










































































 biology
biologySimilar presentations:

")


")
")




Красные и бурые водоросли
1. Красные и бурые водоросли
Лекция № 41
2. ОТДЕЛ КРАСНЫЕ ВОДОРОСЛИ
•почти исключительно морские водоросли.•жизненный цикл - полное отсутствие
жгутиковых стадий
• особая форма оогамного полового процесса.
2
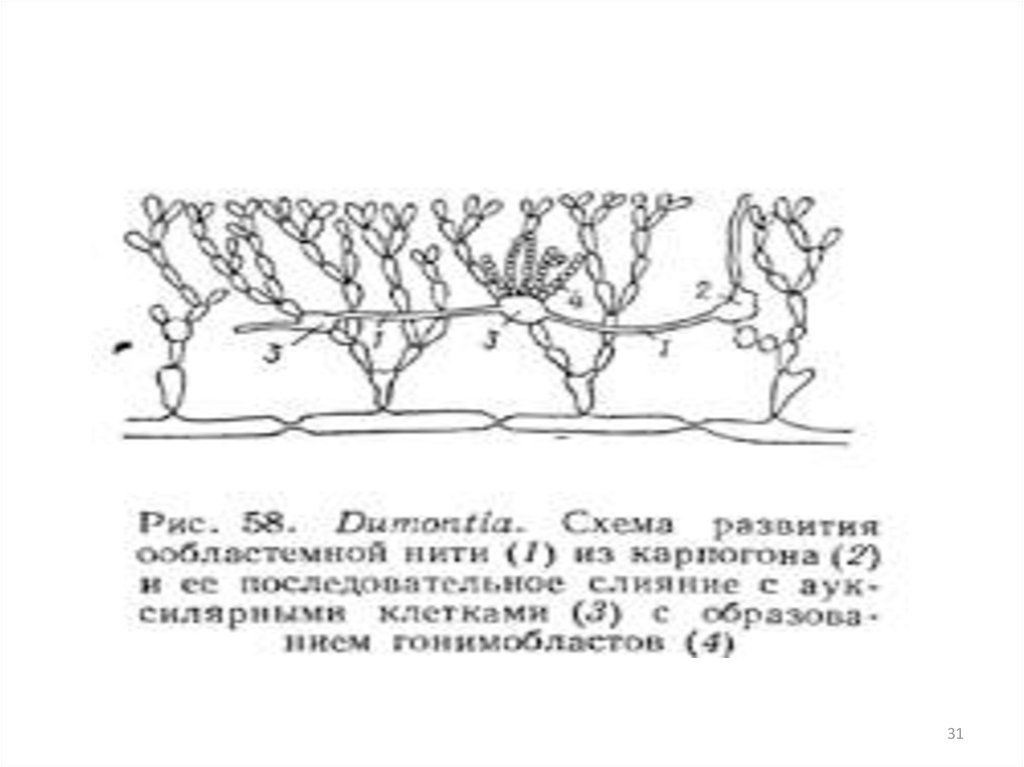
3. Пигменты
• хлорофилл «а» и «в»,• каротиноиды (каротин, зеаксантин,
антераксантин,
• криптоксантин, лютеин, неоксантин)
• фикобилины: фикоэритрины (красный)
• фикоцианины и аллофикоцианин (синий)
• окраска таллома от малиново-красной до
голубовато-стальной (при избытке
фикоцианина)
3
4. Хлоропласты
•оболочка из двух мембран•одиночные тилакоиды, на поверхности фикобилисомы.
•Запасной продукт — полисахарид «багрянковый
крахмал», от иода приобретающий буро-красный
цвет.
•Зерна багрянкового крахмала откладываются в
цитоплазме всегда вне связи с пиреноидами и
хроматофорами.
4
5. Строение таллома
•одноклеточные коккоидные формы(порфиридиум)
•гетеротрихальные и в виде разветвленных
нитей, прикрепленных к субстрату с
помощью ризоидов.
• псевдопаренхиматозные талломы, переплетение боковых ветвей
• Пластинчатые талломы паренхиматозного
строения (порфира).
5
6. Клетка
•одета оболочкой•пектин - гемицеллюлозные компоненты
набухают
•в стенках откладывается известь.
• Клетки одно- и многоядерные
• Хроматофоры париетальные,
многочисленные, в виде зерен или
пластинок.
6
7. Бесполое размножение
посредством неподвижных клеток
моноспоры - одна в спорангии
Тетраспоры - четырех
Тетраспоры - на диплоидных бесполых
растениях — спорофитах .
• В спорангиях перед образованием
тетраспор - мейоз.
7
8. Половой процесс
• Оогамный• Женский орган — карпогон
• у большинства -из расширенной базальной
части — брюшка - яйцеклетка, и отростка —
трихогины.
• Карпогон развивается на особой короткой
карпогониальной ветви .
8
9.
• Антеридии — мелкие бесцветные клетки• содержат безжгутиковые спермации
• спермации пассивно переносятся токами
воды и прилипают к трихогине.
• В месте контакта спермация и трихогины их
стенки растворяются
• ядро спермация перемещается по
трихогине
• в брюшной части карпогона сливается с
9
10. образование карпоспор
• После оплодотворения базальная частькарпогона отделяется перегородкой от
трихогины – отмирает
• Развитие - образование карпоспор
• содержимое зиготы (оплодотворенного
карпогона)
1. непосредственно делится с образованием
неподвижных голых спор — карпоспор,
10
11. образование карпоспор
2. из оплодотворенного карпогонавырастают ветвящиеся нити —
гонимобласты, клетки их превращаются в
карпоспорангии, производящие по одной
карпоспоре.
11
12. образование карпоспор
• у большинства гонимобласты развиваются ненепосредственно из брюшка оплодотворенного
карпогона
• из а у к с и л я р н ы х клеток.
• могут быть удалены от карпогона или
располагаются на талломе в непосредственной
близости
• когда ауксилярные клетки удалены от карпогона,
из его брюшка после оплодотворения вырастают с
о е д и н и т е л ь н ы е , или о о б л ас т е м н ы е ,
нити.
12
13.
клетки ообластемных нитей содержат, диплоидные ядра. Ообластемные нити
подрастают к ауксилярным клеткам, в точке контакта оболочки растворяются и между
клеткой ообластемной нити и ауксилярной клеткой устанавливается сообщение. Это
слияние клеток не сопровождается слиянием их ядер (диплоидного ядра клетки ообластемной нити
и гаплоидного ядра ауксилярной).
• слияние с ауксилярной клеткой
стимулирует деление диплоидного ядра
клетки ообластемной нити и развитие
гонимобластов
• содержат диплоидные ядра и продуцируют
диплоидные карпоспоры
• Гонимобласты - особая генерация —
13
14.
• У наиболее высокоорганизованных красных водорослей ауксилярныеклетки развиваются только после оплодотворения карпогона и в
непосредственной от него близости.
• Совокупность ауксилярной клетки (или клеток) с карпогоном носит
специальное название прокарпий .
• Здесь нет надобности в образовании длинных ообластемных нитей,
связывающих карпогон и ауксилярные клетки, ауксилярная клетка
просто сливается с брюшком оплодотворенного карпогона, после чего
из нее развиваются гонимобласты с карпоспорами
• Карпоспорангии часто располагаются тесными группами —
цистокарпиями, одеты псевдопаренхиматозной оболочкой,
развивающейся из соседних с карпогоном клеток.
14
15.
1516. Отдел Красные водоросли Rhodophyta
Отдел Красные водорослиRhodophyta
Красные водоросли делят на два класса:
• бангиевые
• флоридеи
16
17. КЛАСС БАНГИЕВЫЕ
• Клетки часто со звездчатым хроматофороми пиреноидом.
• Поры между клетками, как правило,
отсутствуют.
• Карпогон без трихогины, после
оплодотворения содержимое карпогона
непосредственно делится с образованием
карпоспор. Бесполое размножение —
моноспорами
17
18. Представитель — род порфира
1819.
1920. род компсопогон
• Пресноводный широко распространенный втропиках занесена с аквариумными
растениями
• Таллом гетеротрихальный
• Размножение моноспорами. Моноспорангии
отчленяются косо идущей перегородкой от
любой клетки таллома и имеют зернистое от
обилия багрянкового крахмала содержимое,
которое превращается в одну неподвижную
голую моноспору
20
21.
2122. КЛАСС ФЛОРИДЕИ
• Порядок немалионовые• Порядок криптонемиевые
22
23. КЛАСС ФЛОРИДЕИ
• Клетки большей частью с париетальными хроматофорами безпиреноидов
• Между клетками имеются поры.
• Карпогон с трихогиной.
• После оплодотворения развиваются гонимобласты или
непосредственно из брюшка оплодотворенного карпогона, или из
ауксилярных клеток после слияния их с ообластемными нитями.
• Бесполое размножение большинства— тетраспорами.
• Гонимобласты, несущие карпоспорангии, рассматриваются как особая
генерация — карпоспорофит.
• Детали развития карпоспорофита (из оплодотворенного карпогона
или из ауксилярных клеток), а также время дифференцировки
ауксилярных клеток (до или после оплодотворения), их положение на
талломе и т. п. кладутся в основу деления флоридей на шесть
порядков
23
24. Порядок немалионовые
• Для представителей характерно отсутствиеауксилярных клеток
• гонимобласты развиваются
непосредственно из оплодотворенного
карпогона
24
25. Род леманеа
2526. род немалион
2627. род батрахоспермум
2728.
• Род леманеа встречается в быстро текущих речках• с холодной водой. Таллом, имеющий вид неветвящейся щетинки 10—
15 см длиной и 1 мм толщиной, темно-фиолетового или оливково• бурого цвета с узловатыми вздутиями. Прикрепляется к субстрату
• при помощи подошвы из стелющихся нитей. По оси таллома проходит
• одна нить из вытянутых бесцветных клеток. От верхней части каждой
• клетки центральной оси отходит мутовка большей частью из четырех
• расходящихся по радиусам ветвей. Базальные клетки каждой такой
• ветви крупные, вытянутые. От их дистального (морфологически верх• него) конца отходят ветви второго порядка, которые в свою очередь
• многократно ветвятся. Конечные разветвления срастаются в
многослой• ную кору. Ее наружные клетки мелкие и заполнены хроматофорами,
28
29. Порядок криптонемиевые
имеются ауксилярные клетки, которые развиваются до оплодотворения
карпогона и рассеяны по таллому на известном расстоянии от карпогона.
Из карпогона после оплодотворения к ауксилярным клеткам растут более или
менее длинные многоклеточные соединительные, или ообластемные, нити.
После слияния ауксилярной клетки с клеткой ообластемной нити и перехода
диплоидного ядра клетки нити в ауксилярную клетку возникают
гонимобласты
Развивающиеся на гонимобластах (карпоспорофите) карпоспоры содержат
диплоидное ядро и прорастают в диплоидные растения — тетраспорофиты,
производящие только органы бесполого размножения — тетраспорангии.
При образовании тетраспор происходит мейоз и гаплоидные тетраспоры
прорастают в гаплоидные растения— гаметофиты, несущие половые органы.
Гаметофит и тетраспорофит внешне (морфологически) не различаются.
изоморфная смена генераций, усложненная диплоидным карпоспорофптом.
29
30. род дюренеа
• распространенный в южных морях.• Таллом имеет вид разветвленных
слизистых кустиков розового цвета.
30
31.
3132.
3233.
3334. Род кораллина
3435. Род литотамнион
3536. Порядок церамиевые
• высоко организованы• много видов
• ауксилярные клетки дифференцируются
только после того, как произошло
оплодотворение карпогона
• Длинных ообластемных нитей не образуется:
брюшко карпогона или непосредственно
сливается с рядом расположенной
ауксилярной клеткой (клетками), или через
короткие выросты, формируя цистокарпии
36
37. род каллитамнион
3738. род делессерия
3839. эволюция в пределах отдела
Бангиевые более примитивны.
у карпогона еще не выработалась типичная форма, мало отличается от обычных вегетативных
клеток.
После оплодотворения содержимое карпогона непосредственно делится на
карпоспоры.
Флоридеи - более эволюционировавшая группа
Карпогоном с трихогиной.
Из класса флоридей наиболее простой порядок - немалионовые, у которых нет ауксилярных
клеток, и гонимобласты, на которых развиваются карпоспорангии, образуются
непосредственно из брюшной части оплодотворенного карпогона.
Криптонемиевые - следующий шаг прогрессивной эволюции: имеют ауксилярные клетки,
способствующие увеличению продукции карпоспор, так как образуется не один цистокарпий,
а много — по числу ауксилярных клеток. При разбросанных беспорядочно по таллому
ауксилярных клетках необходимы более или менее длинные ообластемные нити.
Наивысшей ступени эволюции достигли церамиевые, у которых имеется прокарпий и
ауксилярные клетки дифференцируются только после того, как произошло оплодотворение.
Непосредственное соседство карпогона и ауксилярной клетки (клеток) в прокарпии облегчает
образование цпстокарпиев. Этот порядок наиболее богат видами.
39
40.
• За исключением немногих, преимущественнопростейших представителей живущих в быстро
текущих речках с чистой водой, красные
водоросли — морские обитатели.
• Они всегда прикреплены к камням, ракушкам,
другим водорослям, являясь, как правило,
эпифитами, однако известны и эндофиты, и даже
паразиты. Многие красные водоросли живут на
больших глубинах, и глубоководные формы
отличаются особенно яркой красной окраской.
40
41.
• Древнейшая окаменелость, идентифицируемая каккрасная водоросль является также самой древней
окаменелостью-эукариотом, принадлежащей к
конкретному современному таксону.Bangiomorpha
pubescens, окаменелость многоклеточного организма
найденная в арктической Канаде, лишь немногим
отличается от современной красной водоросли
рода Бангия (Bangia), несмотря на то, что она была
отложена в слоях 1,2 миллиардной давности.
41
42. ОТДЕЛ БУРЫЕ ВОДОРОСЛИ РНАЕОРНУТА
• морские, особенно много - в холодных водах северного июжного полушария.
• По морфологической и анатомической дифференцировке
таллома - на более высоком уровне, чем ранее
рассмотренные группы водорослей.
• неизвестны ни одноклеточные, ни колониальные формы,
ни талломы в виде простой неразветвленной нити.
• самые простейшие - гетеротрихальные
• талломы крупные, ложно- или истиннотканевого
строения.
43.
сильно ослизняющиеся клеточные стенки
одно ядро,
одну или много вакуолей
обычно постенные хроматофоры различной формы.
хроматофоры окружены сложной системой мембран - в
прямой связи с оболочкой ядра — «хлоропластной
эндоплазматической сетью»
• Матрикс хлоропласта пересекается параллельными
трехтилакоидными ламеллами
• Свободный от тилакоидов пиреноид выступает из
хлоропласта в виде почки
44. Пигменты
• хроматофоры окрашены в бурый цвет• хлорофиллы «а» и «с» (хлорофилл «в»
отсутствует)
• в-каротин
• Много бурых ксантофиллов, особенно
фукоксантина.
44
45.
• Запасной полисахарид — ламинарин• спирт маннит
• жиры
• откладывается вне хлоропласта в цитоплазме.
• Монадные клетки (зооспоры и гаметы) имеют
глазок и жгутики. Глазок — часть пластиды,
связан со жгутиковым аппаратом.
• Жгутики - гетероморфные.
45
46. Организация
4647.
48. Размножение
• вегетативное, бесполое и половое.• Вегетативное размножение -участками таллома. У некоторых специализированные веточки (выводковые почки), отламываются -в
новые талломы.
• Бесполое размножение - зооспоры, образующихся в одногнездных
или однокамерных спорангиях на диплоидных растениях
(спорофитах) перед формированием зооспор ядро редукционно
делится
• Гаплоидные зооспоры - в гаплоидные половые растения —
гаметофиты, на которых образуются половые органы.
• У простейших - половой процесс изогамный, гаметыразвиваются в
многогнездных или многокамерных спорангиях
• У наиболее высокоорганизованных бурых водорослей половой
процесс оогамный. В оогониях и антеридиях, как правило, образуется
по одной гамете (яйцеклетке и сперматозоиду соответственно).
• Яйцеклетка оплодотворяется всегда вне оогония.
• Зигота без периода покоя прорастает в диплоидное растение.
49. ОТДЕЛ БУРЫЕ ВОДОРОСЛИ РНАЕОРНУТА
• КЛАСС ИЗОГЕНЕРАТНЫЕ• КЛАСС ГЕТЕРОГЕНЕРАТНЫЕ
• КЛАСС ЦИКЛОСПОРОВЫЕ
49
50. КЛАСС ИЗОГЕНЕРАТНЫЕ
• Порядок эктокарповые• род эктокарпус
- самые
примитивные бурые водоросли.
• Распространены во всех морях, особенно
холодных, и растут на подводных
предметах и других более крупных
водорослях.
50
51.
5152.
5253. Порядок сфацеляриевые
• Род сфацелярия имеет небольшиегетеротрихальные талломы.
53
54.
5455. Порядок кутлериевые
• Порядок включает только два рода: кутлерия изанардиния
• Кутлерия распространена по всему побережью
Европы, занардиния — преимущественно в
Средиземном море, а также в Черном.
• Оба рода обнаруживают чередование
поколений: у кутлерии смена генераций
гетероморфная, у занардинии — изоморфная
55
56. кутлерия
5657.
5758.
5859. Порядок диктиотовые
• род диктиота распространен вАтлантическом океане по берегам Европы
59
60.
6061. род падина
6162.
6263. КЛАСС ГЕТЕРОГЕНЕРАТНЫЕ
Порядок ламинариевые
Гаметофиты мало различаются между собой и представлены
микроскопическими, часто редуцированными до нескольких клеток
нитчатыми растеньицами-заростками, которые несут половые органы. На
мужских гаметофитах образуются антеридии в виде мелких клеток,
развивающих по одному сперматозоиду, на женских — оогонии, в которых
формируется по одной яйцеклетке.
Спорофиты разных родов резко различаются и представляют собой самые
крупные низшие растения, достигающие в длину 60—100 м,
со значительным морфологическим расчленением и сложным
анатомическим строением. В большинстве случаев спорофиты ламинариевых
расчленяются на листовидную пластинку, «ствол» и ризоиды, с помощью
которых все растение прикрепляется к подводным камням и скалам. В месте
перехода листовой пластинки в стволик находится интеркалярная
меристема, за счет деятельности которой нарастает как листовая
пластинка, так и ствол.
род ламинария, виды которого широко распространены в северных морях.
63
64.
65.
6566.
6667.
6768.
6869.
• У нереоцистиса длинный (до 25 м) сравнительно тонкийпервичный ствол на верхушке вздувается в крупный
воздушный пузырь, от которого отходят листовые
пластинки (до 5 м длины), каждая с коротким вторичным
стебельком.
• У макроцистиса, нереоцистиса, как и у ламинарии,
зооспорангии
• располагаются в сорусах на листовых пластинках.
69
70. КЛАСС ЦИКЛОСПОРОВЫЕ
Порядок фукусовые• характеризуется
–
–
–
–
верхушечным ростом таллома
Отсутствием бесполого размножения
оогамным половым процессом
половые органы располагаются в углублениях таллома — концептакулах,
или скафидиях
• Имеется вегетативное размножение, приводящее в некоторых
участках океана к образованию громадных скоплений талломов
саргассума (примером может служить Саргассово море).
71. род саргассум
• сложное морфологическое расчленение таллома• широко распространен в южном полушарии, особенно в теплых
морях.
• Стволик, прикреплен к субстрату подошвой
• ветвится
• несет уплощенные листовидные образования, шаровидные
воздушные пузыри на специальных стебельках
• разветвленные плодущие веточки
71
72.
73. род фукус
• В северных морях широко распространен73
74.
75.
Бурые водоросли — естественная группа, по своей морфологии хорошо отличающаяся от остальных отделов водорослей. Однако, как уже
говорилось, по ряду признаков (наличию хлорофиллов «а» и «с», отсутствию хлорофилла «Ь», расположению тилакоидов в хлоропластах —
трехтилакоидные ламеллы, по сходным продуктам запаса — ламинарии, хризоламинарин, по строению монадных клеток с гетероконтными
и гетероморфными жгутиками) бурые водоросли обнаруживают сходство с золотистыми, желтозелеными, диатомовыми и пирофитовыми водорослями. На этом основании некоторые авторы низводят бурые и
другие перечисленные группы водорослей до ранга классов в пределах
одного большого отдела СЬгошорЬу1а. Существование монадных клеток в жизненном цикле бурых водорослей позволяет выводить их, так
же как остальные перечисленные группы, из каких-то первичных фотосинтезирующих жгутиконосцев с преобладанием бурых пигментов. Эти
жгутиковые эволюционировали несколькими путями, один из которых
и привел к возникновению бурых водорослей. Однако у бурых водорослей связь со жгутиковыми более отдаленная, чем у сходных с ними
по ряду признаков золотистых, разножгутиковых, диатомовых, пирофитовых, а также у зеленых водорослей, так как прямых переходных
форм здесь нет. Среди бурых водорослей не прослеживается последовательное развитие типов организации таллома от монадного к нитчатому и пластинчатому, как это легко наблюдается у зеленых, желтозеленых, золотистых и пирофйтбвых водорослей. Как уже отмечалось,
среди бурых водорослей представлены только высшие ступени морфологической дифференцировки таллома — гетеротрихальная и пластинчатая. Возможно, более простые формы (монадные, коккоидные,
простые нитчатые) полностью утрачены на протяжении длительной эволюции, которую проделали бурые водоросли — древняя группа, известная уже; из отложений силура и девона.
Относительно родственных связей порядков в пределах отдела бурых водорослей и расположения их в системе единого взгляда нет.
Согласно одной из схем филогенеза бурых водорослей, построенной с
учетом сходства строения таллома на самых ранних стадиях развития,
бурые водоросли — группа дифилетическая: одна линия эволюции объединяет (из упомянутых в изложении) сфацеляриевые, диктиотовые и
фукусовые, другая — эктокарповые, кутлериевые, ламинариевые. Обе
219"
75
76.
группы порядков различаются и по способу роста талломов: первая характеризуется верхушечным ростом, вторая — интеркалярным. Килин
(Н. Ку 1ш), система которого наиболее распространена в настоящее
время, от общего предка бурых водорослей выводит три эволюционные
линии, которые представлены им как классы изогенератные, гетерогенератные и циклоспоровые; в основу их положены различия в жизненных циклах и морфологическом строении водорослей.
Крупные бурые водоросли находят большое применение в хозяйстве человека. Из них добывают альгин — клеящее вещество, использующееся в текстильной, пищевой и ряде других отраслей промышленности. В приморских странах выбросы морских водорослей, богатые
калием и азотом, употребляют в качестве удобрения, а также используют на корм скоту. Некоторые, например ламинарии («морская капуста»), съедобны.
76