(«затравочное» количество). При этом образуется α-(1–>4)связь между первым атомом С добавляемого остатка глюкозы и 4гидроксильной
группой
остатка
глюкозы
цепи.
Эта реакция катализируется
гликогенсинтазой. Необходимо еще раз
подчеркнуть, что реакция, катализируемая гликогенсинтазой, возможна
только при условии, что полисахаридная цепь уже содержит более 4 остатков
D-глюкозы.
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким
образом весь цикл превращений глюкозо-1-фосфата начинается сначала.
44.
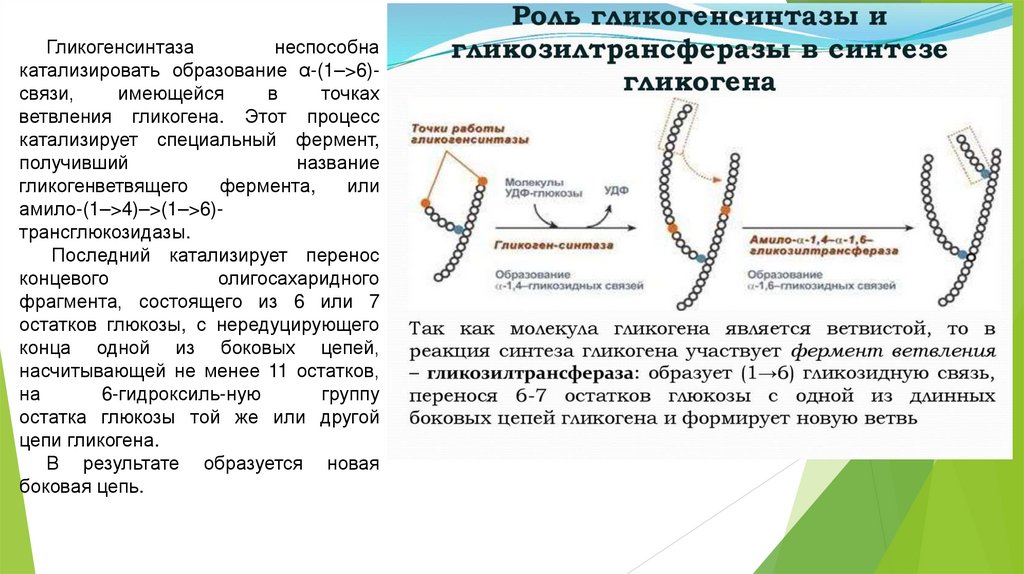
Гликогенсинтазанеспособна
катализировать образование α-(1–>6)связи,
имеющейся
в
точках
ветвления гликогена. Этот процесс
катализирует специальный фермент,
получивший
название
гликогенветвящего
фермента,
или
амило-(1–>4)–>(1–>6)трансглюкозидазы.
Последний катализирует перенос
концевого
олигосахаридного
фрагмента, состоящего из 6 или 7
остатков глюкозы, с нередуцирующего
конца одной из боковых цепей,
насчитывающей не менее 11 остатков,
на
6-гидроксиль-ную
группу
остатка глюкозы той же или другой
цепи гликогена.
В результате образуется новая
боковая цепь.